

Pathogenic and Genetic Characterization of Fusarium verticillioides Strains Collected from Maize and Sorghum Kernels

Dipartimento Territorio e Sistemi Agro-Forestali, Università di Padova, AGRIPOLIS, Viale dell’Università 16, 35020 Legnaro, PD, Italy
*
Author to whom correspondence should be addressed.
Agriculture 2023, 13(1), 105; https://doi.org/10.3390/agriculture13010105
Submission received: 9 December 2022
/
Revised: 27 December 2022
/
Accepted: 27 December 2022
/
Published: 29 December 2022
(This article belongs to the Special Issue Integrated Management of Fungal Diseases in Crops)
Abstract
:Commercial and sanitary qualities of cereals are important and contribute to the determination of the final application and destination of the grain and, thus, farmer profit. Although considered a minor crop in Italy, in the last years, the cultivation area of sorghum (Sorghum bicolor L. Moench) has been increasing and is often adjacent or subsequent to the most common maize (Zea mays L.) fields, with the risk of sharing mycotoxigenic fungi that are common to both cereals, such as Fusarium verticillioides. The present work aims to clarify if a fungal physiological characteristic such as fumonisin production and some plant growth parameters such as effects on germination, root and shoot development and on tissue necrosis may be common and shared among F. verticillioides strains belonging to maize and sorghum, especially if present on adjacent fields at the same time. Sorghum kernels are commonly less colonized and contaminated by F. verticillioides and fumonisins than maize, and, at least in part, this aspect could be explained by the higher frequency of low-producing strains obtained from sorghum compared to those obtained from maize. Moreover, although no clear pathogen specificity was found, the results relating to the parameters analyzed suggest a host preference by F. verticillioides isolated from maize that affects maize seedlings with greater severity as well as F. verticillioides strains from sorghum versus sorghum. Five microsatellite markers showed minor differences between the two sets of strains but could not reliably discriminate between hosts. The resulting data suggest that in adjacent maize and sorghum crops, the population of F. verticillioides selected during sorghum cultivation may not represent a particular mycotoxigenic or pathogenic issue for maize cultivation.
1. Introduction
Cereals are increasing in importance due to the increased worldwide demand for human food, livestock feed, raw materials for industrial production and sources of biomass for energy production [1]. In recent years, these aspects led farmers to increase the cultivation of cereals, making the presence of maize (603,000 ha in 2020) [2] and sorghum (52,000 ha in 2020) [2] in Italy more frequent in crop succession as well as in adjacent fields and areas. Since these cereals share some common pathogens, the growth of maize and sorghum plants in close proximity, or in rotation, could increase the occurrence of diseases and mycotoxin accumulation, as well, due to shared pathogens, increasing the risk to reduce yield and quality production [3]. Fusarium verticillioides is an important fungal pathogen with a wide range of cereal hosts [4]. Although F. verticillioides is the main pathogen for maize in Italy, being responsible for Fusarium ear rot, this species is also an inhabitant of sorghum and is responsible for diseases in this plant species such as Sorghum grain mold and stalk and root rot [5,6,7,8]. Cereal colonization by F. verticillioides is frequently associated with the accumulation of fumonisins (FBs) produced in the field or during storage [9], representing a threat for its consumption as food and feed [10]. The occurrence of FBs in maize kernels has been largely documented, while the level detected in sorghum is generally low; on the other hand, the co-occurrence of different mycotoxins in sorghum samples (aflatoxin, fumonisin, trichothecenes) is not uncommon [11,12]. The generally low colonization of F. verticillioides on sorghum has been associated with limited systemic growth in plant tissues [13] and production of fungitoxic compounds (i.e., phytoalexins) that inhibit pathogen growth [14]. In addition, some positive grain characteristics, such as hardness and polyphenol accumulation in the pericarp, contribute to resistance to fungal infection [15]. These traits are not usually associated with commercial maize hybrids, which, in fact, are more susceptible to disease and mycotoxins. Therefore, this study was conducted to evaluate possible differences in pathogenicity, FB production and genetic variability among strains of F. verticillioides that colonize maize and sorghum. Since these cereals cultivated in succession or in proximity could pose a risk of F. verticillioides-related disease and FBs occurrence, the comprehension of these aspects can help their management.
2. Materials and Methods
2.1. Fusarium spp. Isolation
Maize ears (Kefrancos; KWS Italia) and sorghum panicles (KSH4G04, KWS Italia) were randomly collected at physiological maturity (BBCH 89, fully ripe; approximately 3 kg of kernels per three samples, each) from adjacent fields in Rovigo (Italy). Samples were homogenized and ground to flour with a Grindomix GM200 Laboratory Mill (Retsch Italy, Torre Boldone, Italy), and serial dilutions were spread on Potato Dextrose Agar (PDA (Difco, Detroit, MI, USA) plates to estimate fungal incidence [16]. Five plates for each dilution were made. Plates were incubated for seven days at 25 °C in the dark, and developed colonies were counted to estimate fungal colonization (CFU∙g−1). Colonies were morphologically identified [17], and F. verticillioides isolates were subcultured to obtain single spore cultures [18]. Approximately 100 and 70 single-spore isolates were obtained from maize and sorghum kernels, respectively. Twenty-five F. verticillioides isolates belonging to maize and sorghum samples (hereafter named M- and S-strains), were randomly selected for subsequent analyses. Other Fusarium spp. (F. proliferatum, F. oxysporum, F. semitectum, F. nygamai, F. equiseti and F. crookwellense) were morphologically identified according to Leslie and Summerell [17].
2.2. Molecular Identification and Mating Type Determination
To molecularly confirm the identification of F. verticilliodes at the species level, the M- and S-strains were grown in 50 mL Potato Dextrose Broth (PDB Difco, Detroit, MI, USA) for 7 days (25 °C in the dark under 100 rpm rotatory shaking); mycelia were collected, washed with sterile water, squeezed and stored at −80 °C before DNA extraction. Genomic DNA was extracted from 60 isolates (30 M-strains and 30 S-strains) according to Doyle [19], and a molecular analysis was performed to confirm strain identity at the species level by PCR applying a specific species assay with VER1 and VER2 primers [20]. Twenty-five isolates from both cereals detected as F. verticillioides strains by PCR were selected for the analyses. Mating type identification for each strain was tested by multiplex PCR by using primers to amplify the MAT-1.1 and MAT-1.2 idiomorphs according to Steenkamp et al. [21]. All of the strains produced a fragment of ~200-bp or 800 bp for MAT-1.1 and MAT-1.2, respectively.
2.3. Total Fumonisin Production
Conidial suspensions for FBs production (FB1, FB2 and FB3) and subsequent inoculations were obtained by growing Fusarium verticillioides strains (M- and S- strains) on Spezieller Nährstoffarmer Agar (SNA) plates for 15 days at 25 °C in the dark. Conidia were collected with sterile water and 0.02% Tween-20 (Sigma–Aldrich, Milan, Italy), and the concentration was adjusted to 106 conidia mL−1. FBs production by selected strains was carried out according to Vismer et al. [12] with some modification. Briefly, twenty grams of maize flour with a low FBs content (<100 μg kg−1) were put in glass Petri dishes and autoclaved for 1 h at 121 °C. To each plate, 20 mL of sterile water was added and left to equilibrate overnight to obtain a moisture percentage of 50%. Plate inoculation was performed by three droplets of each fungal strain (50 μL of a 106 CFU mL−1, each) equally outdistanced. Thirty plates per strain as replicates and three inoculations per plate were used to minimize small variations in starting inoculum and toxin production (Vismer et al. 2019). Plates were incubated at 25 °C for 21 days in the dark and then dried overnight at 60 °C. After drying, the 30 replicates for each strain were pooled, mixed well, and ground to powder in a Laboratory Mill (Retsch Italy, Torre Boldone, Italy) The mill was thoroughly cleaned following grinding of every different strain. Ground powder was stored at 4 °C until analyzed. FBs were extracted by suspending 5 g of maize flour in extraction buffer (VICAM Milford, MA, USA) according to the manufacturer’s instructions. FBs content was then quantified by using the immunostrip Fumo-V Aqua test from VICAM (Milford, MA, USA). The extraction solution of a sample exceeding the maximum range of 100,000 ppb was further diluted 1:1 with extraction buffer. FBs content linearity in diluted samples was verified with internal controls and HPLC-certified samples in endowment to our laboratory.
2.4. Pathogenicity Assays
Fusarium verticillioides strains were analyzed on maize and sorghum seedlings for their capacity to inhibit the kernel germination and the root and leaf development and to produce necrotic lesions on detached leaves (DLA). Kernels were surface sterilized with H2O2 10% for 15 min (maize) or 5 min (sorghum) and rinsed thoroughly with sterile water. Thirty sterile kernels from maize and sorghum for each isolated strain (three replicate of ten kernels) were soaked for 30 min in a 106 mL−1 conidial suspension and placed into Petri dishes on sterile water-saturated filter paper according to Yates et al. [22]. Uninoculated kernels were soaked in sterile water and used as the control. After 7 days, pathogenetic abilities were estimated by determining the percentage of not-germinated kernels calculated as seed with radicle >5 mm long (germination inhibition; Gi%) and reduced length of roots (root inhibition; Ri%) and shoots (shoot inhibition; Si%) in comparison to the same tissues from the uninfected control.
DLA was performed according to Browne and Cooke [23]. Maize and sorghum sterile kernels were grown at 22 °C under a 16 h of photoperiod and 75% RH up to the fourth true leaf stage. Leaf portions of approximately 10 cm (15- and 21-day old seedlings for maize and sorghum, respectively) were detached from the second and third true leaves and immediately used in the experiment. Leaves were disinfected with ethanol 10% and placed into sterile Petri dishes on filter paper saturated with water, three leaves per Petri dish (150 mm diameter), five dishes as replicates. For each strain, to avoid the diffusion of inoculum, two SNA plugs of actively growing mycelium were applied to opposite parts of the adaxial surface of each leaf. Plates containing inoculated leaves were placed at 25 °C for two days in the dark and subsequently in the light to favor fungal pigmentation and obtain a better image contrast. Seven days after inoculation, leaves were digitalized, and area extension with necrosis or colonizing mycelium were measured through ImageJ 1.8.0 software.
2.5. Microsatellite Marker Analysis
Genetic similarity through microsatellite analysis was carried out by amplification of Fv-98, Fv-114, Fv-120, Fv-312 [24] and Fv-338 markers [25], the sequences of which are shown in Table 1. Markers were selected on the basis of high polymorphism produced and for their validation confirmed by two different studies [24,25]. Moreover Fv-114, Fv-120 and Fv-312 markers do not amplify Fusarium thapsinum and Fusarium andiyazi [25], common species in sorghum morphologically similar to F. verticillioides [5], providing a further confirmation of the appropriate classification of selected strains as F. verticillioides. Approximately 1 ng DNA was used to amplify selected markers by PCR amplification (0.3 mM dNTP, 0.2 μM of each primer, 1.5 mM MgCl2 and 0.5 U of Taq DNA polymerase; total volume of 20 μL) in an Eppendorf Mastercycler (Eppendorf, Hamburg, Germany). The following amplification program was used: 5 min initial denaturation at 94 °C, followed by 35 cycles of 30 s denaturation at 94 °C, 30 s annealing at 59 °C, and 30 s extension at 72 °C. PCR was completed with a 7 min final extension at 72 °C. The amplified fragments were separated by electrophoresis at 3.5 V/cm in a 3.5% (wt/vol) MS-8 agarose gel (Thermo Scientific, Milan, Italy) in 1× TAE buffer. The GeneRuler 100 bp DNA Ladder (Thermo scientific, Milan, Italy) was used. Fragment sizing was determined with the Gel Analyzer 19.1 software (www.gelanalyzer.com) on the basis of two independent electrophoretic separations. A marker was considered to be polymorphic if the amplified product was present at different lengths, present in some strains or absent in others. To avoid misinterpretation among close PCR fragments (bands), only those varying in size by 4 bp or more were considered different, and so bands were scored as presence (1) or absence of the band (0). Marker’s profile was used to calculate diversity index, which included Shannon’s index (H’; diversity of polymorphism in a population), Pielou’s index (J; evenness of polymorphism distribution) and the Bray Curtis dissimilarity index (BC; difference of polymorphism between two populations). Diversity indexes were estimated according to their respective authors [26,27,28].
2.6. Statistical Analysis
Data belonging to FBs production and pathogenicity assays were analyzed for their differences by using t-test, ANOVA test or nonparametric Mann–Whitney U test. Each challenge among strains and cereals was considered a pathogenic interaction. Differences in strain frequencies between the two populations were analyzed with Yates’s chi square test. Correlations between total FBs production for each strain and their pathogenic abilities were performed by calculating the Pearson’s coefficients and their probabilities. Principal component analysis (PCA) performed on data obtained by pathogenicity assays was carried out by using Pearson’s correlation matrix with the only correlated variables (FBs, mGi%, sGi%, mRi%, sRi% and sNa). Statistical calculations were performed in XLSTAT 2016 software (Addinsoft, Paris, France). Genetic distances on the basis of microsatellite profiles were obtained by using the Dice coefficient similarity matrix (for binary data). Resulting distances were applied to DendroUPGMA [29] to draw a dendrogram constructed according to the Unweighted Pair Group Method with Arithmetic Mean (UPGMA) algorithm with 100 bootstrap. Numbers on the branches show bootstrap support greater than 30%.
3. Results
3.1. Incidence of Fusarium verticillioides and Related Species in Maize and Sorghum Samples
Quantification of Fusarium colonies in kernel (Table 2) revealed that maize samples were significantly more colonized than those of sorghum (16.66-fold; p < 0.001). F. verticillioides, F. proliferatum and F. oxysporum accounted for the majority of Fusarium species in kernels (99.0% for maize and 90.1% for sorghum) but with a strong prevalence of F. verticillioides. Sorghum kernels were also colonized at a low level (9.9%) by F. semitectum, F. nygamai, F. equiseti and F. crookwellense, while these species were almost absent (1.0%) in maize samples.
3.2. Mating Type Determination
Determination of F. verticillioides mating type (Table 3) revealed that, among the M-strains, fifteen (60%) carried the MAT-1 allele and ten (40%) the MAT-2 allele. The same analysis conducted on S-strains shown that sixteen (64%) and nine (36%) were MAT-1 and MAT-2 strains, respectively. Statistical analyses revealed no differences in MAT frequencies between the two groups (p = 0.77).
3.3. Fumonisin Quantification
Preliminary quantification of mycotoxin in cereal samples (3 kg of maize and sorghum kernels per three samples, each) collected at the same time from adjacent fields and used for fungal isolation revealed a higher FBs contamination for maize (4050 μg kg−1 on average) with respect to sorghum samples (<200 μg kg−1 on average).
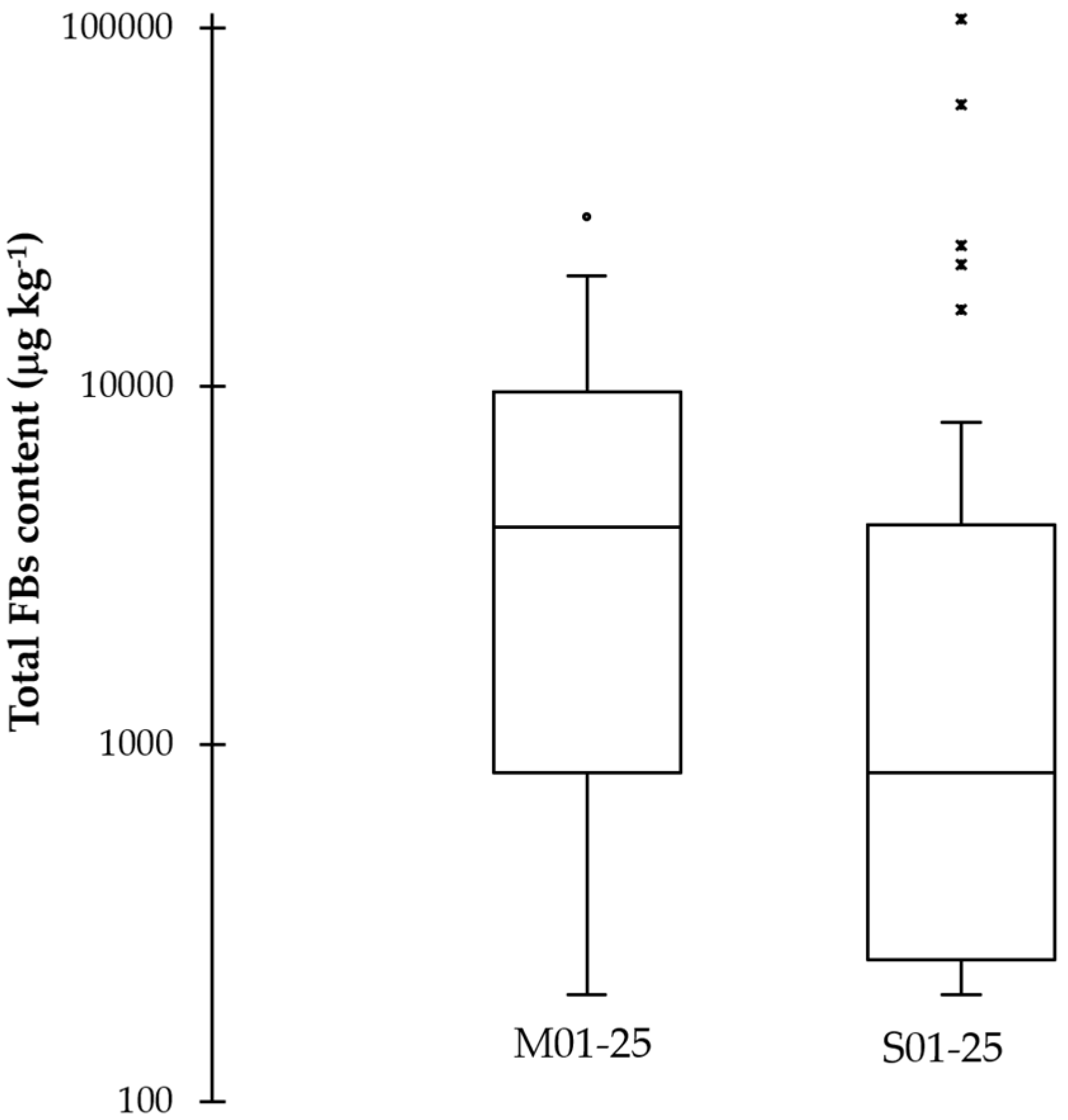
FBs determination of monoconidial strains carried out on producing the toxins under in vitro conditions showed a great variability among strains (Table 3). In particular, three M-strains (M05, M01 and M25) produced low amounts of FBs (<500 μg kg−1), while ten and twelve M-strains produced medium (500–5000 μg kg−1) and high amounts (>5000 μg kg−1), respectively. Eleven, eight and six S-strains produced low, medium and high amounts of FBs, with the maximum amount produced by S16 with 106,300 μg kg−1. The frequency of low FBs-producing strains was higher (p = 0.027) for strains isolated from sorghum (11/25) than strains isolated from maize (3/25). Concerning the FBs production by the two F. verticilliodes populations (Figure 1), statistical analyses revealed the presence of one (M19: 29,600 μg kg−1) and five strains (S16: 106,300; S06: 61,200; 21,900, S04: 24,800 and S25: 16,400 μg kg−1) with outlier values for maize and sorghum, respectively. FBs production, as a whole, did not determine significant differences (p = 0.651) between the two-average production (average 6820 ± 1543 μg kg−1 for M-strains and 10,244 ± 4810 μg kg−1 for S-strains). In contrast, the same analysis, performed omitting the outlier values, differentiated (p = 0.002) maize strains (average 5867 ± 1269 μg kg−1) from sorghum ones (average 1275 ± 432 μg kg−1).
3.4. Effect of Fusarium verticillioides Strains on Seed Germination
F. verticillioides strains belonging to maize and sorghum were tested on kernels to evaluate the differential effects on seed germination. Germination effects may have little impact on fumonisin contamination of the ear, but susceptibility of young tissues could result in increased environmental inoculum and an increased infection during ear ripening. Results were reported as percentage of germination inhibition of maize (mGi%) and sorghum (sGi%) germination compared to the uninfected control (Supplementary Materials S1). The germination of maize and sorghum seeds infected with M-strains was significantly affected by ten (22.7% average inhibition) and eleven strains (31.8% average inhibition), respectively, with respect to uninfected seeds. With regard to the test conducted with S-strains, only seven strains affect maize seed germination (16.7% average inhibition), while five strains were able to significantly reduce sorghum seed germination (28.0% average inhibition). Moreover, the comparison of data revealed that only six strains (M07, M13, M22, M24, S06 and S16) were able to reduce germination in both cereal species. Regarding seed germination, S-strains were found to be more pathogenic on sorghum germination (p ≤ 0.001), but no preference for M-strains was evidenced (p = 0.563). No correlation was observed between the ability of the strains to produce FBs and their effect on seed germination (p = 0.328 for maize; p = 0.097 for sorghum).
3.5. Effect of Fusarium verticillioides Strains on Root Development
Root length development was evaluated to identify a detrimental effect by F. verticillioides infection on roots. Results were reported as percentage of inhibition of root development of maize (mRi%) and sorghum (sRi%) compared to roots from the uninfected control. Concerning the root evaluation after infection with M-strain, 56% of strains reduced root length on maize seedlings, and 36% of strains reduced root length on sorghum seedlings, with an average inhibition of 60.0 and 36.5%, respectively. Regarding S-strains, 65.4% of strains reduced root length on maize seedlings (23.5% reduction) and 72% on sorghum seedlings but with a lower inhibition (17.0% reduction). From the comparison of data, four M-strains (M03, M04, M07 and M16) and ten S-strains (S01, S04, S05, S06, S08, S09, S12, S15, S24 and S25) significantly reduced root length in both plant species. Conversely, six M- and two S-strains did not affect root development (Supplementary Material S2). With respect to root development, the inhibition of root length by M-strains was higher in both maize (p = 0.001) and sorghum plants (p ≤ 0.0001) when compared to S-strains. A weak but positive correlation was observed between FBs and the effect on maize root length (rho = 0.105; p = 0.022); however, the same correlation was not found on sorghum roots (rho = 0.004; p = 0.767).
3.6. Effect of Fusarium verticillioides Strains on Shoot Development
Shoot length development was evaluated to identify a detrimental effect by F. verticillioides infection on shoots. Results were reported as percentage inhibition of maize and sorghum shoot development (mSi% and sSi%, respectively) compared to shoots from the uninfected control. Shoot length measured after infection with M-strains indicates that, on average, a significant inhibition for maize and sorghum seedlings was attributed to sixteen and six strains (on average mSi: 44.8% and sSi: 42.5%), respectively. Similarly to the effect on roots, a large number of S-strains reduced shoot length (60% of strains of maize, 68% of strains of sorghum), with an average inhibition value calculated for significant interactions as 46.7% (mSi%) and 35.4% (sSi%), respectively. From the comparison of the data, 12% of M-strains (M02, M06 and M24) and 44% of S-strains (S01, S02, S03, S04, S10, S11, S15, S16, S19, S23 and S24) significantly reduced the length of the shoots in both plant species. Conversely, ten strains did not affect root development (Supplementary Materials S3). No differences were observed between M- and S-strains with respect to their ability to reduce shoot development in maize (p = 0.838) and sorghum (p = 0.541). No correlation was observed between shoot development and FBs production for both maize (rho = 0.057; p = 0.696) and sorghum (rho = 0.089; p = 0.497).
3.7. Effect of Fusarium verticillioides Strains on Detached Leaf Assays (DLA)
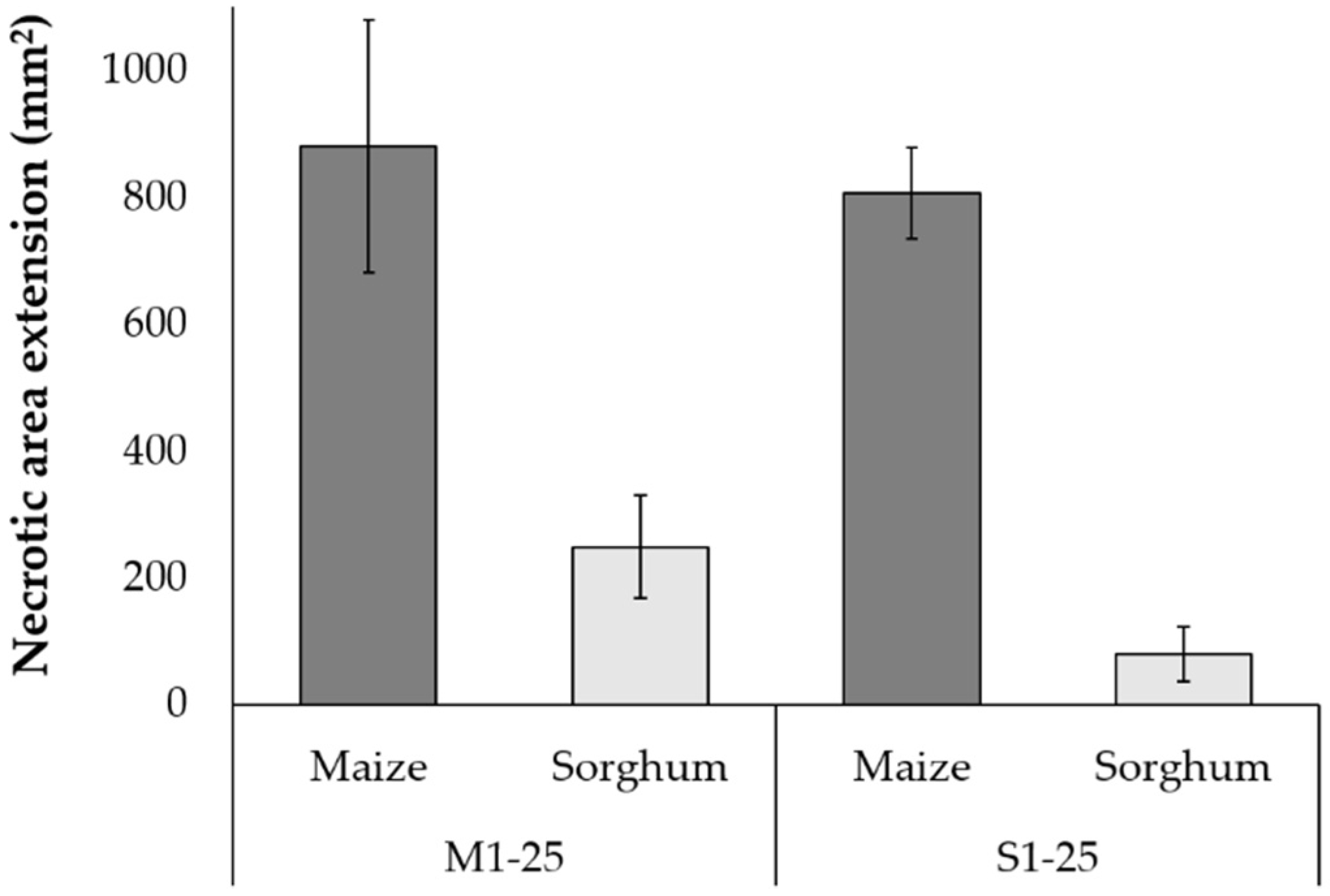
Necrotic area (NA) was evaluated through DLA to identify a detrimental effect by F. verticillioides infection on leaves. Results were reported as necrotic extension area for maize (mNA) and sorghum (sNA) (Supplementary Materials S4). Results from DLA assays showed that necrotic areas produced by M-strains were generally larger (p ≤ 0.0001) on maize (average 131.8 mm2; range from 4.9 to 350.5 mm2) than on sorghum leaves (on average 26.5 mm2; range from 0.0 to 129.1 mm2). Moreover, while all M-strains produced at least a small necrotic area on maize leaves, twelve produced little or no symptom on sorghum. Similarly, fifteen S-strains produced significant necrotic areas on maize leaves (average 103.2 mm2; range from 13.5 to 151.4 mm2), while only five of them significantly affected sorghum leaves (average 39.8 mm2; range from 0.0 to 95.4 mm2), and twenty strains produced no symptoms. Five M-strains (M02, M03, M06, M10, M12 and M20) and two S-strains (S23 and S25) significantly affected both cereals. On average, M-strains (p ≤ 0.0001) and S-strains (p ≤ 0.0001) produced a larger necrosis on maize leaves rather than on sorghum ones (Figure 2). However, based on the overall average NA values, it was not possible to detect any difference between maize leaves colonized by M- or S-strains (Figure 2). The bars corresponding to the NA values on sorghum leaves (Figure 2) also contain the effect of twelve M-strains and twenty S-strains unable to cause symptoms on these tissues. No correlation was observed between FBs production and necrotic area development in maize (rho = 0.017; p = 0.363) and sorghum (rho = 0.022; p = 0.878).
3.8. Clusterization of Fusarium verticillioides Strains Based on Pathogenic Parameters
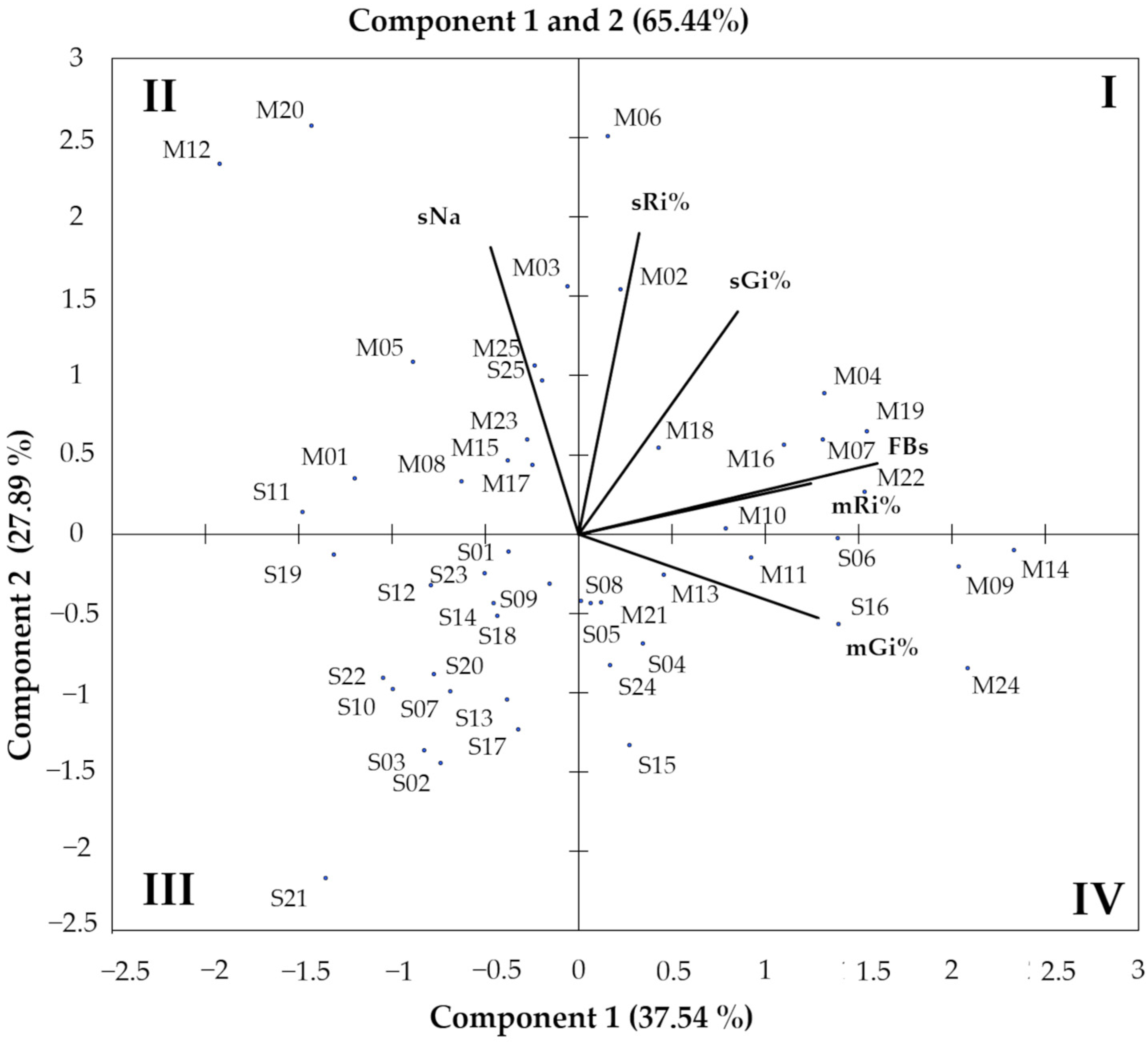
FBs production, inhibition of seed germination, root and shoot development and leaf necrosis were the pathogenic parameters considered for the characterization of the strains. Principal component analysis (PCA) was performed to visually investigate these different characteristics and the similarity between the variable analyzed (Figure 3). No significant correlation was found between the inhibition of shoot development in both cereals and maize necrotic areas with the other considered parameters (Figure 3). Cluster analysis showed that germination and roots inhibition in maize and sorghum (sGi%; mGi%; mRi%; sRi%) were positively correlated with FBs production with different correlation coefficients. Positioning of F. verticillioides strains on the biplot, resulting from PCA, showed that most of the M-strains were located in the same direction of variables (quadrant I, II and IV), while S-strains were mainly located in or near the quadrant III.
In Figure 3, quadrant I includes only M-strains characterized by a high FBs production and a low inhibition of maize seed germination. Quadrant II consists of strains with a low production of fumonisins and with a small inhibitory effect on maize germination and sorghum root development but with significant necrotic areas on sorghum (ten M-strains; two S-strains). Quadrant III includes 16 out of 25 S-strains characterized by low pathogenic effects for each considered parameters. Quadrant IV consists of strains from both populations (seven S-strains; six M-strains) characterized by high FBs production and maize seed germination inhibition but low inhibitory effects on sorghum.
3.9. Microsatellite Marker Analysis
F. verticillioides strains belonging to maize and sorghum were analyzed for their microsatellite profiles to verify a closer similarity among F. verticillioides isolated by different hosts. Five microsatellite markers were analyzed by PCR, and all fungal strains amplified a single band (Supplementary Materials S5). Data in Table 4 show that for Fv-98, Fv-114, Fv-312 and Fv-338 markers, the S-strains presented generally higher values in terms of polymorphism and diversity index (Shannon’s index H’ and Pielou’s evenness index J’) than M-strains. On the contrary, polymorphisms identified for Fv-120 marker were higher for the M-strains. Minimum and maximum values calculated for Bray Curtis dissimilarity values (BC) were produced by Fv-98 (BC 0.5) and Fv-114 (BC 0.84), respectively. Polymorphic profile for each marker and strain are reported in Supplementary Materials S6 and S7.
3.10. Fusarium verticillioides Clusterization Based on Microsatellite Markers
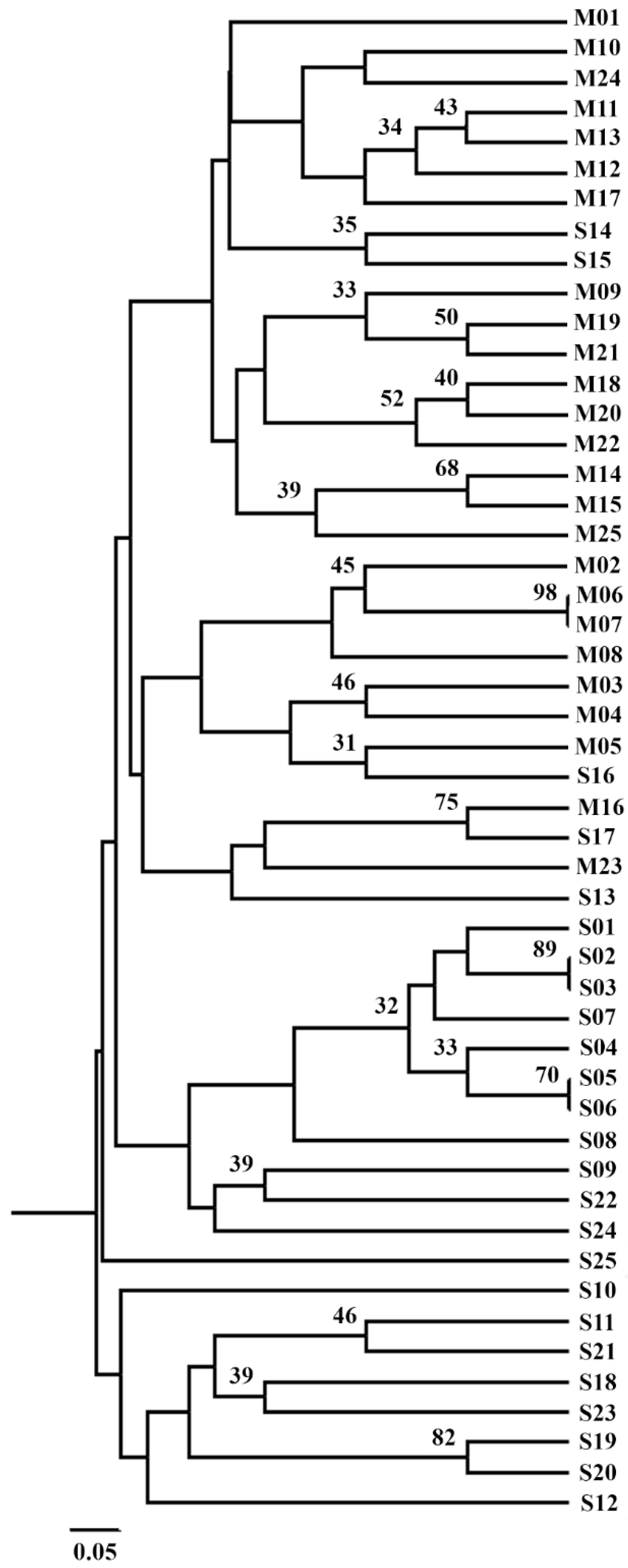
F. verticillioides strains belonging to maize and sorghum were genotyped using five microsatellite markers, and these data were analyzed by UPGMA to cluster together strains with similar microsatellite pattern (Figure 4). The low bootstrap support did not allow us to identify different clusters. However, genetic relationships obtained by microsatellite analyses showed that M-strains tended to group separately from S-strains, but without a statistical support. The cophenetic correlation coefficient [30] was calculated, and a value of 0.767 indicated a good representation of the similarity matrix.
4. Discussion
Sorghum and maize were infected with a pool of fungal pathogens and mycotoxigenic species belonging to Fusarium genus. Since Fusarium verticillioides in Italy represents the most common mycotoxigenic species of maize, this study aimed to elucidate if the Fusarium inoculum isolated from maize and sorghum can reciprocally represent a pathogenic and toxigenic issue. To date, maize and sorghum rotation or succession have been studied for agronomical purpose [31,32], and very few studies have investigated pathogenic aspects [33,34]. The isolation of Fusarium species from maize and sorghum kernels, grown in adjacent fields, shows that F. verticillioides is the most common species in these grains [35,36,37]. In particular, in this study, F. verticillioides represents 88.8% of Fusarium spp. in maize kernels and 59.3% of that in sorghum kernels. Moreover, while F. verticillioides, F. proliferatum and F. oxysporum represent almost all the Fusarium species occurring in maize, the kernels in sorghum have been colonized by other less abundant Fusarium species, usually present in this cereal worldwide [38,39]. A further difference is represented by fungal colonization, which is, in our samples, 16.5-fold higher in maize than in sorghum and may contribute to the different FBs content in the collected samples. Random selection of the two F. verticillioides populations isolated from maize and sorghum samples led us to determine the mating types for sexual reproduction [40,41,42] and revealed a very similar MAT1:MAT2 proportion, demonstrating that any potential pathogenic trait could be randomly transferred between these Fusaria populations. Indeed, through sexual reproduction, fungi recombine their genetic material and increase the diversity in the offspring, which may provide some benefits for the survival of species under different environmental conditions. A previous study [43] conducted in Brazil on F. verticillioides isolated from maize and sorghum samples reported instead a quite different MAT1:MAT2 proportion equal to 5:17 and 14:7 for strains from maize and sorghum, respectively.
With respect to the average amount of fumonisins produced by the F. verticilliodes strains analyzed, no difference was found between the two populations. Yet, the percentage of low-producing strains (<500 μg kg−1) was higher for sorghum (44%) than for maize (12%). Moreover, due to lateral flow sensitivity (≥200 μg kg−1), we were unable to determine for some strains whether no fumonisin was produced at all or the production was at a very low amount; nonetheless, the percentage of putative no-producing strains found in maize appeared not so different from other studies conducted in Italy [44] and worldwide [45,46]. On the contrary, six S-strains (24%) were found to be characterized by a high fumonisin production (>5000 μg kg−1), whereas in a study carried out in India, only a little number of F. verticilliodes strains isolated from sorghum (around 6%) were characterized by a similar fumonisin production [45]. As previously observed for maize [44], present data, also considering the outlier values on fumonisins, support findings that F. verticillioides strains collected from sorghum in a single location might show a great variability in fumonisin production. Data obtained with this study showed a weak but positive correlation between reduced maize root development and mycotoxin content; however, no other correlations have been identified between fumonisin biosynthesis and pathogenesis in the different tissues analyzed of maize and sorghum. With respect to pathogenic aggressiveness between sorghum and maize strains, a clear infection pattern was not identified, and the ability to produce symptoms was different for each strain and tissue challenged by infection, and different strains were able to induce different symptoms. Seed germination has been less influenced by F. verticillioides strains both from maize and sorghum, even if seed decay and germination reduction are symptoms observed under stressful conditions [44,47,48]. In particular, our data suggest that a limited number of strains may affect germination of maize and sorghum seeds, although in general at a low level. Only six strains (four M and two S) were able to significantly reduce seed germination in both cereals, while 42% of M-strains significantly affected maize or sorghum germination, and the percentage decreased to 24% for S-strains. Furthermore, maize strains affected germination of both cereals with similar severity, and strains belonging to sorghum demonstrated to be more pathogenic on sorghum seeds. Conversely to germination, root and shoot development appeared more influenced by pathogens, with an increased number of strains able to reduce tissue development. Although total significant pathogenic interactions for S-strains (66% for root and shoot as a whole) were higher than those for M-strains (45%), the latter generally showed a higher severity, mainly due to the negative impact on root development. As previously suggested, at least for root development in maize, a role could be played by the higher fumonisin production determined in M-strains.
One of the most significant parameters, however, could be represented by necrotic areas produced on leaves. All strains were able to produce at least one small necrotic lesion on maize, while less than half of the strains induced symptoms in sorghum. These results highlight a greater susceptibility to F. verticillioides infection of maize rather than sorghum, which could represent an important aspect to be taken into account in the planning of crop successions. The resistance to fungal colonization in sorghum has been associated with both the constitutive presence [49] and induced accumulation of phenolic acids, condensed tannins and flavonoids [50,51,52] and to antimicrobial peptides [53], all together responsible for the reaction of the plant to biotic and abiotic stress [54,55]. Moreover, sorghum appears more resistant than maize to other stress conditions such as drought; in fact, in conditions of dehydration during the flowering stage, a higher drought resistance has been associated with a greater capacity of the antioxidant–enzymatic system of sorghum with respect to that of maize [56]. The features in pathogenicity of the F. verticillioides strains were analyzed through the principal component analysis (PCA), and the pathogenic parameters analyzed (fumonisin production, seed germination, root and shoot development and leaf necrosis) between and within strains highlighted similarities and differences among M- and S-strains. PCA distribution showed that the totality of the M-strains appeared distributed in the three quadrants that contained the considered variables.
Although the M-strains showed a large variability in terms of pathogenic capacity, the same was not supported by data in terms of genetic variability. In fact, four out of five microsatellite markers analyzed (Fv-98, Fv-114, Fv-312 and Fv-338) resulted as more polymorphic for S-strains than M-strains. Although obtained by exploring different genomic traits, these results showed that in our collection area of the F. verticillioides strains obtained from sorghum had a higher genetic diversity than those isolated from maize, as was similarly observed in Brazil [43], highlighting that strains coming from both local or distant regions can share similar peculiarities. This was also manifested by the higher degree of polymorphism detected in the collected S-strains. The higher diversity for S-strains demonstrated by Shannon’s index (that measures the diversity of species in a community) and Pielou’s evenness (that measures diversity along with species richness) suggested a good distribution of markers within groups. Furthermore, Bray Curtis dissimilarity (used to quantify the differences in species composition and abundance between two populations) calculated for each marker showed that, with the exclusion of Fv-98 (BC 0.50), more than half of the strains were different between the two collected groups with a maximum dissimilarity value of 0.84 for Fv-114. The higher genetic diversity observed for S-strains could suggest that the analysis based on the similarity calculated by the biplot would discriminate the strains between two host cereals, but this diversity was not sufficient to achieve this purpose. The resulting dendrogram seemed to differentiate strain isolates from maize and sorghum in most cases, but the bootstrap support for key nodes never exceeded 30%; hence, typical genetic profiles was not identified. The observation that no pattern was specific for F. verticillioides collected from maize and sorghum may suggest that the genetic population feature could be the result of an active genetic recombination occurring in the field, as observed in the extensive area of cultivation of cereals affected by this fungus [57]. Moreover, no similarities were observed among pathogenicity and microsatellite clusterization. Our findings were not surprising since, as previously reported [44], even using different markers the genetic variability in F. verticillioides was not correlated to pathogenicity. The pathogenicity and mycotoxin parameters shown by F. verticillioides strains isolated from sorghum, which were generally characterized by a lower FBs production, indicate that such strains should not pose a substantial risk for fumonisin accumulation in maize kernels; on the other hand, an active genetic recombination cannot be excluded that could select, over time, the development of more aggressive or toxigenic strains.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/agriculture13010105/s1; S1 Germination percentage of maize and sorghum seeds upon F. verticillioides challenge; S2 Root development of maize and sorghum seedlings upon F. verticillioides challenge; S3 Shoot development of maize and sorghum seedlings upon F. verticillioides challenge; S4 Necrotic areas on maize and sorghum detached leaves (DLA assay) upon F. verticillioides challenge; S5 Representative electrophoretic separation of polymorphic bands amplified with Fv-120 primer pairs; S6 Polymorphic bands identified on F. verticillioides strains through microsatellite markers; S7 Microsatellite polymorphic pattern for investigated F. verticillioides strains.
Author Contributions
Conceptualization, D.F. and A.R.; methodology, D.F. and M.M.; formal analysis, D.F. and A.R.; investigation, D.F. and M.M.; resources, A.R.; data curation, D.F.; writing—original draft preparation, D.F.; writing—review and editing, A.R.; supervision, A.R.; funding acquisition, A.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Institutional Grants of University of Padova (DOR2158522/21; DOR2294335/22).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The authors declare that the data supporting the findings of this study are available within the paper. Raw data are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Warr, B.; Schandl, H.; Ayres, R.U. Long Term Trends in Resource Exergy Consumption and Useful Work Supplies in the UK, 1900 to 2000. Ecol. Econ. 2008, 68, 126–140. [Google Scholar] [CrossRef]
- Italian National Institute of Statistics. Estimate of Crop, Flower and Pot Plant Production and Area 2020. Available online: http://dati.istat.it/Index.aspx?lang=en&SubSessionId=38f169a7-56dc-470f-9fcd-d89bc3eff2c5 (accessed on 18 July 2022).
- Bruns, H.A. Controlling Aflatoxin and Fumonisin in Maize by Crop Management. J. Toxicol. Toxin Rev. 2003, 22, 153–173. [Google Scholar] [CrossRef]
- Deepa, N.; Sreenivasa, M.Y. Fumonisins: A Review on Its Global Occurrence, Epidemiology, Toxicity and Detection. J. Vet. Med. Res. 2017, 4, 1093. [Google Scholar]
- Ackerman, A.; Wenndt, A.; Boyles, R. The Sorghum Grain Mold Disease Complex: Pathogens, Host Responses, and the Bioactive Metabolites at Play. Front. Plant Sci. 2021, 12, 660171. [Google Scholar] [CrossRef] [PubMed]
- Mohamed Nor, N.M.I.; Salleh, B.; Leslie, J.F. Fusarium Species from Sorghum in Thailand. Plant Pathol. J. 2019, 35, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Palmero Llamas, D.; Gil Serna, J.; Galvez Patón, L.; Curt Fernández de la Mora, M.; Cara Garcia, M.; de Tello Marquina, J. First Report of Fusarium verticillioides Causing Stalk and Root Rot of Sorghum in Spain. Plant Dis. 2012, 96, 584.2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesso, T.T.; Ochanda, N.; Little, C.R.; Claflin, L.; Tuinstra, M.R. Analysis of Host Plant Resistance to Multiple Fusarium Species Associated with Stalk Rot Disease in Sorghum [Sorghum bicolor (L.) Moench]. Field Crops Res. 2010, 118, 177–182. [Google Scholar] [CrossRef]
- Deepa, N.; Nagaraja, H.; Sreenivasa, M.Y. Prevalence of Fumonisin Producing Fusarium verticillioides Associated with Cereals Grown in Karnataka (India). Food Sci. Hum. Wellness 2016, 5, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wen, J.; Tang, Y.; Shi, J.; Mu, G.; Yan, R.; Cai, J.; Long, M. Research Progress on Fumonisin B1 Contamination and Toxicity: A Review. Molecules 2021, 26, 5238. [Google Scholar] [CrossRef]
- Hanvi, D.M.; Lawson-Evi, P.; De Boevre, M.; Goto, C.E.; De Saeger, S.; Eklu-Gadegbeku, K. Natural Occurrence of Mycotoxins in Maize and Sorghum in Togo. Mycotoxin Res. 2019, 35, 321–327. [Google Scholar] [CrossRef]
- Vismer, H.F.; Shephard, G.S.; Rheeder, J.P.; van der Westhuizen, L.; Bandyopadhyay, R. Relative Severity of Fumonisin Contamination of Cereal Crops in West Africa. Food Addit. Contam. Part A 2015, 32, 1952–1958. [Google Scholar] [CrossRef] [PubMed]
- Dastjerdi, R.; Karlovsky, P. Systemic Infection of Maize, Sorghum, Rice, and Beet Seedlings with Fumonisin-Producing and Nonproducing Fusarium verticillioides Strains. Plant Pathol. J. 2015, 31, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Snyder, B.A.; Nicholson, R.L. Synthesis of Phytoalexins in Sorghum as a Site-Specific Response to Fungal Ingress. Science 1990, 248, 1637–1639. [Google Scholar] [CrossRef]
- Jambunathan, R.; Kherdekar, M.S.; Stenhouse, J.W. Sorghum Grain Hardness and Its Relationship to Mold Susceptibility and Mold Resistance. J. Agric. Food Chem. 1992, 40, 1403–1408. [Google Scholar] [CrossRef] [Green Version]
- Ben-David, A.; Davidson, C.E. Estimation Method for Serial Dilution Experiments. J. Microbiol. Methods 2014, 107, 214–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 0-470-27646-0. [Google Scholar]
- Tuite, J. Plant Pathological Methods. Fungi and Bacteria; CABI: Wallingford, UK, 1969; p. 239. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Mulè, G.; Susca, A.; Stea, G.; Moretti, A. A Species-Specific PCR Assay Based on the Calmodulin Partial Gene for Identification of Fusarium verticillioides, F. proliferatum and F. subglutinans. Eur. J. Plant Pathol. 2004, 110, 495–502. [Google Scholar] [CrossRef]
- Steenkamp, E.T.; Wingfield, B.D.; Coutinho, T.A.; Zeller, K.A.; Wingfield, M.J.; Marasas, W.F.; Leslie, J.F. PCR-Based Identification of MAT-1 and MAT-2 in the Gibberella fujikuroi Species Complex. Appl. Environ. Microbiol. 2000, 66, 4378–4382. [Google Scholar] [CrossRef] [Green Version]
- Yates, I.E.; Arnold, J.W.; Hinton, D.M.; Basinger, W.; Walcott, R.R. Fusarium verticillioides Induction of Maize Seed Rot and Its Control. Can. J. Bot. 2003, 81, 422–428. [Google Scholar] [CrossRef]
- Browne, R.A.; Cooke, B.M. Development and Evaluation of an in Vitro Detached Leaf Assay Forc Pre-Screening Resistance to Fusarium Head Blight in Wheat. Eur. J. Plant Pathol. 2004, 110, 91–102. [Google Scholar] [CrossRef]
- Xu, R.; ZhenDong, Z.; HongJie, L.; CanXing, D.; XiaoMing, W. SSR marker development and analysis of genetic diversity of Fusarium verticillioides isolated from maize in China. Sci. Agric. Sin. 2012, 45, 52–66. [Google Scholar]
- Leyva-Madrigal, K.Y.; Larralde-Corona, C.P.; Calderón-Vázquez, C.L.; Maldonado-Mendoza, I.E. Genome Distribution and Validation of Novel Microsatellite Markers of Fusarium verticillioides and Their Transferability to Other Fusarium Species. J. Microbiol. Methods 2014, 101, 18–23. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley-Inter-Science: New York, NY, USA, 1969. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Vallvé, S.; Palau, J.; Romeu, A. Horizontal Gene Transfer in Glycosyl Hydrolases Inferred from Codon Usage in Escherichia coli and Bacillus subtilis. Mol. Biol. Evol. 1999, 16, 1125–1134. [Google Scholar] [CrossRef] [Green Version]
- Sokal, R.R.; Rohlf, F.J. The Comparison of Dendrograms by Objective Methods. Taxon 1962, 11, 33–40. [Google Scholar] [CrossRef]
- Sauerborn, J.; Sprich, H.; Mercer-Quarshie, H. Crop Rotation to Improve Agricultural Production in Sub-Saharan Africa. J. Agron. Crop Sci. 2000, 184, 67–72. [Google Scholar] [CrossRef]
- Wu, A.-L.; Jiao, X.-Y.; Fan, F.-F.; Wang, J.-S.; Guo, J.; Dong, E.-W.; Wang, L.-G.; Shen, X.-M. Effect of Continuous Sorghum Cropping on the Rhizosphere Microbial Community and the Role of Bacillus amyloliquefaciens in Altering the Microbial Composition. Plant Growth Regul. 2019, 89, 299–308. [Google Scholar] [CrossRef]
- Jaime-Garcia, R.; Cotty, P.J. Crop Rotation and Soil Temperature Influence the Community Structure of Aspergillus flavus in Soil. Soil Biol. Biochem. 2010, 42, 1842–1847. [Google Scholar] [CrossRef]
- Phillips, N.J.; Uyemoto, J.K.; Wilson, D.L. Maize Chlorotic Mottle Virus and Crop Rotation: Effect of Sorghum on Virus Incidence. Plant Dis. 1982, 66, 376–379. [Google Scholar] [CrossRef]
- Cuevas, H.E.; Fermin-Pérez, R.A.; Prom, L.K.; Cooper, E.A.; Bean, S.; Rooney, W.L. Genome-Wide Association Mapping of Grain Mold Resistance in the US Sorghum Association Panel. Plant Genome 2019, 12, 170099. [Google Scholar] [CrossRef] [Green Version]
- Lanza, F.E.; Zambolim, L.; da Costa, R.V.; Queiroz, V.A.V.; Cota, L.V.; da Silva, D.D.; de Souza, A.G.C.; Figueiredo, J.E.F. Prevalence of Fumonisin-Producing Fusarium Species in Brazilian Corn Grains. Crop Prot. 2014, 65, 232–237. [Google Scholar] [CrossRef]
- Silva, J.J.; Viaro, H.P.; Ferranti, L.S.; Oliveira, A.L.M.; Ferreira, J.M.; Ruas, C.F.; Ono, E.Y.S.; Fungaro, M.H.P. Genetic Structure of Fusarium Verticillioides Populations and Occurrence of Fumonisins in Maize Grown in Southern Brazil. Crop Prot. 2017, 99, 160–167. [Google Scholar] [CrossRef]
- Divakara, S.T.; Santosh, P.; Aiyaz, M.; Venkata Ramana, M.; Hariprasad, P.; Nayaka, S.C.; Niranjana, S.R. Molecular Identification and Characterization of Fusarium Spp. Associated with Sorghum Seeds. J. Sci. Food Agric. 2014, 94, 1132–1139. [Google Scholar] [CrossRef]
- Yassin, M.A.; El-Samawaty, A.-R.; Bahkali, A.; Moslem, M.; Abd-Elsalam, K.A.; Hyde, K.D. Mycotoxin-Producing Fungi Occurring in Sorghum Grains from Saudi Arabia. Fungal Divers. 2010, 44, 45–52. [Google Scholar] [CrossRef]
- Fraser, J.A.; Heitman, J. Chromosomal Sex-Determining Regions in Animals, Plants and Fungi. Curr. Opin. Genet. Dev. 2005, 15, 645–651. [Google Scholar] [CrossRef]
- Fraser, J.A.; Heitman, J. Evolution of Fungal Sex Chromosomes. Mol. Microbiol. 2004, 51, 299–306. [Google Scholar] [CrossRef]
- Martin, S.H.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Structure and Evolution of the Fusarium Mating Type Locus: New Insights from the Gibberella fujikuroi Complex. Fungal Genet. Biol. 2011, 48, 731–740. [Google Scholar] [CrossRef] [Green Version]
- da Silva, V.N.; Fernandes, F.M.C.; Cortez, A.; Ribeiro, D.H.B.; de Almeida, A.P.; Hassegawa, R.H.; Corrêa, B. Characterization and Genetic Variability of Fusarium verticillioides Strains Isolated from Corn and Sorghum in Brazil Based on Fumonisins Production, Microsatellites, Mating Type Locus, and Mating Crosses. Can. J. Microbiol. 2006, 52, 798–804. [Google Scholar] [CrossRef]
- Covarelli, L.; Stifano, S.; Beccari, G.; Raggi, L.; Lattanzio, V.M.T.; Albertini, E. Characterization of Fusarium verticillioides Strains Isolated from Maize in Italy: Fumonisin Production, Pathogenicity and Genetic Variability. Food Microbiol. 2012, 31, 17–24. [Google Scholar] [CrossRef]
- Das, I.K.; Kumar, B.S.V.; Ratnavathi, C.V.; Komala, V.V.; Annapurna, A.; Seetharama, N. Toxigenicity of Fusarium Isolates and Fumonisin B1 Contamination in Rainy Season Sorghum (Sorghum bicolor). Indian J. Agric. Sci. 2010, 80, 724–729. [Google Scholar]
- Deepa, N.; Rakesh, S.; Sreenivasa, M.Y. Morphological, Pathological and Mycotoxicological Variations among Fusarium verticillioides Isolated from Cereals. 3 Biotech 2018, 8, 105. [Google Scholar] [CrossRef]
- Venturini, G.; Assante, G.; Toffolatti, S.L.; Vercesi, A. Pathogenicity Variation in Fusarium verticillioides Populations Isolated from Maize in Northern Italy. Mycoscience 2013, 54, 285–290. [Google Scholar] [CrossRef]
- Jensen, F. Relationships between Seed Germination, Fumonisin Content, and Fusarium verticillioides Infection in Selected Maize Samples from Different Regions of Costa Rica. Plant Pathol. 1998, 47, 609–614. [Google Scholar]
- Dykes, L.; Rooney, L.W. Sorghum and Millet Phenols and Antioxidants. J. Cereal Sci. 2006, 44, 236–251. [Google Scholar] [CrossRef]
- Maurya, S.; Singh, R.; Singh, D.P.; Singh, H.B.; Srivastava, J.S.; Singh, U.P. Phenolic Compounds of Sorghum vulgare in Response to Sclerotium rolfsii Infection. J. Plant Interact. 2007, 2, 25–29. [Google Scholar] [CrossRef]
- Shen, S.; Huang, R.; Li, C.; Wu, W.; Chen, H.; Shi, J.; Chen, S.; Ye, X. Phenolic Compositions and Antioxidant Activities Differ Significantly among Sorghum Grains with Different Applications. Molecules 2018, 23, 1203. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Fang, Z. Sorghum Grain: From Genotype, Nutrition, and Phenolic Profile to Its Health Benefits and Food Applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2025–2046. [Google Scholar] [CrossRef] [Green Version]
- Nida, H.; Lee, S.; Li, Y.; Mengiste, T. Transcriptome Analysis of Early Stages of Sorghum Grain Mold Disease Reveals Defense Regulators and Metabolic Pathways Associated with Resistance. BMC Genom. 2021, 22, 295. [Google Scholar] [CrossRef]
- Cheynier, V. Phenolic Compounds: From Plants to Foods. Phytochem. Rev. 2012, 11, 153–177. [Google Scholar] [CrossRef]
- Dicko, M.H.; Gruppen, H.; Barro, C.; Traore, A.S.; van Berkel, W.J.H.; Voragen, A.G.J. Impact of Phenolic Compounds and Related Enzymes in Sorghum Varieties for Resistance and Susceptibility to Biotic and Abiotic Stresses. J. Chem. Ecol. 2005, 31, 2671–2688. [Google Scholar] [CrossRef] [PubMed]
- Takele, A.; Farrant, J. Enzymatic Antioxidant Defence Mechanisms of Maize and Sorghum after Exposure to and Recovery from Pre- and Post-Flowering Dehydration. Acta Agron. Hung. 2009, 57, 445–459. [Google Scholar] [CrossRef]
- Ortiz, C.S.; Richards, C.; Terry, A.; Parra, J.; Shim, W. Genetic Variability and Geographical Distribution of Mycotoxigenic Fusarium verticillioides Strains Isolated from Maize Fields in Texas. Plant Pathol. J. 2015, 31, 203–211. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Box plot of average FBs content (μg kg−1) produced in vitro on a maize-based medium by strains isolated from maize (M01-25; 6820 μg kg−1) and sorghum (S01-25; 10,244 μg kg−1). Quartile values are reported for M- and S-strains, respectively (Q1: 830 and 250 μg kg−1; Q2: 4050 and 830 μg kg−1; Q3: 9640 and 4100 μg kg−1). One dot and five asterisks represent outlier values calculated on the basis of p < 0.05.
Figure 1.
Box plot of average FBs content (μg kg−1) produced in vitro on a maize-based medium by strains isolated from maize (M01-25; 6820 μg kg−1) and sorghum (S01-25; 10,244 μg kg−1). Quartile values are reported for M- and S-strains, respectively (Q1: 830 and 250 μg kg−1; Q2: 4050 and 830 μg kg−1; Q3: 9640 and 4100 μg kg−1). One dot and five asterisks represent outlier values calculated on the basis of p < 0.05.

Figure 2.
Necrotic areas (mean ± SE) on maize and sorghum leaves challenged with F. verticillioides isolated from maize (M1-25) and sorghum (S1-25). Null interactions for challenged sorghum were twelve and twenty for M- and S-strains, respectively.
Figure 2.
Necrotic areas (mean ± SE) on maize and sorghum leaves challenged with F. verticillioides isolated from maize (M1-25) and sorghum (S1-25). Null interactions for challenged sorghum were twelve and twenty for M- and S-strains, respectively.

Figure 3.
Biplot of principal component analysis computed from fumonisin production (FBs), percentage inhibition of germination of maize (mGi%) and sorghum (sGi%), root inhibition of maize (mRi%) and sorghum (sRi%) and sorghum necrotic area (sNa) of F. verticillioides strains isolated from maize (M-strains) and sorghum (S-strains). Shoot development in both cereals and maize necrotic areas were not correlated to the other considered parameters.
Figure 3.
Biplot of principal component analysis computed from fumonisin production (FBs), percentage inhibition of germination of maize (mGi%) and sorghum (sGi%), root inhibition of maize (mRi%) and sorghum (sRi%) and sorghum necrotic area (sNa) of F. verticillioides strains isolated from maize (M-strains) and sorghum (S-strains). Shoot development in both cereals and maize necrotic areas were not correlated to the other considered parameters.

Figure 4.
UPGMA clusterization based on the Dice similarity index calculated for M- and S-strains on the basis of genetic profile defined by five microsatellite markers. Numbers on the branches show bootstrap support greater than 30%.
Figure 4.
UPGMA clusterization based on the Dice similarity index calculated for M- and S-strains on the basis of genetic profile defined by five microsatellite markers. Numbers on the branches show bootstrap support greater than 30%.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Locus name, repeated motif and primer sequence used to amplify five polymorphic microsatellite loci in Fusarium verticillioides.
Table 1.
Locus name, repeated motif and primer sequence used to amplify five polymorphic microsatellite loci in Fusarium verticillioides.
| Locus Name | Repeated Motif | Primer Name and Sequence (5′–3′) |
|---|---|---|
| Fv-98 | ATCC | Fv-98F AAACAAGATGCGGTCCATTC |
| Fv-98R GGATCGGAGGAGAATCAACA | ||
| Fv-114 | GTCT | Fv-114F CGAATGCCTTGATCTGCTTC |
| Fv-114R GAGAATCCTGTTTGCGTGGT | ||
| Fv-120 | TTG | Fv-120F GTAGCGCGGTAAGAAGATGC |
| Fv-120R AGTCGAAGCCCAACTGAAGA | ||
| Fv-312 | CAGA | Fv-312F TTTCCGAATTCCTGGATCTG |
| Fv-312R GACGCAGTTTGCACAAGGTA | ||
| Fv-338 | AGCAG | Fv-338F TAGACCAGGCAGACGAGACA |
| Fv-338R TGTGAGTGGGTGAGAGTGGA |
Table 2.
Fusarium species from maize and sorghum samples and total fumonisin production by F. verticillioides strains.
Table 2.
Fusarium species from maize and sorghum samples and total fumonisin production by F. verticillioides strains.
| F. verticillioides | F. proliferatum | F. oxysporum | Other Fusaria * | FBs Content | |||||
|---|---|---|---|---|---|---|---|---|---|
| CFU g−1 | % | CFU g−1 | % | CFU g−1 | % | CFU g−1 | % | μg kg−1 | |
| Maize | 9757 ± 3063 | 88.8 | 461 ± 145 | 4.2 | 654 ± 205 | 6.0 | 110 ± 18 | 1.0 | 4050 ± 304 |
| Sorghum | 388 ± 68 | 58.8 | 186 ± 102 | 28.2 | 20 ± 18 | 3.0 | 65 ± 19 | 9.9 | <200 |
Data represent fungal colonization of kernels (CFU g−1) and the percentage of each species with respect to the total Fusarium spp. colonization. Fungal colonization value is the average ± standard error (SE) of three samples. * Indicates Fusarium species other than F. verticillioides, F. proliferatum and F. oxysporum.
Table 3.
Mating type determination and in vitro FBs production (μg kg−1) on maize substrate.
| Maize | Sorghum | |||
|---|---|---|---|---|
| Strain | FBs μg kg−1 | Strain | FBs μg kg−1 | |
| MAT-1 | M06 | 20,300 | S04 | 24,800 |
| M17 | 13,250 | S09 | 21,900 | |
| M22 | 11,170 | S25 | 16,400 | |
| M14 | 7700 | S02 | 2490 | |
| M15 | 6880 | S08 | 1350 | |
| M04 | 6430 | S07 | 1340 | |
| M02 | 2950 | S05 | 830 | |
| M09 | 2380 | S12 | 310 | |
| M18 | 1820 | S10 | 300 | |
| M11 | 830 | S21 | 270 | |
| M13 | 700 | S23 | 240 | |
| M12 | 570 | S03 | 230 | |
| M05 | 240 | S22 | 230 | |
| M01 | 220 | S24 | 230 | |
| M25 | 200 | S11 | 200 | |
| - | - | S20 | 200 | |
| MAT-2 | M19 | 29,600 | S16 | 106,300 |
| M24 | 19,400 | S06 | 61,200 | |
| M08 | 16,780 | S13 | 7950 | |
| M23 | 9640 | S17 | 4100 | |
| M16 | 5570 | S15 | 2180 | |
| M21 | 5140 | S01 | 1930 | |
| M07 | 4050 | S14 | 590 | |
| M10 | 3100 | S18 | 290 | |
| M03 | 850 | S19 | 250 | |
| M20 | 740 | - | - | |
Table 4.
Microsatellite polymorphism for Fusarium verticillioides isolated from maize and sorghum.
| Microsatellite Marker | Strains | Polymorphic Bands/Tot | H’ Index | J’ Index | BC Dissimilarity |
|---|---|---|---|---|---|
| Fv-98 | M1-25 | 3/7 | 1.040 | 0.946 | 0.50 |
| S1-25 | 6/7 | 1.223 | 0.679 | ||
| Fv-114 | M1-25 | 5/9 | 1.332 | 0.828 | 0.84 |
| S1-25 | 6/9 | 1.555 | 0.868 | ||
| Fv-120 | M1-25 | 9/11 | 2.027 | 0.926 | 0.56 |
| S1-25 | 6/11 | 1.658 | 0.925 | ||
| Fv-312 | M1-25 | 6/10 | 1.432 | 0.799 | 0.68 |
| S1-25 | 9/10 | 1.921 | 0.875 | ||
| Fv-338 | M1-25 | 6/12 | 1.457 | 0.813 | 0.68 |
| S1-25 | 11/12 | 2.123 | 0.885 |
Table shows for each marker the number/total of polymorphic bands, Shannon’s index (H’ index), Evenness (J’ index) and Bray Curtis (BC) dissimilarity calculated on the basis of microsatellite markers for M- and S-strains.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ferrigo, D.; Mondin, M.; Raiola, A. Pathogenic and Genetic Characterization of Fusarium verticillioides Strains Collected from Maize and Sorghum Kernels. Agriculture 2023, 13, 105. https://doi.org/10.3390/agriculture13010105
AMA Style
Ferrigo D, Mondin M, Raiola A. Pathogenic and Genetic Characterization of Fusarium verticillioides Strains Collected from Maize and Sorghum Kernels. Agriculture. 2023; 13(1):105. https://doi.org/10.3390/agriculture13010105
Chicago/Turabian StyleFerrigo, Davide, Massimiliano Mondin, and Alessandro Raiola. 2023. "Pathogenic and Genetic Characterization of Fusarium verticillioides Strains Collected from Maize and Sorghum Kernels" Agriculture 13, no. 1: 105. https://doi.org/10.3390/agriculture13010105
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.