
Intercropped Plants Provide a Reservoir of Predatory Mites in Coffee Crop
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
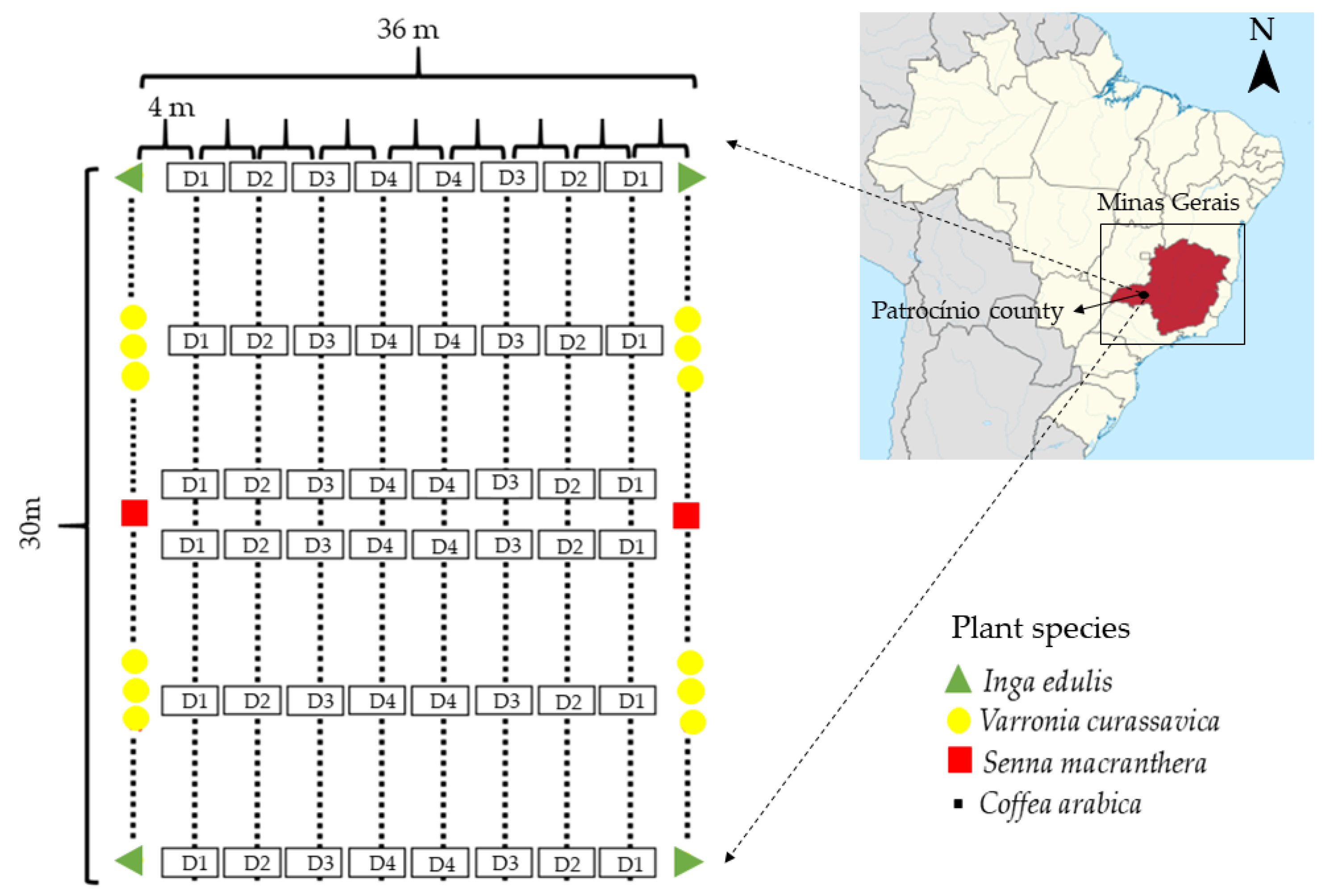
2.1. Study Area
2.2. Data Collection
2.3. Statistical Analyses
3. Results
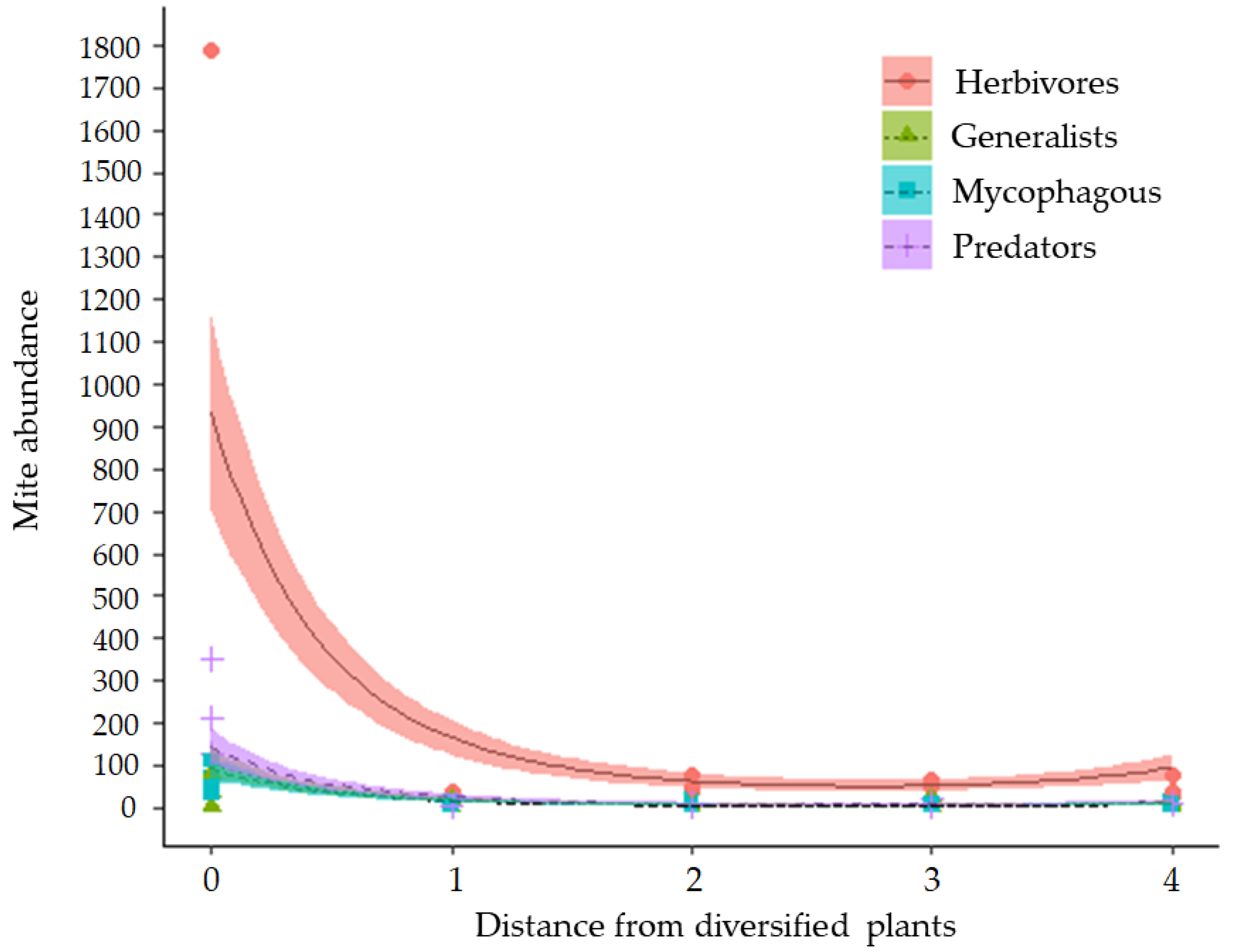
3.1. Abundance and Richness of Mites
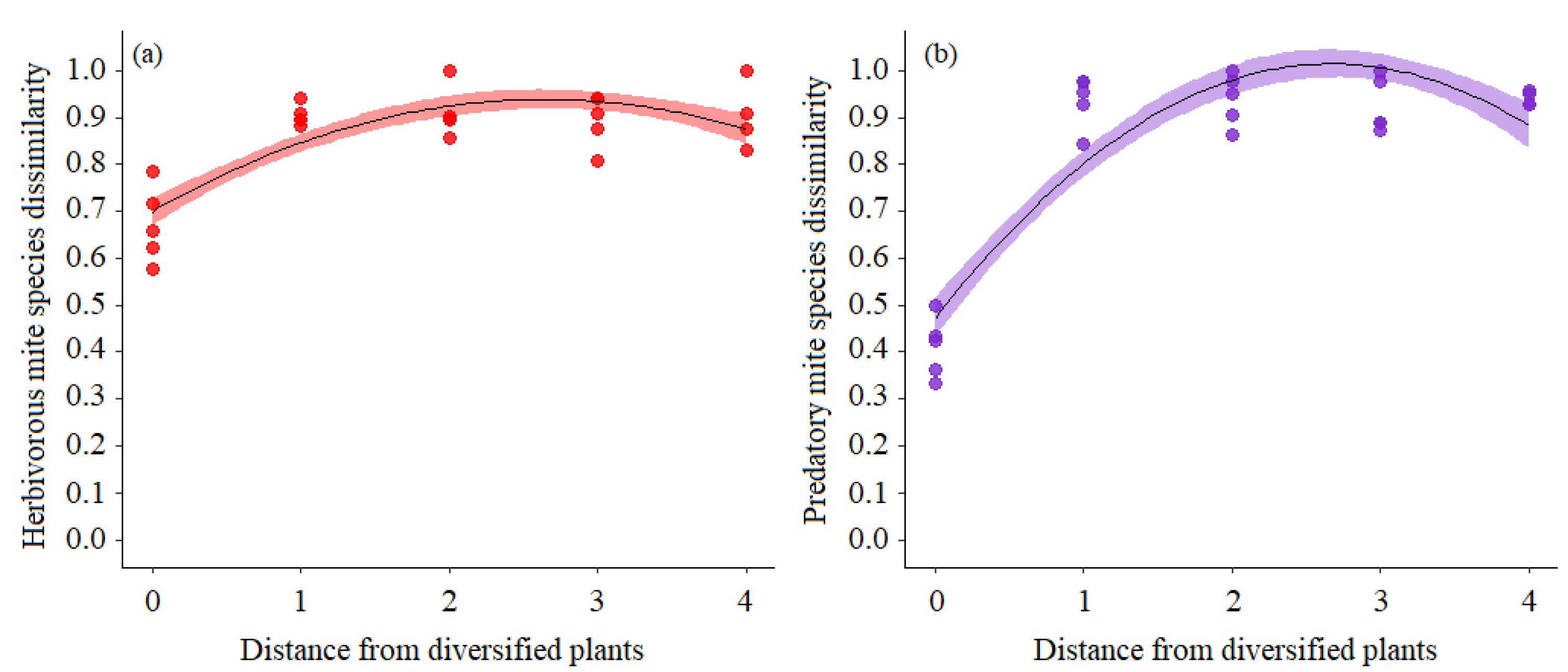
3.2. Dissimilarity of Mites
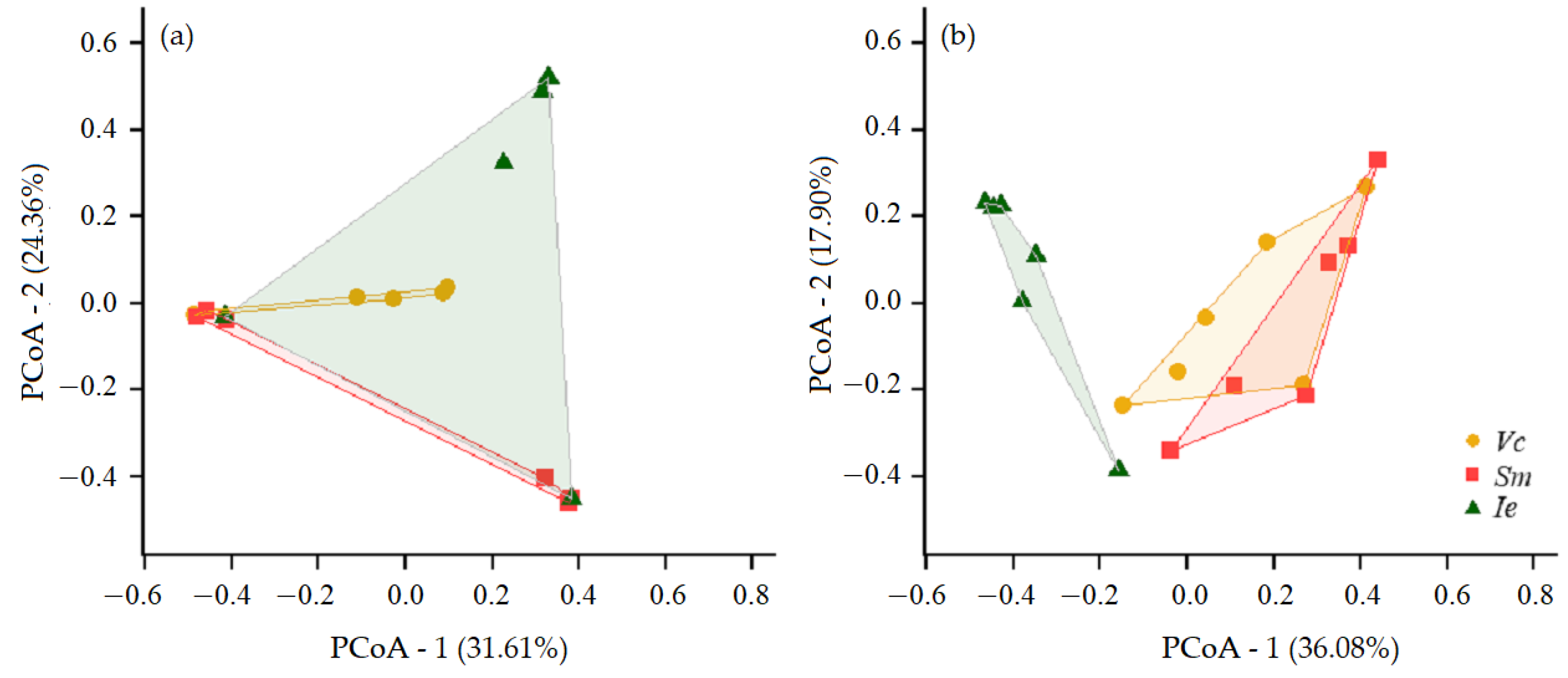
3.3. Communities of Herbivorous and Predatory Mites on the IPS
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CONAB. Acompanhamento Da Safra Brasileira. Safra 2021. 2021, Volume 8, p. 59. Available online: http://www.sapc.embrapa.br/arquivos/consorcio/levantamento/Conab_safra_2021_n2.pdf (accessed on 13 February 2021).
- Avelino, J.; Allinne, C.; Cerda, R.; Willocquet, L.; Savary, S. Multiple-Disease System in Coffee: From Crop Loss Assessment to Sustainable Management. Annu. Rev. Phytopathol. 2018, 56, 611–635. [Google Scholar] [CrossRef] [PubMed]
- de Aparecido, L.E.O.; de Rolim, G.S.; da Silva Cabral De Moraes, J.; Costa, C.T.S.; de Souza, P.S. Machine Learning Algorithms for Forecasting the Incidence of Coffea arabica Pests and Diseases. Int. J. Biometeorol. 2020, 64, 671–688. [Google Scholar] [CrossRef] [PubMed]
- Spongoski, S.; Reis, P.R.; Zacarias, M.S. Acarofauna Da Cafeicultura de Cerrado em Patrocínio, Minas Gerais. Cienc. Agrotec. 2005, 29, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Tasei, J. Impact of Agrochemicals on Non-Apis Bees. In Honey Bees: Estimating the Environmental Impact of Chemicals; Taylor & Francis: London, UK, 2002; pp. 101–131. Available online: https://books.google.com.au/books?hl=zh-TW&lr=&id=yov_MkEt6EwC&oi=fnd&pg=PA101&ots=uJUEKmhDvT&sig=mBRHh51nr4hZkRxdi0GNBIQ9Qak#v=onepage&q&f=false (accessed on 15 March 2021).
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The Sublethal Effects of Pesticides on Beneficial Arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Giglio, A.; Cavaliere, F.; Giulianini, P.G.; Mazzei, A.; Talarico, F.; Vommaro, M.L.; Brandmayr, P. Impact of Agrochemicals on Non-Target Species: Calathus fuscipes Goeze 1777 (Coleoptera: Carabidae) as Model. Ecotoxicol. Environ. Saf. 2017, 142, 522–529. [Google Scholar] [CrossRef]
- Perfecto, I.; Rice, R.A.; Greenberg, R.; van der Voort, M.E. Shade Coffee: A Disappearing Refuge for Biodiversity. Bioscience 1996, 46, 598–608. [Google Scholar] [CrossRef] [Green Version]
- Perfecto, I.; Vandermeer, J. Biodiversity Conservation in Tropical Agroecosystems: A New Conservation Paradigm. Ann. N. Y. Acad. Sci. 2008, 1134, 173–200. [Google Scholar] [CrossRef]
- Khan, S.; Khan, M.; Hanjra, M.; Mu, J. Pathways to Reduce the Environmental Footprints of Water and Energy Inputs in Food Production. Food Policy 2009, 34, 141–149. [Google Scholar] [CrossRef]
- Pedlowski, M.A.; Canela, M.C.; da Costa Terra, M.A.; Ramos de Faria, R.M. Modes of Pesticides Utilization by Brazilian Smallholders and Their Implications for Human Health and the Environment. Crop Prot. 2012, 31, 113–118. [Google Scholar] [CrossRef]
- Sales, E.F.; Méndez, V.E.; Caporal, F.R.; Faria, J.C. Agroecological Transition of Conilon Coffee (Coffea canephora) Agroforestry Systems in the State of Espírito Santo, Brazil. Agroecol. Sustain. Food Syst. 2013, 37, 405–429. [Google Scholar] [CrossRef]
- Tixier, M.S. Predatory Mites (Acari: Phytoseiidae) in Agro-Ecosystems and Conservation Biological Control: A Review and Explorative Approach for Forecasting Plant-Predatory Mite Interactions and Mite Dispersal. Front. Ecol. Evol. 2018, 6, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Ratnadass, A.; Fernandes, P.; Avelino, J.; Habib, R. Plant Species Diversity for Sustainable Management of Crop Pests and Diseases in Agroecosystems: A Review. Agron. Sustain. Dev. 2012, 32, 273–303. [Google Scholar] [CrossRef] [Green Version]
- Letourneau, D.K.; Armbrecht, I.; Rivera, B.S.; Lerma, J.M.; Jimé, E.; Carmona, N.; Daza, M.C.; Escobar, S.; Vi´, V.; Galindo, V.; et al. Does Plant Diversity Benefit Agroecosystems? A Synthetic Review. Ecol. Appl. 2011, 21, 9–21. [Google Scholar] [CrossRef]
- Venzon, M.; Amaral, D.S.S.; Togni, P.H.B.; Rezende, M.Q.; Perez, A.L. Manejo de Pragas Na Agricultura Orgânica. In Tecnologias para Produção Orgânica; EPAMIG Zona da Mata: Viçosa, Brazil, 2011; pp. 107–128. [Google Scholar]
- Zehnder, G.; Gurr, G.M.; Kühne, S.; Wade, M.R.; Wratten, S.D.; Wyss, E. Arthropod Pest Management in Organic Crops. Annu. Rev. Entomol. 2006, 52, 57–80. [Google Scholar] [CrossRef] [Green Version]
- Begg, G.S.; Cook, S.M.; Dye, R.; Ferrante, M.; Franck, P.; Lavigne, C.; Lövei, G.L.; Mansion-Vaquie, A.; Pell, J.K.; Petit, S.; et al. A Functional Overview of Conservation Biological Control. Crop Prot. 2017, 97, 145–158. [Google Scholar] [CrossRef]
- Venzon, M.; Sujii, E.R. Conservative Biological Control. Inf. Agropecu. 2009, 30, 7–16. [Google Scholar]
- Pemberton, R.; Vandenberg, N. Extrafloral Nectar Feeding by Ladybird Beetles (Coleoptera: Coccinellidae). Proc. Entomol. Soc. Washingt. 1993, 95, 139–151. [Google Scholar]
- van Rijn, P.C.J.; Tanigoshi, L.K. The Contribution of Extrafloral Nectar to Survival and Reproduction of the Predatory Mite Iphiseius degenerans on Ricinus communis. In Ecology and Evolution of the Acari; Springer: Dordrecht, The Netherlands, 1999; pp. 405–417. [Google Scholar]
- Rezende, M.Q.; Venzon, M.; Perez, A.L.; Cardoso, I.M.; Janssen, A. Extrafloral Nectaries of Associated Trees Can Enhance Natural Pest Control. Agric. Ecosyst. Environ. 2014, 188, 198–203. [Google Scholar] [CrossRef]
- Rezende, M.Q.; Venzon, M.; dos Santos, P.S.; Cardoso, I.M.; Janssen, A. Extrafloral Nectary-Bearing Leguminous Trees Enhance Pest Control and Increase Fruit Weight in Associated Coffee Plants. Agric. Ecosyst. Environ. 2021, 319, 107538. [Google Scholar] [CrossRef]
- Botti, J.M.C. Role of Green Lacewings and Ants on Coffee Berry Borer Predation. Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, Brazil, 2021. [Google Scholar]
- da Rosado, M.C.; de Araújo, G.J.; Pallini, A.; Venzon, M. Cover Crop Intercropping Increases Biological Control in Coffee Crops. Biol. Control 2021, 160, 104675. [Google Scholar] [CrossRef]
- Martins, E.F.; Franzin, M.L.; Perez, A.L.; Schmidt, J.M.; Venzon, M. Is Ceraeochrysa cubana a Coffee Leaf Miner Predator? Biol. Control 2021, 160, 104691. [Google Scholar] [CrossRef]
- Wäckers, F.; van Rijn, P.; Bruin, J. Plant-Provided Food for Carnivorous Insects: A Protective Mutualism and Its Applications; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Cardoso, I.M.; Guijt, I.; Franco, F.S.; Carvalho, A.F.; Ferreira Neto, P.S. Continual Learning for Agroforestry System Design: University, NGO and Farmer Partnership in Minas Gerais, Brazil. Agric. Syst. 2001, 69, 235–257. [Google Scholar] [CrossRef]
- Grossman, J.M. Inga Oerstediana in Organic Coffee Agroecosystems: Nitrogen Fixation, Nodulation and Farmer Knowledge of Soil Processes in Chiapas, Mexico—ProQuest; University of Minnesota: Minneapolis, MN, USA, 2003. [Google Scholar]
- Souza, H.N.; Cardoso, I.M.; Fernandes, J.M.; Garcia, F.C.P.; Bonfim, V.R.; Santos, A.C.; Carvalho, A.F.; Mendonça, E.S. Selection of Native Trees for Intercropping with Coffee in the Atlantic Rainforest Biome. Agrofor. Syst. 2010, 80, 1–16. [Google Scholar] [CrossRef]
- Koptur, S. Floral and Extrafloral Nectars of Costa Rican Inga Trees: A Comparison of Their Constituents and Composition. Biotropica 1994, 26, 276–284. [Google Scholar] [CrossRef]
- Melo, Y.; Córdula, E.; Machado, S.; Alves, M. Morfologia de Nectários em Leguminosae Senso Lato em Áreas de Caatinga No Brasil. Acta Bot. Bras. 2010, 24, 1034–1045. [Google Scholar] [CrossRef]
- Brandão, D.S.; Mendes, A.D.R.; Santos, R.R.; Rocha, S.M.G.; Leite, G.L.D.; Martins, E.R. Biologia Floral e Sistema Reprodutivo Da Erva-Baleeira (Varronia curassavica Jacq.). Rev. Bras. Plantas Med. 2015, 17, 562–569. [Google Scholar] [CrossRef] [Green Version]
- Teodoro, A.V.; Klein, A.M.; Tscharntke, T. Temporally Mediated Responses of the Diversity of Coffee Mites to Agroforestry Management. J. Appl. Entomol. 2009, 133, 659–665. [Google Scholar] [CrossRef]
- Androcioli, H.G.; Hoshino, A.T.; de Menezes Júnior, A.O.; Morais, H.; Bianco, R.; Caramori, P.H. Coffee Leaf Miner Incidence and Its Predation by Wasp in Coffee Intercropped with Rubber Trees. Coffee Sci. 2018, 13, 389–400. [Google Scholar] [CrossRef] [Green Version]
- de Souza, G.S.; Alves, D.I.; Dan, M.L.; de Lima, J.S.S.; da Fonseca, A.L.C.C.; Araújo, J.B.S.; de Guimarães, L.A.O.P. Soil Physico-Hydraulic Properties under Organic Conilon Coffee Intercropped with Tree and Fruit Species. Pesqui. Agropecu. Bras. 2017, 52, 539–547. [Google Scholar] [CrossRef] [Green Version]
- Mcmurtry, J.A.; Croft, B.A. Life-Styles of Phytoseiid Mites and Their Roles in Biological Control. Annu. Rev. Entomol. 1997, 42, 291–321. [Google Scholar] [CrossRef]
- McMurtry, J.A.; De Moraes, G.J.; Sourassou, N.F. Revision of the Lifestyles of Phytoseiid Mites (Acari: Phytoseiidae) and Implications for Biological Control Strategies. Syst. Appl. Acarol. 2013, 18, 297–320. [Google Scholar] [CrossRef] [Green Version]
- Croft, B.; Blackwood, J.; McMurtry, J. Classifying Life-Style Types of Phytoseiid Mites:Diagnostic Traits. Exp. Appl. Acarol. 2004, 33, 247–260. [Google Scholar] [CrossRef]
- Pemberton, R. Observations of Extrafloral Nectar Feeding by Predaceous and Fungivorous Mites. Proc. Entomol. Soc. Washingt. 1993, 95, 642–643. [Google Scholar]
- White, N.D.; Laing, J.E. Some Aspects of the Biology and a Laboratory Life Table of the Acarine Predator Zetzellia mali. Can. Entomol. 1977, 109, 1275–1281. [Google Scholar] [CrossRef]
- Rodríguez-Cruz, F.A.; Venzon, M.; Pinto, C.M.F. Performance of Amblyseius herbicolus on Broad Mites and on Castor Bean and Sunnhemp Pollen. Exp. Appl. Acarol. 2013, 60, 497–507. [Google Scholar] [CrossRef]
- Villanueva, R.; Childers, C. Phytoseiidae Increase with Pollen Deposition on Citrus Leaves. Florida Entomol. 2014, 87, 609–611. [Google Scholar] [CrossRef]
- Peixoto, M.L.; Fernandes, L.G.; Carvalho, M.A.C.; de Oliveira, M.L.; Putti, F.F.; Reis, A.R. Assessment of Mite Fauna in Different Coffee Cropping Systems in Brazil. Biocontrol Sci. Technol. 2017, 27, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Pallini, A. Acarofauna e Predação de Ácaros Fitófagos Por Ácaros Predadores em Cafeeiro (Coffea arabica L.) No Sul de Minas Gerais. Ph.D. Thesis, niversidade Federal de Lavras, Lavras, MG, Brazil, 1991. [Google Scholar]
- Reis, P.; Chiavegato, L.; Alves, E.; Sousa, E. Ácaros Da Família Phytoseiidae Associados Aos Citros No Município de Lavras, Sul de Minas Gerais. An. Soc. Entomol. Bras. 2000, 29, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, M.M. Ácaros Do Cafeeiro em Minas Gerais Com Chave de Identificação. Master’s Thesis, Universidade Federal de Viçosa, Viçosa, MG, Brazil, 2008. [Google Scholar]
- De Carvalho Mineiro, J.L.; Sato, M.E.; Raga, A.; De Souza Filho, M.F.; Spongoski, S. Mite Fauna on Coffee Plants Cv. Catuaí Amarelo. Bragantia 2008, 67, 197–201. [Google Scholar] [CrossRef]
- Silva, E.A.; Reis, P.R.; Zacarias, M.S.; Marafeli, P.P. Phytoseiids (Acari: Phytoseiidae) Associated to Coffee Plantations and Adjacent Forest Fragments. Cienc. Agrotec. 2010, 34, 1146–1153. [Google Scholar] [CrossRef]
- Berton, L.H.C.; de Mineiro, J.L.C.; Sato, M.E.; de Joaquim, A.A.F.; Raga, A. Mite Fauna of a Coffee Agroecosystem (Coffea arabica L.) in the Municipality of Monte Alegre Do Sul, São Paulo State, Brazil. Part I. Acarologia 2019, 59, 542–550. [Google Scholar] [CrossRef]
- Mineiro, J.; Sato, M.; Matioli, A.; Raga, A. Ácaros Plantícolas e Edáficos em Agroecossistema Cafeeiro, em Atibaia, Estado de São Paulo. In Proceedings of the Simpósio de Pesquisa dos Cafés do Brasil, Brasilia, Brazil, January/June 2009; Available online: http://www.biologico.agricultura.sp.gov.br/uploads/docs/bio/v70_1/mineiro.pdf (accessed on 20 January 2022).
- Mineiro, J.; Sato, M.; Raga, A.; Arthur, V.; Cangani, K.; Barbosa, F. Diversidade de Ácaros (Arachnida: Acari) em Cinco Cultivares de Duas Espécies de Cafeeiros (Coffea Spp.) em Garça, Estado de São Paulo. Arq. Inst. Biol. 2006, 73, 333–341. [Google Scholar] [CrossRef]
- Reis, P.; Souza, J. Pragas Do Cafeeiro. In Cultura do Cafeeiro: Fatores que Afetam a Produtividade; Piracicaba: Associação Brasileira Para Pesquisa da Potássio e do Fosfato: Piracicaba, Brazil, 1986; pp. 323–378. [Google Scholar]
- Reis, P. Ácaro-Vermelho. In Cultivar Grandes Culturas; 2005; Volume 7, pp. 14–17. [Google Scholar]
- Reis, P.; Zacarias, M. Ácaros em Cafeeiro. EPAMIG. 2007, pp. 1–76. Available online: http://www.sapc.embrapa.br/arquivos/consorcio/boletins_tecnicos/Acaros_em_cafeeiro.pdf (accessed on 10 July 2021).
- Chagas, C. Associação Do Ácaro Brevipalpus phoenicis (Geijskes) à Mancha Anular Do Cafeeiro. Biológico 1973, 39, 229–232. [Google Scholar]
- Navia, D.; Mendonça, R.S.; Ferragut, F.; Miranda, L.C.; Trincado, R.C.; Michaux, J.; Navajas, M. Cryptic Diversity in Brevipalpus Mites (Tenuipalpidae). Zool. Scr. 2013, 42, 406–426. [Google Scholar] [CrossRef]
- Beard, J.; Ochoa, R.; Braswell, W.; Bauchan, G. Brevipalpus phoenicis (Geijskes) Species Complex (Acari: Tenuipalpidae)—A Closer Look. Zootaxa 2015, 3944, 1–67. [Google Scholar] [CrossRef] [Green Version]
- Nunes, M.A.; de Carvalho Mineiro, J.L.; Rogerio, L.A.; Ferreira, L.M.; Tassi, A.; Novelli, V.M.; Kitajima, E.W.; Freitas-Astúa, J. First Report of Brevipalpus papayensis as Vector of Coffee Ringspot Virus and Citrus Leprosis Virus C. Plant Dis. 2018, 102, 1046. [Google Scholar] [CrossRef]
- Mineiro, J.L.D.C.; Sato, M.E. Distribuição de Brevipalpus papayensis Baker e Brevipalpus yothersi Baker (Acari: Tenuipalpidae) em Cafeeiros No Estado de São Paulo. In X Simpósio de Pesquisa dos Cafés do Brasil; Centro de Convenções de Vitoria: Vitória, Brazil, 2019; Available online: http://www.consorciopesquisacafe.com.br/ojs/index.php/SimposioCafe2019/article/view/321 (accessed on 11 May 2021).
- Flechtmann, C.H. Os Ácaros Do Cafeeiro. An. Esc. Super. Agric. Luiz Queiroz 1967, 24, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Pena, J.E.; Bullock, R.C. Effects of Feeding of Broad Mite (Acari: Tarsonemidae) on Vegetative Plant Growth. Florida Entomol. 1994, 77, 180–184. [Google Scholar] [CrossRef]
- Amaral, J. A Infestação de Ácaros Nos Cafezais. Biológico 1951, 17, 130. [Google Scholar]
- Jeppson, L.; Keifer, H.; Baker, E. Mites Injurious to Economic Plants; University of California Press: Berkeley, CA, USA, 1975. [Google Scholar]
- Matiello, J. Novas Condições de Ocorrência de Mancha Anular Do Cafeeiro. In Proceedings of the 14 Congresso Brasileiro de Pesquisas Cafeeiras, Rio de Janeiro, Brazil; 1987. [Google Scholar]
- IBGE Estado de Minas Gerais Meso e Microrregiões Do IBGE. Available online: https://www.mg.gov.br/sites/default/files/paginas/arquivos/2016/ligminas_10_2_04_listamesomicro.pdf (accessed on 27 July 2021).
- Souza, J.C.; Reis, P.; de Rigitano, R.L.O. Bicho-Mineiro Do Cafe: Biologia, Dano e Manejo Integrado. Bol. Técnico EPAMIG 1998, 37, 1–29. Available online: http://www.sapc.embrapa.br/arquivos/consorcio/boletins_tecnicos/bt_bicho_mineiro.pdf (accessed on 2 July 2021).
- Krantz, G.W.; Walter, D.E. (Eds.) A Manual of Acarology, 3rd ed.; Texas Tech University Press: Lubbock, TX, USA, 2009. [Google Scholar]
- Andre, H. A Generic Revision of the Family Tydeidae (Acari: Actinedida). IV. Generic Descriptions, Keys and Conclusions. Ann. Soc. R. Zool. Belgie 1980, 116, 103–168. [Google Scholar]
- Baker, E.; Tuttle, D. A Guide to the Spider Mites (Tetranychidae) of the United States; Indira Publishing House: West Bloomfield, MI, USA, 1994; ISBN 9780930. [Google Scholar]
- Chant, D.; McMurtry, J. Illustrated Keys and Diagnoses for the Genera and Subgenera of the Phytoseiidae of the World (Acari: Mesostigmata); Indira Publishing House: West Bloomfield, MI, USA, 2007; ISBN 0930337220. [Google Scholar]
- Beard, J.J.; Ochoa, R.; Bauchan, G.R.; Trice, M.D.; Redford, A.J.; Walters, T.W.; Mitter, C. Lucid Key: Brevipalpus of the World. Available online: https://idtools.org/id/mites/flatmites/key.php?key=genera_key (accessed on 3 October 2022).
- Fan, Q.H.; Flechtmann, C.H.W.; De Moraes, G.J. Annotated Catalogue of Stigmaeidae (Acari: Prostigmata), with a Pictorial Key to Genera. Zootaxa 2016, 4176, 199. [Google Scholar] [CrossRef]
- Chant, D.; McMurtry, J. A Review of the Subfamilies Phytoseiinae and Typhlodrominae (Acari: Phytoseiidae). Int. J. Acarol. 1994, 20, 223–310. [Google Scholar] [CrossRef]
- Paktinat-Saeij, S.; Bagheri, M.; Noronha, A.C.D.S. A New Species of Agistemus Summers (Acari: Trombidiformes: Stigmaeidae) from Brazil, with a Key to the American Species. Syst. Appl. Acarol. 2016, 21, 813–819. [Google Scholar] [CrossRef]
- Beard, J.; Ochoa, R.; Bauchan, G.; Trice, M.; Redford, A.; Walters, T.; Mitter, C.; Lucid Key: Flat Mite Genera of the World. Lucid Key: Flat Mite Genera of the World. Available online: http://idtools.org/id/mites/flatmites/ (accessed on 18 November 2022).
- Amrine, J.; Manson, D. Preparation, Mounting and Descriptive Study of Eriophyoid Mites. In Eriophyoid Mites—Their Biology, Natural Enemies and Control; Lindquist, E., Sabelis, M., Bruin, J., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1996; pp. 383–396. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2014, 67, 1–48. [Google Scholar] [CrossRef]
- Mazerolle, M.; Linden, D. AICcmodavg: Model Selection and Multimodel Inference Based on (Q) AIC (c)(2.2-2). 2019, pp. 1–223. Available online: https://cran.r-project.org/web/packages/AICcmodavg/AICcmodavg.pdf (accessed on 11 March 2021).
- Burnham, K.; Anderson, D. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A General and Simple Method for Obtaining R2 from Generalized Linear Mixed-Effects Models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Sage Publications: Thousands Oaks, CA, USA, 2018. [Google Scholar]
- Lenth, R.V. Least-Squares Means: The R Package Lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; De Cáceres, M. Beta Diversity as the Variance of Community Data: Dissimilarity Coefficients and Partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.; Hara, R.; Simpson, G.; Solymos, P. Community Ecology Package. R Package Version 2.5–6. 2019. Available online: http://cran.r-project.org/ (accessed on 12 March 2021).
- Zhang, D. Rsq: R-Squared and Related Measures. R Package Version. 2018, Volume 1. Available online: https://cran.r-project.org/web/packages/rsq/rsq.pdf (accessed on 12 March 2021).
- Oksanen, J. Vegan: Community Ecology Package. R Package Version 1.8-5. 2007. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 12 March 2021).
- Arbizu, M. Pairwise Multilevel Comparison Using Adonis R Package Version 0.3. 2019. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing. 2013. Available online: https://www.gbif.org/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 12 March 2021).
- de Silva, V.C.; Perdoná, M.J.; Soratto, R.P.; Negrisoli, E. Ocorrência de Plantas Daninhas em Cultivo Consorciado de Café e Nogueira-Macadâmia. Pesqui. Agropecu. Trop. 2013, 43, 441–449. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Hessein, N.A.; Perring, T.M. Homeopronematus anconai (Baker) (Acari: Tydeidae) Predation on Citrus Flat Mite, Brevipalpus lewisi McGregor (Acari: Tenuipalpidae). Int. J. Acarol. 1988, 14, 89–90. [Google Scholar] [CrossRef]
- Thistlewood, H.M.A.; Clements, D.R.; Harmsen, R. Chapter 2.2 Stigmaeidae. World Crop Pests 1996, 6, 457–470. [Google Scholar] [CrossRef]
- Abou-Awad, B.A.; El-Sawaf, B.M.; Kader, A.A.A. Life History and Life Table of Pronematus ubiquitus (McGregor) as a Predator of Eriophyoid Mites in Egypt (Acari: Tydeidae). Acarologia 1999, 40, 29–32. [Google Scholar]
- Horn, T.B.; Johann, L.; Ferla, N.J. Ecological Interactions between Phytophagous and Predaceous Mites in Citrus Agroecosystems in Taquari Valley, Rio Grande Do Sul, Brazil. Syst. Appl. Acarol. 2011, 16, 133–144. [Google Scholar] [CrossRef]
- Van De Velde, V.; Duarte, M.V.A.; Benavente, A.; Vangansbeke, D.; Wäckers, F.; De Clercq, P. Quest for the Allmitey: Potential of Pronematus ubiquitus (Acari: Iolinidae) as a Biocontrol Agent against Tetranychus urticae and Tetranychus evansi (Acari: Tetranychidae) on Tomato (Solanum lycopersicum L.). bioRxiv 2021. [Google Scholar] [CrossRef]
- Gerson, U.; Smiley, R.; Ochoa, R. Mites (Acari) for Pest Control; Blackwell Science: Oxford, UK, 2008. [Google Scholar]
- Reis, P.R.; Chiavegato, L.G.; Alves, E.B. Biologia de Iphiseiodes zuluagai Denmark & Muma (Acari: Phytoseiidae). An. Soc. Entomol. Bras. 1998, 27, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Reis, P.R.; Sousa, E.O.; Teodoro, A.V.; Pedro Neto, M. Effect of Prey Density on the Functional and Numerical Responses of Two Species of Predaceous Mites (Acari: Phytoseiidae). Neotrop. Entomol. 2003, 32, 461–467. [Google Scholar] [CrossRef]
- Yamamoto, P.T.; Gravena, S. Influência Da Temperatura e Fontes de Alimento No Desenvolvimento e Oviposição de Iphiseiodes zuluagai Denmark & Muma (Acari: Phytoseiidae). An. Soc. Entomol. Bras. 1996, 25, 109–115. [Google Scholar] [CrossRef]
- De Albuquerque, F.A.; De Moraes, G.J. Perspectivas Para a Criação Massal de Iphiseiodes zuluagai Denmark & Muma (Acari: Phytoseiidae). Neotrop. Entomol. 2008, 37, 328–333. [Google Scholar] [CrossRef] [Green Version]
- Andre, H.; Fan, A. Phylogeny, Ontogeny and Adaptive Radiation in the Superfamily Tydeoidea (Acari: Actinedida), with a Reappraisal of Morphological Characters. Zool. J. Linn. Soc. 2000, 130, 405–448. [Google Scholar] [CrossRef]
- White, N. Some Aspects of the Biology of the Predaceous Mite Zetzellia Mali (Ewing) (Acarine: Stigmaeidae) Found in Southern Ontario Apple Orchards. Master’s Thesis, University of Guelph, Guelph, ON, Canada, 1976. [Google Scholar]
- O’Dowd, D. Mite Association with the Leaf Domatia of Coffee (Coffea arabica) in North Queensland, Australia. Bull. Entomol. Res. 1994, 84, 361–366. [Google Scholar] [CrossRef]
- Franco, R.A.; Reis, P.R.; Zacarias, M.S.; Altoé, B.F.; Neto, M.P. Population Dynamics of Oligonychus ilicis (McGregor, 1917) (Acari: Tetranychidae) in Coffee Plants and of Their Associated Phytoseiids. Coffee Sci. 2008, 3, 38–46. [Google Scholar]
- Landis, D.; Menalled, F.; Costamagna, A.C.; Wilkinson, T.K. Manipulating Plant Resources to Enhance Beneficial Arthropods in Agricultural Landscapes. Weed Sci. 2005, 53, 902–908. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feeding Behavior | Mite Family | Mite Species | Plant Species | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Vc | Sm | Ie | Ca1 | Ca2 | Ca3 | Ca4 | |||
| Generalists | Tydeidae | Lorryia formosa Cooreman | 1 | 1 | 48 | 1 | 1 | ||
| Tydeidae | Lorryia sp1 | 3 | 34 | 8 | 14 | 7 | 22 | 10 | |
| Tydeidae | Lorryia sp2 | 5 | 1 | 1 | |||||
| Mycophagous | Acaridae | Tyrophagus putrescentiae (Schrank) | 26 | 27 | 84 | 7 | 10 | 12 | 16 |
| Astigmatina * | Astigmatina sp. | 3 | |||||||
| Glycyphagidae | Lepidoglyphus destructor (Schrank) | 1 | 1 | ||||||
| Tarsonemidae | Daidalotarsonemus savanicus Rezende, Lofego and Ochoa | 4 | 1 | ||||||
| Tarsonemidae | Tarsonemus confusus Ewing | 7 | 6 | 11 | 1 | 2 | |||
| Winterschmidtiidae | Winterschmidtiidae sp. | 4 | 1 | ||||||
| Herbivores | Eriophyidae | Aculus sp. | 1200 | ||||||
| Eriophyidae | Eriophyidae sp1 | 4885 | |||||||
| Eriophyidae | Eriophyidae sp2 | 70 | |||||||
| Eriophyidae | Eriophyidae sp3 | 97 | |||||||
| Diptilomiopidae | Diptilomiopidae sp. | 18 | |||||||
| Tarsonemidae | Polyphagotarsonemus latus (Banks) | 65 | |||||||
| Tenuipalpidae | Brevipalpus yothersi Baker | 15 | 144 | 77 | 95 | 152 | 110 | 100 | |
| Tetranychidae | Atrichoproctus uncinatus Flechtmann | 12 | 7 | 12 | |||||
| Tetranychidae | Mononychellus planki (McGregor) | 3 | |||||||
| Tetranychidae | Oligonychus coffeae (Nietner) | 7 | 4 | 12 | 4 | ||||
| Tetranychidae | Oligonychus ilicis (McGregor) | 1 | 1 | 1 | 4 | 2 | |||
| Tetranychidae | Tetranychus sp. | 2 | |||||||
| Predators | Iolinidae | Pseudopronematulus nadirae Silva, Da-Costa and Ferla | 41 | 61 | 1 | 5 | 7 | 10 | 4 |
| Iolinidae | Pausia sp. | 5 | 7 | ||||||
| Phytoseiidae | Amblyseius aff. chiapensis | 1 | 2 | ||||||
| Phytoseiidae | Amblyseius aff. impressus | 1 | |||||||
| Phytoseiidae | Amblyseius sp. | 1 | |||||||
| Phytoseiidae | Euseius alatus DeLeon | 4 | |||||||
| Phytoseiidae | Euseius citrifolius (Denmark and Muma) | 5 | 6 | 7 | 4 | 2 | 2 | 1 | |
| Phytoseiidae | Euseius concordis (Chant) | 1 | 1 | 11 | 4 | 5 | 3 | 3 | |
| Phytoseiidae | Euseius sibelius (De Leon) | 1 | 6 | 1 | |||||
| Phytoseiidae | Iphiseiodes zuluagai Denmark and Muma | 32 | 2 | 2 | |||||
| Phytoseiidae | Galendromus annectens (De Leon) | 4 | 2 | ||||||
| Phytoseiidae | Neoseiulus tunus (De Leon) | 6 | |||||||
| Phytoseiidae | Typhlodromalus aripo DeLeon | 4 | 4 | 1 | |||||
| Phytoseiidae | Typhlodromips mangleae De Leon | 1 | 4 | ||||||
| Stigmaeidae | Agistemus floridanus Gonzalez | 38 | 13 | 78 | 1 | 1 | 1 | ||
| Stigmaeidae | Agistemus brasiliensis Matioli, Ueckermann and Oliveira | 2 | |||||||
| Feeding Behavior | Mite Family | Plant Species | ||||||
|---|---|---|---|---|---|---|---|---|
| Vc | Sm | Ie | Ca1 | Ca2 | Ca3 | Ca4 | ||
| Generalists | Tydeidae | 6 | 37 | 76 | 35 | 19 | 50 | 22 |
| Mycophagous | Acaridae | 32 | 37 | 109 | 19 | 17 | 19 | 25 |
| Astigmatina * | 3 | |||||||
| Glycyphagidae | 1 | 1 | ||||||
| Tarsonemidae | 111 | 14 | 23 | 7 | 12 | 3 | 4 | |
| Winterschmidtiidae | 4 | 1 | ||||||
| Herbivores | Eriophyidae | 167 | 1200 | 4885 | ||||
| Diptilomiopidae | 18 | |||||||
| Tarsonemidae | 108 | |||||||
| Tenuipalpidae | 15 | 158 | 84 | 130 | 208 | 171 | 147 | |
| Tetranychidae | 19 | 14 | 18 | 16 | 14 | 27 | 39 | |
| Predators | Iolinidae | 85 | 134 | 2 | 8 | 12 | 20 | 11 |
| Phytoseiidae | 37 | 53 | 192 | 16 | 15 | 15 | 21 | |
| Stigmaeidae | 48 | 20 | 129 | 1 | 1 | 1 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferla, J.J.; Araújo, G.J.d.; Venzon, M.; Nascimento, P.H.M.G.; Kalile, M.O.; Pancieri, S.D.; Cardoso, A.C.; Martins, E.F.; Ferla, N.J.; Pallini, A. Intercropped Plants Provide a Reservoir of Predatory Mites in Coffee Crop. Agriculture 2023, 13, 285. https://doi.org/10.3390/agriculture13020285
Ferla JJ, Araújo GJd, Venzon M, Nascimento PHMG, Kalile MO, Pancieri SD, Cardoso AC, Martins EF, Ferla NJ, Pallini A. Intercropped Plants Provide a Reservoir of Predatory Mites in Coffee Crop. Agriculture. 2023; 13(2):285. https://doi.org/10.3390/agriculture13020285
Chicago/Turabian StyleFerla, Júlia J., Gustavo J. de Araújo, Madelaine Venzon, Pedro H. M. G. Nascimento, Milena O. Kalile, Shauanne D. Pancieri, André C. Cardoso, Elem F. Martins, Noeli J. Ferla, and Angelo Pallini. 2023. "Intercropped Plants Provide a Reservoir of Predatory Mites in Coffee Crop" Agriculture 13, no. 2: 285. https://doi.org/10.3390/agriculture13020285