
Different Host Plants Distinctly Influence the Adaptability of Myzus persicae (Hemiptera: Aphididae)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials
2.1.1. Insects
2.1.2. Plants
2.2. Test Methods
2.2.1. EPG Parameter Measure
2.2.2. Life Table Parameter Measure
2.2.3. Life Table Analysis
2.2.4. Population Development
2.2.5. Data Analysis
3. Results
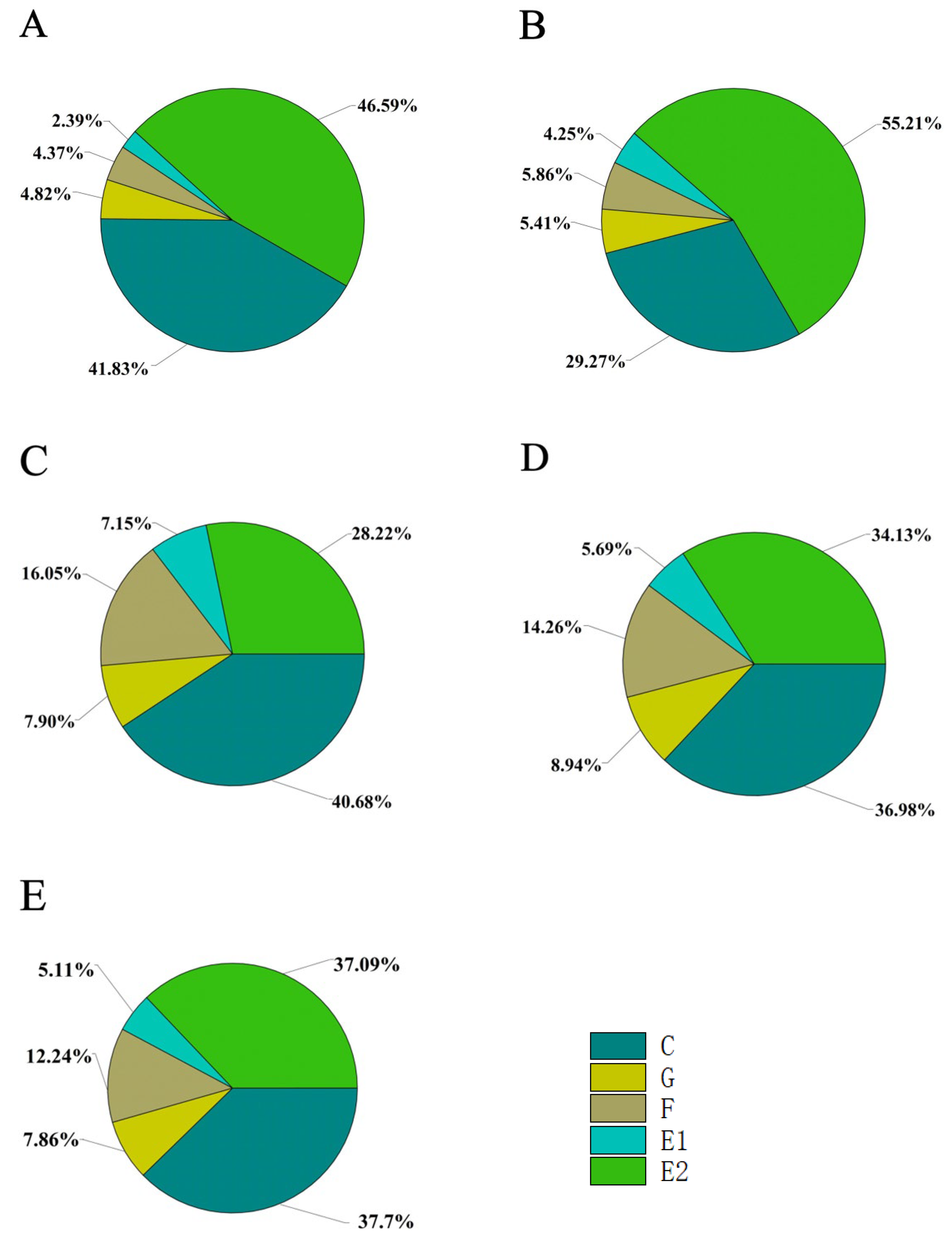
3.1. Probing Behavior of M. persicae on Five Different Host Plants
3.2. Pathway Activities
3.3. Phloem and Xylem Activities
3.4. Life History Statistics of M. persicae on Five different Host Plants
3.5. Population Parameters of M. persicae Reared on Five Host Plants
3.6. Population Dynamics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peng, L.; Yan, Y.; Liu, W.X.; Wan, F.H.; Wang, J.J. Counter-defense mechanisms of phytophagous insects towards plant defense. Acta Entomol. Sin. 2010, 53, 572–580. [Google Scholar] [CrossRef]
- Fang, Y.; Qiao, G.X.; Zhang, G.X. Morphological adaptation of aphid species on different host plant leaves. Acta Entomol. Sin. 2011, 54, 157–178. [Google Scholar] [CrossRef]
- Li, F.Q. The Population Dynamics and Different Expressed Genes of Myzus persicae (Sulzer) on Different Host Plants. Master’s Thesis, Henan Agricultural University, Henan, China, 2013. [Google Scholar]
- Zhou, M. Principle and Application of Insect Resistance in Crops; Beijing Agricultural University Press: Beijing, China, 1992. [Google Scholar]
- Khokhlova, I.S.; van der Mescht, L.; Warburton, E.M.; Stavtseva, N.A.; Krasnov, B.R. Adaptation to a novel host and performance trade-off in host-generalist and host-specific insect ectoparasites. Insect Sci. 2021, 29, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Santos, N.A.; Teixeira, N.C.; Valim, J.O.S.; Almeida, E.F.A.; Oliveira, M.G.A.; Campos, W.G. Sulfur fertilization increases defense metabolites and nitrogen but decreases plant resistance against a host-specific insect. Bull. Entomol. Res. 2017, 108, 479–486. [Google Scholar] [CrossRef] [PubMed]
- He, Y.Q.; Zhang, Y.Q.; Chen, J.N.; Chen, W.L.; Zeng, X.Y.; Chen, H.T.; Ding, W. Effects of Aphidius gifuensis on the feeding behavior and potato virus Y transmission ability of Myzus persicae. Insect Sci. 2017, 25, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.A.; Radcliffe, E.B.; Ragsdale, D.W. Resistance to green peach aphid, Myzus persicae (Sulzer), and potato aphid, Macrosiphum euphorbiae (Thomas), in potato cultivars. Am. J. Potato Res. 2007, 84, 259–269. [Google Scholar] [CrossRef]
- Bai, T.Q.; Zhang, L.M.; Gu, X.H.; Ji, S.G.; Sun, Y.X.; Yang, S.Y.; Zhang, H.R. Rearing of Myzus persicae and Aphidius gifuensis on Cruciferous Crops. J. Yunnan Agric. Univ. 2015, 30, 541–546. [Google Scholar] [CrossRef]
- Cao, Y.; Zeng, L.; Cui, Z.X.; Wang, Y.B.; Chen, W.S. Study of the effect of host plants on peach aphid on lily. J. Southwest Agric. Univ. (Nat. Sci.) 2004, 26, 165–168. [Google Scholar]
- Zhou, L.F. Influence of Hosts on Population of Myzus persicae (Sulzer) and Coat Protein Gene and Sequence Analysis of TuMV, TMV and CMV. Master’s Thesis, Southwest University, Chongqing, China, 2007. [Google Scholar]
- Ng, J.C.K.; Perry, K.L. Transmission of plant viruses by aphid vectors. Mol. Plant Pathol. 2004, 5, 505–511. [Google Scholar] [CrossRef]
- Whitfield, A.E.; Falk, B.W.; Rotenberg, D. Insect vector-mediated transmission of plant viruses. Virology 2015, 479–480, 278–289. [Google Scholar] [CrossRef] [Green Version]
- Jhou, Y.S.; Poovendhan, S.; Huang, L.H.; Tsai, C.W. Host Acceptance and Plant Resistance: A Comparative Behavioral Study of Myzus persicae and Acyrthosiphon pisum. Insects 2021, 12, 975. [Google Scholar] [CrossRef] [PubMed]
- Tjallingii, W.F.; Esch, T.H. Fine structure of aphid stylet routes in plant tissues in correlation with EPG signals. Physiol. Entomol. 1993, 18, 317–328. [Google Scholar] [CrossRef]
- Martín, B.; Collar, J.L.; Tjallingii, W.F.; Fereres, A. Intracellular ingestion and salivation by aphids may cause the acquisition and inoculation of non-persistently transmitted plant viruses. J. Gen. Virol. 1997, 78, 2701–2705. [Google Scholar] [CrossRef] [PubMed]
- Tjallingii, W.F. Membrane potentials as an indication for plant cell penetration by aphid stylets. Entomol. Exp. Appl. 1985, 38, 187–193. [Google Scholar] [CrossRef]
- Tjallingii, W.F.; Garzo, E.; Fereres, A. New structure in cell puncture activities by aphid stylets: A dual-mode EPG study. Entomol. Exp. Appl. 2010, 135, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Tjallingii, W.F. Electrical nature of recorded signals during stylet penetration by aphids. Entomol. Exp. Appl. 1985, 38, 177–186. [Google Scholar] [CrossRef]
- Liu, X.D.; Zhang, X.X.; Zhai, B.P. The EPG Recording of Feeding Behavior of Aphis gossypii After Inter transforming between Cotton and Cucurbits. J. Cotton Sci. 2002, 14, 33–35. [Google Scholar]
- Liu, X.D. Studies on Host and Migration Biotypes of Aphis Gossypii Glover. Ph.D. Thesis, Nanjing Agricultural University, Jiangsu, China, 2003. [Google Scholar]
- Lu, H. Effect of Host Alteration on Life Cycle, Feeding Behavior and Salivary Gland Gene Expression of Pea Aphid. Ph.D. Thesis, Shandon Agricultural University, Shandon, China, 2016. [Google Scholar]
- Chi, H.; Su, H.Y. Age-Stage, Two-Sex Life Tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and Its Host Myzus persicae (Sulzer) (Homoptera: Aphididae) with Mathematical Proof of the Relationship Between Female Fecundity and the Net Reproductive Rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Govindan, B.N.; Hutchison, W.D. Influence of Temperature on Age-Stage, Two-Sex Life Tables for a Minnesota-Acclimated Population of the Brown Marmorated Stink Bug (Halyomorpha halys). Insects 2020, 11, 108. [Google Scholar] [CrossRef] [Green Version]
- Tjallingii, W.F. Electronic recording of penetration behaviour by aphids. Entomol. Exp. Appl. 1978, 24, 721–730. [Google Scholar] [CrossRef]
- Spiller, N.J.; Koenders, L.; Tjallingii, W.F. Xylem ingestion by aphids—A strategy for maintaining water balance. Entomol. Exp. Appl. 1990, 55, 101–104. [Google Scholar] [CrossRef]
- Garzo, E.; Moreno, A.; Plaza, M.; Fereres, A. Feeding Behavior and Virus-transmission Ability of Insect Vectors Exposed to Systemic Insecticides. Plants 2020, 9, 895. [Google Scholar] [CrossRef] [PubMed]
- Goodman, D. Optimal Life Histories, Optimal Notation, and the Value of Reproductive Value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-Table Analysis Incorporating Both Sexes and Variable Development Rates Among Individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis; National Chung Hsing University: Taichung, Taiwan, 2022; Available online: http://140.120.197.173/Ecology/ (accessed on 8 March 2021).
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; Chapman and Hall/CRC Press: New York, NY, USA, 1994. [Google Scholar] [CrossRef]
- Huang, Y.B.; Chi, H. Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Sci. 2012, 19, 263–273. [Google Scholar] [CrossRef]
- Liu, P.; Ma, H.; Zhu, Q.S.; Chen, B.C.; Gao, J.; Lin, X.Q. Research Progress of Insect Adaptability to Their Host Plants. Biol. Disaster Sci. 2016, 39, 250–254. [Google Scholar] [CrossRef]
- Yan, F.M.; Wang, M.Q. Insect Electrical Penetration Graph and Its Applications; Henan Science and Technology Press: Zhengzhou, China, 2017. [Google Scholar]
- Zeng, X.Y.; He, Y.Q.; Wu, J.X.; Tang, Y.M.; Gu, J.; Ding, W.; Zhang, Y.Q. Sublethal Effects of Cyantraniliprole and Imidacloprid on Feeding Behavior and Life Table Parameters of Myzus persicae (Hemiptera: Aphididae). J. Econ. Entomol. 2016, 109, 1595–1602. [Google Scholar] [CrossRef]
- Alvarez, A.E.; Tjallingii, W.F.; Garzo, E.; Vleeshouwers, V.; Dicke, M.; Vosman, B. Location of resistance factors in the leaves of potato and wild tuber-bearing Solanum species to the aphid Myzus persicae. Entomol. Exp. Appl. 2006, 121, 145–157. [Google Scholar] [CrossRef]
- Ten Broeke, C.J.M.; Dicke, M.; van Loon, J.J.A. Performance and feeding behaviour of two biotypes of the black currant-lettuce aphid, Nasonovia ribisnigri, on resistant and susceptible Lactuca sativa near-isogenic lines. Bull. Entomol. Res. 2013, 103, 511–521. [Google Scholar] [CrossRef]
- Powell, G.; Tosh, C.R.; Hardie, J. Host Plant Selection by Aphids: Behavioral, Evolutionary, and Applied Perspectives. Annu. Rev. Entomol. 2006, 51, 309–330. [Google Scholar] [CrossRef] [PubMed]
- Wójcicka, A. Surface Waxes as a Plant Defense Barrier Towards Grain Aphid. Acta Biol. Crac. S. Bot. 2015, 57, 95–103. [Google Scholar] [CrossRef]
- Björkman, M.; Klingen, I.; Birch, A.N.E.; Bones, A.M.; Bruce, T.J.A.; Johansen, T.J.; Meadow, R.; Mølmann, J.; Seljåsen, R.; Smart, L.E.; et al. Phytochemicals of Brassicaceae in plant protection and human health—Influences of climate, environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.N.; Wu, C.X.; He, Y.Q.; Yu, C.; Liu, J.F.; Li, T.S.; Zhou, C.Y.; Chen, W.L. Different Host Plants Distinctly Influence the Feeding Ability of the Brown Citrus Aphid Toxoptera citricida. Insects 2021, 12, 864. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.H.; Wu, J.; Zhang, Z.F.; Liu, T.X. Phloem nutrition of detached cabbage leaves varies with leaf age and influences performance of the green peach aphid, Myzus persicae. Entomol. Exp. Appl. 2018, 166, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Miao, J.; Han, B.Y.; Zhang, Q.H. Probing behavior of Empoasca vitis (Homoptera: Cicadellidae) on resistant and susceptible cultivars of tea plants. J. Insect Sci. 2014, 14, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Zhou, C.; Long, G.; Yang, X.; Wei, Z.; Liao, Y.; Yang, H.; Hu, C. Fitness of fall armyworm, Spodoptera frugiperda to three solanaceous vegetables. J. Integr. Agric. 2021, 20, 755–763. [Google Scholar] [CrossRef]
- Sousa, F.F.; Mendes, S.M.; Santos-Amaya, O.F.; Araújo, O.G.; Oliveira, E.E.; Pereira, E.J.G. Life-History Traits of Spodoptera frugiperda Populations Exposed to Low-Dose Bt Maize. PLoS ONE 2016, 11, e0156608. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.J.; Zhang, L.; Cheng, Y.X.; Jiang, X.F. Age-stage two-sex life table for laboratory populations of fall armyworm, Spodoptera frugiperda at different temperatures. Plant Prot. 2019, 45, 20–27. [Google Scholar] [CrossRef]
- Özgökçe, M.S.; Chi, H.; Atlıhan, R.; Kara, H. Demography and population projection of Myzus persicae (Sulz.) (Hemiptera: Aphididae) on five pepper (Capsicum annuum L.) cultivars. Phytoparasitica 2018, 46, 153–167. [Google Scholar] [CrossRef]
- Goundoudaki, S.; Tsitsipis, J.A.; Margaritopoulos, J.T.; Zarpas, K.D.; Divanidis, S. Performance of the tobacco aphid Myzus persicae (Hemiptera: Aphididae) on Oriental and Virginia tobacco varieties. Agric. For. Entomol. 2003, 5, 285–291. [Google Scholar] [CrossRef]
- Bahram, N.; Fatemeh, H.; Asgar, E.; Fatemeh, S. Physicochemical traits of Vicia faba L. seed cultivars affect oviposition preference and demographic parameters of Callosobruchus maculatus (F.) (Coleoptera: Chrysomelidae). J. Stored Prod. Res. 2022, 95, 101924. [Google Scholar] [CrossRef]
- Hon, F. Research about the Occurrence of Alatus Tobacco Aphids and Tobacco Mosaic Disease in Fields, and the Life Table of Myzus persicae on Different Host Plants. Ph.D. Thesis, Southwest University, Chongqing, China, 2017. [Google Scholar]



{kind=link}
{kind=link}
| EPG Parameter | Tobacco | Radish | Chinese | Brassica oleracea | Rape |
|---|---|---|---|---|---|
| Cabbage | |||||
| Count probes | 7.47 ± 0.72 ab | 5.13 ± 1.30 b | 6.87 ± 1.21 ab | 10.13 ± 1.97 a | 6.87 ± 0.94 ab |
| Sum time of np wave (min) | 41.93 ± 6.80 b | 62.29 ± 16.80 ab | 106.16 ± 24.24 a | 93.09 ± 13.61 a | 74.06 ± 13.63 ab |
| Time to 1st probe (min) | 3.69 ± 1.43 b | 2.02 ± 1.08 b | 10.87 ± 3.06 a | 6.89 ± 2.62 ab | 5.89 ± 3.55 ab |
| Duration of 1st probe (min) | 42.02 ± 7.58 | 58.31 ± 10.75 | 46.16 ± 11.24 | 31.70 ± 7.40 | 54.28 ± 14.24 |
| Number of probes to the 1st E1 | 5.53 ± 0.94 | 4.87 ± 0.89 | 6.00 ± 0.97 | 6.93 ± 0.96 | 5.8 ± 0.72 |
| Number of pd | 89.93 ± 8.34 ab | 68.87 ± 15.08 b | 81.67 ± 11.36 b | 121.93 ± 15.87 a | 71.4 ± 8.14 b |
| Mean duration of pd (s) | 5.14 ± 0.30 | 5.64 ± 0.25 | 5.39 ± 0.27 | 5.74 ± 0.24 | 5.80 ± 0.23 |
| Number of short probes C (<3 min) | 3.93 ± 0.51 | 3.07 ± 0.59 | 2.87 ± 0.61 | 4.73 ± 1.08 | 3.67 ± 0.73 |
| Total duration of C (min) | 133.06 ± 12.73 a | 87.12 ± 10.05 b | 103.13± 16.07 ab | 98.75 ± 8.84 b | 107.78 ± 9.25 ab |
| Number of G | 0.8 ± 0.17 | 0.93 ± 0.27 | 1.20 ± 0.30 | 1.00 ± 0.26 | 1.33 ± 0.30 |
| Duration of G (min) | 15.35 ± 2.86 | 16.12 ± 3.23 | 20.01 ± 3.44 | 23.87 ± 4.38 | 22.48 ± 3.20 |
| Number of F | 0.67 ± 0.19 b | 0.6 ± 0.19 b | 1.93 ± 0.38 a | 2.00 ± 0.35 a | 1.87 ± 0.32 a |
| Duration of F (min) | 13.91 ± 4.09 b | 17.46 ± 3.43 b | 40.67 ± 7.51 a | 38.08 ± 6.94 a | 34.99 ± 5.50 a |
| EPG Parameter | Tobacco | Radish | Chinese | Brassica oleracea | Rape |
|---|---|---|---|---|---|
| Cabbage | |||||
| Number of E1 | 1.07 ± 0.27 b | 1.33 ± 0.35 ab | 2.40 ± 0.42 a | 1.87 ± 0.42 ab | 1.73 ± 0.57 ab |
| Total duration of E1 | 7.61 ± 1.68 b | 12.64 ± 2.39 ab | 18.11 ± 3.09 a | 15.19 ± 3.33 ab | 14.58 ± 2.85 ab |
| Number of E2 | 1.87 ± 0.29 | 1.53 ± 0.31 | 1.27 ± 0.25 | 1.47 ± 0.26 | 1.60 ± 0.43 |
| Time from 1st probe to 1st E2 (min) | 65.75 ± 8.84 b | 71.04 ± 12.03 b | 118.07 ± 14.75 a | 85.34 ± 16.43 ab | 96.43 ± 13.41 ab |
| Number of sustained E2 (>10 min) | 1.27 ± 0.21 | 1.07 ± 0.21 | 0.8 ± 0.2 | 1.00 ± 0.17 | 1.07 ± 0.28 |
| Total duration of E2 (min) | 148.17 ± 11.49 a | 164.39 ± 11.15 a | 71.51 ± 12.68 b | 91.10 ± 11.87 b | 106.04 ± 15.24 b |
| Stage | Tobacco | Radish | Chinese Cabbage | Brassica oleracea | Rape |
|---|---|---|---|---|---|
| First instar (N1) | 1.51 ± 0.11 bc | 1.57 ± 0.05 abc | 1.76 ± 0.07 a | 1.74 ± 0.05 ab | 1.48 ± 0.05 c |
| Second instar (N2) | 1.57 ± 0.09 c | 2.06 ± 0.05 b | 1.65 ± 0.08 c | 1.92 ± 0.06 bc | 2.20 ± 0.05 a |
| Third instar (N3) | 1.57 ± 0.08 b | 2.17 ± 0.05 a | 1.67 ± 0.07 b | 2.01 ± 0.04 a | 2.15 ± 0.04 a |
| Fourth instar (N4) | 1.27 ± 0.07 bc | 1.14 ± 0.06 bc | 1.51 ± 0.08 a | 1.10 ± 0.06 c | 1.34 ± 0.09 ab |
| Preadult duration | 5.85 ± 0.18 c | 6.89 ± 0.08 ab | 6.56 ± 0.14 b | 6.77 ± 0.06 ab | 7.18 ± 0.11 a |
| Adult longevity | 14.52 ± 0.48 c | 19.16 ± 0.37 a | 17.58 ± 0.49 b | 19.12 ± 0.36 a | 19.22 ± 0.50 a |
| Mean longevity | 20.34 ± 0.51 c | 26.02 ± 0.40 a | 24.07 ± 0.50 b | 25.63 ± 0.41 a | 26.21 ± 0.59 a |
| APOP | 0.66 ± 0.05 b | 0.68 ± 0.04 b | 1.20 ± 0.13 a | 0.76 ± 0.06 b | 0.88 ± 0.08 b |
| TPOP | 6.46 ± 0.17 c | 7.57 ± 0.08 b | 7.76 ± 0.16 ab | 7.53 ± 0.10 b | 8.05 ± 013 a |
| Reproduction period | 11.90 ± 0.31 c | 16.55 ± 0.45 a | 14.91 ± 0.45 b | 15.81 ± 0.50 ab | 15.07 ± 0.35 b |
| Mean fecundity | 40.67 ± 2.58 bc | 53.73 ± 1.95 a | 36.52 ± 2.74 c | 47.24 ± 1.91 b | 50.78 ± 2.53 a |
| Parameter | Tobacco | Radish | Chinese Cabbage | Brassica oleracea | Rape |
|---|---|---|---|---|---|
| r (d−1) | 0.3918 ± 0.0092 a | 0.3203 ± 0.0065 b | 0.3224 ± 0.0093 b | 0.3191 ± 0.0059 b | 0.3066 ± 0.0082 b |
| λ (d−1) | 1.4797 ± 0.0136 a | 1.3775 ± 0.0089 b | 1.3805 ± 0.0129 b | 1.3760 ± 0.0080 b | 1.3589 ± 0.0111 b |
| R0 (offspring/individual) | 38.43 ± 2.71 bc | 47.30 ± 3.17 a | 35.16± 2.72 c | 44.63 ± 2.54 ab | 44.71 ± 3.37 ab |
| T (d) | 9.31 ± 0.22 d | 12.04 ± 0.09 ab | 11.04 ± 0.21 c | 11.90 ± 0.10 b | 12.39 ± 0.17 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, W.; Cheng, Q.; Lu, C.; Chen, W.; Zhao, D.; He, Y. Different Host Plants Distinctly Influence the Adaptability of Myzus persicae (Hemiptera: Aphididae). Agriculture 2022, 12, 2162. https://doi.org/10.3390/agriculture12122162
Jiang W, Cheng Q, Lu C, Chen W, Zhao D, He Y. Different Host Plants Distinctly Influence the Adaptability of Myzus persicae (Hemiptera: Aphididae). Agriculture. 2022; 12(12):2162. https://doi.org/10.3390/agriculture12122162
Chicago/Turabian StyleJiang, Wenbin, Qian Cheng, Changhao Lu, Wenlong Chen, Degang Zhao, and Yingqin He. 2022. "Different Host Plants Distinctly Influence the Adaptability of Myzus persicae (Hemiptera: Aphididae)" Agriculture 12, no. 12: 2162. https://doi.org/10.3390/agriculture12122162