

Characterization and Fungicide Screening of a New Pathogen That Causes Leaf Spot on Rehmannia glutinosa
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation
2.2. Pathogenicity Determination
2.3. Morphological Observations of Pathogens
2.4. DNA Extraction and Multiple Sequence Analysis of Pathogens
2.5. Determination of the Optimal Growth Conditions of the Pathogen
2.6. Bioassays
2.7. Data Analyses
3. Results and Analyses
3.1. Pathogen Isolation and Pathogenicity Determination
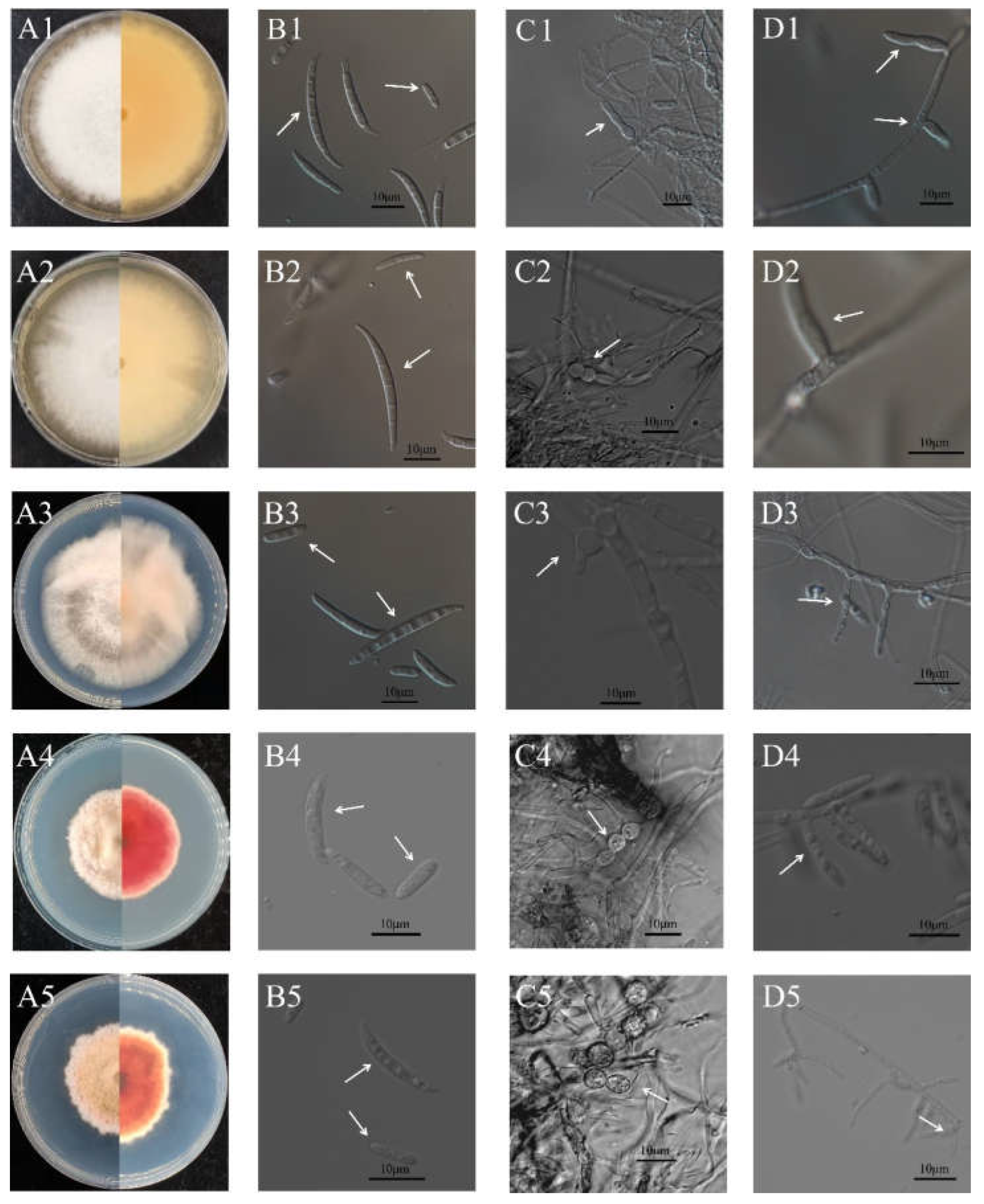
3.2. Morphological Characterization of the Pathogens
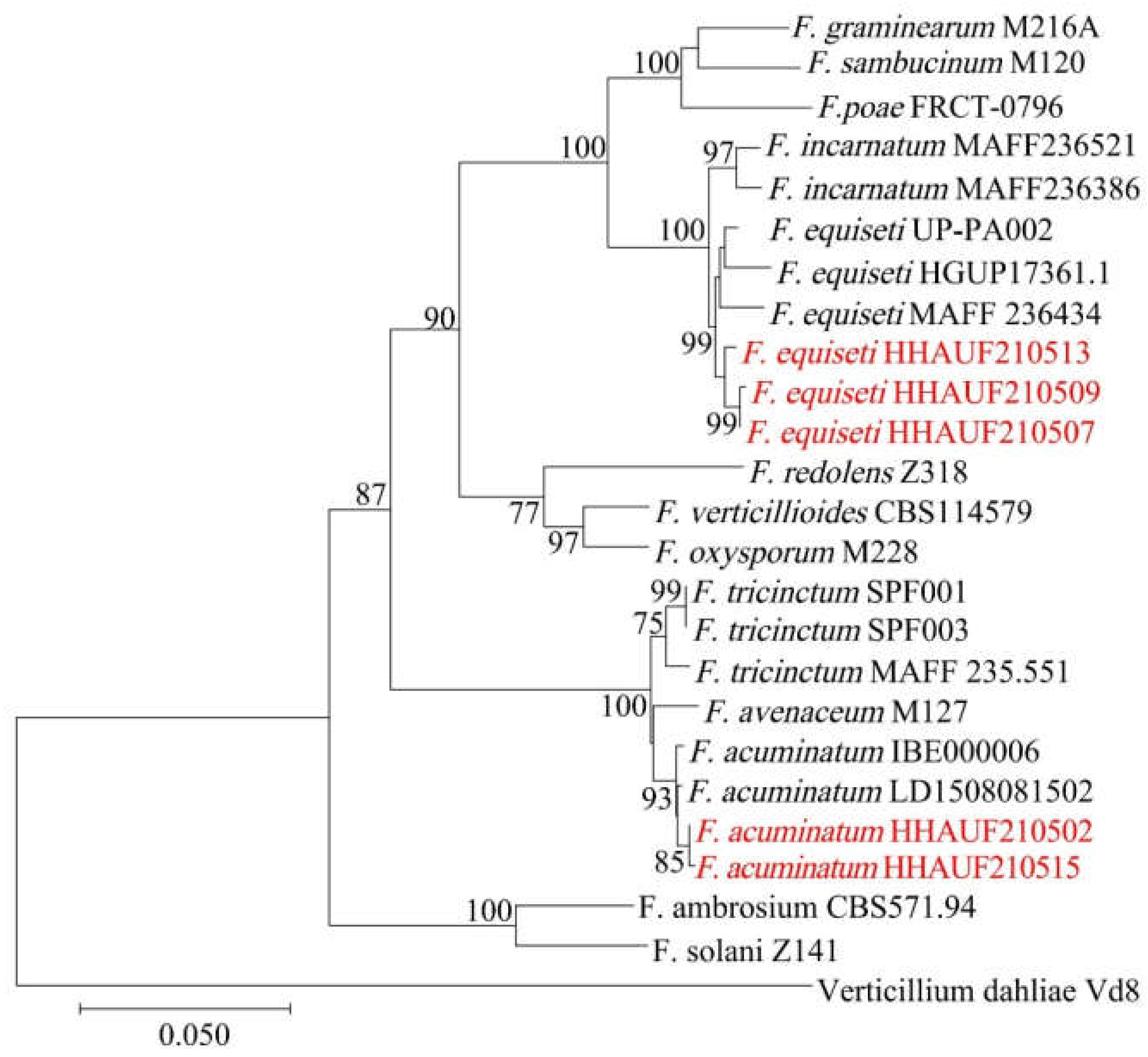
3.3. Phylogenetic Analysis of Pathogens
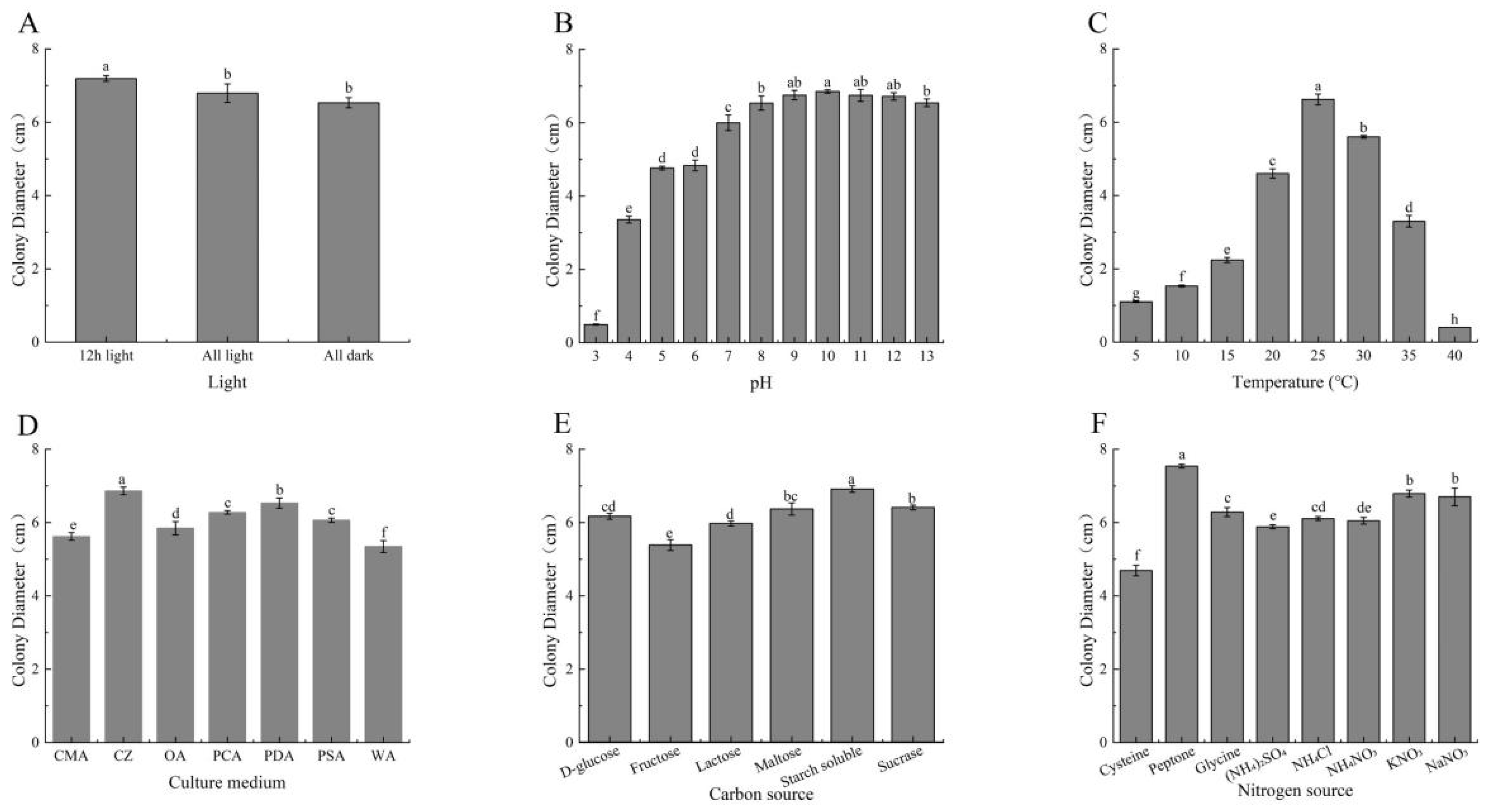
3.4. The Optimal Growth Conditions of F. equiseti
3.5. Activities of Fungicides against F. equiseti and F. acuminatum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, B.; Li, X.; Wang, F.; Li, M.; Zhang, J.; Gu, L.; Zhang, L.; Tu, W.; Zhang, Z. Assaying the potential autotoxins and microbial community associated with Rehmannia glutinosa replant problems based on its ‘autotoxic circle’. Plant Soil. 2016, 407, 307–322. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, M.; Shoyama, Y.; Nishioka, I.; Iwai, H.; Wakimoto, S. Identification of viruses infected in Rehmannia glutinosa Libosch. var purpurea Makino and effect of virus infection on root yield and iridoid glycoside contents. Plant Cell Rep. 1989, 7, 636–638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Li, M.; Jia, Z. Rehmannia glutinosa: Review of botany, chemistry and pharmacology. J. Ethnopharmacol. 2008, 117, 199–214. [Google Scholar] [CrossRef]
- Gong, W.; Zhang, N.; Cheng, G.; Zhang, Q.; He, Y.; Shen, Y.; Zhang, Q.; Zhu, B.; Zhang, Q.; Qin, L. Rehmannia glutinosa Libosch extracts prevent bone loss and architectural deterioration and enhance osteoblastic bone formation by regulating the IGF-1/PI3K/mTOR pathway in streptozotocin-induced diabetic rats. Int. J. Mol. Sci. 2019, 20, 3964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Ma, R.; Wang, L.; Zhu, R.; Liu, H.; Guo, Y.; Zhao, B.; Zhao, S.; Tang, J.; Li, Y.; et al. Rehmanniae Radix in osteoporosis: A review of traditional Chinese medicinal uses, phytochemistry, pharmacokinetics and pharmacology. J. Ethnopharmacol. 2017, 198, 351–362. [Google Scholar] [CrossRef]
- Xia, Z.; Li, C.; Hu, S.; Chen, S.; Xu, Y. The complete chloroplast genome of Chinese medicine cultivar species of Rehmannia glutinosa (Orobanchaceae). Mitochondrial DNA Part B 2021, 6, 290–292. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, X.; Chen, J.; Xu, H.; Zhang, Z. Differential miRNA expression in Rehmannia glutinosa plants subjected to continuous cropping. BMC Plant Biol. 2011, 11, 53. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y. Occurrence and control of common diseases of Rehmannia glutinosa. Henan Agric. 2006, 8, 22. [Google Scholar]
- Wang, R.; Wang, Y.; Yang, Q.; Kang, C.; Li, M. Unraveling the characteristics of the microbial community and potential pathogens in the rhizosphere soil of Rehmannia glutinosa with root rot disease. Appl. Soil Ecol. 2018, 130, 271–279. [Google Scholar] [CrossRef]
- Yan, J.; Ye, H.; Qin, Y.; Wu, G. Investigation on the disease of medicinal plants in Sichuan province and pathogen identification Ⅱ. A list of medicinal plant diseases. Southwest China J. Agric. Sci. 2008, 21, 359–363. [Google Scholar]
- Liu, T. Prevention and control of Rehmannia spot blight and fusarium wilt. Bull. Agric. Sci. Technol. 2002, 10, 31. [Google Scholar]
- Dou, T.; Cai, Y.; Gong, M.; Cui, J.; Du, J.; Wang, Q.; Zhang, Z.; Wang, M.; Liu, X.; Zhang, J. Identification of the pathogen causing Rehmannia glutinosa wilt and screening of fungicides. Acta Phytopathol. Sin. 2022, 52, 638–647. [Google Scholar]
- Du, J.; Nian, W.; Zhou, Z.; Dou, T.; Song, G.; Gu, L.; Li, M.; Wang, M.; Geng, Y.; Zhao, Y.; et al. Leaf Spot Disease Caused by Alternaria alternata on Rehmannia glutinosa in China. Plant Dis. 2020, 104, 3059. [Google Scholar] [CrossRef]
- Guo, M.; Li, B.; Wang, R.; Liu, P.; Chen, Q. Occurrence of dieback disease caused by Fusarium equiseti on Dendrobium officinale in China. Crop Prot. 2020, 137, 105209. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited—ScienceDirect. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef] [Green Version]
- Arif, R.; Pani, D.R.; Zaidi, N.W.; Singh, U.S. PCR-Based identification and characterization of Fusarium sp. associated with mango malformation. Biotechnol. Res. Int. 2011, 2011, 141649. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, S.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microb. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutierrez, S.; Silla-Martinez, J.M.; Gabaldon, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudhir, K.; Glen, S.; Koichiro, T. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar]
- Zhao, Z.; Lu, G. Fusarium sinensis sp. nov., a new species from wheat in China. Mycologia 2008, 100, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Lombard, L.; van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefańczyk, E.; Sobkowiak, S.; Brylińska, M.; Śliwka, J. Diversity of Fusarium spp. associated with dry rot of potato tubers in Poland. Eur. J. Plant Pathol. 2016, 145, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Yonezawa, T.; Sugita-Konishi, Y.; Kamata, Y. Utility of the phylotoxigenic relationships among trichothecene-producing Fusarium species for predicting their mycotoxin-producing potential. Food Addit. Contam. Part A 2013, 30, 1370–1381. [Google Scholar] [CrossRef]
- Nuangmek, W.; Aiduang, W.; Suwannarach, N.; Kumla, J.; Kiatsiriroat, T.; Lumyong, S. First report of fruit rot on cantaloupe caused by Fusarium equiseti in Thailand. J. Gen. Plant Pathol. 2019, 85, 259–300. [Google Scholar] [CrossRef]
- Watanabe, M.; Yonezawa, T.; Lee, K.I.; Kumagai, S.; Hara-Kudo, Y. Molecular phylogeny of the higher and lower taxonomy of the Fusarium genus and differences in the evolutionary histories of multiple genes. BMC Evol. Biol. 2011, 11, 322. [Google Scholar] [CrossRef] [Green Version]
- Aktaruzzaman, M.; Afroz, T.; Lee, Y.G.; Kim, B.S. Morphological and molecular characterization of Fusarium tricinctum causing postharvest fruit rot of pumpkin in Korea. J. Gen. Plant Pathol. 2018, 84, 407–413. [Google Scholar] [CrossRef]
- Watanabe, M.; Yonezawa, T.; Lee, K.-i.; Kumagai, S.; Sugita-Konishi, Y.; Goto, K.; Hara-Kudo, Y. Evaluation of genetic markers for identifying isolates of the species of the genus Fusarium. J. Sci. Food Agric. 2011, 91, 2500–2504. [Google Scholar] [CrossRef]
- Tran, V.T.; Braus-Stromeyer, S.A.; Timpner, C.; Braus, G.H. Molecular diagnosis to discriminate pathogen and apathogen species of the hybrid Verticillium longisporum on the oilseed crop Brassica napus. Appl. Microbiol. Biotechnol. 2013, 97, 4467–4483. [Google Scholar] [CrossRef] [Green Version]
- Machuca, A.; Durán, N. Optimization of some parameters influencing Thermoascus aurantiacus growth: Effects of lignin-related compounds. J. Ind. Microbiol. 1996, 16, 224–229. [Google Scholar] [CrossRef]
- Wang, K.; Jiang, S.; Pu, T.; Fan, L.; Ye, M. Antifungal activity of phenolic monoterpenes and structure-related compounds against plant pathogenic fungi. Nat. Prod. Res. 2019, 33, 1423–1430. [Google Scholar] [CrossRef]
- Cai, Y.; Dou, T.; Gao, F.; Du, J.; Wang, F.; Nian, W.; Wang, M.; Zhang, Z.; Liu, X. Identification and biological characteristics of a pathogen causing leaf blight of Rehmannia glutinosa. China J. Chin. Mater. Med. 2022, 47, 1824–1830. [Google Scholar]
- Swamy, S.D.; Mahadevakumar, S.; Hemareddy, H.B.; Amruthesh, K.N.; Mamathab, S.; Kunjetic, S.G.; Swapnilb, R.; Kumard, T.V.; Lakshmidevi, N. First report of Fusarium equiseti–the incitant of post flowering stalkrot of maize (Zea mays L.) in India. Crop Prot. 2020, 129, 105035. [Google Scholar] [CrossRef]
- Parikh, L.; Kodati, S.; Eskelson, M.J.; Adesemoye, A.O. Identification and pathogenicity of Fusarium spp. In row crops in Nebraska. Crop Prot. 2018, 108, 120–127. [Google Scholar] [CrossRef]
- Avila, C.F.; Moreira, G.M.; Nicolli, C.P.; Gomes, L.B.; Abreu, L.M.; Pfenning, L.H.; Haidukowski, M.; Moretti, A.; Logrieco, A.; Ponte, E. Fusarium incarnatum-equiseti species complex associated with brazilian rice: Phylogeny, morphology and toxigenic potential. Int. J. Food Microbiol. 2019, 306, 108267. [Google Scholar] [CrossRef]
- Hami, A.; Rasool, R.S.; Khan, N.A.; Mansoor, S.; Masoodi, K.Z. Morpho-molecular identification and first report of Fusarium equiseti in causing chilli wilt from Kashmir (Northern Himalayas). Sci. Rep. 2021, 11, 3610. [Google Scholar] [CrossRef]
- Yang, L.; Gao, W.; Wang, W.; Zhang, C.; Wang, Y. First report of stem and root rot of coriander caused by Fusarium equiseti in China. Plant Dis. 2021, 105, 220. [Google Scholar] [CrossRef]
- Moosa, A.; Farzand, A.; Khan, S.A.; Ahmad, T.; Aslam, H.M.U.; Saeed, S.; Shafique, M.; Masroor, A.; Akhtar, S. First report of postharvest Fusarium rot of mandarin (Citrus reticulata ‘Kinnow’) caused by Fusarium equiseti in Pakistan. Plant Dis. 2021, 15, 4170. [Google Scholar] [CrossRef]
- Guo, Z.; Yu, Z.; Li, Q.; Tang, L.; Luo, S. Fusarium species associated with leaf spots of mango in China. Microb. Pathog. 2021, 150, 104736. [Google Scholar] [CrossRef] [PubMed]
- Lazreg, F.; Belabid, L.; Sanchez, J.; Gallego, E.; Garrido-Cardenas, J.A.; Elhaitoum, A. First report of Fusarium equiseti causing damping-off disease on Aleppo pine in Algeria. Plant Dis. 2014, 98, 1268. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Huang, Q.; He, H.; Li, K.; Zhang, Y. First report of black stem of Avicennia marina caused by Fusarium equiseti in China. Plant Dis. 2014, 98, 843. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Punja, Z.K. Molecular and biochemical characterization of defense responses in ginseng (Panax quinquefolius) roots challenged with Fusarium equiseti. Physiol. Mol. Plant Pathol. 2008, 72, 10–20. [Google Scholar] [CrossRef]
- Wen, X.; Qi, H.; Sun, W.; Liu, Y.; Feng, L.; Meng, T.; Han, Z.; Cao, J.; Wang, J. Identification and biological characteristics of the pathogen (Fusarium equiseti) causing shoot blight of Atractylodes chinensis. J. Agric. Sci. Technol. 2020, 22, 115–121. [Google Scholar]
- Jin, S.L.; Guo, C.; Wei, H.; Guo, M.; He, F. Isolation, identification and biological characteristics of Fusarium equiseti from maize ear rot samples. Acta Agrestia Sin. 2014, 22, 182–187. [Google Scholar]
- Li, C.; Fan, S.; Zhang, Y.; Zhang, X.; Luo, J.; Liu, C. Toxicity, bioactivity of triazole fungicide metconazole and its effect on mycotoxin production by Fusarium verticillioides: New perspective from an enantiomeric level. Sci. Total Environ. 2022, 828, 154432. [Google Scholar] [CrossRef]
- Muellenborn, C.; Steiner, U.; Ludwig, M.; Oerke, E. Effect of fungicides on the complex of Fusarium species and saprophytic fungi colonizing wheat kernels. Eur. J. Plant Pathol. 2008, 120, 157–166. [Google Scholar] [CrossRef]
- Mengesha, G.G.; Abebe, S.M.; Lera, Z.T.; Shertore, M.M.; Fedilu, K.B.; Tadesse, Y.B.; Mekonnen, A.A.; Esho, A.G.M.; Cheleko, D.C.; Silassie, A.B.W. Integration of host resistance, fungicides, and spray frequencies for managing Fusarium head blight of bread wheat under field conditions in southern Ethiopia. Heliyon 2021, 7, e07938. [Google Scholar] [CrossRef]
- Cendoya, E.; Nichea, M.J.; Monge, M.d.P.; Zachetti, V.G.L.; Chiacchiera, S.M.; Ramirez, M.L. Effect of fungicides commonly used for Fusarium head blight management on growth and fumonisin production by Fusarium proliferatum. Rev. Argent. Microbiol. 2021, 53, 64–74. [Google Scholar] [CrossRef]
- Mateo, E.M.; Valle-Algarra, F.M.; Mateo, R.; Jimenez, M.; Magan, N. Effect of fenpropimorph, prochloraz and tebuconazole on growth and production of T-2 and HT-2 toxins by Fusarium langsethiae in oat-based medium. Int. J. Food Microbiol. 2011, 151, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Malandrakis, A.; Daskalaki, E.R.; Skiada, V.; Papadopoulou, K.K.; Kavroulakis, N. A Fusarium solani endophyte vs fungicides: Compatibility in a Fusarium oxysporum f.sp. radicis-lycopersici-tomato pathosystem. Fungal Biol. 2018, 122, 1215–1221. [Google Scholar] [CrossRef]
- Liao, X.; Hou, W.; Tang, X.; Zhao, Y. Identification of the roselle root rot pathogen and its sensitivity to different fungicides. Arab. J. Chem. 2022, 15, 103932. [Google Scholar] [CrossRef]
- Ghosal, D.; Tarafder, M.; Datta, B. Fusarium oxysporum isolates showed varied benzimidazole fungicides sensitivity under low temperature and heavy metal stress. Rhizosphere 2020, 15, 100227. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | GenBank Accession Numbers | Reference | ||
|---|---|---|---|---|---|
| ITS | Tub | EF1-α | |||
| F. acuminatum | IBE000006 | EF531232 | EF531244 | EF531237 | [24] |
| F. acuminatum | LD1508081502 | MF523230 | MF523225 | MF523228 | NP |
| F. acuminatum | HHAUF210502 | MZ351204 | MZ366376 | MZ366377 | • |
| F. acuminatum | HHAUF210515 | OP846525 | OP837541 | OP837544 | • |
| F. ambrosium | CBS 571.94 | KM231801 | KM232063 | KM231929 | [25] |
| F. avenaceum | M127 | KP265352 | KP710634 | KP674195 | [26] |
| F. equiseti | HHAUF210507 | MZ543968 | MZ547046 | MZ547045 | • |
| F. equiseti | HHAUF210509 | OP846523 | OP837539 | OP837542 | • |
| F. equiseti | HHAUF210513 | OP846524 | OP837540 | OP837543 | • |
| F. equiseti | HGUP17361.1 | MK069606 | MK069604 | MK069605 | NP |
| F. equiseti | MAFF 236434 | AB586999 | AB587047 | AB674277 | [27] |
| F. equiseti | UP-PA002 | MH521295 | MH521296 | MH521297 | [28] |
| F. graminearum | M216A | KP295509 | KP765707 | KP400687 | [26] |
| F. incarnatum | MAFF 236386 | AB820720 | AB820712 | AB820704 | [27] |
| F. incarnatum | MAFF 236521 | AB586988 | AB587036 | AB674267 | [29] |
| F. oxysporum | M228 | KP264651 | KP674278 | KP400703 | [26] |
| F. poae | FRCT-0796 | AB586983 | AB587072 | AB674301 | [29] |
| F. redolens | Z318 | KP264660 | KP674235 | KP400713 | [26] |
| F. sambucinum | M120 | KP265348 | KP710630 | KP674191 | [26] |
| F. solani | Z141 | KP265362 | KP710644 | KP674205 | [26] |
| F. tricinctum | SPF001 | MG990937 | MG990938 | MG990939 | [30] |
| F. tricinctum | SPF003 | MG704912 | MG704913 | MG704914 | [30] |
| F. tricinctum | MAFF 235.551 | AB587030 | AB587079 | AB674262 | [31] |
| F. verticillioides | CBS 114579 | KU604025 | KU603860 | KU711696 | NP |
| Verticillium dahliae | Vd8 | HE972024 | KF555285 | KM408514 | [32] |
| Medium | Formulation |
|---|---|
| PDA | peeled potato, 200 g; glucose, 20 g; agar powder, 20 g, distilled water to 1 L |
| PSA | peeled potato, 200 g; sucrose, 20 g; agar powder, 20 g, distilled water to 1 L |
| PCA | peeled potato, 20 g; peeled carrot, 20 g; agar powder, 20 g, distilled water to 1 L |
| OA | oatmeal, 30 g; agar powder, 20 g, distilled water to 1 L |
| CMA | corn flour, 50 g; agar powder, 15 g, distilled water to 1 L |
| CZ | NaNO3, 3 g; K2HPO4•3H₂O, 1 g; MgSO4•7H2O, 0.5 g; KCl, 0.5 g; FeSO4•7H₂O, 0.01 g; sucrose, 30 g; agar powder, 20 g, distilled water to 1 L |
| WA | agar powder, 20 g, distilled water to 1 L |
| Fungicides | Manufacturer | Concentration Gradient (mg·L−1) |
|---|---|---|
| Thiophanate-methyl (97% a.i.) | Anhui Guangxin Agrochemical Co., Ltd. | 5, 10, 20, 40, 80, 160 |
| Boscalid (97% a.i.) | Hebei Lansheng Biotechnology Co., Ltd. | 12.5, 25, 50, 100, 200, 400 |
| Prochloraz (97% a.i.) | Jiangxi Huihe Chemical Co., Ltd. | 0.25, 0.5, 1, 2, 4, 8 |
| Flusilazole (97% a.i.) | Shandong Audley Chemical Co., Ltd. | 0.125, 0.25, 0.5, 1, 2, 4 |
| Tebuconazole (97% a.i.) | Xinyi Youlian Chemical Co., Ltd. | 0.25, 0.5, 1, 2, 4, 8 |
| Trifloxystrobin (97% a.i.) | Nanjing Kangmanlin Chemical Co., Ltd. | 1, 2, 4, 8, 16, 32 |
| Pyraclostrobin (98% a.i.) | Hebei Chengyue Chemical Co., Ltd. | 0.25, 0.5, 1, 2, 4, 8 |
| Chlorothalonil (98% a.i.) | Anhui Guangxin Agrochemical Co., Ltd. | 12.5, 25, 50, 100, 200, 400 |
| Fungicides | Regression Equation | EC50 (mg·L−1) | Correlation Coefficient | 95% Confidence Intervals (mg·L−1) |
|---|---|---|---|---|
| Thiophanate-methyl (97% a.i.) | Y = 2.704 + 1.484X | 35.261 | 0.970 | 29.552~42.074 |
| Boscalid (97% a.i.) | Y = 3.637 + 0.590X | 204.807 | 0.990 | 118.740~353.260 |
| Prochloraz (97% a.i.) | Y = 6.140 + 1.329X | 0.139 | 0.986 | 0.082~0.234 |
| Flusilazole (97% a.i.) | Y = 5.769 + 1.548X | 0.318 | 0.980 | 0.257~0.395 |
| Tebuconazole (97% a.i.) | Y = 5.817 + 1.146X | 0.194 | 0.991 | 0.118~0.317 |
| Trifloxystrobin (97% a.i.) | Y = 4.548 + 0.323X | 25.158 | 0.963 | 7.797~81.173 |
| Pyraclostrobin (98% a.i.) | Y = 4.711 + 0.432X | 4.668 | 0.995 | 2.175~10.020 |
| Chlorothalonil (98% a.i.) | Y = 3.266 + 0.743X | 215.960 | 0.987 | 163.783~340.970 |
| Fungicides | Regression Equation | EC50 (mg·L−1) | Correlation Coefficient | 95% Confidence Intervals (mg·L−1) |
|---|---|---|---|---|
| Thiophanate-methyl (97% a.i.) | Y = 2.582 + 1.539X | 37.249 | 0.974 | 19.764~70.204 |
| Boscalid (97% a.i.) | Y = 2.317 + 1.199X | 173.158 | 0.994 | 64.197~467.060 |
| Prochloraz (97% a.i.) | Y = 5.606 + 0.667X | 0.123 | 0.980 | 0.050~3.313 |
| Flusilazole (97% a.i.) | Y = 5.186 + 1.142X | 0.688 | 0.987 | 0.313~1.511 |
| Tebuconazole (97% a.i.) | Y = 5.237 + 1.131X | 0.617 | 0.995 | 0.225~1.690 |
| Trifloxystrobin (97% a.i.) | Y = 4.968 + 0.774X | 21.505 | 0.983 | 3.641~127.012 |
| Pyraclostrobin (98% a.i.) | Y = 4.723 + 0.799X | 2.223 | 0.987 | 0.686~7.203 |
| Chlorothalonil (98% a.i.) | Y = 3.298 + 0.785X | 147.666 | 0.966 | 39.031~558.666 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dou, T.; Cai, Y.; Song, X.; Gao, F.; Zhao, Y.; Du, J.; Wang, F.; Li, X.; An, S.; Yin, X.; et al. Characterization and Fungicide Screening of a New Pathogen That Causes Leaf Spot on Rehmannia glutinosa. Agriculture 2023, 13, 301. https://doi.org/10.3390/agriculture13020301
Dou T, Cai Y, Song X, Gao F, Zhao Y, Du J, Wang F, Li X, An S, Yin X, et al. Characterization and Fungicide Screening of a New Pathogen That Causes Leaf Spot on Rehmannia glutinosa. Agriculture. 2023; 13(2):301. https://doi.org/10.3390/agriculture13020301
Chicago/Turabian StyleDou, Tao, Yubiao Cai, Xuhong Song, Futao Gao, Yajun Zhao, Jiafang Du, Fengqing Wang, Xuanzhen Li, Shiheng An, Xinming Yin, and et al. 2023. "Characterization and Fungicide Screening of a New Pathogen That Causes Leaf Spot on Rehmannia glutinosa" Agriculture 13, no. 2: 301. https://doi.org/10.3390/agriculture13020301