
Survival Analysis of the Green Lacewing, Chrysoperla externa (Hagen) Exposed to Neem-Based Products

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chrysoperla Externa Rearing and Neem-Based Products
2.2. Bioassays
2.3. Statistical Analysis
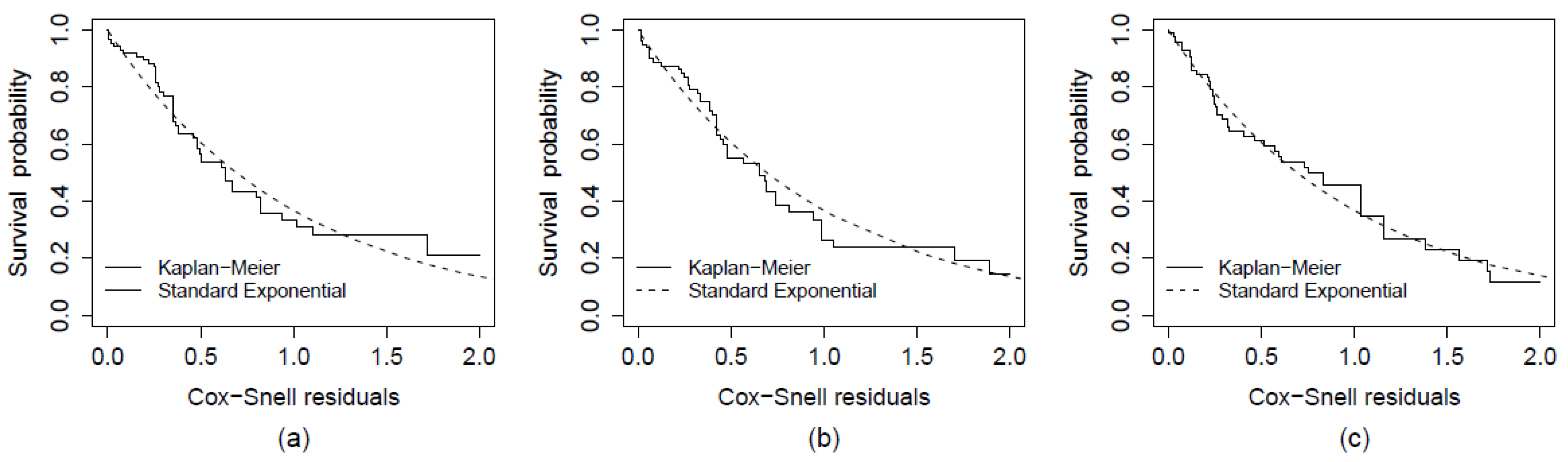
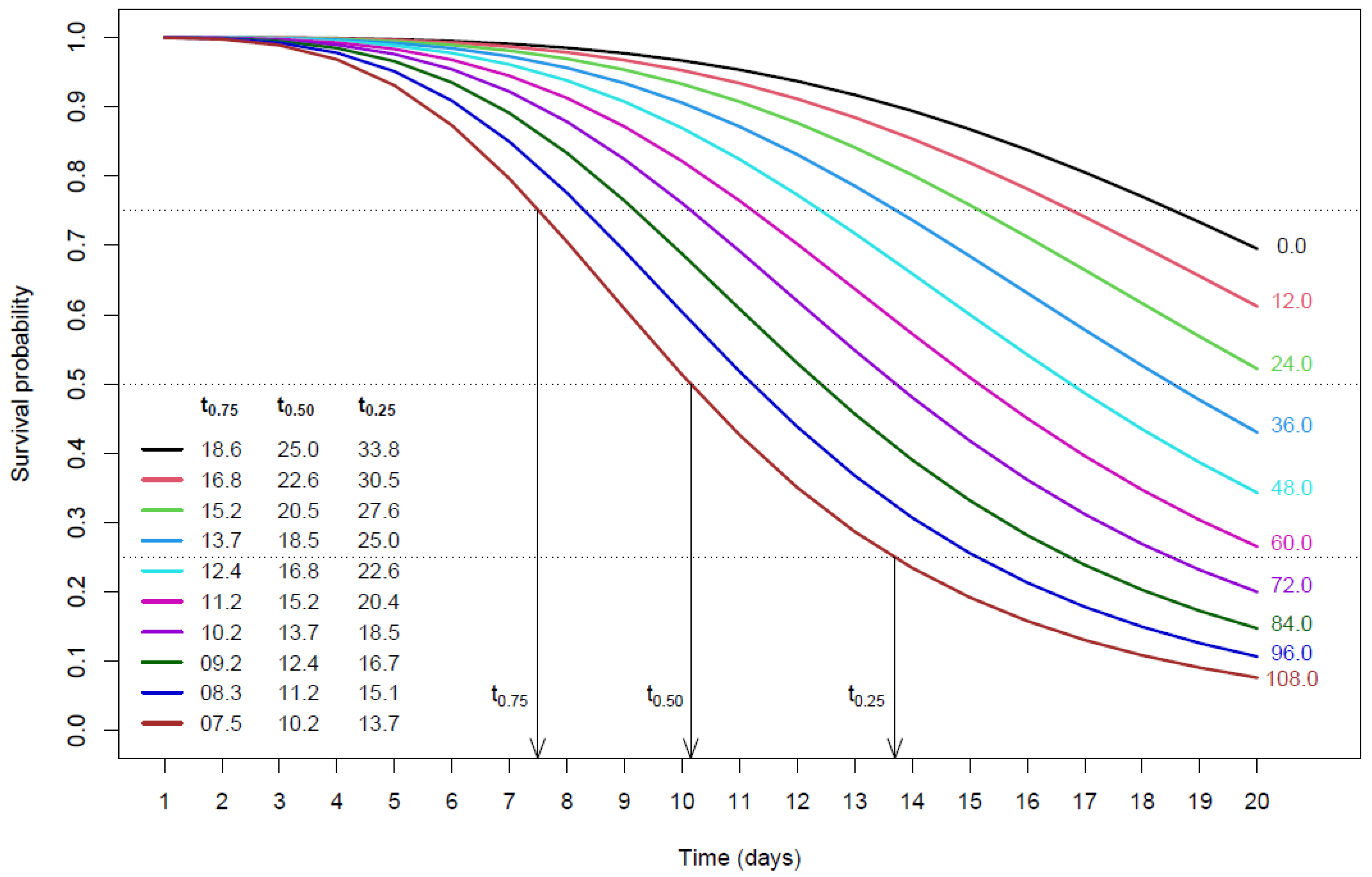
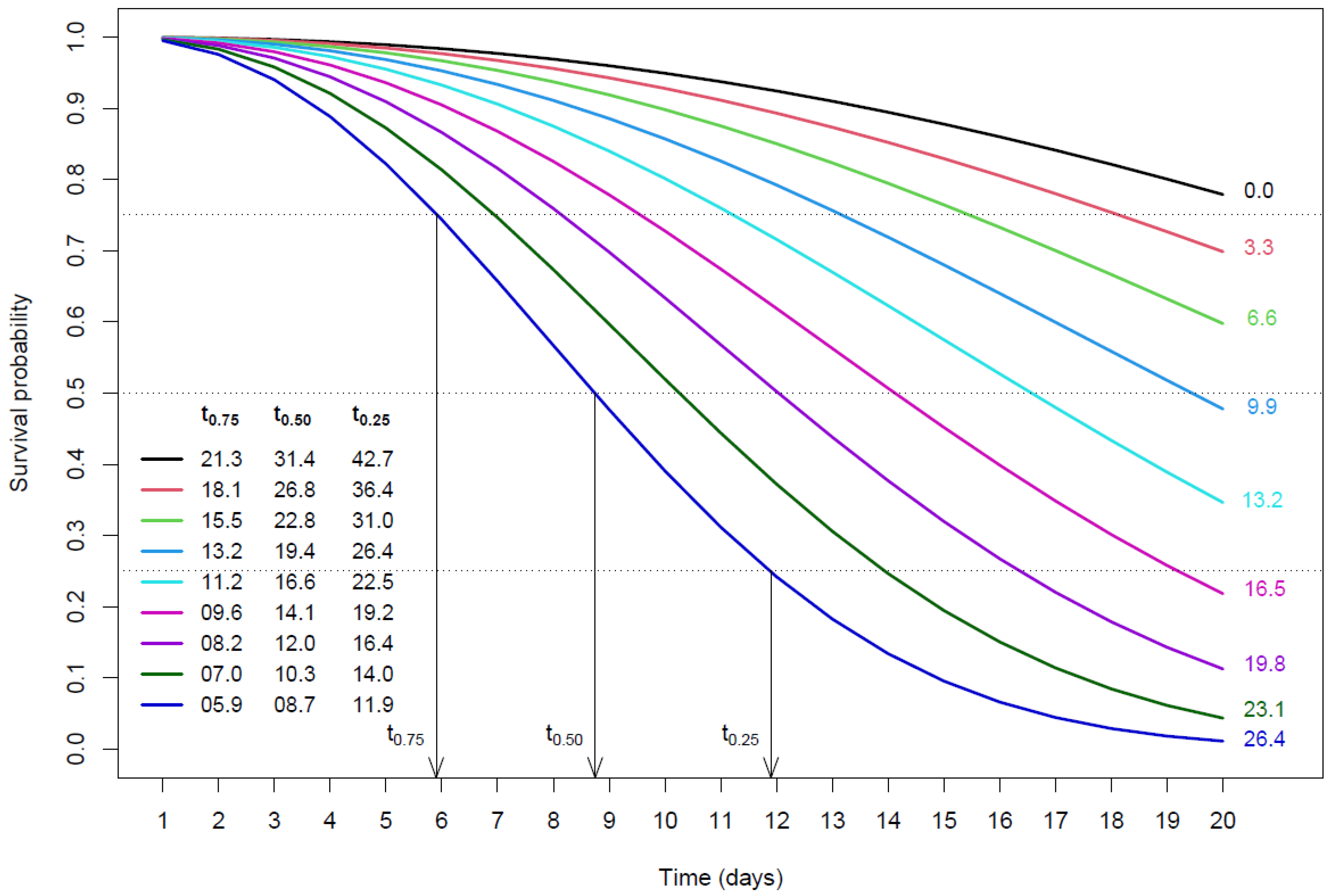
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- ICO—International Coffee Organization. Trade Statistics. 2022. Available online: http://www.ico.org/trade_statistics.asp (accessed on 14 December 2022).
- Davis, A.P.; Gargiulo, R.; Fay, M.F.; Sarmu, D.; Haggar, J. Lost and found: Coffea stenophylla and C. affinis, the forgotten coffee crop species of West Africa. Front. Plant Sci. 2020, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Huded, A.K.C.; Jingade, P.; Bychappa, M.; Mishra, M.K. Genetic diversity and population structure analysis of coffee (Coffea canephora) germplasm collections in Indian Gene Bank employing SRAP and SCoT markers. Int. J. Fruit Sci. 2020, 20, 757–784. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Auad, A.M.; Mendes, S.M.; Frizzas, M.R. Economic impact of exotic insect pests in Brazilian agriculture. J. Appl. Entomol. 2013, 137, 1–15. [Google Scholar] [CrossRef]
- Milligan, M.C.; Johnson, M.D.; Garfinkel, M.; Smith, C.J.; Njoroge, P. Quantifying pest control services by birds and ants in Kenyan coffee farms. Biol. Conserv. 2016, 194, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Avelino, J.; Cerda, R.; Willocquet, L.; Savary, S. Multiple-disease system in coffee: From crop loss assessment to sustainable management. Annu. Rev. Phytopat. 2018, 56, 611–635. [Google Scholar] [CrossRef]
- Cure, J.R.; Rodríguez, D.; Gutierrez, A.P.; Ponti, L. The coffee agroecosystem: Bio-economic analysis of coffee berry borer control (Hypothenemus hampei). Sci. Rep. 2020, 10, 12262. [Google Scholar] [CrossRef]
- Le Pelley, R.H. Las Plagas del Cafeto; Editorial Labor S.A.: Barcelona, Espanha, 1968; 693p. [Google Scholar]
- Reis, P.R.; Souza, J.C.; Venzon, M. Manejo ecológico das principais pragas do cafeeiro. Inf. Agropec. 2002, 23, 83–99. [Google Scholar]
- Vega, F.E.; Infante, F.; Castillo, A.; Jaramillo, J. The coffee berry borer Hypothenemus hampei (Ferrari) (Coleoptera: Curculionidae): A short review, with recent findings and future research directions. Terr. Arthropod. Rev. 2009, 22, 129–147. [Google Scholar] [CrossRef]
- Venzon, M. Agro-Ecological Management of Coffee Pests in Brazil. Front. Sustain. Food Syst. 2021, 5, 721117. [Google Scholar] [CrossRef]
- Fragoso, D.B.; Guedes, R.N.C.; Ladeira, J.A. Seleção na evolução de resistência a organofosforados em Leucoptera coffeella (Guérin-Mèneville) (Lepidoptera: Lyonetiidae). Neotrop. Entomol. 2003, 32, 329–334. [Google Scholar] [CrossRef]
- Reis, M.R.; Fernandes, F.L.; Lopes, E.A.; Gorri, J.E.R.; Alves, F.M. Pesticide residues in coffee agroecosystems. In Coffee in Health and Disease Prevention; Preedy, V.R., Ed.; Academic Press: London, UK, 2015; pp. 235–244. [Google Scholar] [CrossRef]
- Hutter, H.P.; Kundi, M.; Lemmerer, K.; Poteser, M.; Weitensfelder, L.; Wallner, P.; Moshammer, H. Subjective symptoms of male workers linked to occupational pesticide exposure on coffee plantations in the Jarabacoa region, Dominican Republic. Int. J. Environ. Res. Public Health 2018, 15, 2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leite, S.A.; Santos, M.P.; Costa, D.R.; Moreira, A.A.; Guedes, R.N.C.; Castellani, M.A. Time-concentration interplay in insecticide resistance among populations of the Neotropical coffee leaf miner, Leucoptera coffeela. Agric. For. Entomol. 2021, 23, 232–241. [Google Scholar] [CrossRef]
- Leite, S.A.; Santos, M.P.; Resende-Silva, G.A.; Costa, D.R.; Moreira, A.A.; Lemos, O.L.; Guedes, R.N.C.; Castellani, M.A. Area-wide survey of chlorantraniliprole resistance and control failure likelihood of the Neotropical coffee leaf miner Leucoptera coffeella (Lepidoptera: Lyonetiidae). J. Econ. Entomol. 2020, 113, 1399–1410. [Google Scholar] [CrossRef]
- Botti, J.M.C.; Martins, E.F.; Franzin, M.L.; Venzon, M. Predation of coffee berry borer by a green lacewing. Neotrop. Entomol. 2022, 51, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Ecole, C.C.; Silva, R.A.; Louzada, J.N.C.; Moraes, J.C.; Barbosa, L.R.; Ambrogi, B.G. Predação de ovos, larvas e pupas do bicho-mineiro-do-cafeeiro, Leucoptera coffeella (Guérin-Menèville and Perrottet, 1842) (Lepidoptera: Lyonetiidae) por Chrysoperla externa (Hagen, 1861) (Neuroptera: Chrysopidae). Cienc. Agrotec. 2002, 26, 318–324. [Google Scholar]
- Venzon, M.; Lemos, F.; Sarmento, R.A.; Rosado, M.C.; Pallini, A. Predação por coccinelídeos e crisopídeo influenciada pela teia de Tetranychus evansi. Pesq. Agropec. Bras. 2009, 44, 1086–1091. [Google Scholar] [CrossRef] [Green Version]
- Martins, E.F.; Franzin, M.L.; Perez, A.L.; Schmidt, J.M.; Venzon, M. Is Ceraeochrysa cubana a coffee leaf miner predator? Biol. Control. 2021, 160, 104691. [Google Scholar] [CrossRef]
- Kilani-Morakchi, S.; Morakchi-Goudjil, H.; Sifi, K. Azadirachtin-Based Insecticide: Overview, Risk Assessments, and Future Directions. Front. Agron. 2021, 3, 676208. [Google Scholar] [CrossRef]
- Martinez, S.S.; Meneguim, A.M. Redução da oviposição e da sobrevivência de ovos de Leucoptera coffeella causadas pelo óleo emulsionável de nim. Manejo Integr. Plagas Y Agroecol. 2003, 67, 58–62. [Google Scholar]
- Venzon, M.; Rosado, M.C.; Fadini, M.A.M.; Ciociola, A.I., Jr.; Pallini, A. The potential of NeemAzal for the control of coffee leaf pests. Crop Prot. 2005, 24, 213–219. [Google Scholar] [CrossRef]
- Depieri, R.A.; Martinez, S.S.; Menezes Jr, A.O. Compatibility of the fungus Beauveria bassiana (Bals.) Vuill. (Deuteromycetes) with extracts of neem seeds and leaves and the emulsible oil. Neotrop. Entomol. 2005, 34, 601–606. [Google Scholar] [CrossRef] [Green Version]
- Colosimo, E.A.; Giolo, S.R. Análise de Sobrevivência Aplicada; Editora Blücher: São Paulo, Brasil, 2006; 392p. [Google Scholar]
- Cordeiro, E.; Corrêa, A.; Venzon, M.; Guedes, R.N.C. Insecticide survival and behavioral avoidance in the lacewings Chrysoperla externa and Ceraeochrysa cubana. Chemosphere 2010, 81, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- Venzon, M.; Oliveira, R.M.; Perez, A.L.; Rodríguez-Cruz, F.A.; Martins Filho, S. Lime sulfur toxicity to broad mite, to its host plants and to natural enemies. Pest Manag. Sci. 2013, 69, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.R.; Sarmento, R.A.; Venzon, M.; Santos, G.R.; Silveira, M.C.A.C.; Tschoeke, P.H. Lethal and sublethal effects of neem on Aphis gossypii and Cycloneda sanguinea in watermelon. Acta Sci. Agron. 2015, 37, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Amaral, R.L.; Venzon, M.; Martins Filho, S.; Lima, M.A.P. Does ingestion of neem-contaminated diet cause mortality of honey bee larvae and foragers? J. Apic. Res. 2015, 4, 405–410. [Google Scholar] [CrossRef]
- Rimoldi, F.; Fogel, M.N.; Ronco, A.E.; Schneider, M.I. Comparative susceptibility of two Neotropical predators, Eriopis connexa and Chrysoperla externa, to acetamiprid and pyriproxyfen: Short and long-term effects after egg exposure. Environ. Pollut. 2017, 231, 1042–1050. [Google Scholar] [CrossRef]
- Brevik, K.; Schoville, S.D.; Mota-Sanchez, D.; Chen, Y.H. Pesticide durability and the evolution of resistance: A novel application of survival analysis. Pest Manag. Sci. 2018, 74, 1953–1963. [Google Scholar] [CrossRef]
- Francesena, N.; Schneider, M.I. Selectivity assessment of two biorational insecticides, azadirachtin and pyriproxyfen, in comparison to a neonicotinoid, acetamiprid, on pupae and adults of a Neotropical strain Eretmocerus mundus Mercet. Chemosphere 2018, 206, 349–358. [Google Scholar] [CrossRef]
- Duarte, A.D.F.; Pazini, J.B.; Duarte, J.L.P.; Silva, L.R.; Cunha, U.S. Compatibility of pesticides used in strawberry crops with predatory mites Stratiolaelaps scimitus (Womersley) and Cosmolaelaps brevistilis (Karg). Ecotoxicology 2020, 29, 148–155. [Google Scholar] [CrossRef]
- Silva, W.M.; Martínez, L.C.; Plata-Rueda, A.; Serrão, J.E.; Zanuncio, J.C. Exposure to insecticides causes effects on survival, prey consumption, and histological changes in the midgut of the predatory bug, Podisus nigrispinus (Hemiptera: Pentatomidae). Environ. Sci. Pollut. Res. 2021, 28, 57449–57458. [Google Scholar] [CrossRef]
- Venzon, M.; Rosado, M.C.; Euzébio, D.E.; Souza, B.; Schoereder, J.H. Suitability of leguminous cover crop pollens as food source for the green lacewing Chrysoperla externa (Hagen) (Neuroptera: Chrysopidae). Neotrop. Entomol. 2006, 35, 371–376. [Google Scholar] [CrossRef] [Green Version]
- Potter, C. An improved laboratory apparatus for applying direct sprays and surface films, with data on the electrostatic charge on atomized spray fluids. Ann. Appl. Biol. 1952, 39, 1–29. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; pp. 70–76. [Google Scholar]
- Symonds, M.R.E.; Moussalli, A. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav. Ecol. Sociobiol. 2011, 65, 13–21. [Google Scholar] [CrossRef]
- Cox, D.R.; Snell, E.J. A general definition of residuals. J. R. Stat. Soc. Ser. B (Methodol.) 1968, 30, 248–265. [Google Scholar] [CrossRef]
- Therneau, T. A Package for Survival Analysis in R. R package version 3.5-0. 2023. Available online: https://CRAN.R-project.org/package=survival (accessed on 4 December 2022).
- Clavel, J.; Escarguel, G.; Merceron, G. mvMORPH: An R package for fitting multivariate evolutionary models to morphometric data. Methods Ecol. Evol. 2015, 6, 1311–1319. [Google Scholar] [CrossRef]
- Haman, J.; Avery, M. ciTools: Confidence or Prediction Intervals, Quantiles, and Probabilities for Statistical Models. R package version 0.6.1. 2020. Available online: https://CRAN.R-project.org/package=ciTools (accessed on 4 December 2022).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.r-project.org/ (accessed on 23 December 2022).
- Portet, S. A primer on model selection using the Akaike Information Criterion. Infect. Dis. Model. 2020, 5, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.P.; Moeschberger, M.L. Survival Analysis: Techniques for Censored and Truncated Data, 2nd ed.; Springer: New York, NY, USA, 2003; pp. 358–359. [Google Scholar]
- Souza, J.R.; Moreira, L.B.; Lima, L.L.R.; Silva, T.G.; Braga, P.P.M.; Carvalho, A.D. Susceptibility of Chrysoperla externa (Hagen, 1861) (Neuroptera: Crysopidae) to insecticides used in coffee crops. Ecotoxicology 2020, 29, 1306–1314. [Google Scholar] [CrossRef]
- Ribeiro, A.L.P.; Lúcio, A.D.; Costa, E.C.; Bolzan, A.R.; Jovanowichs, R.; Riffel, C.T. Desenvolvimento de Chrysoperla externa alimentada na fase larval com ovos de Bonagota cranaodes. Ciência Rural. 2011, 41, 1571–1577. [Google Scholar] [CrossRef] [Green Version]
- Nagini, S. Neem Limonoids as Anticancer Agents: Modulation of Cancer Hallmarks and Oncogenic Signaling. Enzymes 2014, 36, 131–147. [Google Scholar] [CrossRef]
- Gupta, S.C.; Prasad, S.; Tyagi, A.K.; Kunnumakkara, A.B.; Aggarwal, B.B. (2017). Neem (Azadirachta indica): An indian traditional panacea with modern molecular basis. Phytomedicine 2017, 34, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Venzon, M.; Togni, P.H.; Perez, A.L.; Oliveira, J.M. Control of two-spotted spider mites with neem-based products on a leafy vegetable. Crop Prot. 2020, 128, 105006. [Google Scholar] [CrossRef]
- MAPA Ministério da Agricultura Pecuária e Abastecimento. Available online: https://agrofit.agricultura.gov.br/agrofit_cons/!ap_produto_form_detalhe_cons?p_id_produto_formulado_tecnico=8413&p_tipo_janela=NEW (accessed on 26 December 2022).
- Miranda, M.S. Técnicas não-paramétricas e paramétricas usadas na análise de sobrevivência de Chrysoperla externa (Neuroptera: Chrysopidae). Master’s Thesis, Universidade Federal de Viçosa, Viçosa, MG, Brazil, 13 March 2012. [Google Scholar]
- Brito, H.M.; Gondim Junior, M.G.C.; Oliveira, J.V.D.; Câmara, C.A.G.D. Toxicity of the Natuneem® over the Tetranychus urticae Koch (Acari: Tetranychidae) and mites predators from the family Phytoseiidae. Cienc. Agrotecnol. 2006, 30, 685–691. [Google Scholar] [CrossRef]
- Soto, A.; Venzon, M.; Oliveira, R.M.; Oliveira, H.G.; Pallini, A. Alternative control of Tetranychus evansi Baker & Pritchard (Acari: Tetranychidae) on tomato plants grown in greenhouses. Neotrop. Entomol. 2010, 39, 638–644. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models | Probability Density Function 1 | Survival Function |
|---|---|---|
| Exponential | ||
| Weibull | ||
| Log-normal | ||
| Log-logistic | ||
| Logistic | ||
| Gaussian |
| Parameters | Exponential | Weibull | Log-Normal | Log-Logistic | Logistic | Gaussian | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | SE | Estimate | SE | Estimate | SE | Estimate | SE | Estimate | SE | Estimate | SE | |
| Azamax® | ||||||||||||
| 4.0726 | 0.2992 | 3.4303 | 0.1288 | 3.2001 | 0.1241 | 3.2202 | 0.1079 | 22.5068 | 1.4292 | 22.2546 | 1.5065 | |
| −0.0158 | 0.0042 | −0.0089 | 0.0018 | −0.0078 | 0.0019 | −0.0084 | 0.0016 | −0.1143 | 0.0205 | −0.1055 | 0.0227 | |
| 1.0000 | 0.0000 | 0.3853 | 0.0450 | 0.5270 | 0.0564 | 0.2725 | 0.0333 | 3.6114 | 0.4361 | 6.4241 | 0.6961 | |
| Log L | −212.30 | --- | −190.30 | --- | −190.90 | --- | −187.70 | --- | −196.40 | --- | −196.90 | --- |
| AIC | 428.67 | --- | 386.56 | --- | 387.76 | --- | 381.42 | --- | 398.74 | --- | 399.87 | --- |
| 0.00 | --- | 0.07 | --- | 0.04 | --- | 0.89 | --- | 0.00 | --- | 0.00 | --- | |
| Organic neem® | ||||||||||||
| 4.2724 | 0.3264 | 3.6093 | 0.1632 | 3.3696 | 0.1618 | 3.3835 | 0.1441 | 24.1767 | 1.6901 | 23.8370 | 1.7897 | |
| −0.0788 | 0.0181 | −0.0485 | 0.0090 | −0.0453 | 0.0096 | −0.0459 | 0.0084 | −0.5847 | 0.0967 | −0.5540 | 0.1069 | |
| 1.0000 | 0.0000 | 0.4425 | 0.0548 | 0.6297 | 0.0703 | 0.3320 | 0.0429 | 3.8997 | 0.4950 | 7.0087 | 0.7936 | |
| Log L | −188.00 | --- | −172.70 | --- | −174.70 | --- | −172.80 | --- | −180.40 | --- | −181.10 | --- |
| AIC | 379.98 | --- | 351.45 | --- | 355.50 | --- | 351.54 | --- | 366.73 | --- | 368.23 | --- |
| 0.00 | --- | 0.48 | --- | 0.06 | --- | 0.46 | --- | 0.00 | --- | 0.00 | --- | |
| Natuneem® | ||||||||||||
| 4.9722 | 0.3716 | 4.0718 | 0.2205 | 3.8558 | 0.1939 | 3.7988 | 0.1880 | 28.5799 | 2.0402 | 28.8093 | 2.0433 | |
| −0.0867 | 0.0148 | −0.0549 | 0.0087 | −0.0563 | 0.0083 | −0.0538 | 0.0081 | −0.6350 | 0.0855 | −0.6355 | 0.0874 | |
| 1.0000 | 0.0000 | 0.5041 | 0.0603 | 0.6591 | 0.0712 | 0.3754 | 0.0453 | 4.1101 | 0.4918 | 7.0062 | 0.7705 | |
| Log L | −198.00 | --- | −185.70 | --- | −183.80 | --- | −184.30 | --- | −195.40 | --- | −194.40 | --- |
| AIC | 399.91 | --- | 377.45 | --- | 373.62 | --- | 374.51 | --- | 396.80 | --- | 394.73 | --- |
| 0.00 | --- | 0.08 | --- | 0.56 | --- | 0.36 | --- | 0.00 | --- | 0.00 | --- | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins Filho, S.; Duarte, M.L.; Venzon, M. Survival Analysis of the Green Lacewing, Chrysoperla externa (Hagen) Exposed to Neem-Based Products. Agriculture 2023, 13, 292. https://doi.org/10.3390/agriculture13020292
Martins Filho S, Duarte ML, Venzon M. Survival Analysis of the Green Lacewing, Chrysoperla externa (Hagen) Exposed to Neem-Based Products. Agriculture. 2023; 13(2):292. https://doi.org/10.3390/agriculture13020292
Chicago/Turabian StyleMartins Filho, Sebastião, Marciel Lelis Duarte, and Madelaine Venzon. 2023. "Survival Analysis of the Green Lacewing, Chrysoperla externa (Hagen) Exposed to Neem-Based Products" Agriculture 13, no. 2: 292. https://doi.org/10.3390/agriculture13020292