
Evaluation of the Egg Predator Blattisocius tarsalis (Mesostigmata: Blattisociidae) for the Biological Control of the Potato Tuber Moth Tecia solanivora under Storage Conditions

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Materials and Experimental Conditions
2.2. Prey Acceptance Test and Evaluation of the Predatory Potential at Different Temperatures
2.3. Microcosm
2.4. Host Choice: Tecia Solanivora (Tcs)-Phthorimaea Operculella (Ptm)
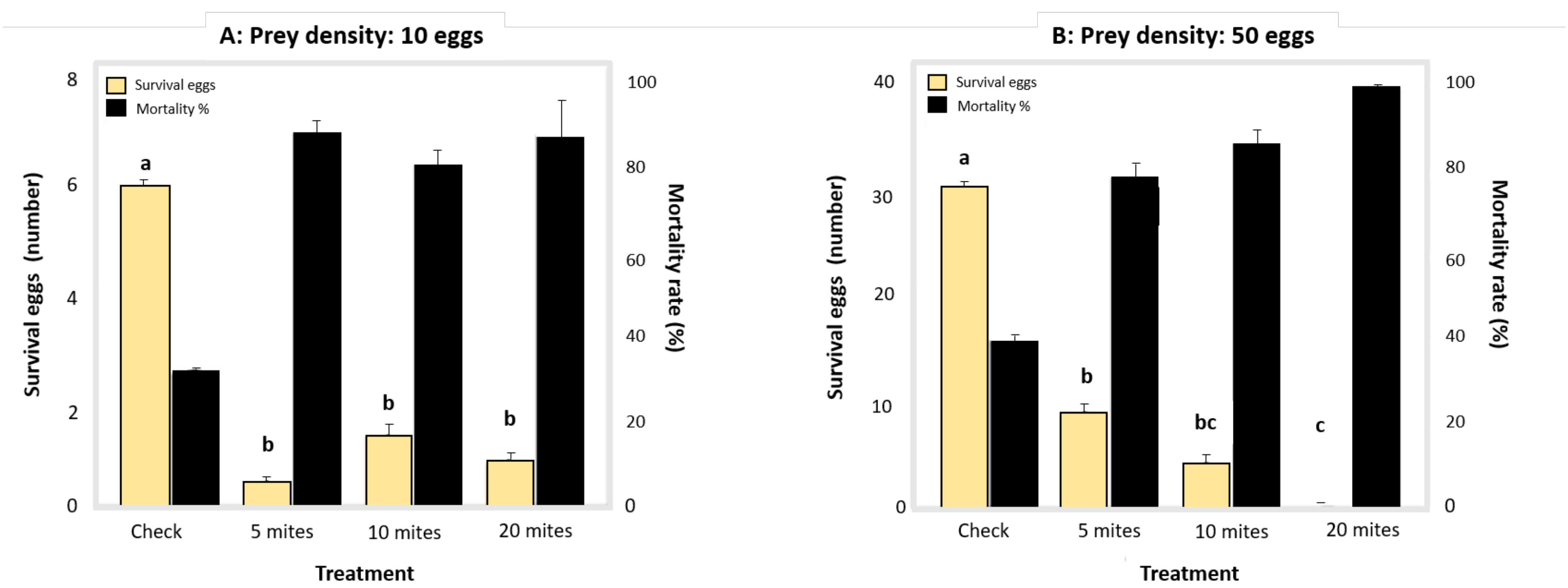
2.5. Functional Response: Predatory Behaviour at Different Prey Densities
- Type I:
- Type II:
- Type III:
3. Results
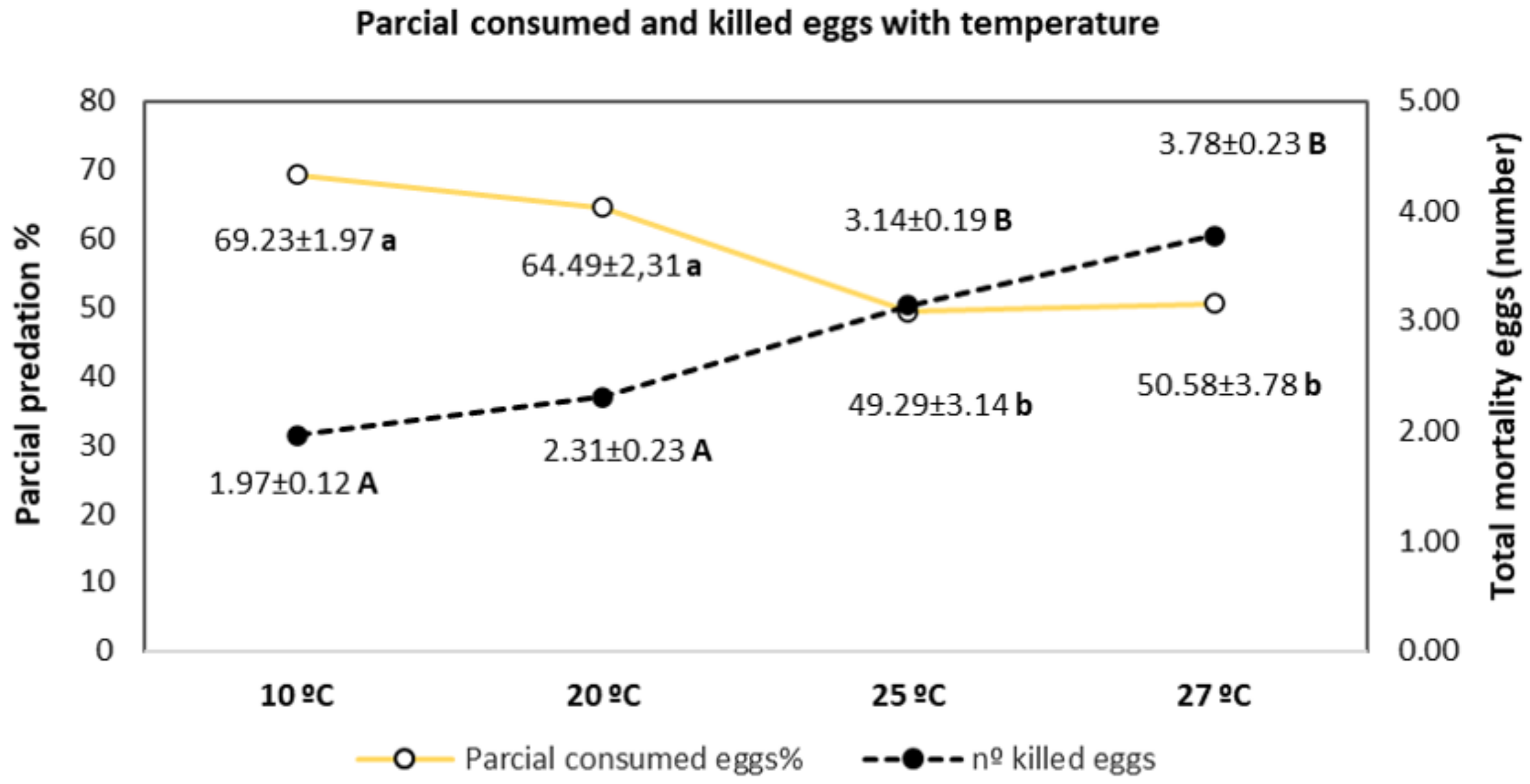
3.1. Evaluation of the Predatory Potential at Different Temperatures
3.2. Microcosm
3.3. Host Choice
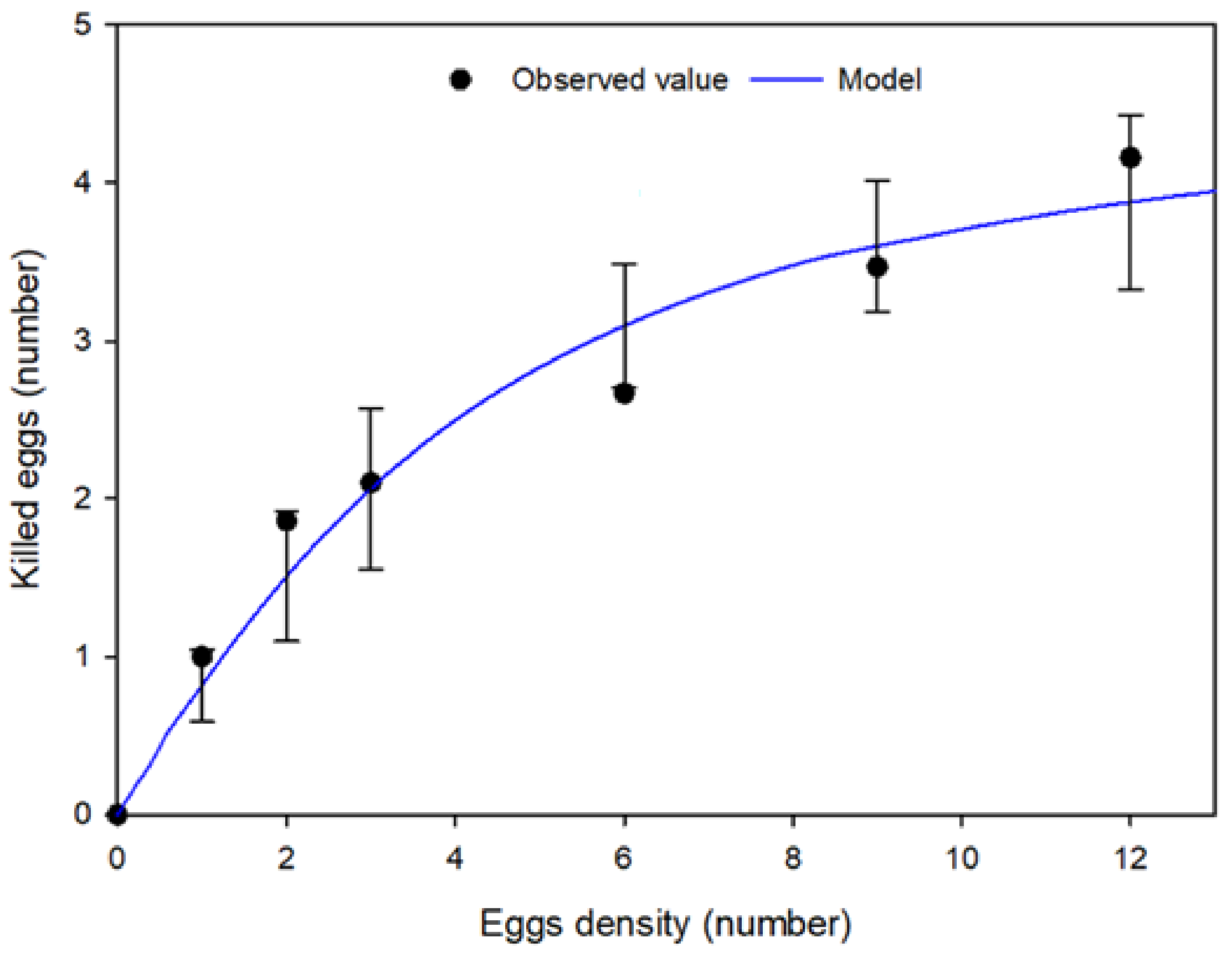
3.4. Functional Response: Predatory Behaviour at Different Prey Densities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sánchez-Aguirre, R.; Palacios, M.L. Eficacia del parasitismo de Copidosoma koehleri en el complejo polilla de la papa. Rev. Peru Entomol. 1995, 38, 59–62. [Google Scholar]
- Carrillo, D.; Torrado-Leon, E. Tecia solanivora Povolny (Lepidoptera: Gelechiidae), an invasive pest of potatoes Solanum tuberosum L. in the Northern Andes. In Potencial Invasive Pest of Agricultural Crops; Peña, J.E., Ed.; CABI: Boston, MA, USA, 2013; pp. 126–136. [Google Scholar] [CrossRef]
- Villanueva, D.F.; Saldamando, C.I. Tecia solanivora, Povolny (Lepidoptera: Gelechiidae): Una revisión sobre su origen, dispersión y estrategias de control biológico. Ing. Cienc. 2013, 18, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Arias, J.; Jaramillo, J.A.; Arévalo, E.; Muñoz, L.; Rocha, N.R. Evaluación de la incidencia y severidad del daño de la polilla gigante de la papa Tecia solanivora en el departamento de Antioquia. Ing. Cienc. Available online: https://repository.agrosavia.co/ (accessed on 2 December 2021).
- Puillandre, N.; Dupas, S.; Dangles, O. Genetic bottleneck in invasive species: The potato tuber moth adds to the list. Biol. Invasions 2008, 10, 319–333. [Google Scholar] [CrossRef]
- Corral, S.; Romero, M.D.; Pereira, A. Assessing the complexity of the spreading processes of agricultural pests: The case of the Guatemalan potato moth in Tenerife. Land Use Policy 2017, 69, 338–348. [Google Scholar] [CrossRef]
- Gallegos, P.; Asaquibay, C.; Cahmorro, F.; Rodríguez, P.; Williams, R. Asolación de los tubérculos de semilla de papa como método de control para la polilla, Tecia solanivora: Una tecnología tradicional para la solución a un nuevo problema. Boletín INIAP 2005, 260. Available online: http://repositorio.iniap.gob.ec/handle/41000/2613 (accessed on 26 November 2021).
- Trujillo, E.; Perera, S. Estudio comparativo de dos feromonas sexuales y de dos tipos de trampas para la captura de adultos de la polilla guatemalteca de la papa (Tecia solanivora). Serv. Técnico Agric. Desarro Rural Cabil. Insul. Tenerife 2011. Available online: https://www.agrocabildo.org/ (accessed on 26 November 2021).
- Lobo, M.G.; González-García, C.; Cabrera, R.; Ríos, D. Development of a Quarantine Postharvest Treatment against Guatemalan Potato Moth (Tecia solanivora Povolny). Agriculture 2021, 11, 801. [Google Scholar] [CrossRef]
- Carnero, A.; Padilla, A.; Perera, S.; Hernández, E.; Trujillo, E. Pest status of Tecia solanivora (Povolny 1973) (Lepidoptera: Gelechiidae), Guatemalan Potato moth, in the Canary Islands. Rev. Peru Entomol. 2008, 31, 336–339. [Google Scholar]
- Gavara, J.; Suárez-Hernández, E.; Piedra-Buena, A. Gestion Integrada de La polilla guatemalteca de la papa (Tecia Solanivora). Agrocabildo, in press.
- RD 197/2017, de 3 de Marzo, Por El Que Se Establece el Programa Nacional de Control y Erradicación de Tecia (Scrobipalpopsis) solanivora (Povolny). Boletín Oficial del Estado (España). Available online: https://www.boe.es/buscar/doc.php?id=BOE-A-2017-2312 (accessed on 2 December 2021).
- Perera, S.; Cubas, F.; Padilla, Á.; Carnero, A.; Hernández, J.; Asensio, M. Ensayo de Eficacia en el Control de la Polilla Guatemalteca de la Papa (Tecia solanivora) Mediante la AplicacióN en Campo de NemáTodos Entomopató Genos. Serv. Técnico Agric. Desarro Rural Cabil. Insul. Tenerife 2009. Available online: https://www.icia.es/ (accessed on 30 November 2021).
- Torres, F. Biología y Manejo Integrado de la Polilla Centroamericana de la Papa Tecia solanivora en Venezuela. Universidad Nacional Experimental del Táchira, Venezuela. Available online: http://www.sidalc.net (accessed on 2 December 2021).
- Niño, L. Revisión sobre la polilla de la papa Tecia solanivora en Centro y Suramérica. Supl. Rev. Latinoam. Papa 2004. Available online: http://www.sidalc.net (accessed on 30 November 2021).
- Herrera, F. La Polilla Guatemalteca de La Papa. Biología, Comportamiento y Practicas de Manejo Integrado. 1998. Available online: https://es.scribd.com/document/22245296/La-polilla-guatemalteca-de-la-papa-Biologia-comportamiento-y-practicas-de-manejo-integrado (accessed on 3 December 2021).
- Schaub, B.; Carhuapoma, P.; Kroschel, J. Guatemalan potato tuber moth, Tecia solanivora (Povolny 1973). In Pest Distribution and Risk Atlas for Africa; 2021; pp. 24–38. Available online: https://cipotato.org/riskatlasforafrica/tecia-solanivora/ (accessed on 10 June 2022).
- Ministerio de Agricultura Pesca y Alimentación. Available online: https://www.mapa.gob.es/es/agricultura/temas/sanidad-vegetal/productosfitosanitarios/registro/menu.asp (accessed on 3 December 2021).
- Guzmán, C. Cambio ClimáTico y Control Biológico de Plagas: Efecto de las condiciones abióticas en las Interacciones entre enemigos naturales presentes en el agro-ecosistema del aguacate. Ph.D. Thesis, Málaga University, Málaga, Spain, 2013. [Google Scholar]
- Ferragut, F.; Garcia-Marí, F.; Costa-Comelles, J.; Laborda, R. Influence of food and temperature on development and oviposition of Euseius stipulatus and Typhlodromus phialatus (Acari: Phytoseiidae). Exp Appl Acarol. 1987, 3, 317–329. [Google Scholar] [CrossRef]
- Nielsen, P.S. Developmental time of Blattisocius tarsalis (Acari: Ascidae) at different temperatures. Exp Appl Acarol. 2001, 25, 605–608. [Google Scholar] [CrossRef]
- Nielsen, P.S. The impact of temperature on activity and consumption rate of moth eggs by Blattisocius tarsalis (Acari: Ascidae). Exp Appl Acarol. 1999, 23, 149–157. [Google Scholar] [CrossRef]
- Pinhero, R.G.; Coffin, R.; Yada, R.Y. Post-Harvest Storage of Potatoes. In Advances in Potato Chemistry and Technology; Singh, J., Kaur, L., Eds.; Elsevier: London, UK, 2009; pp. 338–370. [Google Scholar]
- Benítez, K.; Duarte, A.d.P. Prospección de patógenos con potencial para el control biològico de la polilla guatemalteca de la papa Tecia solanivora, FAM Gelechiidae. Bachelor Thesis, Santo Tomás University, Bogotá, Colombia, 2016. Available online: https://repository.usta.edu.co/handle/11634/2594?show=full (accessed on 22 June 2022).
- Martinelli, C. Resultados de investigación acerca de la biología de Tecia solanivora Povolny. Internal report. Servicio Técnico agricultura y Desarro Rural Cabildo Insular Tenerife, 2004. [Google Scholar]
- Osorio, P.; Espitia, E.; Luque, E. Reconocimiento de enemigos naturales de Tecia solanivora (Lepidoptera:Gelechiidae) en localidades productoras de papa en Colombia. Rev. Colomb. Entomol. 2001, 27, 177–185. [Google Scholar] [CrossRef]
- Piedra-Buena, A.; Gavara, J.; Bastin, S.; Martín-Toledo, T.; Jiménez Rodríguez, T.; Hernández-Suárez, E.; Cabello, T. Resultados de las prospecciones para la búsqueda de nuevos enemigos naturales de las polillas de la papa en Canarias. In IV Taller Internacional de La Polilla Guatemalteca, Tecia solanivora; Agrocabildo, La Orotava: Canary Islands, Spain, 2019; pp. 81–89. Available online: https://www.agrocabildo.org/ (accessed on 8 December 2021).
- Villamil, J.E.; Martínez, J.W. Evaluación de aislamientos nativos de Beauveria spp. sobre Tecia solanivora (Lepidoptera: Gelechiidae) in vitro. Rev. Cienc. Agríc. 2013, 31, 92. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Arias, A.C.; López Pazos, S.A.; Chaparro-Giraldo, A. Actividad biológica de Bacillus thuringiensis sobre la polilla guatemalteca de la papa, Tecia solanivora Povolny (Lepidoptera: Gelechiidae). Rev. Mutis. 2013, 3, 31–42. [Google Scholar] [CrossRef]
- Villamizar, L.; Espinel, C.; Grijalba, E. Reconocimiento, selección y evaluación de aislamientos nativos de virus de la granulosis para el control biológico de la polilla guatemalteca de la papa. Bol. Téc. CORPOICA 2006. Available online: https://www.agrosavia.co/biblioteca (accessed on 8 December 2021).
- Gómez-Bonilla, Y.; López-Ferber, M.; Caballero, P.; Murillo, R.; Muñoz, D. Granulovirus formulations efficiently protect stored and field potatoes from Phthorimaea operculella and Tecia solanivora in Costa Rica. BioControl 2013, 58, 215–224. [Google Scholar] [CrossRef]
- Rubio, S.A.; Vargas, B.I.; López-Ávila, A. Evaluation of the efficiency of Trichogramma lopezandinensis (Hymenoptera: Irichogrammatidae) to control Tecia solanivora (Lepidoptera:) in storage potato. Rev. Colomb. Entomol. 2004, 30, 107–114. [Google Scholar] [CrossRef]
- Gallego, J.R.; Gamez, M.; Cabello, T. Potential of the Blattisocius mali Mite (Acari: Blattisociidae) as a Biological Control Agent of Potato Tubermoth (Lepidoptera: Gelechiidae) in Stored Potatoes. Potato Res. 2020, 63, 241–251. [Google Scholar] [CrossRef]
- Gallego, J.R.; Caicedo, O.; Gamez, M.; Hernandez, J.; Cabello, T. Selection of predatory mites for the biological control of potato tuber moth in stored potatoes. Insects 2020, 11, 196. [Google Scholar] [CrossRef] [Green Version]
- Gavara, J.; Piedra-Buena, A.; Hernandez-Suarez, E.; Gamez, M.; Cabello, T.; Gallego, J.R. Potential for the Postharvest Biological Control of Phthorimaea operculella (Lepidoptera, Gelechiidae) by Blattisocius tarsalis (Mesostigmata, Blattisociidae). Agronomy 2021, 11, 288. [Google Scholar] [CrossRef]
- Haines, C.P. Laboratory studies on the role of an egg predator, Blattisocius tarsalis (Berlese) (Acari: Ascidae), in relation to the natural control of Ephestia Cautella (Walker) (Lepidoptera: Pyralidae) in Warehouses. Bull. Entomol. Res. 1987, 71, 557–574. [Google Scholar] [CrossRef]
- Healy, M.J.R. A table of Abbott’s correction for natural mortality. Ann. Appl. Biol. 1952, 39, 211–212. [Google Scholar] [CrossRef]
- Manly, B.F.J.; Miller, P.; Cook, L.M. Analysis of a Selective Predation Experiment. Am. Nat. 1972, 106, 719–736. [Google Scholar] [CrossRef]
- Chesson, J. The Estimation and Analysis of Preference and Its Relatioship to Foraging Models. Ecology 1983, 64, 1297–1304. [Google Scholar] [CrossRef]
- Juliano, S.A. Nonlinear Curve Fitting: Predation and Functional Response Curves. In Design and Analysis of Ecological Experiments; Scheiner, S., Gurevitch, J., Eds.; Oxford University Press: Oxford, UK, 2001; pp. 178–196. [Google Scholar]
- Hassell, M.P. Arthropod Predator-Prey System; Princeton University Press: Princeton, NJ, USA, 1978. [Google Scholar]
- Cabello, T.; Gamez, M. An improvement of the holling type III functional response in entomophagous species model. J. Biol. Syst. 2007, 5, 515–524. [Google Scholar] [CrossRef]
- Nielsen, P.S. The use of Blattisocius (Acari: Ascidae) for biological control in flour mils. In Proceedings of the 7th International Working Conference on Stored-Product Protection, Beijing, China, 14–19 October 1998; Sichuan Publishing House of Science and Technology: Chengdu, China, 1999; ISBN 7536440987. [Google Scholar]
- Rall, B.C.; Vucic-Pestic, O.; Ehnes, R.B.; Emmerson, M.; Brose, U. Temperature, predator-prey interaction strength and population stability. Glob. Change Biol. 2010, 16, 2145–2157. [Google Scholar] [CrossRef]
- Kroschel, J.; Schaub, B. Biology and Ecology of Potato Tuber Moths as Major Pests of Potato. In Insect Pests of Potato: Global Perspectives on Biology and Management; Giordanengo, P.V.C., Alyokhin, A., Eds.; Elsevier: Waltham, MA, USA, 2013; pp. 165–192. [Google Scholar]
- Richards, O.W.; Thomson, W.S. A contribution to the study of the genera Ephestia, gn. (including strymax, dyar), and Plodia, gn. (lepidoptera, phycitidae), with notes on parasites of the larvae. In Ecological Entomology; Gilbert, F., Roitberg, B., Cotter, S.C., Mennéndez, R., Eds.; Empire Marketing Board: London, UK, 2022; pp. 137–168. [Google Scholar] [CrossRef]
- Andreadis, S.; Spanoudis, C.; Zakka, G.; Aslanidou, B.; Noukari, S.; Savopoulou-Soultani, M. Effect of temperature on rate of development, survival and adult longevity of Phthorimaea operculella (Lepidoptera: Gelechiidae). Eur. J. Entomol. 2017, 114, 35–41. [Google Scholar] [CrossRef]
- Riudavets, J.; Quero, R. Prey preference of the predatory mite Blattisocius tarsalis (Acari: Ascidae). In Advances in Stored Product Protection; Credland, P.F., Armitage, D.M., Bell, C.H., Cogan, P.M., Highley, E., Eds.; CABI Publishing: Wallingford, UK, 2002; pp. 297–299. [Google Scholar]
- Mills, N.J. Satiation and the functional response: A test of a new model. Ecol. Entomol. 1982, 7, 305–315. [Google Scholar] [CrossRef]
- Riudavets, J.; Maya, M. Monserrat M. Predation by Blattisocius tarsalis (Acari: Ascidae) on stored product pests. IOBC WPRS Bull. 2002, 25. Available online: http://iobc-wprs.org (accessed on 15 March 2022).
- Arbogast, R.T.; Lecato, G.L.; Van Byrd, R. External Morphology of Some Eggs of Stored-Product Moths (Lepidoptera). J. Insect Morphol. Embryol. 1980, 9, 165–167. [Google Scholar] [CrossRef]
- Kučerová, Z.; Stejskal, V. External egg morphology of two stored-product anobiids, Stegobium paniceum and Lasioderma serricorne (Coleoptera: Anobiidae). J. Stored Prod. Res. 2010, 46, 202–205. [Google Scholar] [CrossRef]
- Martínez, C.; Soria, C.A.; León V., R. Morfología comparativa del corión del huevo de Symmetrischema tangolias (Gyen 1913) y Tecia solanivora (Povolny 1973), polillas del cultivo de Solanum tuberosum ). Rev. Ecuat. Med. Cienc. Biol. 2019, 40. [Google Scholar] [CrossRef]
- Díaz, F. Evaluación del efecto del huevo huésped de cría en la biología del parasitoide Trichogramma achaeae (Hym.: Trichogrammatidae). Degree Final Project, University of Almería, Almería, June 2013. Available online: http://repositorio.ual.es/ (accessed on 26 March 2022).
- Blum, M.; Hilker, M. Chemical protection of insect eggs. Chemoecology of Insect Eggs and Egg Deposition; Hilker, M., Meiners, T., Eds.; Blackwell Publishing: Malden, MA, USA, 2002; pp. 61–90. [Google Scholar]
- Thomas, H.Q.; Zalom, F.G.; Nicola, N.L. Laboratory studies of Blattisocius keegani (Fox) (Acari: Ascidae) reared on eggs of navel orangeworm: Potential for biological control. Bull. Entomol. Res. 2011, 101, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Ghoneim, K. Parasitic insects and mites as potential biocontrol agents for a devastative pest of tomato, Tuta absoluta Meyrick (Lepidoptera: Gelechiidae) in the world: A review. J. Adv. Res. 2010, 2, 81–115. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature °C | Surviving Eggs (nº ± SE) | Mortality (% ±SE) | Efficacy % | ||
|---|---|---|---|---|---|
| Treatment | Check | Treatment | Check | ||
| 10 | 3.15 ± 0.35 a | 4.30 ± 0.39 b | 40.00 ± 2.34 | 13.10 ± 1.80 | 33.52 ± 2.44 |
| 20 | 2.87 ± 0.31 a | 4.53 ± 0.18 b | 45.33 ± 4.06 | 3.45 ± 1.43 | 40.14 ± 3.54 |
| 25 | 1.90 ± 0.25 a | 4.62 ± 0.40 b | 64.67 ± 4.15 | 7.33 ± 1.83 | 59.26 ± 4.59 |
| 27 | 1.17 ± 0.20 a | 4.4 ± 0.38 b | 76.67 ± 4.40 | 12.41 ± 2.51 | 75.19 ± 4.64 |
| Host | Eggs Mortality | Manly Index |
|---|---|---|
| T. solanivora | 1.77 ± 0.43 a | 0.57 ± 0.07 a |
| P. operculella | 1.46 ± 0.23 a | 0.43 ± 0.07 a |
| Parameter | Value | SE | 95% Confidence Level | F.R. Type | |
|---|---|---|---|---|---|
| (Intercept) | 1.1707 | 0.0598 | 0.9805 | 1.3609 | II |
| (Linear) | −0.1661 | 0.025 | −0.0865 | −0.0865 | II |
| Type | Fit Curve Parameters (±SE) | Statistical Parameters | ||||
|---|---|---|---|---|---|---|
| d.f. | ||||||
| I | 0.4999 | - | - | 6 | 0.7237 | −2.2467 |
| II | 2.0502 | 0.2091 | - | 5 | 0.9457 | −6.1924 |
| III | - | 0.2934 | 0.7833 | 5 | 0.8358 | 1.8271 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gavara, J.; Cabello, T.; Gallego, J.R.; Hernández-Suarez, E.; Piedra-Buena Díaz, A. Evaluation of the Egg Predator Blattisocius tarsalis (Mesostigmata: Blattisociidae) for the Biological Control of the Potato Tuber Moth Tecia solanivora under Storage Conditions. Agriculture 2022, 12, 920. https://doi.org/10.3390/agriculture12070920
Gavara J, Cabello T, Gallego JR, Hernández-Suarez E, Piedra-Buena Díaz A. Evaluation of the Egg Predator Blattisocius tarsalis (Mesostigmata: Blattisociidae) for the Biological Control of the Potato Tuber Moth Tecia solanivora under Storage Conditions. Agriculture. 2022; 12(7):920. https://doi.org/10.3390/agriculture12070920
Chicago/Turabian StyleGavara, Jorge, Tomás Cabello, Juan Ramón Gallego, Estrella Hernández-Suarez, and Ana Piedra-Buena Díaz. 2022. "Evaluation of the Egg Predator Blattisocius tarsalis (Mesostigmata: Blattisociidae) for the Biological Control of the Potato Tuber Moth Tecia solanivora under Storage Conditions" Agriculture 12, no. 7: 920. https://doi.org/10.3390/agriculture12070920