
Arbuscular Mycorrhizal Fungi Isolated from Highly Saline “Sabkha Habitat” Soil Alleviated the NaCl-Induced Stress and Improved Lasiurus scindicus Henr. Growth

 ,
, 
 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
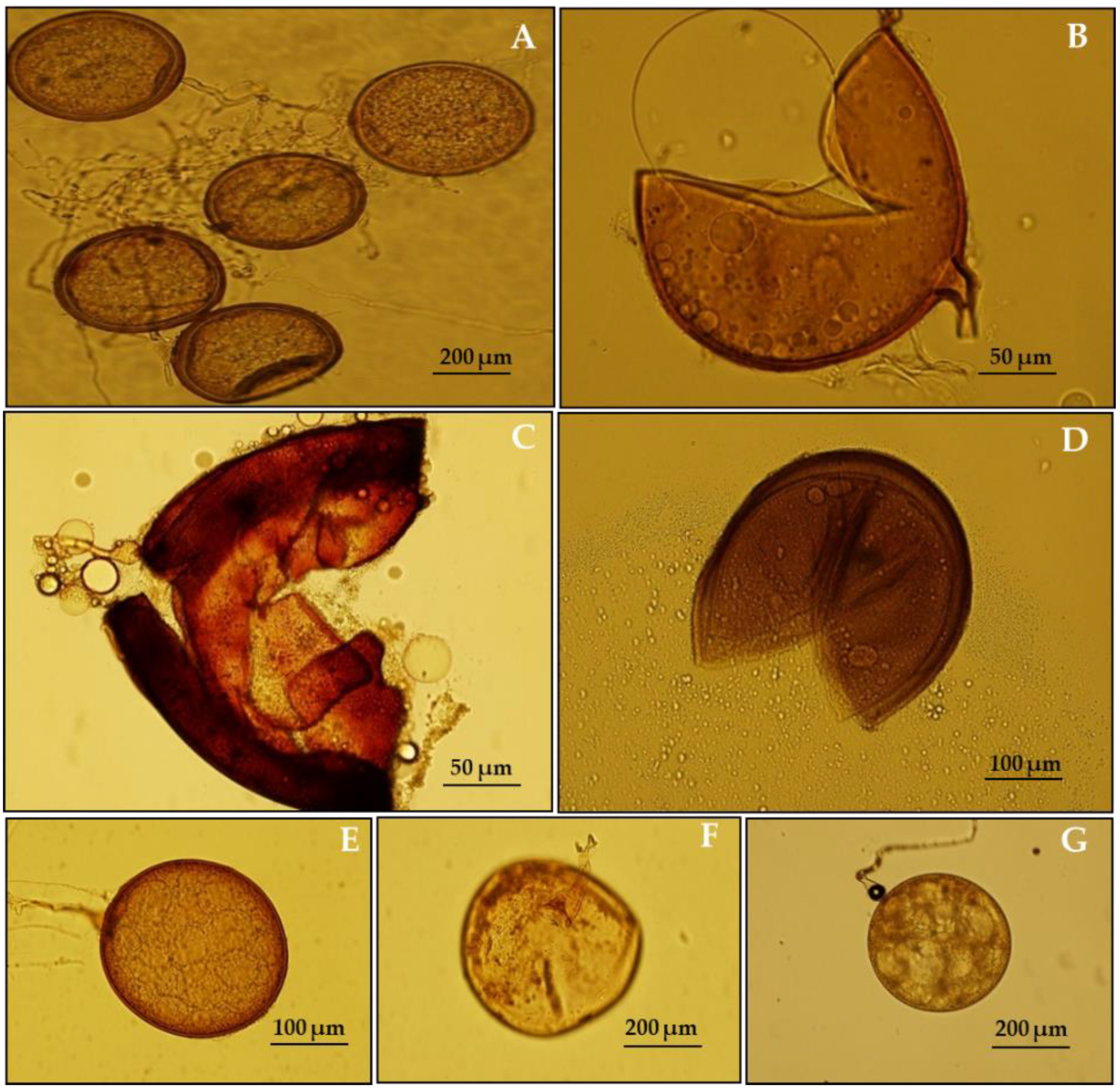
2.1. Sampling, AMF Inoculum Production, and Identification
2.2. Collection of L. scindicus Seeds and Preparation of Soil Mixture
2.3. Experimental Design
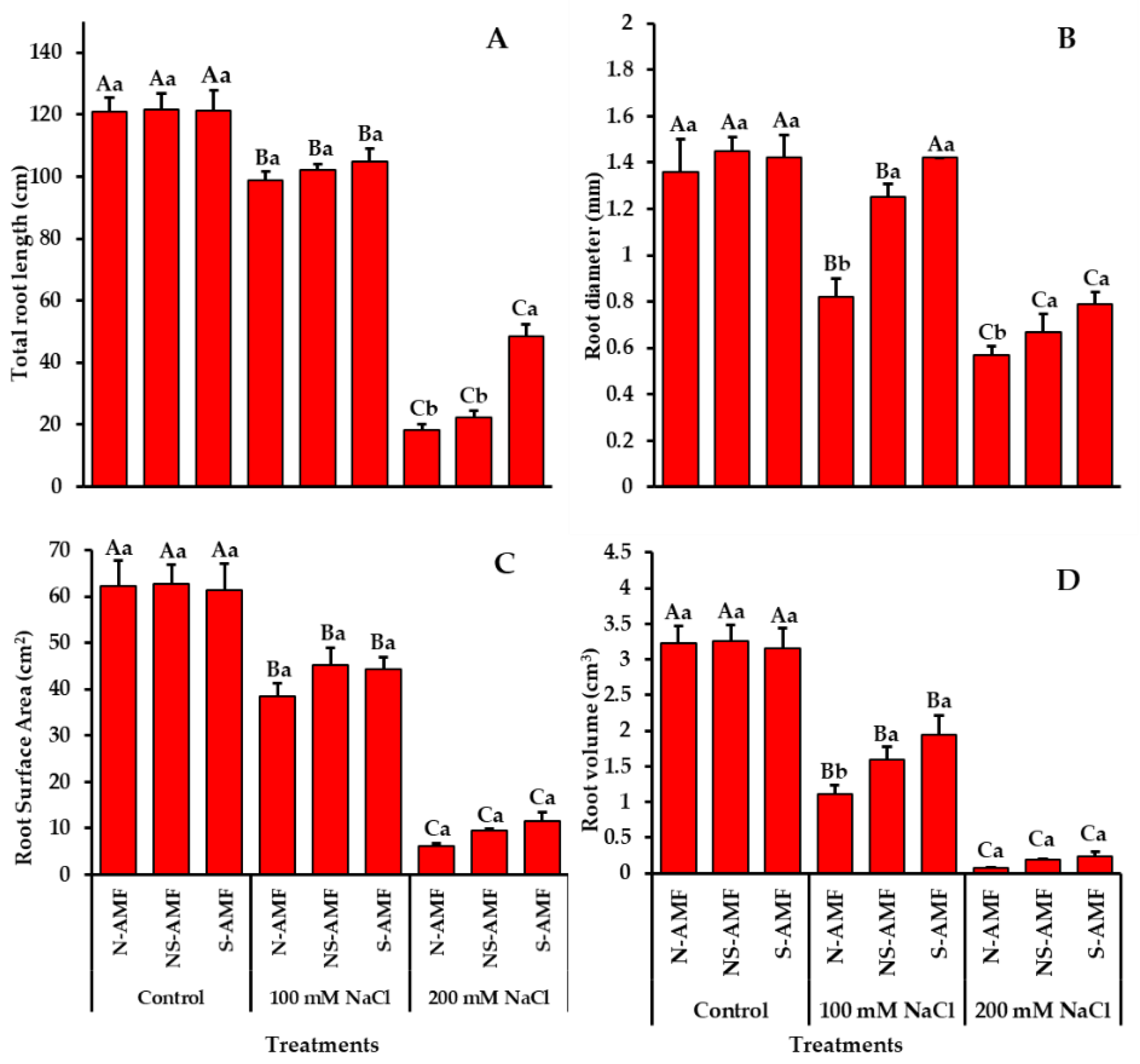
2.4. Growth Parameters
2.5. Symbiotic Development and Spore Count
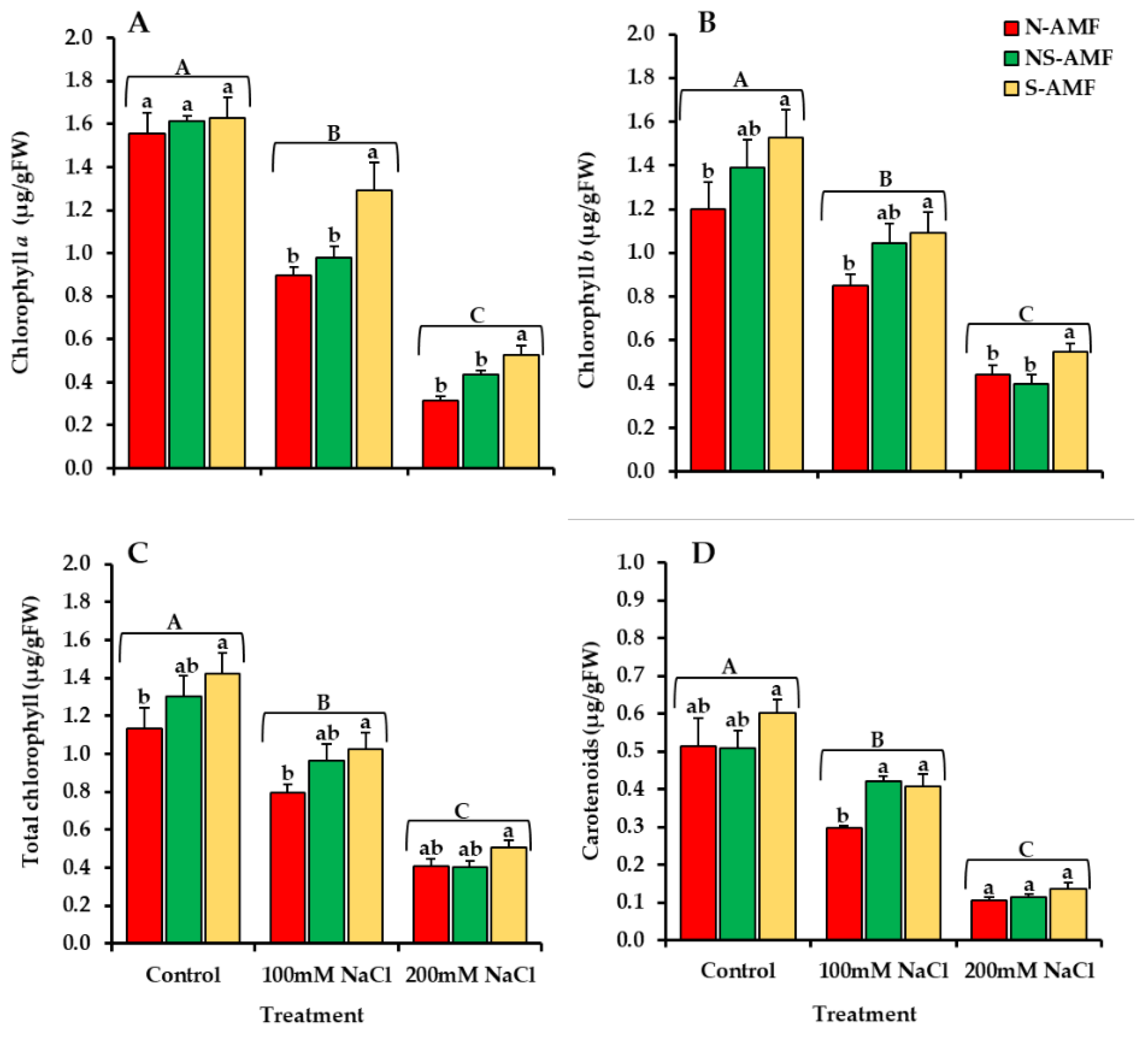
2.6. Chlorophyll Contents and Carotenoids
2.7. Estimation of Proline
2.8. Total Phenolic Content in L. scindicus
2.9. Determination of Hydrogen Peroxide (H2O2) Content
2.10. Protein Estimation and Antioxidant Enzyme Activity
2.10.1. Superoxide-Dismutase (SOD, EC 1.15.1.1)
2.10.2. The Catalase (CAT, EC 1.11.1.6)
2.10.3. Ascorbate-Peroxidase (APX, EC 1.11.1.11)
2.10.4. Glutathione-Reductase (G.R., EC 1.6.4.2)
2.10.5. Monodehydroascorbate-Reductase (MDHAR, EC 1.6.5.4)
2.11. Statistical Analysis
3. Results
3.1. Effect of Salinity Stress on AMF Colonization of L. scindicus Plants and Total Spore Count
3.2. Growth Parameters of S-AMF- and NS-AMF- Inoculated L. scindicus Plants under Different Levels of NaCl Stress
3.3. Chlorophyll and Carotenoid Contents of L. scindicus
3.4. Proline and Total Phenol Contents
3.5. Hydrogen Peroxide (H2O2) Content
3.6. Protein Content and Antioxidant Enzyme Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alotaibi, M.O.; Sonbol, H.S.; Alwakeel, S.S.; Suliman, R.S.; Fodah, R.A.; Jaffal, A.S.A.; AlOthman, N.I.; Mohammed, A.E. Microbial diversity of some sabkha and desert sites in Saudi Arabia. Saudi J. Biol. Sci. 2020, 27, 2778–2789. [Google Scholar] [CrossRef] [PubMed]
- Al-Amoudi, O.S.B. Studies on Soil-Foundation Interaction in the Sabkha Environment of Eastern Province of Saudi Arabia. Ph.D. Thesis, King Fahd University of Petroleum and Minerals, Dhahran, Saudi Arabia, 1992. [Google Scholar]
- Abd-ElGawad, A.M.; Assaeed, A.M.; Al-Rowaily, S.L.; Dar, B.M.; Malik, J.A. Moisture and Salinity Drive the Vegetation Composition of Wadi Hargan, Riyadh, Saudi Arabia. Diversity 2021, 13, 587. [Google Scholar] [CrossRef]
- Al-Fredan, M.A. Sand dune and sabkha vegetations of Eastern Saudi Arabia. Int. J. Bot. 2008, 4, 196–204. [Google Scholar] [CrossRef]
- Etesami, H.; Noori, F. Soil salinity as a challenge for sustainable agriculture and bacterial-mediated alleviation of salinity stress in crop plants. In Saline Soil-Based Agriculture by Halotolerant Microorganisms; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–22. [Google Scholar]
- Ivushkin, K.; Bartholomeus, H.; Bregt, A.K.; Pulatov, A.; Kempen, B.; De Sousa, L. Global mapping of soil salinity change. Remote Sens. Environ. 2019, 231, 111260. [Google Scholar] [CrossRef]
- Din, B.U.; Sarfraz, S.; Xia, Y.; Kamran, M.A.; Javed, M.T.; Sultan, T.; Munis, M.F.H.; Chaudhary, H.J. Mechanistic elucidation of germination potential and growth of wheat inoculated with exopolysaccharide and ACC-deaminase producing Bacillus strains under induced salinity stress. Ecotoxicol. Environ. Saf. 2019, 183, 109466. [Google Scholar]
- Lakhdar, A.; Rabhi, M.; Ghnaya, T.; Montemurro, F.; Jedidi, N.; Abdelly, C. Effectiveness of compost use in salt-affected soil. J. Hazard. Mater. 2009, 171, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Rizwan, M.; Qayyum, M.F.; Ok, Y.S.; Ibrahim, M.; Riaz, M.; Arif, M.S.; Hafeez, F.; Al-Wabel, M.I.; Shahzad, A.N. Biochar soil amendment on alleviation of drought and salt stress in plants: A critical review. Environ. Sci. Pollut. Res. 2017, 24, 12700–12712. [Google Scholar] [CrossRef] [PubMed]
- Nan, X.; Huihui, Z.; Haixiu, Z.; Yining, W.; Jinbo, L.; Li, X.; Zepeng, Y.; Wenxu, Z.; Yi, Q.; Guangyu, S. The response of photosynthetic functions of F1 cutting seedlings from Physocarpus amurensis Maxim (♀)× Physocarpus opulifolius “Diabolo”(♂) and the parental seedlings to salt stress. Front. Plant Sci. 2018, 9, 714. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, M.; Poyraz, İ.; Çavdar, A.; Özgen, Y.; Beyaz, R. Plant Responses to Salt Stress. In Plant Breeding-Current Future Views; Intechopen: London, UK, 2020. [Google Scholar]
- Kumar, S.; Li, G.; Yang, J.; Huang, X.; Ji, Q.; Liu, Z.; Ke, W.; Hou, H. Effect of salt stress on growth, physiological parameters, and ionic concentration of water dropwort (Oenanthe javanica) cultivars. Front. Plant Sci. 2021, 12, 660409. [Google Scholar] [CrossRef] [PubMed]
- Fayaz, F.; Zahedi, M. Beneficial effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) nutritional status and tolerance indices under soil salinity stress. J. Plant Nutr. 2021, 45, 185–201. [Google Scholar] [CrossRef]
- Liang, S.; Jiang, Y.; Li, M.; Zhu, W.; Xu, N.; Zhang, H. Improving plant growth and alleviating photosynthetic inhibition from salt stress using AMF in alfalfa seedlings. J. Plant Interact. 2019, 14, 482–491. [Google Scholar] [CrossRef] [Green Version]
- Sofy, M.R.; Aboseidah, A.A.; Heneidak, S.A.; Ahmed, H.R. ACC deaminase containing endophytic bacteria ameliorate salt stress in Pisum sativum through reduced oxidative damage and induction of antioxidative defense systems. Environ. Sci. Pollut. Res. 2021, 28, 40971–40991. [Google Scholar] [CrossRef] [PubMed]
- Sofy, M.; Mohamed, H.; Dawood, M.; Abu-Elsaoud, A.; Soliman, M. Integrated usage of Trichoderma harzianum and biochar to ameliorate salt stress on spinach plants. Arch. Agron. Soil Sci. 2021, 1–22. [Google Scholar] [CrossRef]
- Wu, H.; Shabala, L.; Zhou, M.; Stefano, G.; Pandolfi, C.; Mancuso, S.; Shabala, S. Developing and validating a high-throughput assay for salinity tissue tolerance in wheat and barley. Planta 2015, 242, 847–857. [Google Scholar] [CrossRef]
- Silva, P.C.C.; Azevedo Neto, A.D.d.; Gheyi, H.R.; Ribas, R.F.; Silva, C.R.d.R.; Cova, A.M.W. Salt tolerance induced by hydrogen peroxide priming on seed is related to improvement of ion homeostasis and antioxidative defense in sunflower plants. J. Plant Nutr. 2020, 44, 1207–1221. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [Green Version]
- Goddard, M.-L.; Belval, L.; Martin, I.R.; Roth, L.; Laloue, H.; Deglène-Benbrahim, L.; Valat, L.; Bertsch, C.; Chong, J. Arbuscular mycorrhizal symbiosis triggers major changes in primary metabolism together with modification of defense responses and signaling in both roots and leaves of Vitis vinifera. Front. Plant Sci. 2021, 25, 1675. [Google Scholar] [CrossRef]
- Borde, M.; Dudhane, M.; Kulkarni, M. Role of arbuscular mycorrhizal fungi (AMF) in salinity tolerance and growth response in plants under salt stress conditions. In Mycorrhiza-Eco-Physiology, Secondary Metabolites, Nanomaterials; Springer: Berlin/Heidelberg, Germany, 2017; pp. 71–86. [Google Scholar]
- Klinsukon, C.; Lumyong, S.; Kuyper, T.W.; Boonlue, S. Colonization by arbuscular mycorrhizal fungi improves salinity tolerance of eucalyptus (Eucalyptus camaldulensis) seedlings. Sci. Rep. 2021, 11, 4362. [Google Scholar] [CrossRef]
- Lenoir, I.; Fontaine, J.; Sahraoui, A.L.-H. Arbuscular mycorrhizal fungal responses to abiotic stresses: A review. Phytochemistry 2016, 123, 4–15. [Google Scholar] [CrossRef]
- Giovannini, L.; Palla, M.; Agnolucci, M.; Avio, L.; Sbrana, C.; Turrini, A.; Giovannetti, M. Arbuscular mycorrhizal fungi and associated microbiota as plant biostimulants: Research strategies for the selection of the best performing inocula. Agronomy 2020, 10, 106. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.-Y.; Liu, R.-J.; Lin, X.-G.; Zhou, J.-M. Arbuscular mycorrhizal status of wild plants in saline-alkaline soils of the Yellow River Delta. Mycorrhiza 2004, 14, 133–137. [Google Scholar] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Evelin, H.; Devi, T.S.; Gupta, S.; Kapoor, R. Mitigation of salinity stress in plants by arbuscular mycorrhizal symbiosis: Current understanding and new challenges. Front. Plant Sci. 2019, 10, 470. [Google Scholar] [CrossRef] [Green Version]
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J.M. Salinity stress alleviation using arbuscular mycorrhizal fungi. A review. Agron. Sustain. Dev. 2012, 32, 181–200. [Google Scholar] [CrossRef] [Green Version]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Aldubise, A.; Egamberdieva, D. Arbuscular mycorrhizal fungi enhances salinity tolerance of Panicum turgidum Forssk by altering photosynthetic and antioxidant pathways. J. Plant Interact. 2015, 10, 230–242. [Google Scholar] [CrossRef] [Green Version]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.-B.F.; Aldehaish, H.A.; Egamberdieva, D.; Abd_Allah, E.F. Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 2018, 25, 1102–1114. [Google Scholar] [CrossRef] [PubMed]
- Beltrano, J.; Ruscitti, M.; Arango, M.C.; Ronco, M. Effects of arbuscular mycorrhiza inoculation on plant growth, biological and physiological parameters and mineral nutrition in pepper grown under different salinity and p levels. J. Soil Sci. Plant Nutr. 2013, 13, 123–141. [Google Scholar] [CrossRef] [Green Version]
- Porcel, R.; Redondo-Gómez, S.; Mateos-Naranjo, E.; Aroca, R.; Garcia, R.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal symbiosis ameliorates the optimum quantum yield of photosystem II and reduces non-photochemical quenching in rice plants subjected to salt stress. J. Plant Physiol. 2015, 185, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Assaeed, A.M.; Al-Doss, A.A. Seedling competition of Lasiurus scindicus and Rhazya stricta in response to water stress. J. Arid. Environ. 2001, 49, 315–320. [Google Scholar] [CrossRef]
- Cunningham, P.L. Plants included in the diet of Arabian sand gazelle (Reem) from Saudi Arabia. J. King Saud Univ.-Sci. 2013, 25, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Sanadya, S.K.; Shekhawat, S.S.; Sahoo, S. Sewan Grass: A Potential Forage Grass in Arid Environments. In Grasses and Grassland—New Perspectives; Intechopen: London, UK, 2021. [Google Scholar]
- Al-Rowaily, S.L.; Abd-ElGawad, A.M.; Alghanem, S.M.; Al-Taisan, W.A.A.; El-Amier, Y.A. Nutritional Value, Mineral Composition, Secondary Metabolites, and Antioxidant Activity of Some Wild Geophyte Sedges and Grasses. Plants 2019, 8, 569. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, S.A.; Al-Jowaid, A.A.A. Vegetation of the Kingdom of Saudi Arabia; National Agricultural and Water Research Center, Ministry of Agriculture And Water: Ibaraki, Saudi Arabia, 1999. [Google Scholar]
- Chauhan, S.S. Desertification control and management of land degradation in the Thar desert of India. Environmentalist 2003, 23, 219–227. [Google Scholar] [CrossRef]
- Naz, N.; Rafique, T.; Hameed, M.; Ashraf, M.; Batool, R.; Fatima, S. Morpho-anatomical and physiological attributes for salt tolerance in sewan grass (Lasiurus scindicus Henr.) from Cholistan Desert, Pakistan. Acta Physiol. Plant. 2014, 36, 2959–2974. [Google Scholar] [CrossRef]
- Gadi, B.; Goswami, B. Effect of salt stress on early seedling growth and chlorophyll stability index of Lasiurus sindicus Henr: Endemic to Indian Thar desert. Biochem. Cell Arch. 2016, 16, 103–106. [Google Scholar]
- El-Keblawy, A.; Al-Ansari, F.; Al-Shamsi, N. Effects of temperature and light on salinity tolerance during germination in two desert glycophytic grasses, Lasiurus scindicus and Panicum turgidum. Grass Forage Sci. 2011, 66, 173–182. [Google Scholar] [CrossRef]
- Naz, N.; Batool, R.; Fatima, S.; Hameed, M.; Ashraf, M.; Ahmad, F.; Ahmad, M.S.A. Adaptive components of tolerance to salinity in a saline desert grass Lasiurus scindicus Henrard. Ecol. Res. 2015, 30, 429–438. [Google Scholar] [CrossRef]
- Ashraf, M.; Hameed, M.; Arshad, M.; Ashraf, Y.; Akhtar, K. Salt tolerance of some potential forage grasses from Cholistan desert of Pakistan. In Ecophysiology of High Salinity Tolerant Plants; Springer: Berlin/Heidelberg, Germany, 2008; pp. 31–54. [Google Scholar]
- Estrada, B.; Aroca, R.; Barea, J.M.; Ruiz-Lozano, J.M. Native arbuscular mycorrhizal fungi isolated from a saline habitat improved maize antioxidant systems and plant tolerance to salinity. Plant Sci. 2013, 201, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Outamamat, E.; Bourhia, M.; Dounas, H.; Salamatullah, A.M.; Alzahrani, A.; Alyahya, H.K.; Albadr, N.A.; Al Feddy, M.N.; Mnasri, B.; Ouahmane, L. Application of Native or Exotic Arbuscular Mycorrhizal Fungi Complexes and Monospecific Isolates from Saline Semi-Arid Mediterranean Ecosystems Improved Phoenix dactylifera’s Growth and Mitigated Salt Stress Negative Effects. Plants 2021, 10, 2501. [Google Scholar] [CrossRef] [PubMed]
- Gerdemann, J.; Nicolson, T.H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Walker, C. Spore Extraction by Centrifugation-Sugar Flotation; Biological Research and Imaging Laboratory: Hampshire, UK, 1997. [Google Scholar]
- Redecker, D.; Schüßler, A.; Stockinger, H.; Stürmer, S.L.; Morton, J.B.; Walker, C. An evidence-based consensus for the classification of arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza 2013, 23, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Schüssler, A.; Walker, C. The Glomeromycota: A Species List with New Families and New Gener. In The Glomeromycota; CreateSpace Independent Publishing Platform: Scotts Valley, CA, USA, 2010. [Google Scholar]
- Phillips, J.M.; Hayman, D. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.; Gianinazzi-Pearson, V. Evaluation of VA infection levels in root systems. Research for estimation methods having a functional significance. Physiol. Genet. Asp. Mycorrhizae 1986, 1, 217–221. [Google Scholar]
- Al-Qarawi, A.; Mridha, M.; Alghamdi, O. Diversity of structural colonization and spore population of arbuscular mycorrhizal fungi in some plants from Riyadh, Saudi Arabia. J. Pure Appl. Microbiol. 2012, 6, 1119–1125. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajput, R.; Patil, R. The comparative study on spectrophotometric analysis of chlorophyll and carotenoids pigments from non-leguminous fodder crops. Int. J. Innov. Sci. Eng. Technol. 2017, 4, 140–148. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Sergiev, I.; Alexieva, V.; Karanov, E. Effect of spermine, atrazine and combination between them on some endogenous protective systems and stress markers in plants. Comptes Rendus De L’académie Bulg. Des Sci. 1997, 51, 121–124. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Schaedle, M.; Bassham, J.A. Chloroplast glutathione reductase. Plant Physiol. 1977, 59, 1011–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, S.G.; Ju, Z.; van Santen, E.; Qiu, J.; Weaver, D.B.; Prior, S.A.; Rogers, H.H. The influence of elevated CO2 on the activities of antioxidative enzymes in two soybean genotypes. Funct. Plant Biol. 2000, 27, 1061–1068. [Google Scholar] [CrossRef]
- Miransari, M. Contribution of arbuscular mycorrhizal symbiosis to plant growth under different types of soil stress. Plant Biol. 2010, 12, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Yarahmadi, M.A.; Shahsavani, S.; Akhyani, A.; Dorostkar, V. Pomegranate growth affected by arbuscular mycorrhizae, phosphorus fertilizer, and irrigation water salinity. Commun. Soil Sci. Plant Anal. 2018, 49, 478–488. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Kim, K.; Kim, C.; Sa, T. Changes of arbuscular mycorrhizal traits and community structure with respect to soil salinity in a coastal reclamation land. Soil Biol. Biochem. 2014, 72, 1–10. [Google Scholar] [CrossRef]
- Sharifi, M.; Ghorbanli, M.; Ebrahimzadeh, H. Improved growth of salinity-stressed soybean after inoculation with salt pre-treated mycorrhizal fungi. J. Plant Physiol. 2007, 164, 1144–1151. [Google Scholar] [CrossRef]
- Aliasgharzadeh, N.; Rastin, S.N.; Towfighi, H.; Alizadeh, A. Occurrence of arbuscular mycorrhizal fungi in saline soils of the Tabriz Plain of Iran in relation to some physical and chemical properties of soil. Mycorrhiza 2001, 11, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Alqarawi, A.A.; Abd Allah, E.; Hashem, A. Alleviation of salt-induced adverse impact via mycorrhizal fungi in Ephedra aphylla Forssk. J. Plant Interact. 2014, 9, 802–810. [Google Scholar] [CrossRef] [Green Version]
- Elhindi, K.M.; El-Din, A.S.; Elgorban, A.M. The impact of arbuscular mycorrhizal fungi in mitigating salt-induced adverse effects in sweet basil (Ocimum basilicum L.). Saudi J. Biol. Sci. 2017, 24, 170–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif, P.; Seyedsalehi, M.; Paladino, O.; Van Damme, P.; Sillanpää, M.; Sharifi, A. Effect of drought and salinity stresses on morphological and physiological characteristics of canola. Int. J. Environ. Sci. Technol. 2018, 15, 1859–1866. [Google Scholar] [CrossRef]
- Meriem, B.F.; Kaouther, Z.; Chérif, H.; Tijani, M.; André, B. Effect of priming on growth, biochemical parameters and mineral composition of different cultivars of coriander (Coriandrum sativum L.) under salt stress. J. Stress Physiol. Biochem. 2014, 10, 84–109. [Google Scholar]
- Taibi, K.; Taibi, F.; Abderrahim, L.A.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. S. Afr. J. Bot. 2016, 105, 306–312. [Google Scholar] [CrossRef]
- Guo, R.; Zhou, J.; Hao, W.; Gu, F.; Liu, Q.; Li, H.; Xia, X.; Mao, L. Germination, growth, chlorophyll fluorescence and ionic balance in linseed seedlings subjected to saline and alkaline stresses. Plant Prod. Sci. 2014, 17, 20–31. [Google Scholar] [CrossRef] [Green Version]
- Abeer, H.; Abd_Allah, E.; Alqarawi, A.; El-Didamony, G.; Alwhibi, M.; Egamberdieva, D.; Ahmad, P. Alleviation of adverse impact of salinity on faba bean (Vicia faba L.) by arbuscular mycorrhizal fungi. Pak. J. Bot. 2014, 46, 2003–2013. [Google Scholar]
- Ahmad, P.; Abdel Latef, A.A.; Hashem, A.; Abd_Allah, E.F.; Gucel, S.; Tran, L.-S.P. Nitric oxide mitigates salt stress by regulating levels of osmolytes and antioxidant enzymes in chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Li, Z.; Wu, F.; Tang, M. Comparative photochemistry activity and antioxidant responses in male and female Populus cathayana cuttings inoculated with arbuscular mycorrhizal fungi under salt. Sci. Rep. 2016, 6, 37663. [Google Scholar] [CrossRef] [PubMed]
- Mehr, Z.; Khajeh, H.; Bahabadi, S.; Sabbagh, S. Changes on proline, phenolic compounds and activity of antioxidant enzymes in Anethum graveolens L. under salt stress. Int. J. Agron. Plant Prod. 2012, 3, 710–715. [Google Scholar]
- Rezazadeh, A.; Ghasemnezhad, A.; Barani, M.; Telmadarrehei, T. Effect of salinity on phenolic composition and antioxidant activity of artichoke (Cynara scolymus L.) leaves. Res. J. Med. Plant 2012, 6, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Bors, W.; Michel, C.; Saran, M. Flavonoid antioxidants: Rate constants for reactions with oxygen radicals. Methods Enzymol. 1994, 234, 420–429. [Google Scholar] [PubMed]
- Sarabi, B.; Bolandnazar, S.; Ghaderi, N.; Tabatabaei, S.J. Multivariate analysis as a tool for studying the effects of salinity in different melon landraces at germination stage. Not. Bot. Horti Agrobot. 2016, 44, 264–271. [Google Scholar] [CrossRef] [Green Version]
- Ait-El-Mokhtar, M.; Baslam, M.; Ben-Laouane, R.; Anli, M.; Boutasknit, A.; Mitsui, T.; Wahbi, S.; Meddich, A. Alleviation of detrimental effects of salt stress on date palm (Phoenix dactylifera L.) by the application of arbuscular mycorrhizal fungi and/or compost. Front. Sustain. Food Syst. 2020, 4, 131. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A.; Linn, S. DNA damage and oxygen radical toxicity. Science 1988, 240, 1302–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Wirth, S.; Egamberdieva, D. Arbuscular mycorrhizal fungi alleviate salt stress in lupine (Lupinus termis Forsik) through modulation of antioxidant defense systems and physiological traits. Legume Res. -Int. J. 2016, 39, 198–207. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, M.; Boopathi, T.; Manivannan, P. Comprehensive Assessment of Ameliorative Effects of AMF in Alleviating Abiotic Stress in Tomato Plants. J. Fungi 2021, 7, 303. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Alyemeni, M.N.; Al-Huqail, A.A.; Alqahtani, M.A.; Wijaya, L.; Ashraf, M.; Kaya, C.; Bajguz, A. Zinc oxide nanoparticles application alleviates arsenic (As) toxicity in soybean plants by restricting the uptake of as and modulating key biochemical attributes, antioxidant enzymes, ascorbate-glutathione cycle and glyoxalase system. Plants 2020, 9, 825. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Status | Spores/100 g Dry Soil |
|---|---|---|
| Control | N-AMF | 0.00 |
| NS-AMF | 211 ± 14.799 B,a | |
| S-AMF | 196.7 ± 14.495 B,a | |
| 100 mM NaCl | N-AMF | 0.00 |
| NS-AMF | 292 ± 8.622 A,a | |
| S-AMF | 282.7 ± 11.865 A,a | |
| 200 mM NaCl | N-AMF | 0.00 |
| NS-AMF | 35.3 ± 4.096 C,b | |
| S-AMF | 76 ± 7.371 C,a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malik, J.A.; AlQarawi, A.A.; Dar, B.A.; Hashem, A.; Alshahrani, T.S.; AlZain, M.N.; Habib, M.M.; Javed, M.M.; Abd_Allah, E.F. Arbuscular Mycorrhizal Fungi Isolated from Highly Saline “Sabkha Habitat” Soil Alleviated the NaCl-Induced Stress and Improved Lasiurus scindicus Henr. Growth. Agriculture 2022, 12, 337. https://doi.org/10.3390/agriculture12030337
Malik JA, AlQarawi AA, Dar BA, Hashem A, Alshahrani TS, AlZain MN, Habib MM, Javed MM, Abd_Allah EF. Arbuscular Mycorrhizal Fungi Isolated from Highly Saline “Sabkha Habitat” Soil Alleviated the NaCl-Induced Stress and Improved Lasiurus scindicus Henr. Growth. Agriculture. 2022; 12(3):337. https://doi.org/10.3390/agriculture12030337
Chicago/Turabian StyleMalik, Jahangir A., AbdulAziz A. AlQarawi, Basharat A. Dar, Abeer Hashem, Thobayet S. Alshahrani, Mashail N. AlZain, Muhammad M. Habib, Muhammad M. Javed, and Elsayed Fathi Abd_Allah. 2022. "Arbuscular Mycorrhizal Fungi Isolated from Highly Saline “Sabkha Habitat” Soil Alleviated the NaCl-Induced Stress and Improved Lasiurus scindicus Henr. Growth" Agriculture 12, no. 3: 337. https://doi.org/10.3390/agriculture12030337