
Strigolactone (GR24) Application Positively Regulates Photosynthetic Attributes, Stress-Related Metabolites and Antioxidant Enzymatic Activities of Ornamental Sunflower (Helianthus annuus cv. Vincent’s Choice) under Salinity Stress

 , , ,
, , ,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Area, Seed Material and Layout
2.2. Chemical Materials
2.3. Application of Salinity and Strigolactone (GR24)
2.4. Measurement of Photosynthetic Attributes and Water Relations
2.5. Determination of Stress-Related Metabolites (Proline and Malondialdehyde)
2.6. Determination of Glycinebetaine (GB), Total Soluble Protein and Hydrogen Peroxide (H2O2)
2.7. Determination of Antioxidant Enzymatic Activities
2.8. Mineral Ion (Na+, K+ and Ca2+) Quantification
2.9. Statistical Analysis
3. Results
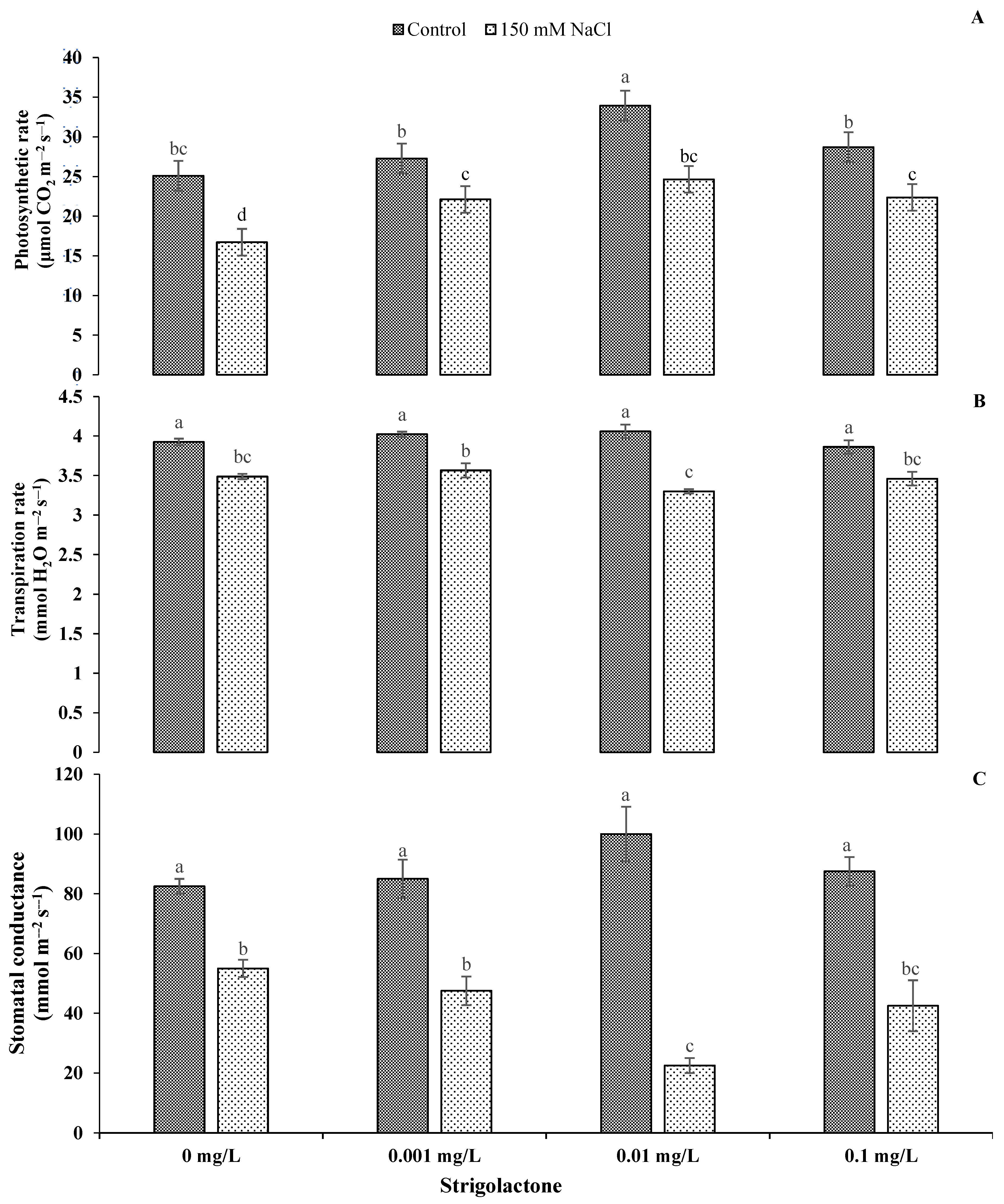
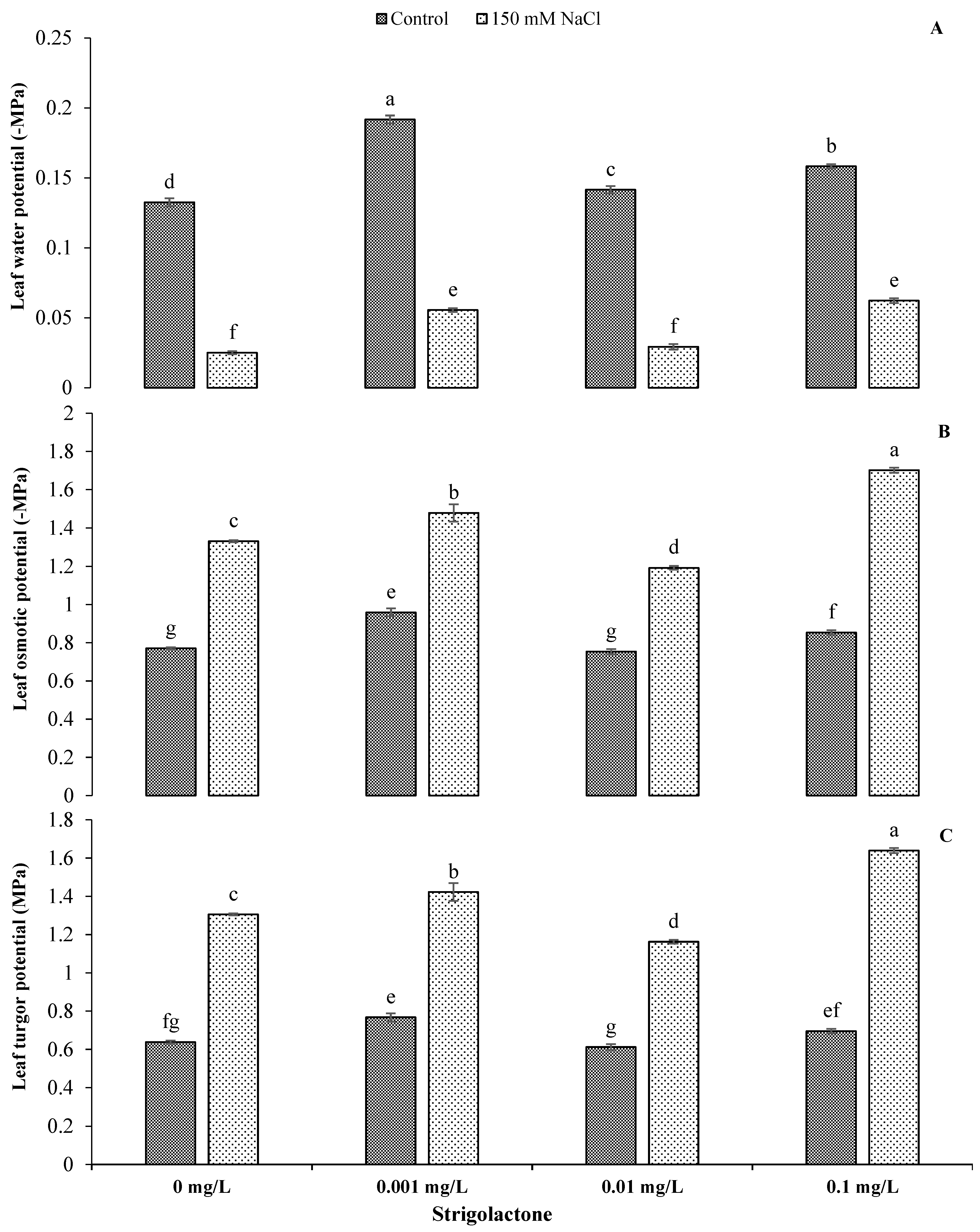
3.1. Role of Exogenous GR24 Application in Photosynthetic Attributes and Water Relations of Sunflower under Salt Stress
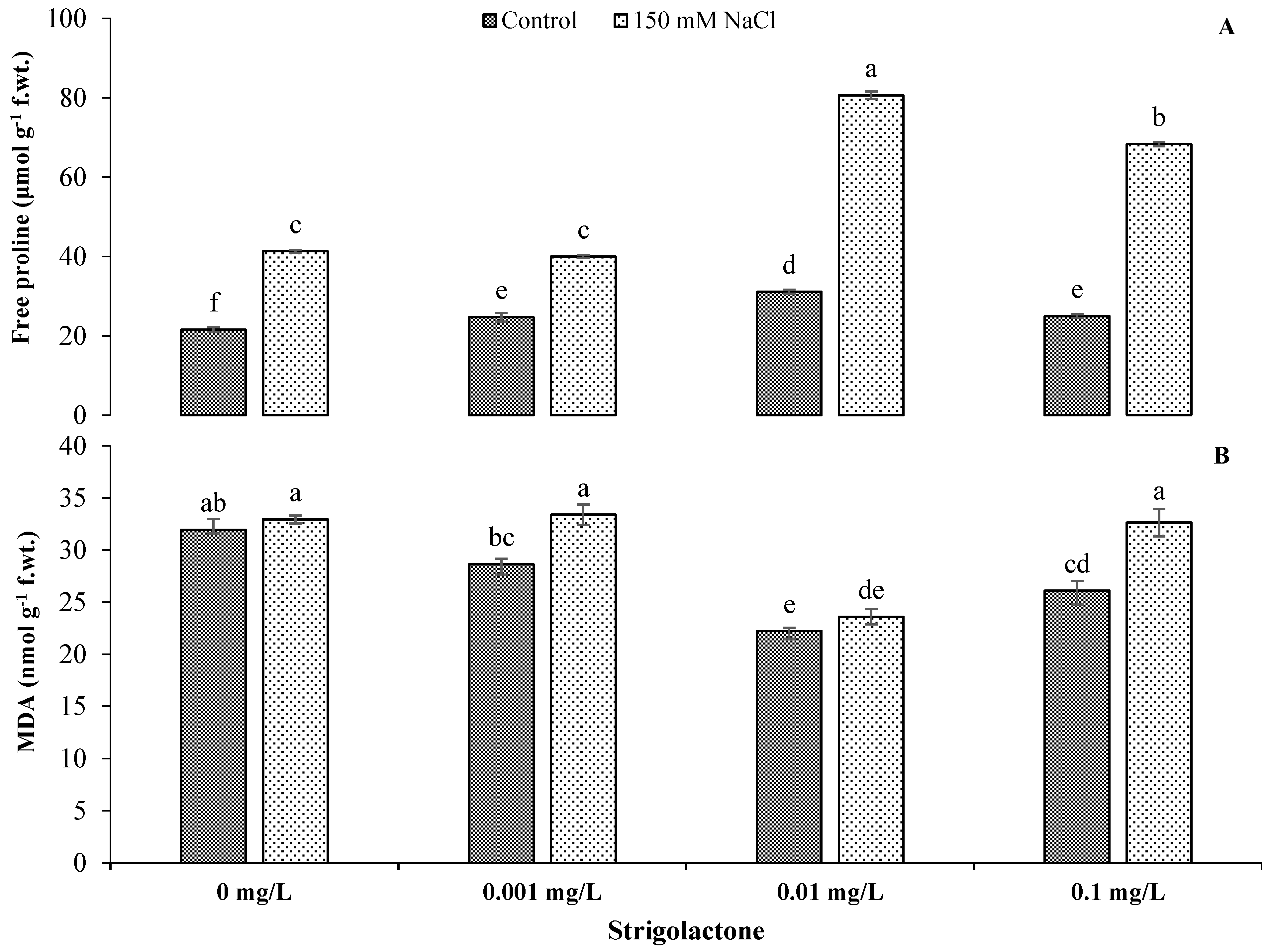
3.2. Effect of Exogenous GR24 Application on Stress-Related Metabolites (Free Proline and MDA) of Sunflower under Salt Stress
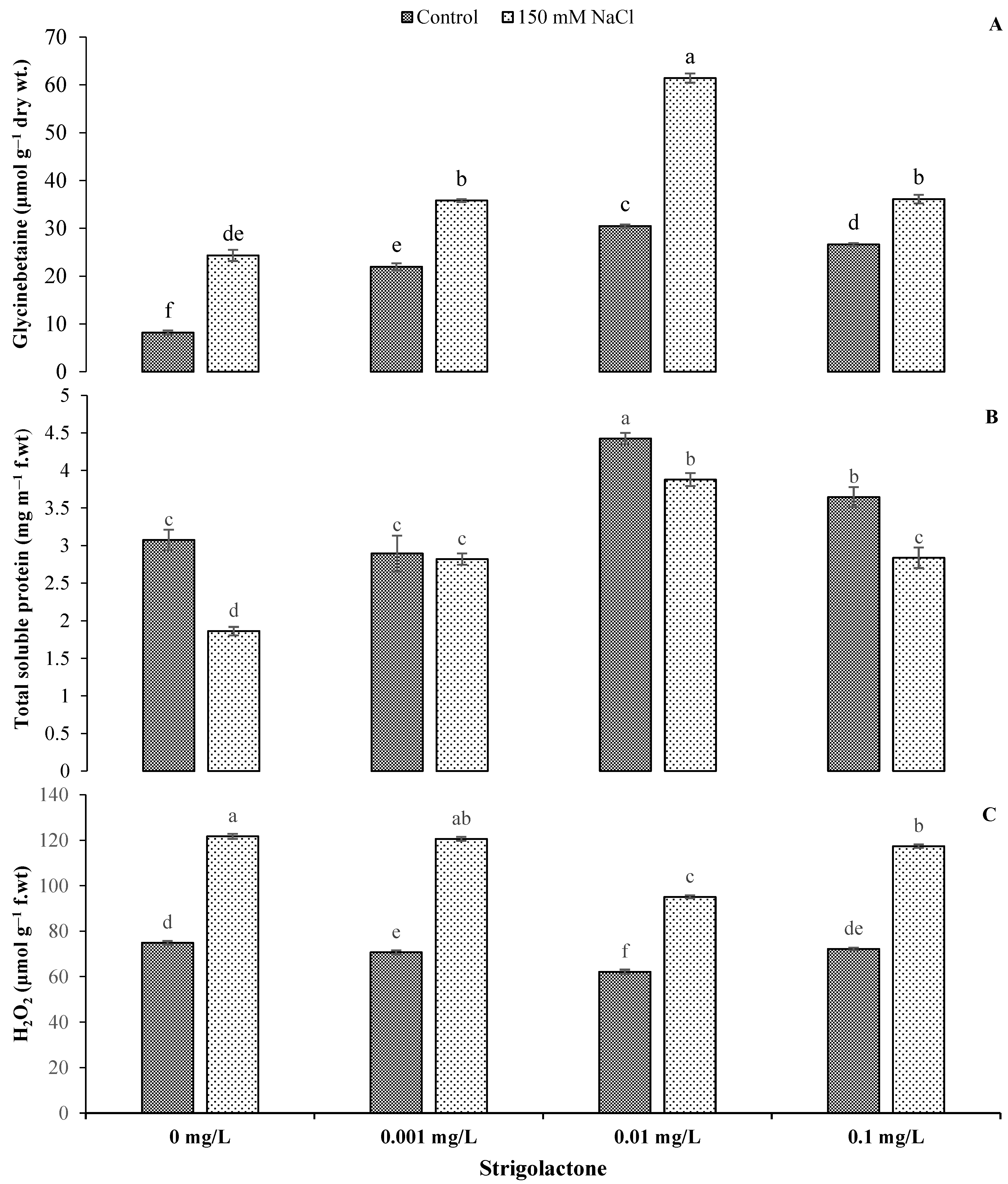
3.3. Impact of GR24 Application on GB, Total Soluble Protein and H2O2 under Salt Stress
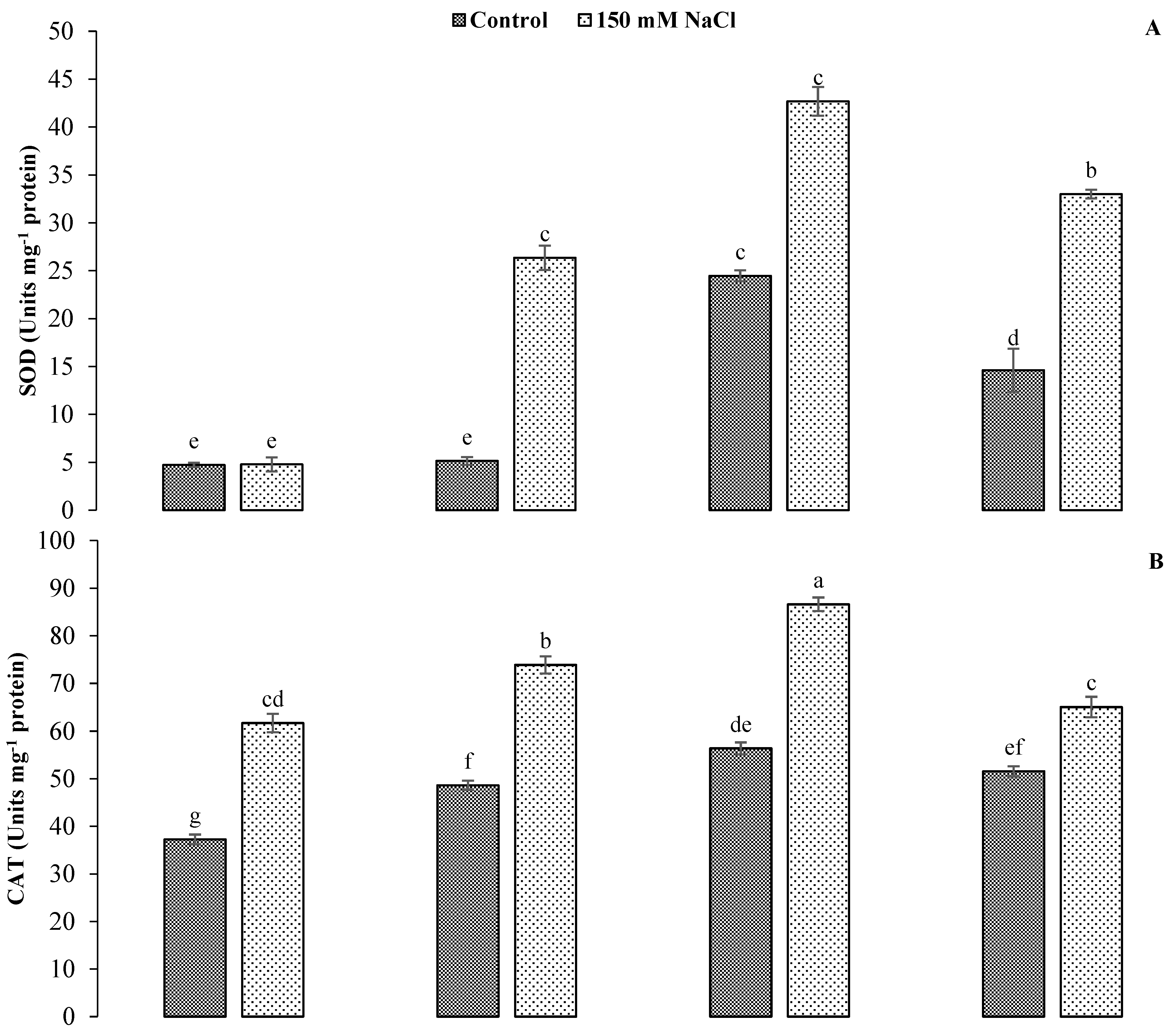
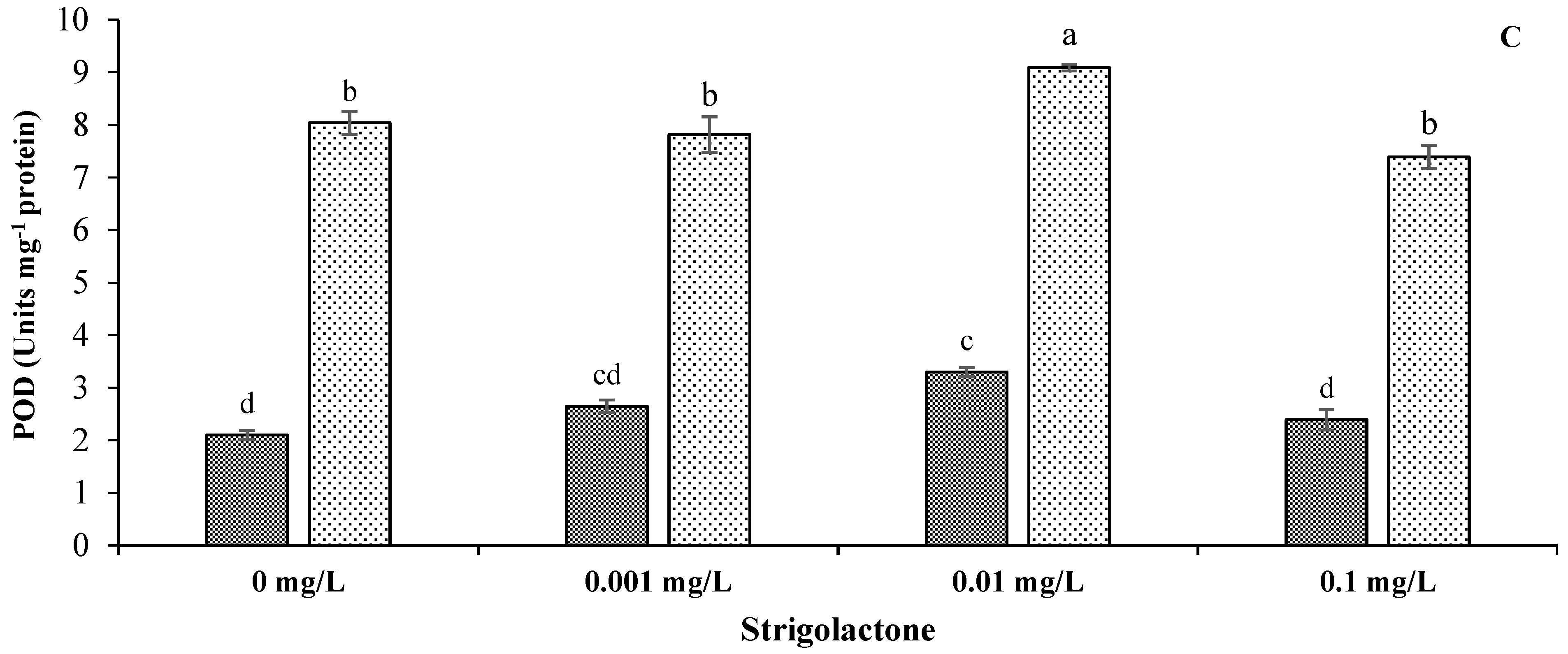
3.4. Impact of GR24 on Antioxidant Enzymatic Activities of Sunflower under Salinity Stress
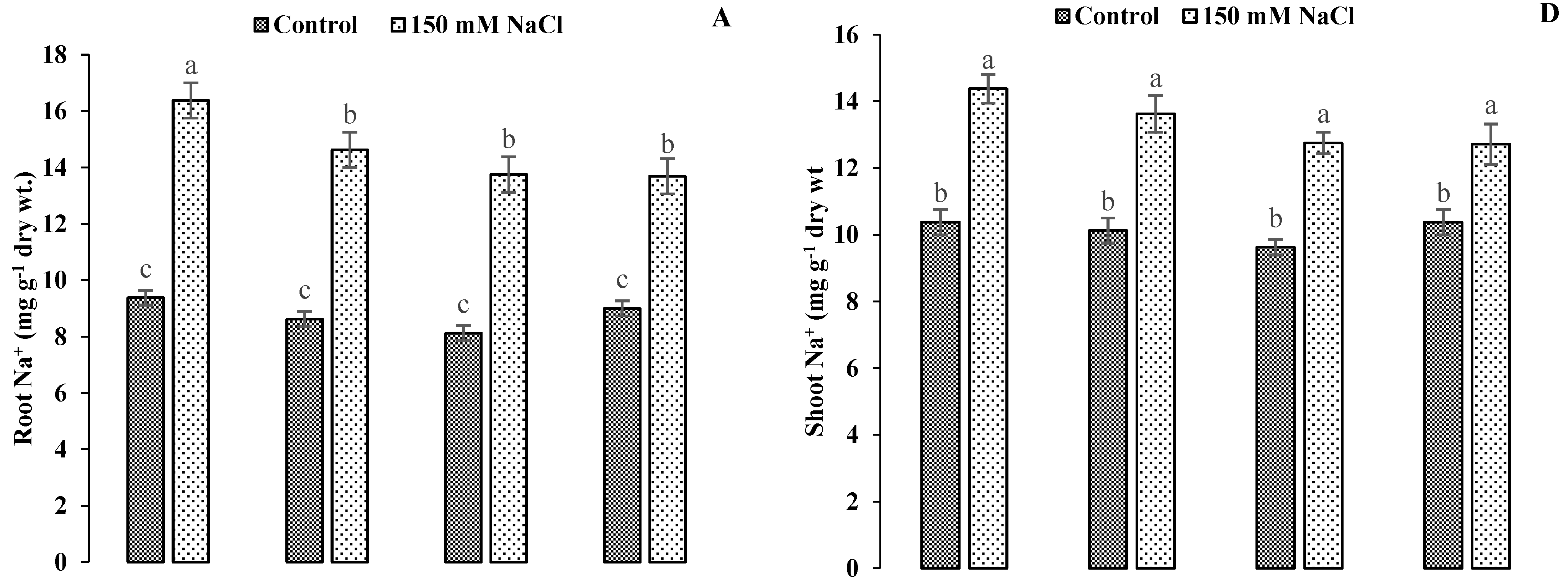
3.5. Impact of GR24 Application on Quantification of Mineral Ions in Roots and Shoots of Ornamental Sunflower under Salt Stress
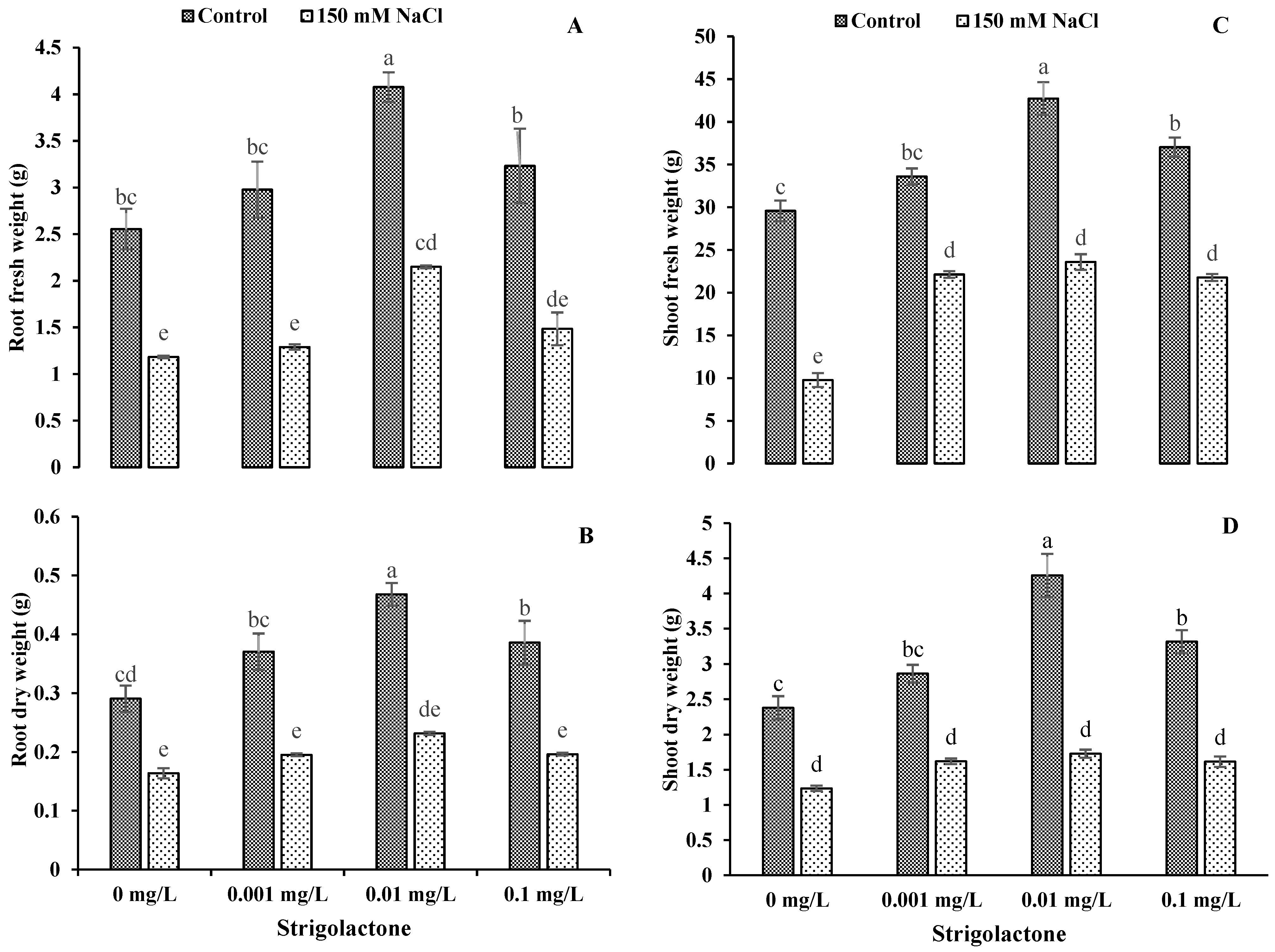
3.6. Impact of GR24 Application on Plant Biomass of Ornamental Sunflower under Salt Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J.T. An overview of hazardous impacts of soil salinity in crops, tolerance mechanisms, and amelioration through selenium supplementation. Int. J. Mol. Sci. 2019, 21, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, F.; Su, Q.; Jiang, H.; Cui, J.; He, X.; Wu, Z.; Zhang, Z.; Liu, J.; Zhao, Y. Effects of strigolactone on photosynthetic and physiological characteristics in salt-stressed rice seedlings. Sci. Rep. 2020, 10, 6183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalarukh, I.; Shahbaz, M. Response of antioxidants and lipid peroxidation to exogenous application of alpha-tocopherol in sunflower (Helianthus annuus L.) under salt stress. Pak. J. Bot. 2020, 52, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Schillaci, M.; Walker, R.; Smith, P.M.C.; Watt, M.; Roessner, U. Alleviation of salinity stress in plants by endophytic plant-fungal symbiosis: Current knowledge, perspectives and future directions. Plant Soil. 2021, 461, 219–244. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, C.; Xue, Y.; Liu, X.; Chen, S.; Song, C.P.; Yang, Y.; Guo, Y. Calcium-activated 14-3-3 proteins as a molecular switch in salt stress tolerance. Nat. Commun. 2019, 10, 1199. [Google Scholar] [CrossRef] [Green Version]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New insights on plant salt tolerance Mechanisms and their potential use for breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef] [Green Version]
- Hong, C.Y.; Chao, Y.Y.; Yang, M.Y.; Cheng, S.Y.; Cho, S.C.; Kao, C.H. NaCl-induced expression of glutathione reductase in roots of rice (Oryza sativa L.) seedlings is mediated through hydrogen peroxide but not abscisic acid. Plant Soil. 2009, 320, 103–115. [Google Scholar] [CrossRef]
- Grieve, C.M.; Poss, J.A. Response of ornamental sunflower cultivars ‘sunbeam’ and ‘moonbright’ to irrigation with saline wastewaters. J. Plant Nut. 2010, 11, 1579–1592. [Google Scholar] [CrossRef]
- Bosnjak, D.; Rodić, V.; Karapandžin, J. Possibilities for the improvement of the soybean production in Serbia. Contemp. Agric. 2013, 62, 266–275. [Google Scholar]
- Brito, C.L.L.; Matsumoto, S.N.; Santos, J.L.; Gonçalves, D.N.; Ribeiro, A.F.F. Effect of paclobutrazol on the development of ornamental sunflower plants. J. Agric. Sci. 2016, 39, 153–160. [Google Scholar] [CrossRef]
- Mladenovic, E.; Cvejic, S.; Čukanovic, J.; Žeravica, G.; Jocic, S. Evaluation of sunflower genotypes for ornamental use. Contemp. Agric. 2016, 65, 39–43. [Google Scholar] [CrossRef] [Green Version]
- Menzel, C.; González-Martínez, C.; Chiralt, A.; Vilaplana, F. Antioxidant starch films containing sunflower hull extracts. Carbohydr. Polym. 2019, 214, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Brewer, P.B.; Koltai, H.; Beveridge, C.A. Diverse roles of strigolactones in plant development. Mol. Plant 2013, 6, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwack, P.J.; Rashotte, A.M. Interactions between cytokinin signaling and abiotic stress response. J. Exp. Bot. 2015, 66, 4863–4871. [Google Scholar] [CrossRef] [Green Version]
- Cook, C.E.; Whichard, L.P.; Monroe, W.E.; Egley, G.H.; Coggon, P.; Luhan, P.A.; McPhail, A.T. Germination stimulants. II. Structure of strigol, a potent seed germination stimulant for witchweed (Striga lutea). J. Am. Chem. Soc. 1972, 94, 6198–6199. [Google Scholar] [CrossRef]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Magome, H.; Kamiya, Y.; Shirasu, K.; Kyozuka, J.; Yamaguchi, S. Inhibition of shoot branching by new terpenoid plant hormones. Nature 2008, 455, 195–200. [Google Scholar] [CrossRef]
- Zheng, X.; Li, Y.; Xi, X.; Ma, C.; Sun, Z.; Yang, X.; Li, X.; Tian, Y.; Wang, C. Exogenous Strigolactones alleviate KCl stress by regulating photosynthesis, ROS migration and ion transport in Malus hupehensis Rehd. Plant Physiol. Biochem. 2021, 159, 113–122. [Google Scholar] [CrossRef]
- Rani, K.; Zwanenburg, B.; Sugimoto, Y.; Yoneyama, K.; Bouwmeester, H.J. Biosynthetic considerations could assist the structure elucidation of host plant produced rhizosphere signaling compounds (strigolactones) for arbuscular mycorrhizal fungi and parasitic plants. Plant Physiol. Bioch. 2008, 46, 617–626. [Google Scholar] [CrossRef]
- Kohlen, W.; Tatsiana, C.; Liu, Q.; Bours, R.; Domagalska, M.A.; Beguerie, S.; Verstappen, F.; Leyser, O.; Bouwmeester, H.; Ruyter-Spira, C. Strigolactones are transported through the xylem and play a key role in shoot architectural response to phosphate deficiency in nonarbuscular mycorrhizal host Arabidopsis. Plant Physiol. 2011, 155, 974–987. [Google Scholar] [CrossRef] [Green Version]
- Koltai, H.; Prandi, C. Strigolactones: Biosynthesis, synthesis and functions in plant growth and stress responses. In Phytohormones: A Window to Metabolism, Signaling and Biotechnological Applications; Springer: New York, NY, USA, 2014; pp. 265–288. [Google Scholar]
- Liu, H.; Li, C.; Yan, M.; Zhao, Z.; Huang, P.; Wei, L.; Wu, X.; Wang, C.; Liao, W. Strigolactone is involved in nitric oxide-enhanced the salt resistance in tomato seedlings. J. Plant Res. 2022, 135, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Hu, C.; Wan, L.; Hu, Q.; Xiong, J.; Zhang, C. Strigolactones improve plant growth, photosynthesis and alleviate oxidative stress under salinity in rapeseed (Brassica napus L.) by regulating gene expression. Front. Plant Sci. 2017, 8, 1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarwar, Y.; Shahbaz, M. Modulation in growth, photosynthetic pigments, gas exchange attributes and inorganic ions in sunflower (Helianthus annuus l.) by strigolactones (GR24) achene priming under saline conditions. Pak. J. Bot. 2020, 52, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Tsago, Y.; Andargie, M.; Takele, A. In vitro screening for drought tolerance in different sorghum (Sorghum bicolor (L.) Moench) varieties. J. Stress Physiol. Biochem. 2013, 9, 72–83. [Google Scholar]
- Al-Jibouri, A.M.J.; Altahan, S.F.; Al-Anii, T.A. Evaluation of three sunflower (Helianthus annuus L.) hybrids for salt tolerance in vitro. JLS 2011, 5, 1037–1041. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil; California Agricultural Experiment Station Circular; University of California: Los Angeles, CA, USA, 1950; Volume 347, pp. 1–32. [Google Scholar]
- Bates, L.E.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Annu. Rev. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective roles of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutase in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A. Assay of catalase and peroxidase. Methods Enzymol. 1955, 2, 764–817. [Google Scholar]
- Kausar, F.; Shahbaz, M. Influence of strigolactone (GR24) as a seed treatment on growth, gas exchange and chlorophyll fluorescence of wheat under saline conditions. Int. J. Agric. Biol. 2017, 19, 321–327. [Google Scholar] [CrossRef]
- Zhou, X.; Tan, Z.; Zhou, Y.; Guo, S.; Sang, T.; Wang, Y.; Shu, S. Physiological mechanism of strigolactone enhancing tolerance to low light stress in cucumber seedlings. BMC Plant Biol. 2022, 22, 30. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, G.H.; Akhtar, J.; Anwar-Ul-Haq, M.; Ali, S.; Chen, Z.; Malik, W. Exogenous potassium differentially mitigates salt stress in tolerant and sensitive maize hybrids. Pak. J. Bot. 2014, 46, 135–146. [Google Scholar]
- Kausar, F.; Shahbaz, M. Interactive effect of foliar application of nitric oxide (NO) and salinity on wheat (Triticum aestivum L.). Pak. J. Bot. 2013, 45, 67–73. [Google Scholar]
- Zhang, X.; Zhang, L.; Ma, C.; Su, M.; Wang, J.; Zheng, S.; Zhang, T. Exogenous strigolactones alleviate the photosynthetic inhibition and oxidative damage of cucumber seedlings under salt stress. Sci. Hortic. 2022, 297, 110962. [Google Scholar] [CrossRef]
- Perveen, S.; Iqbal, M.; Parveen, A.; Akram, M.S.; Shahbaz, M.; Akber, S.; Mehboob, A. Exogenous triacontanol mediated increase in phenolics, proline, activity of nitrate reductase and shoot k+ confers salt tolerance in maize (Zea mays L.). Braz. J. Bot. 2017, 40, 1–11. [Google Scholar] [CrossRef]
- Chen, F.; Jiang, L.; Zheng, J.; Wang, R. Identification of differentially expressed proteins and phosphorylated proteins in rice seedlings in response to strigolactone treatment. PLoS ONE 2014, 9, e93947. [Google Scholar] [CrossRef] [Green Version]
- Cha-um, S.; Ashraf, M.; Kirdmanee, C. Screening upland rice (Oryza sativa L. ssp. indica) genotypes for salt-tolerance using multivariate cluster analysis. Afr. J. Biotechnol. 2010, 9, 4731–4740. [Google Scholar]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and Antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.H.; Ali, S.; Rehman, M.; Rizwan, M.; Kamran, M.; Ibrahim, A.A.M.; Khan, Z.; Bamagoos, A.A.; Alharby, H.F.; Hakeem, K.R. Individual and combined application of EDTA and citric acid assisted phytoextraction of copper using jute (Corchorus capsularis L.) seedlings. Environ. Technol. Innov. 2020, 19, 100895. [Google Scholar] [CrossRef]
- Li, L.; Xia, W.; Li, H.; Zeng, H.; Wei, B.; Han, S.; Yin, C. Salinity inhibits rice seed germination by reducing α-amylase activity via decreased bioactive gibberellin content. Front. Plant Sci. 2018, 9, 275. [Google Scholar] [CrossRef] [PubMed]
- Khosla, A.; Nelson, D.C. Strigolactones, super hormones in the fight against Striga. Curr. Opin. Plant Biol. 2016, 33, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Li, Z.; Jiang, X.; Qin, Y. Effects of salt stress on photosynthetic fluorescence characteristics, antioxidant system, and osmoregulation of Coreopsis tinctoria Nutt. Hortscience 2021, 56, 1066–1072. [Google Scholar] [CrossRef]
- Naveed, S.; Mustafa, A.; Niamat, B.; Ahmad, Z.; Yaseen, M.; Kamran, M.; Rafique, M.; Ahmar, S.; Chen, J.T. Alleviation of salinity-induced oxidative stress, improvement in growth, physiology and mineral nutrition of canola (Brassica napus L.) through calcium-fortified composted animal manure. Sustainability 2020, 12, 846. [Google Scholar] [CrossRef] [Green Version]
- Parveen, A.; Hamzah, S.M.; Kamran, M.; Haider, M.Z.; Chen, J.T.; Hur, G.; Javed, M.T.; Azeem, M. Effect of citric acid on growth, ecophysiology, chloroplast ultrastructure, and phytoremediation potential of jute (Corchorus capsularis l.) seedlings exposed to copper stress. Biomolecules 2020, 10, 592. [Google Scholar] [CrossRef]
- Mohamed, A.N.; Ismail, M.R. Changes in organic and inorganic solutes of in vitro tomato cultivars under NaCl stress. Aust. J. Crop Sci. 2011, 5, 939–944. [Google Scholar]
- Hu, Y.; Xia, S.; Su, Y.; Wang, H.; Luo, W.; Xiao, L. Brassinolide increases potato root growth in vitro in a dose-dependent way and alleviates salinity stress. Biomed Res. Int. 2016, 2016, 8231873. [Google Scholar] [CrossRef]
- Zulfiqar, H.; Shahbaz, M.; Ahsan, M.; Nafees, M.; Nadeem, H.; Akram, M.; Maqsood, A.; Ahmar, S.; Kamran, M.; Alamri, S.; et al. Strigolactone (GR24) induced salinity tolerance in sunflower (Helianthus annuus L.) by ameliorating morpho-physiological and biochemical attributes under in-vitro conditions. J. Plant Growth Regul. 2021, 40, 2079–2091. [Google Scholar] [CrossRef]
- Saleem, M.H.; Ali, S.; Rehman, M.; Rana, M.S.; Rizwan, M.; Kamran, M.; Imran, K.; Riaz, M.; Soliman, M.H.; Elkelish, E.; et al. Influence of phosphorus on copper phytoextraction via modulating cellular organelles in two jute (Corchorus capsularis L.) varieties grown in a copper mining soil of Hubei Province, China. Chemosphere 2020, 248, 126032. [Google Scholar] [CrossRef]
- Shi, Q.; Song, X.; Liu, Z.; Wang, Y.; Fu, J.; Su, C.; Xia, X.; Song, E.; Song, Y. Quinones derived from polychlorinated biphenyls induce ROS-dependent autophagy by evoking an Autophagic flux and inhibition of mTOR/p70S6k. Chem. Res. Toxicol. 2016, 29, 1160–1171. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.H.; Kamran, M.; Zhou, Y.; Parveen, A.; Mustafa, A.; Anjum, R.M.A.; Wang, B.; Liu, L. Appraising growth, oxidative stress and copper phytoextraction potential of flax (Linum usitatissimum L.) grown in soil differentially spiked with copper. J. Environ. Manag. 2020, 257, 109994. [Google Scholar] [CrossRef] [PubMed]
- Torres-Vera, R.; Garcia, J.M.; Lopez-Raez, J.A. Do strigolactones contribute to plant defence. Mol. Plant Pathol. 2014, 15, 211–216. [Google Scholar] [CrossRef]
- Marzec, M. Strigolactones as part of the plant defence system. Trends Plant Sci. 2016, 21, 900–903. [Google Scholar] [CrossRef]
- Koren, D.; Resnick, N.; Gati, E.M.; Belausov, E.; Weininger, S.; Kapulnik, Y.; Koltai, H. Strigolactone signaling in the endodermis is sufficient to restore root responses and involves SHORT HYPOCOTYL 2 (SHY2) activity. New Phytol. 2013, 198, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Jiang, D.; Liu, F.; Dai, T.; Liu, W.; Jing, Q.; Cao, W. Exogenous nitric oxide improves seed germination in wheat against mitochondrial oxidative damage induced by high salinity. Environ. Exp. Bot. 2009, 67, 222–227. [Google Scholar] [CrossRef]
- Quintero, J.M.; Fournier, J.M.; Benlloch, M.; Rodríguez-Navarro, A. Na+ accumulation in root symplast of sunflower plants exposed to moderate salinity is transpiration-dependent. J. Plant Physiol. 2008, 165, 1248–1254. [Google Scholar] [CrossRef]
- Eisa, S.; Hussain, S.; Geissler, N.; Koyro, H.W. Effect of NaCl salinity on water relations, photosynthesis and chemical composition of quinoa (Chenopodium quinoa Willd.) as a potential cash crop halophyte. Aust. J. Crop Sci. 2012, 6, 357–368. [Google Scholar]
- Mujahid, N.; Muhammad, S.; Aysha, K.; Muhammad, A.W. Modulations Induced by seed priming of strigolactone (GR24) in morpho physiological and biochemical attributes of ajwain (Trachyspermum ammi L.) under salt stress. J. Plant Growth Regul. 2022, 1–14. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Photosynthetic Rate (μmol CO2 m−2 s−1) | Transpiration Rate (mmol H2O m−2 s−1) | Stomatal Conductance (mmol m−2 s−1) | Ψw (−MPa) | Ψs (−MPa) | Ψp (−MPa) | Proline (μmol g−1 f.wt.) |
|---|---|---|---|---|---|---|---|
| Salt stress | 424.86 ** | 2.12 ** | 17578.13 ** | 0.102 ** | 2.797 ** | 3.968 ** | 8173.56 ** |
| GR24 | 94.55 ** | 0.03 ns | 78.13 ns | 0.003 ** | 0.161 ** | 0.124 ** | 1114.92 ** |
| S × GR24 | 7.07 ns | 0.05 * | 936.46 ** | 5.736 ** | 0.063 ** | 0.056 ** | 577.219 ** |
| Error | 5.879 | 0.017 | 132.291 | 1.767 | 0.001 | 0.001 | 1.780 |
| Source of variation | MDA (μmol g−1 f.wt.) | GB (μmol g−1 dry wt.) | TSP (mg m−1 f.wt.) | H2O2 (μmol g−1 f.wt) | SOD (Units mg−1 protein) | POD (Units mg−1 protein) | CAT (Units mg−1 protein) |
| Salt stress | 93.213 ** | 2478.406 ** | 3.485 ** | 15292.74 ** | 1674.03 ** | 239.92 ** | 4368.37 ** |
| GR24 | 142.011 ** | 1185.009 ** | 4.155 ** | 639.68 ** | 1194.86 ** | 2.71 ** | 659.46 ** |
| S × GR24 | 14.346 ** | 173.373 ** | 0.453 ** | 110.83 ** | 188.41 ** | 0.42 ns | 99.37 ** |
| Error | 2.888 | 2.054 | 0.066 | 2.783 | 5.076 | 0.141 | 9.178 |
| Source of variation | Root Na+ (mg g−1 Dry wt.) | Root K+ (mg g−1 Dry wt.) | Root Ca2+ (mg g−1 dry wt.) | Shoot Na+ (mg g−1 Dry wt.) | Shoot K+ (mg g−1 Dry wt.) | Shoot Ca2+ (mg g−1 Dry wt.) | Root fresh wt. (g) | Root dry wt. (g) | Shoot fresh wt. (g) | Shoot dry wt. (g) |
|---|---|---|---|---|---|---|---|---|---|---|
| Salt stress | 300.125 ** | 202.507 ** | 45.125 ** | 99.757 ** | 205.031 ** | 23.632 ** | 22.680 ** | 0.264 ** | 2156.767 ** | 21.912 ** |
| GR24 | 5.177 ** | 5.591 ** | 1.302 * | 2.091 ns | 5.718 ** | 6.716 ** | 2.294 ** | 0.020 ** | 258.936 ** | 1.944 ** |
| S × GR24 | 0.729 ns | 0.403 ns | 0.187 ns | 0.257 ns | 0.468 ns | 1.153 ns | 0.106 ns | 0.004 ns | 29.849 ** | 0.797 ** |
| Error | 0.703 | 0.617 | 0.380 | 0.752 | 0.890 | 0.565 | 0.175 | 0.001 | 4.562 | 0.088 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahsan, M.; Zulfiqar, H.; Farooq, M.A.; Ali, S.; Tufail, A.; Kanwal, S.; Shaheen, M.R.; Sajid, M.; Gul, H.; Jamal, A.; et al. Strigolactone (GR24) Application Positively Regulates Photosynthetic Attributes, Stress-Related Metabolites and Antioxidant Enzymatic Activities of Ornamental Sunflower (Helianthus annuus cv. Vincent’s Choice) under Salinity Stress. Agriculture 2023, 13, 50. https://doi.org/10.3390/agriculture13010050
Ahsan M, Zulfiqar H, Farooq MA, Ali S, Tufail A, Kanwal S, Shaheen MR, Sajid M, Gul H, Jamal A, et al. Strigolactone (GR24) Application Positively Regulates Photosynthetic Attributes, Stress-Related Metabolites and Antioxidant Enzymatic Activities of Ornamental Sunflower (Helianthus annuus cv. Vincent’s Choice) under Salinity Stress. Agriculture. 2023; 13(1):50. https://doi.org/10.3390/agriculture13010050
Chicago/Turabian StyleAhsan, Muhammad, Hira Zulfiqar, Muhammad Ansar Farooq, Sajjad Ali, Aasma Tufail, Shamsa Kanwal, Muhammad Rashid Shaheen, Mateen Sajid, Hera Gul, Aftab Jamal, and et al. 2023. "Strigolactone (GR24) Application Positively Regulates Photosynthetic Attributes, Stress-Related Metabolites and Antioxidant Enzymatic Activities of Ornamental Sunflower (Helianthus annuus cv. Vincent’s Choice) under Salinity Stress" Agriculture 13, no. 1: 50. https://doi.org/10.3390/agriculture13010050