
Non-Structural Carbohydrates, Foliar Nutrients, Yield Components and Oxidative Metabolism in Pecan Trees in Response to Foliar Applications of Growth Regulators

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Orchard Management and Experimental Design
2.2. Sampling of Leaflets, Buds and Nuts
2.3. Non-Structural Carbohydrates (Leaflets and Buds)
2.4. Leaf Mineral Nutrients
2.5. Yield Components (Yield and Nut Quality)
2.6. Oxidative Metabolism (Enzymatic Activity and Antioxidant Capacity)
2.7. Statistical Analyses
3. Results
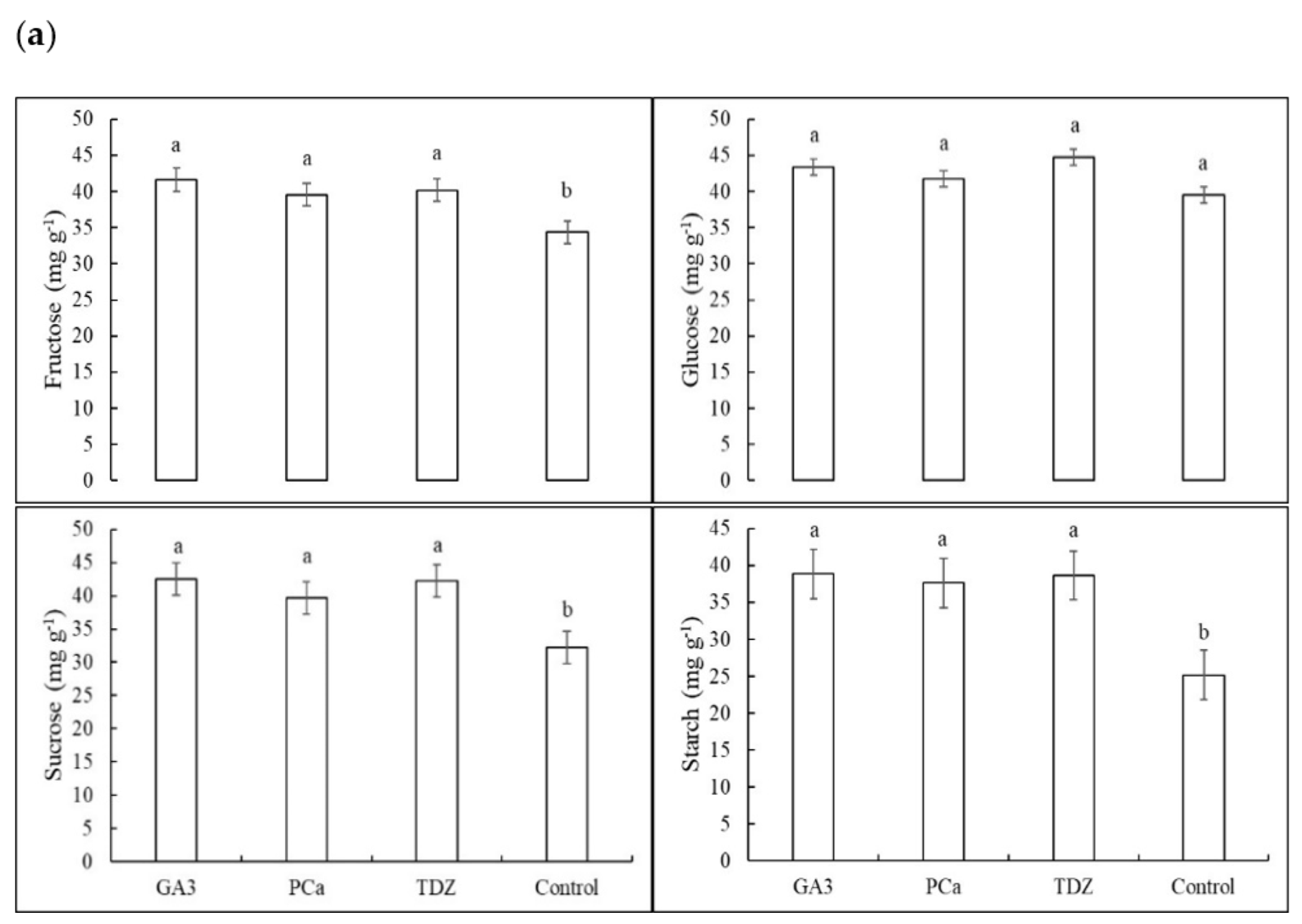
3.1. Non-Structural Carbohydrates in cv. Wichita Pecan Leaflets and Buds
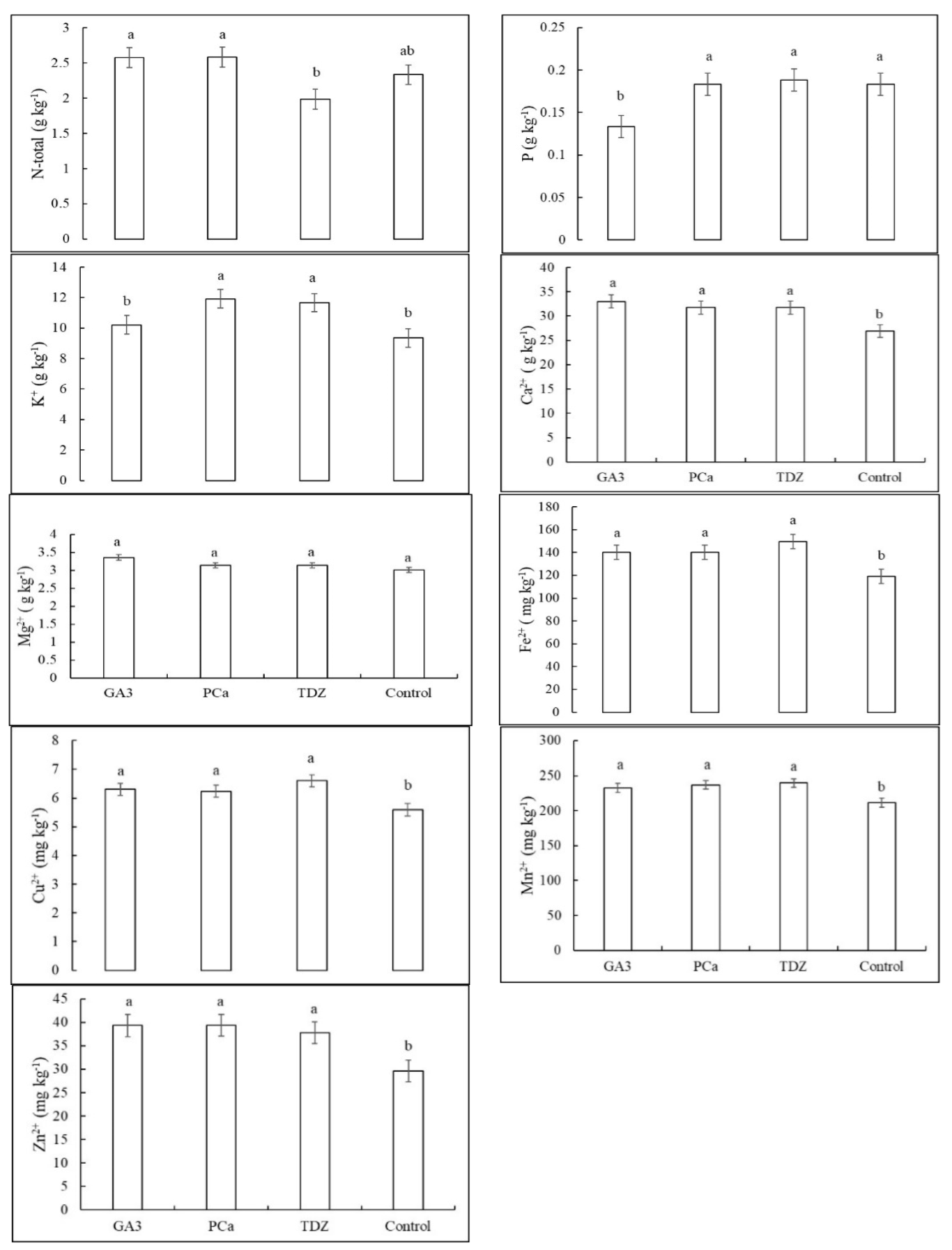
3.2. Concentrations of Nutrients in Pecan Leaflets
3.3. Yield Components (Yield and Nut Quality)
3.4. Oxidative Metabolism and Antioxidant Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Heerema, R.J.; Vanleeuwen, D.; Thompson, M.Y.; Sherman, J.D.; Comeau, M.J.; Walworth, J.L. Soil-application of Zinc-EDTA increases leaf photosynthesis of immature ‘Wichita’ pecan trees. J. Am. Soc. Hortic. Sci. 2017, 142, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.; Singh, S.K.; Mahato, A.K.; Ravishankar, H.; Dubey, A.K.; Singh, N.K. Physiological and molecular basis of alternate bearing in perennial fruit crops. Sci. Hortic. 2019, 243, 214–225. [Google Scholar] [CrossRef]
- Conner, P.J.; Worley, R.E. Alternate bearing intensity of pecan cultivars. HortScience 2000, 35, 1067–1069. [Google Scholar] [CrossRef] [Green Version]
- Abu-Zahra, T.; Al-Dmoor, H. Seasonal changes in endogenous plant hormones and alternate bearing of Nabali olive (Olea europea L.) trees. Asian J. Plant Sci. 2013, 12, 241. [Google Scholar] [CrossRef]
- Noperi-Mosqueda, L.C.; Soto-Parra, J.M.; Sánchez, E.; Navarro-León, E.; Pérez-Leal, R.; Flores-Cordova, M.A.; Salas-Salazar, N.A.; Yáñez-Muñoz, R.M. Yield, quality, alternate bearing and long-term yield index in pecan, as a response to mineral and organic nutrition. Not. Bot. Horti Agrobot. 2020, 48, 342–353. [Google Scholar] [CrossRef] [Green Version]
- Randall, J.J.; Rascon, A.; Heerema, R.J.; Potter, M.T. Molecular mechanisms of pecan flower induction. Acta Hortic. 2015, 1070, 89–99. [Google Scholar] [CrossRef]
- Zea-Hernández, L.O.; Saucedo-Veloz, C.; Cruz-Huerta, N.; Ramírez-Guzmán, M.A.; Robles-González, M.M. Evaluation of post-harvest applications of gibberellic acid on the quality and shelf life of three varieties of Mexican lime. Rev. Chapingo Ser. Hortic. 2016, 22, 17–26. [Google Scholar] [CrossRef]
- Wood, B.W. Influence of plant bioregulators on pecan flowering and implications for regulation of pistillate flower initiation. HortScience 2011, 46, 870–877. [Google Scholar] [CrossRef] [Green Version]
- Ionescu, I.A.; Møller, B.L.; Sánchez-Pérez, R. Chemical control of flowering time. J. Exp. Bot. 2017, 68, 369–382. [Google Scholar] [CrossRef]
- Dahal, K.C.; Bhattarai, S.P.; Midmore, D.J.; Oag, D.R.; Walsh, K.B. Temporal yield variability in subtropical table grape production. Sci. Hortic. 2019, 246, 951–956. [Google Scholar] [CrossRef]
- Orozco-Meléndez, L.R.; Hernández-Rodríguez, O.A.; Cruz-Alvarez, O.; Benavidez-Mendoza, A.; Calderón-Jurado, M.; Ojeda-Barrios, D.L. Does the application of growth bioregulators improve the foliar concentration of nutrients, non-structural carbohydrates and yield in pecan? Ciênc. Agrotecnologia 2021, 45, e004721. [Google Scholar] [CrossRef]
- Orozco-Meléndez, L.R.; Hernández-Rodríguez, O.A.; Cruz-Alvarez, O.; Cano-Medrano, R.; Jacobo-Cuellar, J.L.; Parra-Quezada, R.A.; Jiménez-Castro, J.; Ojeda-Barrios, D.L. Foliar application of some growth bioregulators and their effect on the yield and nut quality in pecan. J. Elem. 2021, 26, 407–417. [Google Scholar] [CrossRef]
- Valenzuela-Núñez, L.M.; Briceño-Contreras, E.A.; Esparza-Rivera, J.R.; Rodríguez-Bautista, G.; Núñez-Colima, J.A. Seasonal changes in soluble sugar concentration in pecan perennial organs (Carya illinoiensis [Wangenh.] Koch). Acta Univ. 2019, 29, e2423. [Google Scholar]
- Pasa, M.S.; Silva, C.P.D.; Carra, B.; Brighenti, A.F.; Souza, A.L.K.D.; Petri, J.L. Thidiazuron (TDZ) increases fruit set and yield of “Hosui” and “Packham”s Triumph’ pear trees. Acad. Brasil. Ciências 2017, 89, 3103–3110. [Google Scholar] [CrossRef] [Green Version]
- Marino, G.; La Mantia, M.; Caruso, T.; Marra, F.P. Seasonal dynamics of photosynthesis and total carbon gain in bearing and nonbearing pistachio (Pistacia vera L.) shoots. Photosynthetica 2018, 56, 932–941. [Google Scholar] [CrossRef]
- Rohla, C.T.; Smith, M.W.; Maness, N.O. Influence of cluster thinning on return bloom, nut quality, and concentrations of potassium, nitrogen, and non-structural carbohydrates. J. Am. Soc. Hortic. Sci. 2007, 132, 158–165. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [Green Version]
- García-Fuentes, J.E.; Herrera-Castellanos, B.F.; Rivas-Martínez, E.N.; Narvaéz-Ortiz, W.A.; Benavidez-Mendoza, A.; Medrano-Macías, J. Outcomes of foliar iodine application on growth, minerals and antioxidants in tomato plants under salt stress. Folia Hortic. 2022; in press. [Google Scholar] [CrossRef]
- Balandrán-Valladares, M.I.; Cruz-Alvarez, O.; Jacobo-Cuellar, J.L.; Hernández-Rodríguez, O.A.; Flores-Córdova, M.A.; Parra-Quezada, R.A.; Sánchez-Chávez, E.; Ojeda-Barrios, D.L. Changes in nutrient concentration and oxidative metabolism in pecan leaflets at different doses of zinc. Plant Soil Environ. 2021, 67, 33–39. [Google Scholar] [CrossRef]
- Sariñana-Aldaco, O.; Sánchez-Chávez, E.; Troyo-Diéguez, E.; Tapia-Vargas, L.M.; Díaz-Pérez, J.C.; Preciado-Rangel, P. Foliar aspersion of salicylic acid improves nutraceutical quality and fruit yield in tomato. Agriculture 2020, 10, 482. [Google Scholar] [CrossRef]
- Sánchez, E.; Soto, J.M.; García, P.C.; López-Lefebre, L.R.; Rivero, R.M.; Ruiz, J.M.; Romero, L. Phenolic compounds and oxidative metabolism in green bean plants under nitrogen toxicity. Funct. Plant Biol. 2000, 27, 973–978. [Google Scholar] [CrossRef]
- Wood, B.W.; Conner, P.J.; Worley, R.E. Insight into alternate bearing of pecan. In Proceedings of the XXVI International Horticultural Congress: Key Processes in the Growth and Cropping of Deciduous Fruit and Nut Trees, Toronto, ON, Canada, 30 April 2004. [Google Scholar]
- Sánchez, E.; Soto, J.M.; Núñez, A.; Ruiz, J.M.; Romero, L. Biosynthesis of non-structural carbohydrates and their distribution in green bean plants (Phaseolus vulgaris L. Cv. Strike): Deficiency vs toxicity of nitrogen. Rev. Fitotec. Mex. 2005, 28, 55–61. [Google Scholar]
- Cruz-Alvarez, O.; Hernández-Rodríguez, A.O.; Jacobo-Cuellar, J.L.; Ávila-Quezada, G.; Morales-Maldonado, E.; Parra-Quezada, R.A.; Robles-Hernández, L.; Ojeda-Barrios, D.L. Nitrogen fertilization in pecan and its effect on leaf nutrient concentration, yield and nut quality. Rev. Chapingo Ser. Hortic. 2020, 26, 163–173. [Google Scholar] [CrossRef]
- Comenuez. Available online: http://www.comenuez.com/info-del-cultivo.html (accessed on 2 November 2021).
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutase occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L.W.T. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Hanusz, Z.; Tarasińska, J. Normalization of the Kolmogorov–Smirnov and Shapiro–Wilk tests of normality. Biom. Lett. 2015, 52, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Nzima, M.D.; Martin, G.C.; Nishijima, C. Seasonal changes in total nonstructural carbohydrates within branches and roots of naturally “Off” and “On” Kerman’ pistachio trees. J. Am. Soc. Hortic. Sci. 1997, 122, 856–862. [Google Scholar] [CrossRef] [Green Version]
- Wood, B.W.; Conner, P.J.; Worley, R.E. Relationship of alternate bearing intensity in pecan to fruit and canopy characteristics. HortScience 2003, 38, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Mirsoleimani, A.; Shahsavar, A.R.; Kholdebarin, B. Seasonal changes of mineral nutrient concentrations of leaves and stems of ‘Kinnow’mandarin trees in relation to alternate bearing. Int. J. Fruit Sci. 2014, 14, 117–132. [Google Scholar] [CrossRef]
- Cruz-Crespo, E.; Can-Chulim, Á.; Bugarín-Montoya, R.; Pineda-Pineda, J.; Flores-Canales, R.; Juárez-López, P.; Alejo-Santiago, G. Concentración nutrimental foliar y crecimiento de chile serrano en función de la solución nutritiva y el sustrato. Rev. Fitotec. Mex. 2014, 37, 289–295. [Google Scholar] [CrossRef]
- Pond, A.P.; Walworth, J.L.; Kilby, M.W.; Gibson, R.D.; Call, R.E.; Nunez, H. Leaf nutrient levels for pecans. HortScience 2006, 41, 1339–1341. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.B.; Treeby, M.; Stephenson, R.A. Fruits, vines and nuts. In Plant Analysis: An Interpretation Manual; Reuter, D.J., Robinson, J.B., Eds.; CSIRO Publishing: Victoria, Australia, 1997; pp. 347–349. [Google Scholar]
- Jones, J.B.; Wolf, B.; Mills, H.A. Plant Analysis Handbook; Micro-Macro Publishing: Athens, Greece, 1991; pp. 154–196. [Google Scholar]
- Conner, P.J. Performance of 19 pecan cultivars and selections in Southern Georgia. HortTechnology 2004, 24, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Grauke, L.J.; Thompson, T.E. Variability in pecan flowering. Fruit Var. J. 1996, 50, 140–150. [Google Scholar]
- Thompson, M.Y.; Randall, J.; Heerema, R.J.; VanLeeuwen, D. Exogenous plant growth regulators show promise for management of alternate bearing in pecan. HortScience 2019, 54, 1204–1207. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Organ | GR (mg L−1) | Fructose 1 | Glucose | Sucrose | Starch | ||||
|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | ||
| Leaflets | GA3 (50) | 40.9 a | 42.4 a | 43.5 a | 46.9 a | 40.4 a | 44.7 a | 36.9 a | 40.7 a |
| PCa (500) | 38.1 a | 40.9 a | 41.6 a | 42.6 a | 37.7 a | 41.7 a | 35.9 a | 39.3 a | |
| TDZ (10) | 38.5 a | 41.8 a | 41.9 a | 46.2 a | 40.4 a | 44.2 a | 37.0 a | 40.3 a | |
| Control | 33.1 a | 35.6 b | 33.6 a | 37.6 b | 25.7 a | 38.7 b | 23.4 a | 28.3 b | |
| Buds | GA3 (50) | 41.7 a | 45.9 a | 40.5 a | 44.3 a | 43.9 a | 46.2 a | 43.6 a | 45.8 a |
| PCa (500) | 40.2 a | 45.1 a | 38.8 a | 41.4 a | 41.1 a | 44.7 a | 41.0 a | 45.5 a | |
| TDZ (10) | 41.73 a | 45.2 a | 38.6 a | 42.6 a | 41.6 a | 44.5 a | 42.5 a | 45.1 a | |
| Control | 30.3 a | 35.9 b | 28.9 a | 33.8 b | 31.9 a | 35.7 a | 31.6 a | 34.9 b | |
| GR 1 | g kg−1 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N-Total | P | K+ | Ca2+ | Mg2+ | ||||||
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | |
| GA3 | 28.4 a | 23.1 a | 1.1 a | 1.5 a | 9.9 a | 10.5 a | 34.1 a | 31.9 a | 3.3 a | 3.5 a |
| PCa | 26.8 a | 24.9 a | 1.8 a | 1.9 a | 12.9 a | 10.9 b | 31.6 a | 31.8 a | 3.1 a | 3.2 a |
| TDZ | 20.7 a | 19.0 a | 1.7 a | 2.0 a | 11.4 a | 11.9 a | 31.2 a | 32.4 a | 3.1 a | 3.1 a |
| Control | 23.8 a | 22.9 a | 1.8 a | 1.7 a | 9.3 a | 9.4 a | 26.9 a | 26.9 a | 2.9 a | 3.1 a |
| GR | mg kg−1 | |||||||||
| Fe2+ | Cu2+ | Mn2+ | Zn2+ | |||||||
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | |||
| GA3 | 142.0 a | 138.3 a | 6.2 a | 6.4 a | 229.7 a | 236.3 a | 40.5 a | 38.1 a | ||
| PCa | 138.0 a | 142.3 | 6.3 a | 6.2 a | 236.3 a | 237.4 a | 41.5 a | 37.3 a | ||
| TDZ | 151.7 a | 147.3 a | 6.8 a | 6.3 a | 244.7 a | 234.3 a | 45.1 a | 30.5 ab | ||
| Control | 123.8 a | 114.3 a | 5.6 a | 5.6 a | 217.0 a | 205.6 a | 31.2 a | 28.2 b | ||
| GR (mg L−1) | Yield (kg tree−1) | Nut Quality | ||||
|---|---|---|---|---|---|---|
| Nut Weight Per kg (g) | (%) Kernel | |||||
| 2017 1 | 2018 | 2017 | 2018 | 2017 | 2018 | |
| GA3 (50) | 14.53 a | 16.7 a | 6.7 a | 6.1 a | 61.8 a | 59.9 a |
| PCa (500) | 11.3 a | 13.1 b | 6.4 a | 5.6 b | 60.7 a | 54.3 b |
| TDZ (10) | 13.5 a | 15.7 b | 7.2 a | 5.9 b | 59.5 a | 53.9 b |
| Control | 6.1 a | 9.9 b | 5.5 a | 5.0 b | 58.6 a | 55.1 b |
| GR (mg L−1) | SOD 1 | H2O2 | CAT | GPx | AC |
|---|---|---|---|---|---|
| GA3 (50) | 1.29 b | 0.34 b | 2.43 b | 4.35 b | 60.00 c |
| PCa (500) | 1.18 c | 0.35 b | 2.31 b | 4.20 b | 71.09 b |
| TDZ (10) | 1.44 ab | 0.31 b | 2.33 b | 3.22 c | 75.15 a |
| Control | 1.60 a | 0.42 a | 3.20 a | 5.46 a | 76.78 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ojeda-Barrios, D.L.; Orozco-Meléndez, L.R.; Cano-Medrano, R.; Sánchez-Chávez, E.; Parra-Quezada, R.Á.; Calderón-Jurado, M.; Jacobo-Cuellar, J.L.; Hernández-Ordoñez, E.; Cruz-Álvarez, O. Non-Structural Carbohydrates, Foliar Nutrients, Yield Components and Oxidative Metabolism in Pecan Trees in Response to Foliar Applications of Growth Regulators. Agriculture 2022, 12, 688. https://doi.org/10.3390/agriculture12050688
Ojeda-Barrios DL, Orozco-Meléndez LR, Cano-Medrano R, Sánchez-Chávez E, Parra-Quezada RÁ, Calderón-Jurado M, Jacobo-Cuellar JL, Hernández-Ordoñez E, Cruz-Álvarez O. Non-Structural Carbohydrates, Foliar Nutrients, Yield Components and Oxidative Metabolism in Pecan Trees in Response to Foliar Applications of Growth Regulators. Agriculture. 2022; 12(5):688. https://doi.org/10.3390/agriculture12050688
Chicago/Turabian StyleOjeda-Barrios, Damaris Leopoldina, Laura Raquel Orozco-Meléndez, Raquel Cano-Medrano, Esteban Sánchez-Chávez, Rafael Ángel Parra-Quezada, Marisela Calderón-Jurado, Juan Luis Jacobo-Cuellar, Elizabeth Hernández-Ordoñez, and Oscar Cruz-Álvarez. 2022. "Non-Structural Carbohydrates, Foliar Nutrients, Yield Components and Oxidative Metabolism in Pecan Trees in Response to Foliar Applications of Growth Regulators" Agriculture 12, no. 5: 688. https://doi.org/10.3390/agriculture12050688