
Sustainable Bioconversion of Wetland Plant Biomass for Pleurotus ostreatus var. florida Cultivation: Studies on Proximate and Biochemical Characterization

 , ,
, ,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
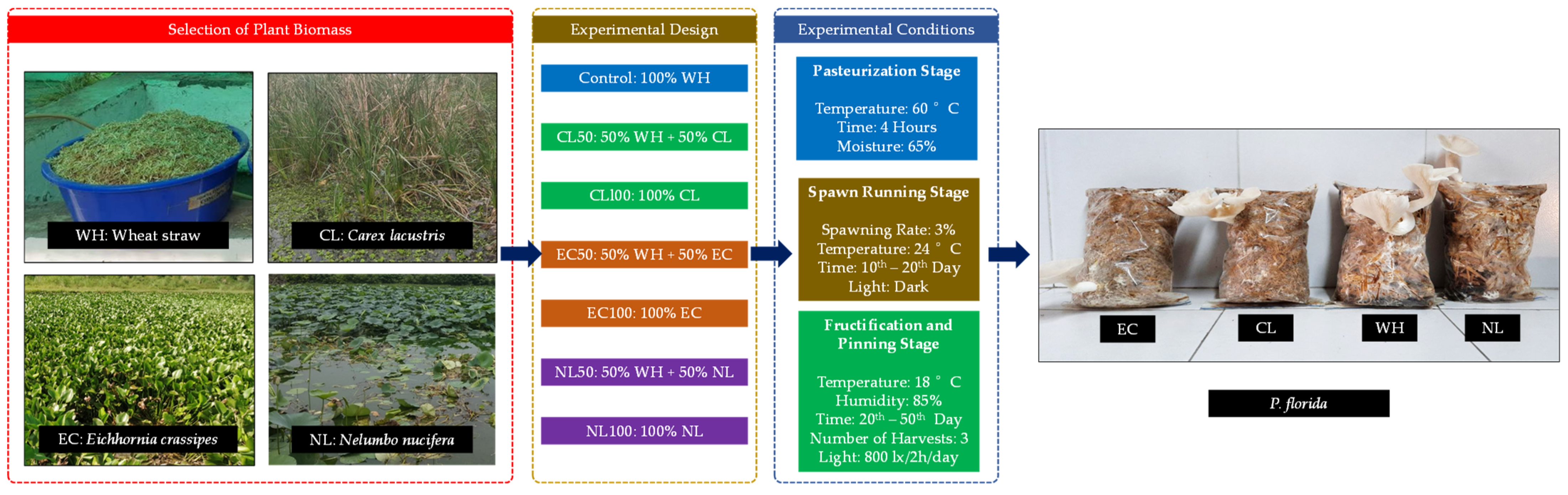
2.1. Collection of Wetland Plant Biomass and Mushroom Spawn
2.2. Experimental Design and Conditions
2.3. Analytical Methods
2.4. Data Analysis
2.5. Software and Tools
3. Results and Discussion
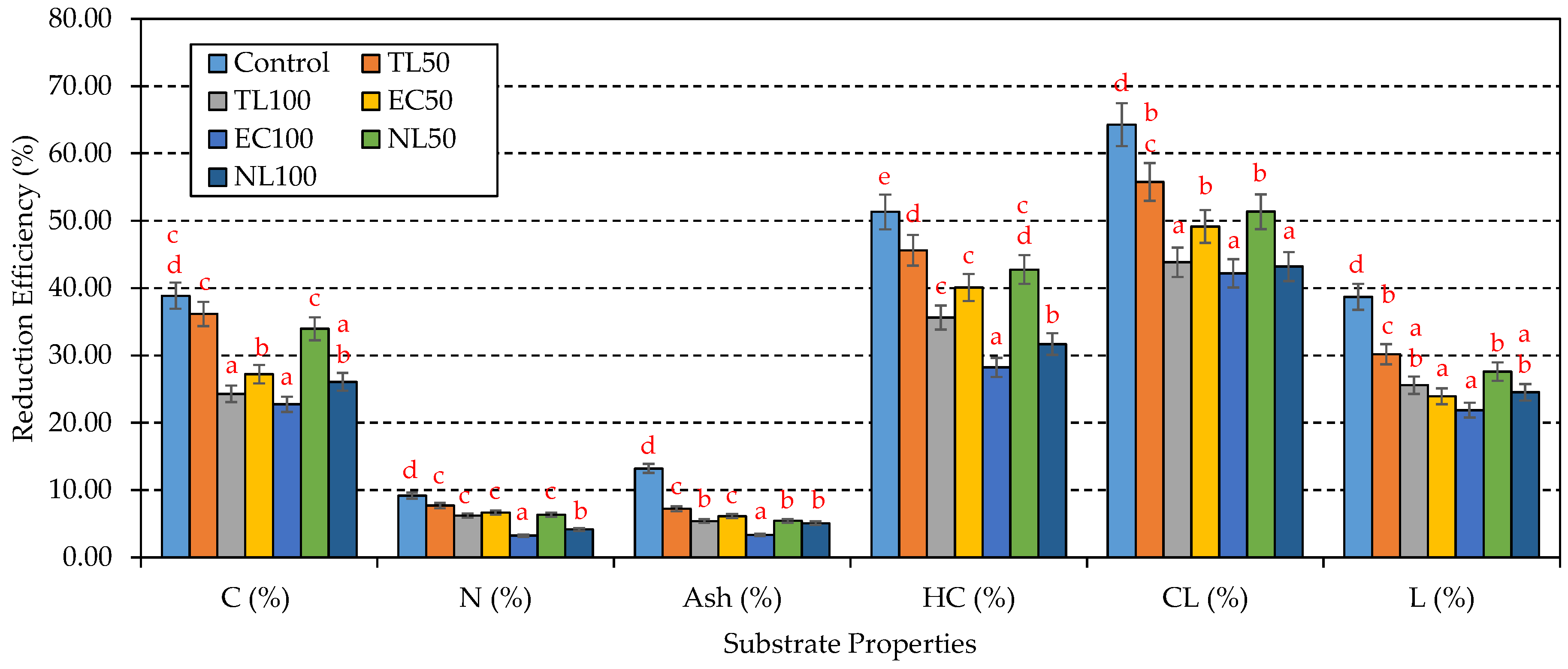
3.1. Changes in Substrate Properties before and after P. florida Cultivation
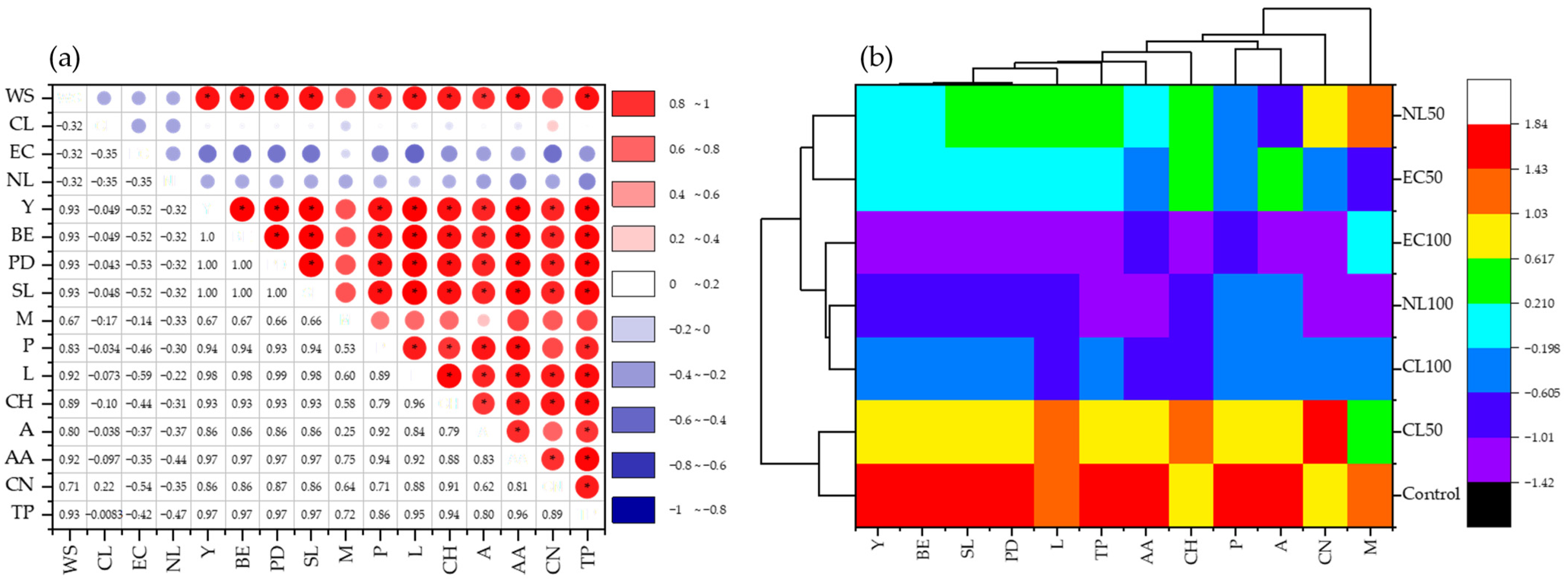
3.2. Effect of Wetland Plant Biomass on Growth and Yield of P. florida
3.3. Effect of Wetland Plant Biomass on Nutrient and Biochemical Composition of P. florida
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hondula, K.L.; Jones, C.N.; Palmer, M.A. Effects of Seasonal Inundation on Methane Fluxes from Forested Freshwater Wetlands. Environ. Res. Lett. 2021, 16, 084016. [Google Scholar] [CrossRef]
- Kao-Kniffin, J.; Freyre, D.S.; Balser, T.C. Methane Dynamics across Wetland Plant Species. Aquat. Bot. 2010, 93, 107–113. [Google Scholar] [CrossRef]
- Robertson, G.P.; Grace, P.R. Greenhouse Gas Fluxes in Tropical and Temperate Agriculture: The Need for a Full-Cost Accounting of Global Warming Potentials. In Tropical Agriculture in Transition—Opportunities for Mitigating Greenhouse Gas Emissions? Springer: Dordrecht, The Netherlands, 2004; pp. 51–63. [Google Scholar]
- Zhao, J.; Liu, C.; Li, H.; Liu, J.; Jiang, T.; Yan, D.; Tong, J.; Dong, L. Review on Ecological Response of Aquatic Plants to Balanced Harvesting. Sustainability 2022, 14, 12451. [Google Scholar] [CrossRef]
- Saeed, T.; Sun, G. A Review on Nitrogen and Organics Removal Mechanisms in Subsurface Flow Constructed Wetlands: Dependency on Environmental Parameters, Operating Conditions and Supporting Media. J. Environ. Manag. 2012, 112, 429–448. [Google Scholar] [CrossRef] [PubMed]
- Ceschin, S.; Abati, S.; Traversetti, L.; Spani, F.; del Grosso, F.; Scalici, M. Effects of the Invasive Duckweed Lemna minuta on Aquatic Animals: Evidence from an Indoor Experiment. Plant Biosyst. 2019, 153, 749–755. [Google Scholar] [CrossRef]
- Kumar, V.; Kumar, P.; Eid, E.M.; Singh, J.; Adelodun, B.; Kumar, P.; Kumari, S.; Choi, K.S. Modeling of Water Hyacinth Growth and Its Role in Heavy Metals Accumulation from Unoperated Old Ganga Canal at Haridwar, India. Rend. Lincei 2021, 32, 805–816. [Google Scholar] [CrossRef]
- Laterra, P.; Booman, G.C.; Picone, L.; Videla, C.; Orúe, M.E. Indicators of Nutrient Removal Efficiency for Riverine Wetlands in Agricultural Landscapes of Argentine Pampas. J. Environ. Manag. 2018, 222, 148–154. [Google Scholar] [CrossRef]
- Hultberg, M.; Prade, T.; Bodin, H.; Vidakovic, A.; Asp, H. Adding Benefit to Wetlands—Valorization of Harvested Common Reed through Mushroom Production. Sci. Total Environ. 2018, 637–638, 1395–1399. [Google Scholar] [CrossRef]
- Bandopadhyay Mukhopadhyay, S. Oyster Mushroom Cultivation on Water Hyacinth Biomass: Assessment of Yield Performances, Nutrient, and Toxic Element Contents of Mushrooms. In An Introduction to Mushroom; IntechOpen: London, UK, 2020. [Google Scholar]
- Shirur, M.; Ahlawat, O.P.; Manikandan, K. Mushroom Consumption and Purchasing Behaviour in India: A Study among Selected Respondents. Mushroom Res. 2014, 23, 225–231. [Google Scholar]
- Sharma, V.P.; Annepu, S.K.; Gautam, Y.; Singh, M.; Kamal, S. Status of Mushroom Production in India. Mushroom Res. 2017, 26, 111–120. [Google Scholar]
- Raman, J.; Lee, S.-K.; Im, J.-H.; Oh, M.-J.; Oh, Y.-L.; Jang, K.-Y. Current Prospects of Mushroom Production and Industrial Growth in India. J. Mushrooms 2018, 4, 239–249. [Google Scholar]
- Thakur, M.P. Advances in Mushroom Production: Key to Food, Nutritional and Employment Security: A Review. Indian Phytopathol. 2020, 73, 377–395. [Google Scholar] [CrossRef]
- Gupta, S.; Summuna, B.; Gupta, M.; Annepu, S.K. Edible Mushrooms: Cultivation, Bioactive Molecules, and Health Benefits. In Bioactive Molecules in Food, 1st ed.; Springer: Cham, Switzerland, 2019; pp. 1815–1847. [Google Scholar] [CrossRef]
- Josephine, M. Solid Waste Management through Mushroom Cultivation—An Eco Friendly Approach. Int. J. Environ. Ecol. Eng. 2014, 8, 115–117. [Google Scholar]
- Shirur, M.; Barh, A.; Annepu, S.K. Sustainable Production of Edible and Medicinal Mushrooms: Implications on Mushroom Consumption. In Climate Change and Resilient Food Systems; Springer: Singapore, 2021; pp. 315–346. [Google Scholar]
- Showkat, Q.A.; Rather, J.A.; Abida, J.; Dar, B.N.; Makroo, H.A.; Majid, D. Bioactive Components, Physicochemical and Starch Characteristics of Different Parts of Lotus (Nelumbo nucifera Gaertn.) Plant: A Review. Int. J. Food Sci. Technol. 2021, 56, 2205–2214. [Google Scholar] [CrossRef]
- Kumar, P.; Eid, E.M.; Al-Huqail, A.A.; Širić, I.; Adelodun, B.; Abou Fayssal, S.; Valadez-Blanco, R.; Goala, M.; Ajibade, F.O.; Choi, K.S.; et al. Kinetic Studies on Delignification and Heavy Metals Uptake by Shiitake (Lentinula edodes) Mushroom Cultivated on Agro-Industrial Wastes. Horticulturae 2022, 8, 316. [Google Scholar] [CrossRef]
- Kurtzman, R.H., Jr. Pateurization of mushroom substrate and other solids. Afr. J. Environ. Sci. 2010, 4, 936–941. [Google Scholar]
- Kumar, P.; Kumar, V.; Adelodun, B.; Bedeković, D.; Kos, I.; Širić, I.; Alamri, S.A.M.; Alrumman, S.A.; Eid, E.M.; Abou Fayssal, S.; et al. Sustainable Use of Sewage Sludge as a Casing Material for Button Mushroom (Agaricus bisporus) Cultivation: Experimental and Prediction Modeling Studies for Uptake of Metal Elements. J. Fungi 2022, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Abou Fayssal, S.; Alsanad, M.A.; Yordanova, M.H.; el Sebaaly, Z.; Najjar, R.; Sassine, Y.N. Effect of Olive Pruning Residues on Substrate Temperature and Production of Oyster Mushroom (Pleurotus ostreatus). Acta Hortic. 2021, 1327, 245–252. [Google Scholar] [CrossRef]
- AOAC Official Methods of Analysis, 21st Edition (2019)—AOAC International. Available online: https://www.aoac.org/official-methods-of-analysis-21st-edition-2019/ (accessed on 17 September 2020).
- Kumar, V.; Valadez-Blanco, R.; Kumar, P.; Singh, J.; Kumar, P. Effects of Treated Sugar Mill Effluent and Rice Straw on Substrate Properties under Milky Mushroom (Calocybe indica P&C) Production: Nutrient Utilization and Growth Kinetics Studies. Environ. Technol. Innov. 2020, 19, 101041. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An Examination of the Degtjareff Method for Determining Soil Organic Matter and a Proposed Modification of the Chromic Acid Titration Method. Soil Sci. 1933, 37, 29–38. [Google Scholar] [CrossRef]
- Chromý, V.; Vinklárková, B.; Šprongl, L.; Bittová, M. The Kjeldahl Method as a Primary Reference Procedure for Total Protein in Certified Reference Materials Used in Clinical Chemistry. I. A Review of Kjeldahl Methods Adopted by Laboratory Medicine. Crit. Rev. Anal. Chem. 2015, 45, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Gessner, M.O. Proximate Lignin and Cellulose. In Methods to Study Litter Decomposition: A Practical Guide; Springer: Berlin/Heidelberg, Germany, 2005; pp. 115–120. ISBN 9781402034664. [Google Scholar]
- Lalotra, P.; Bala, P.; Kumar, S.; Sharma, Y.P. Biochemical Characterization of Some Wild Edible Mushrooms from Jammu and Kashmir. Proc. Natl. Acad. Sci. India Sect. B-Biol. Sci. 2018, 88, 539–545. [Google Scholar] [CrossRef]
- Alam, N.; Khan, A.; Hossain, S.; Amin, R.; Khan, L.A. Nutritional Analysis of Dietary Mushroom- Pleurotus florida Eger and Pleurotus sajor-caju (Fr.) Singer. Bangladesh J. Mushroom 2007, 1, 1–7. [Google Scholar]
- Kumar, P.; Kumar, V.; Goala, M.; Singh, J.; Kumar, P. Integrated use of treated dairy wastewater and agro-residue for Agaricus bisporus mushroom cultivation: Experimental and kinetics studies. Biocatal. Agric. Biotechnol. 2021, 32, 101940. [Google Scholar] [CrossRef]
- Abou Fayssal, S.; Alsanad, M.A.; el Sebaaly, Z.; Ismail, A.I.H.; Sassine, Y.N. Valorization of Olive Pruning Residues through Bioconversion into Edible Mushroom Pleurotus ostreatus (Jacq. Ex Fr.) P. Kumm. (1871) of Improved Nutritional Value. Scientifica 2020, 2020, 39503. [Google Scholar] [CrossRef]
- Kapahi, M.; Sachdeva, S. Mycoremediation Potential of Pleurotus Species for Heavy Metals: A Review. Bioresour. Bioprocess. 2017, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Öztürk, C.; Atila, F. Changes in Lignocellulosic Fractions of Growing Substrates during the Cultivation of Hypsizygus ulmarius Mushroom and Its Effects on Mushroom Productivity. Sci. Hortic. 2021, 288, 110403. [Google Scholar] [CrossRef]
- Hoa, H.T.; Wang, C.L.; Wang, C.H. The Effects of Different Substrates on the Growth, Yield, and Nutritional Composition of Two Oyster Mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology 2015, 43, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Werghemmi, W.; Abou Fayssal, S.; Mazouz, H.; Hajjaj, H.; Hajji, L. Olive and Green Tea Leaves Extract in Pleurotus ostreatus var. florida Culture Media: Effect on Mycelial Linear Growth Rate, Diameter and Growth Induction Index. IOP Conf. Ser. Earth Environ. Sci. 2022, 1090, 012020. [Google Scholar] [CrossRef]
- Sassine, Y.N.; Naim, L.; el Sebaaly, Z.; Abou Fayssal, S.; Alsanad, M.A.; Yordanova, M.H. Nano Urea Effects on Pleurotus ostreatus Nutritional Value Depending on the Dose and Timing of Application. Sci. Rep. 2021, 11, 5588. [Google Scholar] [CrossRef]
- Abou Fayssal, S.; el Sebaaly, Z.; Alsanad, M.A.; Najjar, R.; Bohme, M.; Yordanova, M.H.; Sassine, Y.N. Combined Effect of Olive Pruning Residues and Spent Coffee Grounds on Pleurotus ostreatus Production, Composition, and Nutritional Value. PLoS ONE 2021, 16, e0255794. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kumar, V.; Eid, E.M.; AL-Huqail, A.A.; Adelodun, B.; Abou Fayssal, S.; Goala, M.; Arya, A.K.; Bachheti, A.; Andabaka, Ž.; et al. Spatial Assessment of Potentially Toxic Elements (PTE) Concentration in Agaricus bisporus Mushroom Collected from Local Vegetable Markets of Uttarakhand State, India. J. Fungi 2022, 8, 452. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Hansen, M.; Hobley, T.J.; Jensen, P.R. Valorization of Green Biomass: Alfalfa Pulp as a Substrate for Oyster Mushroom Cultivation. Foods 2022, 11, 2519. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, M.; Singh, G. Effect of Different Pretreatments on the Quality of Mushrooms during Solar Drying. J. Food Sci. Technol. 2013, 50, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Piskov, S.; Timchenko, L.; Avanesyan, S.; Siddiqui, S.A.; Sizonenko, M.; Kurchenko, V.; Rzhepakovsky, I.; Blinov, A.; Nagdalian, A.; Shariati, M.A.; et al. A Comparative Study on the Structural Properties and Lipid Profile of Mushroom (Pleurotus ostreatus) Powder Obtained by Different Drying Methods. Agriculture 2022, 12, 1590. [Google Scholar] [CrossRef]
- Gao, S.; Huang, Z.; Feng, X.; Bian, Y.; Huang, W.; Liu, Y. Bioconversion of Rice Straw Agro-Residues by Lentinula edodes and Evaluation of Non-Volatile Taste Compounds in Mushrooms. Sci. Rep. 2020, 10, 1814–1818. [Google Scholar] [CrossRef] [Green Version]
- Tolera, K.D.; Abera, S. Nutritional Quality of Oyster Mushroom (Pleurotus ostreatus) as Affected by Osmotic Pretreatments and Drying Methods. Food Sci. Nutr. 2017, 5, 989–996. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [Green Version]
- Kozarski, M.; Klaus, A.; Jakovljevic, D.; Todorovic, N.; Vunduk, J.; Petrović, P.; Niksic, M.; Vrvic, M.M.; van Griensven, L. Antioxidants of Edible Mushrooms. Molecules 2015, 20, 19489–19525. [Google Scholar] [CrossRef] [Green Version]
- Soobrattee, M.A.; Neergheen, V.S.; Luximon-Ramma, A.; Aruoma, O.I.; Bahorun, T. Phenolics as Potential Antioxidant Therapeutic Agents: Mechanism and Actions. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2005, 579, 200–213. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Mushroom Substrate Properties | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| pH | EC (dS/m) | C (%) | N (%) | C/N | Ash (%) | H.C.L. (%) | CL (%) | L (%) | ||
| Control | Before | 7.01 ± 0.01 a | 3.01 ± 0.03 a | 48.10 ± 1.90 a | 1.31 ± 0.01 a | 36.72 | 2.12 ± 0.03 a | 24.20 ± 0.56 a | 45.37 ± 2.74 a | 9.07 ± 0.03 a |
| After | 5.10 ± 0.03 b | 2.07 ± 0.05 b | 29.40 ± 2.02 b | 1.19 ± 0.03 b | 24.71 | 1.84 ± 0.04 b | 11.78 ± 1.78 b | 16.21 ± 3.06 b | 5.56 ± 0.09 b | |
| CL50 | Before | 6.96 ± 0.03 a | 2.79 ± 0.06 a | 47.23 ± 0.95 a | 1.30 ± 0.02 a | 36.33 | 2.08 ± 0.02 a | 22.99 ± 1.05 a | 43.69 ± 1.59 a | 8.74 ± 0.12 a |
| After | 5.13 ± 0.02 b | 2.09 ± 0.10 b | 30.15 ± 1.64 b | 1.20 ± 0.01 b | 25.13 | 1.93 ± 0.03 b | 12.50 ± 2.66 b | 19.32 ± 2.20 b | 6.10 ± 0.07 b | |
| CL100 | Before | 6.90 ± 0.03 a | 2.56 ± 0.05 a | 46.35 ± 1.31 a | 1.29 ± 0.02 a | 35.93 | 2.04 ± 0.01 a | 21.78 ± 0.97 a | 42.01 ± 1.11 a | 8.40 ± 0.05 a |
| After | 5.87 ± 0.04 b | 2.12 ± 0.09 b | 35.09 ± 1.05 b | 1.21 ± 0.01 b | 29.00 | 1.93 ± 0.02 b | 14.02 ± 1.32 b | 23.58 ± 3.01 b | 6.25 ± 0.19 b | |
| EC50 | Before | 6.76 ± 0.01 a | 2.91 ± 0.08 a | 38.59 ± 2.10 a | 1.28 ± 0.01 a | 30.27 | 1.96 ± 0.04 a | 21.62 ± 0.63 a | 38.97 ± 2.48 a | 8.99 ± 0.08 a |
| After | 5.49 ± 0.02 b | 2.25 ± 0.03 b | 28.09 ± 1.47 b | 1.19 ± 0.02 b | 23.61 | 1.84 ± 0.03 b | 12.95 ± 1.70 b | 19.81 ± 0.97 b | 6.84 ± 0.10 b | |
| EC100 | Before | 6.50 ± 0.03 a | 2.81 ± 0.04 a | 29.08 ± 0.89 a | 1.24 ± 0.02 a | 23.45 | 1.80 ± 0.02 a | 19.03 ± 1.07 a | 32.57 ± 1.76 a | 8.91 ± 0.04 a |
| After | 5.75 ± 0.05 b | 2.34 ± 0.07 b | 22.47 ± 0.52 b | 1.20 ± 0.01 b | 18.73 | 1.74 ± 0.02 b | 13.66 ± 0.81 b | 18.82 ± 2.55 b | 6.96 ± 0.11 b | |
| NL50 | Before | 6.92 ± 0.02 a | 2.87 ± 0.10 a | 41.02 ± 1.73 a | 1.26 ± 0.01 a | 32.56 | 1.85 ± 0.01 a | 23.75 ± 1.25 a | 41.25 ± 2.02 a | 8.80 ± 0.09 a |
| After | 5.12 ± 0.01 b | 2.18 ± 0.05 b | 27.09 ± 0.86 b | 1.18 ± 0.03 b | 22.96 | 1.75 ± 0.02 b | 13.59 ± 2.04 b | 20.06 ± 1.17 b | 6.37 ± 0.03 b | |
| NL100 | Before | 6.82 ± 0.04 a | 2.73 ± 0.04 a | 33.94 ± 1.24 a | 1.21 ± 0.02 a | 28.05 | 1.58 ± 0.03 a | 23.29 ± 1.42 a | 37.13 ± 2.35 a | 8.52 ± 0.10 a |
| After | 5.78 ± 0.03 b | 2.22 ± 0.06 b | 25.09 ± 0.92 b | 1.16 ± 0.02 b | 21.63 | 1.50 ± 0.01 b | 15.91 ± 0.63 b | 21.09 ± 1.60 b | 6.43 ± 0.04 b | |
| Treatment | Colonization (Days) | Yield (g/kg Fresh Substrate) | Biological Efficiency (%) | Pileus Diameter (cm) | Stipe Length (cm) | |||
|---|---|---|---|---|---|---|---|---|
| First Flush | Second Flush | Third Flush | Total | |||||
| Control | 12.00 ± 1.00 a | 115.46 ± 4.08 e | 93.55 ± 2.77 e | 45.05 ± 1.50 d | 254.06 ± 6.82 f | 72.59 ± 0.80 e | 7.30 ± 0.04 f | 6.39 ± 0.07 e |
| CL50 | 13.00 ± 1.00 a | 95.10 ± 3.57 d | 86.81 ± 4.23 e | 42.18 ± 2.74 d | 224.17 ± 8.10 e | 64.05 ± 1.47 d | 6.79 ± 0.07 e | 5.75 ± 0.05 d |
| CL100 | 18.00 ± 2.00 c | 70.22 ± 5.16 a | 56.63 ± 2.06 b | 30.08 ± 0.98 a | 156.96 ± 5.17 ab | 44.85 ± 1.82 b | 4.66 ± 0.03 b | 4.22 ± 0.18 b |
| EC50 | 15.00 ± 1.00 b | 77.36 ± 2.09 b | 64.09 ± 3.42 c | 34.10 ± 1.26 b | 175.45 ± 7.32 c | 50.13 ± 3.25 c | 5.30 ± 0.10 c | 4.42 ± 0.09 b |
| EC100 | 19.00 ± 2.00 c | 61.09 ± 4.82 a | 46.32 ± 1.85 a | 24.67 ± 2.30 a | 132.09 ± 9.51 a | 37.74 ± 0.78 a | 4.20 ± 0.18 a | 3.46 ± 0.13 a |
| NL50 | 14.00 ± 1.00 b | 82.24 ± 2.70 c | 70.40 ± 2.57 d | 39.30 ± 1.10 c | 191.99 ± 4.08 d | 54.85 ± 1.35 c | 5.82 ± 0.05 d | 4.75 ± 0.06 c |
| NL100 | 14.00 ± 1.00 b | 67.02 ± 3.56 a | 51.06 ± 3.14 a | 26.50 ± 3.12 a | 144.58 ± 8.66 a | 41.31 ± 2.92 b | 4.38 ± 0.12 a | 3.64 ± 0.20 a |
| Treatment | Proximate Constituents (%) | Biochemical Constituents | ||||||
|---|---|---|---|---|---|---|---|---|
| Moisture | Protein | Lipid | Carbohydrates | Ash | Ascorbic Acid (µg/100 g) | Carotenoids (µg/100 g) | Total Phenol (mg/100 g) | |
| Control | 86.30 ± 2.10 ab | 25.14 ± 0.22 c | 2.36 ± 0.05 b | 48.70 ± 0.40 cd | 7.52 ± 0.04 b | 0.49 ± 0.01 d | 0.23 ± 0.01 b | 6.12 ± 0.03 b |
| CL50 | 85.66 ± 1.98 a | 24.42 ± 0.24 c | 2.35 ± 0.05 b | 49.02 ± 0.21 d | 7.48 ± 0.05 b | 0.47 ± 0.02 cd | 0.24 ± 0.01 b | 6.10 ± 0.04 b |
| CL100 | 85.10 ± 2.06 a | 23.15 ± 0.17 b | 2.30 ± 0.06 a | 46.20 ± 0.19 b | 7.38 ± 0.10 a | 0.43 ± 0.02 b | 0.22 ± 0.01 a | 6.05 ± 0.01 a |
| EC50 | 84.90 ± 1.55 a | 23.23 ± 0.30 b | 2.32 ± 0.04 ab | 47.81 ± 0.28 c | 7.44 ± 0.05 ab | 0.44 ± 0.01 c | 0.22 ± 0.02 a | 6.07 ± 0.04 ab |
| EC100 | 85.53 ± 2.91 a | 22.90 ± 0.13 a | 2.28 ± 0.06 a | 45.75 ± 0.13 a | 7.35 ± 0.07 a | 0.43 ± 0.02 b | 0.21 ± 0.01 a | 6.03 ± 0.02 a |
| NL50 | 86.37 ± 1.72 ab | 23.28 ± 0.25 b | 2.33 ± 0.07 ab | 47.94 ± 0.36 c | 7.36 ± 0.03 a | 0.45 ± 0.01 c | 0.23 ± 0.01 ab | 6.08 ± 0.04 ab |
| NL100 | 84.48 ± 1.05 a | 23.20 ± 0.18 b | 2.30 ± 0.04 a | 46.09 ± 0.09 b | 7.39 ± 0.06 a | 0.42 ± 0.02 a | 0.21 ± 0.01 a | 6.02 ± 0.02 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elbagory, M.; El-Nahrawy, S.; Omara, A.E.-D.; Eid, E.M.; Bachheti, A.; Kumar, P.; Abou Fayssal, S.; Adelodun, B.; Bachheti, R.K.; Kumar, P.; et al. Sustainable Bioconversion of Wetland Plant Biomass for Pleurotus ostreatus var. florida Cultivation: Studies on Proximate and Biochemical Characterization. Agriculture 2022, 12, 2095. https://doi.org/10.3390/agriculture12122095
Elbagory M, El-Nahrawy S, Omara AE-D, Eid EM, Bachheti A, Kumar P, Abou Fayssal S, Adelodun B, Bachheti RK, Kumar P, et al. Sustainable Bioconversion of Wetland Plant Biomass for Pleurotus ostreatus var. florida Cultivation: Studies on Proximate and Biochemical Characterization. Agriculture. 2022; 12(12):2095. https://doi.org/10.3390/agriculture12122095
Chicago/Turabian StyleElbagory, Mohssen, Sahar El-Nahrawy, Alaa El-Dein Omara, Ebrahem M. Eid, Archana Bachheti, Pankaj Kumar, Sami Abou Fayssal, Bashir Adelodun, Rakesh Kumar Bachheti, Pankaj Kumar, and et al. 2022. "Sustainable Bioconversion of Wetland Plant Biomass for Pleurotus ostreatus var. florida Cultivation: Studies on Proximate and Biochemical Characterization" Agriculture 12, no. 12: 2095. https://doi.org/10.3390/agriculture12122095