
Complementary Strategies for Biological Control of Aphids and Related Virus Transmission in Sugar Beet to Replace Neonicotinoids

 ,
, 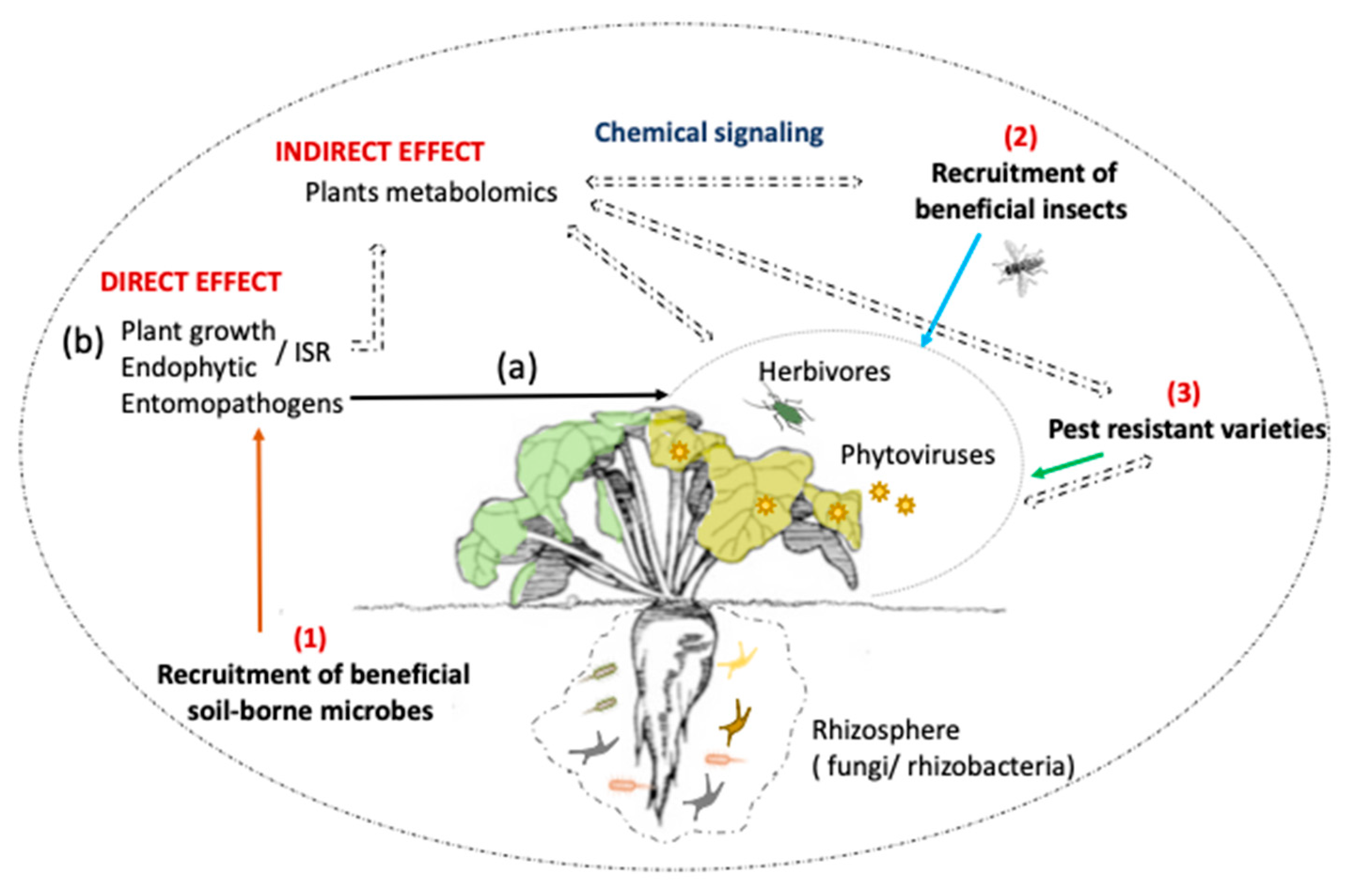{kind=link}
Abstract
:1. Introduction
2. Manipulation of Virus Processes
3. Cross-Protection in Multi-Infections
4. Breeding for Resistant Varieties
5. Selection of Resistant Varieties
6. Use of Semiochemicals
7. Entomopathogenic Fungi, a Biorational Control Agent with Wide Spectrum of Activity
8. Rhizobacteria and Plant Defenses
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Biancardi, E.; Mcgrath, J.M.; Panella, L.W.; Lewellen, R.T.; Stevanato, P. Sugar Beet. In Root and Tuber Crops; Bradshaw, J.E., Ed.; Springer: New York, NY, USA, 2010; pp. 173–219. [Google Scholar]
- Stevens, M.; Patron, N.J.; Dolby, C.A.; Weekes, R.; Hallsworth, P.B.; Lemaire, O.; Smith, H.G. Distribution and Properties of Geographically Distinct Isolates of Sugar Beet Yellowing Viruses. Plant Pathol. 2005, 54, 100–107. [Google Scholar] [CrossRef]
- Hossain, R.; Menzel, W.; Lachmann, C.; Varrelmann, M. New Insights into Virus Yellows Distribution in Europe and Effects of Beet Yellows Virus, Beet Mild Yellowing Virus, and Beet Chlorosis Virus on Sugar Beet Yield Following Field Inoculation. Plant Pathol. 2021, 70, 584–593. [Google Scholar] [CrossRef]
- Stevens, M.; Hallsworth, P.B.; Smith, H.G. The Effects of Beet Mild Yellowing Virus and Beet Chlorosis Virus on the Yield of UK Field-Grown Sugar Beet in 1997, 1999 and 2000. Ann. Appl. Biol. 2004, 144, 113–119. [Google Scholar] [CrossRef]
- Elbert, A.; Haas, M.; Springer, B.; Thielert, W.; Nauen, R. Applied Aspects of Neonicotinoid Uses in Crop Protection. Pest Manag. Sci. 2008, 64, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.; Bub, S.; Petschick, L.L.; Stehle, S.; Wolfram, J. Applied Pesticide Toxicity Shifts toward Plants and Invertebrates, Even in GM Crops. Science 2021, 372, 81–84. [Google Scholar] [CrossRef]
- Hauer, M.; Hansen, A.L.; Manderyck, B.; Olsson, Å.; Raaijmakers, E.; Hanse, B.; Stockfisch, N.; Märländer, B. Neonicotinoids in Sugar Beet Cultivation in Central and Northern Europe: Efficacy and Environmental Impact of Neonicotinoid Seed Treatments and Alternative Measures. Crop Prot. 2017, 93, 132–142. [Google Scholar] [CrossRef]
- Goulson, D. Review: An Overview of the Environmental Risks Posed by Neonicotinoid Insecticides. J. Appl. Ecol. 2013, 50, 977–987. [Google Scholar] [CrossRef]
- Stuligross, C.; Williams, N.M. Past Insecticide Exposure Reduces Bee Reproduction and Population Growth Rate. Proc. Natl. Acad. Sci. USA 2021, 118, e2109909118. [Google Scholar] [CrossRef]
- Francis, F.; Martin, T.; Lognay, G.; Haubruge, E. Role of (E)-Farnesene in Systematic Aphid Prey Location by Episyrphus Balteatus Larvae (Diptera: Syrphidae). Eur. J. Entomol. 2005, 102, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Francis, F.; Lognay, G.; Haubruge, E. Olfactory Responses to Aphid and Host Plant Volatile Releases: (E)-Beta-Farnesene an Effective Kairomone for the Predator Adalia Bipunctata. J. Chem. Ecol. 2004, 30, 741–755. [Google Scholar] [CrossRef]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Infection of Host Plants by Cucumber Mosaic Virus Increases the Susceptibility of Myzus Persicae Aphids to the Parasitoid Aphidius Colemani. Sci. Rep. 2015, 5, 10963. [Google Scholar] [CrossRef] [Green Version]
- Dader, B.; Then, C.; Berthelot, E.; Ducousso, M.; Ng, J.C.K.; Drucker, M. Insect Transmission of Plant Viruses: Multilayered Interactions Optimize Viral Propagation. Insect Sci. 2017, 24, 929–946. [Google Scholar] [CrossRef]
- Carmo-Sousa, M.; Moreno, A.; Plaza, M.; Garzo, E.; Fereres, A. Cucurbit Aphid-Borne Yellows Virus (CABYV) Modifies the Alighting, Settling and Probing Behaviour of Its Vector Aphis Gossypii Favouring Its Own Spread. Ann. Appl. Biol. 2016, 169, 284–297. [Google Scholar] [CrossRef]
- Mauck, K.E.; Chesnais, Q.; Shapiro, L.R.; Mauck, K. Evolutionary Determinants of Host and Vector Manipulation by Plant Viruses. In Advances in Virus Research; Elsevier Inc.: Amsterdam, The Netherlands, 2018; Volume 101, pp. 189–250. [Google Scholar]
- Ziegler-Graff, V. Molecular Insights into Host and Vector Manipulation by Plant Viruses. Viruses 2020, 12, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Ye, J. Manipulation of Jasmonate Signaling by Plant Viruses and Their Insect Vectors. Viruses 2020, 12, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casteel, C.L.; De Alwis, M.; Bak, A.; Dong, H.; Whitham, S.A.; Jander, G. Disruption of Ethylene Responses by Turnip Mosaic Virus Mediates Suppression of Plant Defense against the Green Peach Aphid Vector. Plant Physiol. 2015, 169, 209–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bak, A.; Mackenzie, P.F.; Perilla-Henao, L.M.; Aegerter, B.J.; Casteel, C.L. Ethylene Signaling Mediates Potyvirus Spread by Aphid Vectors. Oecologia 2019, 190, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Ziebell, H.; Carr, J.P. Cross-Protection: A Century of Mystery. Adv. Virus Res. 2010, 76, 211–264. [Google Scholar] [CrossRef]
- Gal-On, A. A Point Mutation in the FRNK Motif of the Potyvirus Helper Component-Protease Gene Alters Symptom Expression in Cucurbits and Elicits Protection Against the Severe Homologous Virus. Phytopathology 2000, 90, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.-Q.; Natsuaki, T.; Kosaka Seiichi Okuda, Y.; Wang, W.; Natsuaki, T.; Okuda, S.; Kosaka, Y. Comparison of the Nucleotide and Amino Acid Sequences of Parental and Attenuated Isolates of Zucchini Yellow Mosaic Virus. J. Gen. Plant Pathol. 2006, 72, 52–56. [Google Scholar] [CrossRef]
- Ziebell, H.; Payne, T.; Berry, J.O.; Walsh, J.A.; Carr, J.P. A Cucumber Mosaic Virus Mutant Lacking the 2b Counter-Defence Protein Gene Provides Protection against Wild-Type Strains. J. Gen. Virol. 2007, 88, 2862–2871. [Google Scholar] [CrossRef] [PubMed]
- Folimonova, S.Y. Developing an Understanding of Cross-Protection by Citrus Tristeza Virus. Front. Microbiol. 2013, 4, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Schlaefli, H.; Marmonier, A.; Chesnais, Q.; Villeroy, C.; Brault, V. La Multi-Infection Virale de La Betterave à Sucre: Effets Sur l’Accumulation Des Virus et Leur Transmission Par Pucerons. In Proceedings of the Journées BAPOA, Nice, France, 17–18 May 2022. [Google Scholar]
- Francis, S.A.; Luterbacher, M.C. Identification and Exploitation of Novel Disease Resistance Genes in Sugar Beet. Pest Manag. Sci. 2003, 59, 225–230. [Google Scholar] [CrossRef]
- Grimmer, M.K.; Bean, K.M.R.; Qi, A.; Stevens, M.; Asher, M.J.C. The Action of Three Beet Yellows Virus Resistance QTLs Depends on Alleles at a Novel Genetic Locus That Controls Symptom Development. Plant Breed. 2008, 127, 391–397. [Google Scholar] [CrossRef]
- James, L.C.; Bean, K.M.R.; Grimmer, M.K.; Barnes, S.; Kraft, T.; Stevens, M. Varieties of the Future: Identification of “broad Spectrum” Genetic Resistance in Sugar Beet. Int. Sugar J. 2012, 114, 164–168. [Google Scholar]
- Zhang, C.L.; Xu, D.C.; Jiang, X.C.; Zhou, Y.; Cui, J.; Zhang, C.X.; Chen, D.F.; Fowler, M.R.; Elliott, M.C.; Scott, N.W.; et al. Genetic Approaches to Sustainable Pest Management in Sugar Beet (Beta Vulgaris). Ann. Appl. Biol. 2008, 152, 143–156. [Google Scholar] [CrossRef]
- Truniger, V.; Aranda, M. Recessive Resistance to Plant Viruses. Adv. Virus Res. 2009, 75, 119–159. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.; Freeman, B.; Liu, H.Y.; Herrbach, E.; Lemaire, O. Beet Poleroviruses: Close Friends or Distant Relatives? Mol. Plant Pathol. 2005, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Laliberté, J.F. The Genome-Linked Protein VPg of Plant Viruses—A Protein with Many Partners. Curr. Opin. Virol. 2011, 1, 347–354. [Google Scholar] [CrossRef]
- Reinbold, C.; Lacombe, S.; Ziegler-Graff, V.; Scheidecker, D.; Wiss, L.; Beuve, M.; Caranta, C.; Brault, V. Closely Related Poleroviruses Depend on Distinct Translation Initiation Factors to Infect Arabidopsis Thaliana. Mol. Plant-Microbe Interact. 2013, 26, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollwage, L.; Hossain, R.; Varrelmann, M. Zuckerrübe Infizierende Poleroviren Interagieren Mit Multiplen Translationsintiationsfaktoren Ihres Wirtes. In Proceedings of the 54. Jahrestagung des DPG ArbeitskreisesViruskrankheit der Pflanzen, Dossenheim, Germany, 21–22 March 2022. [Google Scholar]
- Smith, H.G.; Hallsworth, P.B. The Effects of Yellowing Viruses on Yield of Sugar Beet in Field Trials, 1985 and 1987. Ann. Appl. Biol. 1990, 116, 503–511. [Google Scholar] [CrossRef]
- Dolja, V.V. Beet Yellows Virus: The Importance of Being Different. Mol. Plant Pathol. 2003, 4, 91–98. [Google Scholar] [CrossRef]
- Gray, S.; Gildow, F.E. Luteovirus-Aphid Interactions. Annu. Rev. Phytopathol. 2003, 41, 539–566. [Google Scholar] [CrossRef]
- Heathcote, G.D. Aphids Caught on Sticky Traps in Eastern England in Relation to the Spread of Yellowing Viruses of Sugar-Beet. Bull. Entomol. Res. 1974, 64, 669–676. [Google Scholar] [CrossRef]
- Heathcote, G.D. The Time of Flight and the Relative Importance of Myzus Persicae (Sulz.) and Aphis Fabae Scop. in Relation to the Incidence of Beet Yellows as Shown by Trap Catches at Rothamsted and Broom’s Barn. Bull. Entomol. Res. 1966, 56, 473–480. [Google Scholar] [CrossRef]
- Boissinot, S.; Pichon, E.; Sorin, C.; Piccini, C.; Scheidecker, D.; Ziegler-Graff, V.; Brault, V. Systemic Propagation of a Fluorescent Infectious Clone of a Polerovirus Following Inoculation by Agrobacteria and Aphids. Viruses 2017, 9, 166. [Google Scholar] [CrossRef] [Green Version]
- Thielemann, R.; Nagi, A. Welche Bedeutung Haben Die Zur “Aphis Fabae-Gruppe” Gehörenden Blattlausstämme für die Übertragung des Schwachen Vergilbungsvirus Auf Beta-Rüben? Z. Pflanz. Pflanzenschutz 1979, 86, 161–168. [Google Scholar]
- Mathers, T.C.; Chen, Y.; Kaithakottil, G.; Legeai, F.; Mugford, S.T.; Baa-Puyoulet, P.; Bretaudeau, A.; Clavijo, B.; Colella, S.; Collin, O.; et al. Rapid Transcriptional Plasticity of Duplicated Gene Clusters Enables a Clonally Reproducing Aphid to Colonise Diverse Plant Species. Genome Biol. 2017, 18, 27. [Google Scholar] [CrossRef] [Green Version]
- Bass, C.; Zimmer, C.T.; Riveron, J.M.; Wilding, C.S.; Wondji, C.S.; Kaussmann, M.; Field, L.M.; Williamson, M.S.; Nauen, R. Gene Amplification and Microsatellite Polymorphism Underlie a Recent Insect Host Shift. Proc. Natl. Acad. Sci. USA 2013, 110, 19460–19465. [Google Scholar] [CrossRef] [Green Version]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex Pheromones and Their Impact on Pest Management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef]
- Smart, L.E.; Aradottir, G.I.; Bruce, T.J.A. Chapter 6—Role of Semiochemicals in Integrated Pest Management. In Integrated Pest Management: Current Concepts and Ecological Perspective; Abrol, D.P., Ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 93–109. [Google Scholar]
- Sharma, A.; Sandhi, R.K.; Reddy, G.V.P. A Review of Interactions between Insect Biological Control Agents and Semiochemicals. Insects 2019, 10, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregg, P.C.; Del Socorro, A.P.; Landolt, P.J. Advances in Attract-and-Kill for Agricultural Pests: Beyond Pheromones. Annu. Rev. Entomol. 2018, 63, 453–470. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.-b.; Chen, J.-l.; Liu, Y.; Francis, F.; Haubruge, E.; Bragard, C.; Sun, J.; Cheng, D. Influence of Garlic Intercropping or Active Emitted Volatiles in Releasers on Aphid and Related Beneficial in Wheat Fields in China. J. Integr. Agric. 2013, 12, 467–473. [Google Scholar] [CrossRef]
- Xu, Q.; Hatt, S.; Lopes, T.; Zhang, Y.; Bodson, B.; Chen, J.; Francis, F. A Push–Pull Strategy to Control Aphids Combines Intercropping with Semiochemical Releases. J. Pest Sci. 2018, 91, 93–103. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, L.; Liu, Y.; Chen, J.; Francis, F. Use of Slow-Release Plant Infochemicals to Control Aphids: A First Investigation in a Belgian Wheat Field. Sci. Rep. 2016, 6, 31552. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Hatt, S.; Han, Z.; Francis, F.; Chen, J. Combining E-β-Farnesene and Methyl Salicylate Release with Wheat-Pea Intercropping Enhances Biological Control of Aphids in North China. Biocontrol. Sci. Technol. 2018, 28, 883–894. [Google Scholar] [CrossRef]
- Hardie, J.; Isaacs, R.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Methyl Salicylate and (−)-(1R,5S)-Myrtenal Are Plant-Derived Repellents for Black Bean Aphid, Aphis Fabae Scop. (Homoptera: Aphididae). J. Chem. Ecol. 1994, 20, 2847–2855. [Google Scholar] [CrossRef]
- Pettersson, J.; Pickett, J.A.; Pye, B.J.; Quiroz, A.; Smart, L.E.; Wadhams, L.J.; Woodcock, C.M. Winter Host Component Reduces Colonization by Bird-Cherry Oat Aphid, Rhopalosiphum Padi (L.) (Homoptera, Aphididae), and Other Aphids in Cereal Fields. J. Chem. Ecol. 1994, 20, 2565–2574. [Google Scholar] [CrossRef]
- Birkett, M.A.; Campbell, C.A.M.; Chamberlain, K.; Guerrieri, E.; Hick, A.J.; Martin, J.L.; Matthes, M.; Napier, J.A.; Pettersson, J.; Pickett, J.A.; et al. New Roles for Cis-Jasmone as an Insect Semiochemical and in Plant Defense. Proc. Natl. Acad. Sci. USA 2000, 97, 9329–9334. [Google Scholar] [CrossRef] [Green Version]
- Ninkovic, V.; Ahmed, E.; Glinwood, R.; Pettersson, J. Effects of Two Types of Semiochemical on Population Development of the Bird Cherry Oat Aphid Rhopalosiphum Padi in a Barley Crop. Agric. Entomol. 2003, 5, 27–34. [Google Scholar] [CrossRef]
- Herrbach, E. Effect of Dodecanoic Acid on the Colonisation of Sugar Beet by Aphids and the Secondary Spread of Virus Yellows. Ann. Appl. Biol. 1987, 111, 477–482. [Google Scholar] [CrossRef]
- Boomsma, J.J.; Jensen, A.B.; Meyling, N.V.; Eilenberg, J. Evolutionary Interaction Networks of Insect Pathogenic Fungi. Annu. Rev. Entomol. 2014, 59, 467–485. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect Pathogens as Biological Control Agents: Back to the Future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, G. Review on Safety of the Entomopathogenic Fungus Metarhizium Anisopliae. Biocontrol. Sci. Technol. 2007, 17, 879–920. [Google Scholar] [CrossRef]
- Berbee, M.L. The Phylogeny of Plant and Animal Pathogens in the Ascomycota. Physiol. Mol. Plant Pathol. 2001, 59, 165–187. [Google Scholar] [CrossRef] [Green Version]
- Vega, F.E.; Meyling, N.V.; Luangsa-Ard, J.J.; Blackwell, M. Fungal Entomopathogens. In Insect Pathology; Vega, F., Kaya, H., Eds.; Academic Press: San Diego, CA, USA, 2012; pp. 171–220. ISBN 9780123849847. [Google Scholar]
- Gange, A.C.; Koricheva, J.; Currie, A.F.; Jaber, L.R.; Vidal, S. Meta-Analysis of the Role of Entomopathogenic and Unspecialized Fungal Endophytes as Plant Bodyguards. New Phytol. 2019, 223, 2002–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyser, C.A.; Jensen, B.; Meyling, N.V. Dual Effects of Metarhizium Spp. and Clonostachys Rosea against an Insect and a Seed-Borne Pathogen in Wheat. Pest Manag. Sci. 2016, 72, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Mantzoukas, S.; Eliopoulos, P.A. Endophytic Entomopathogenic Fungi: A Valuable Biological Control Tool against Plant Pests. Appl. Sci. 2020, 10, 360. [Google Scholar] [CrossRef] [Green Version]
- Rasool, S.; Vidkjær, N.H.; Hooshmand, K.; Jensen, B.; Fomsgaard, I.S.; Meyling, N.V. Seed Inoculations with Entomopathogenic Fungi Affect Aphid Populations Coinciding with Modulation of Plant Secondary Metabolite Profiles across Plant Families. New Phytol. 2021, 229, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Saranya, S.; Ushakumari, R.; Jacob, S.; Philip, B.M. Efficacy of Different Entomopathogenic Fungi against Cowpea Aphid, Aphis Craccivora (Koch). J. Biopestic. 2010, 3, 138–142. [Google Scholar]
- Shan, L.T.; Feng, M.G. Evaluation of the Biocontrol Potential of Various Metarhizium Isolates against Green Peach Aphid Myzus Persicae (Homoptera: Aphididae). Pest Manag. Sci. 2010, 66, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Mweke, A.; Ulrichs, C.; Nana, P.; Akutse, K.S.; Fiaboe, K.K.M.; Maniania, N.K.; Ekesi, S. Evaluation of the Entomopathogenic Fungi Metarhizium Anisopliae, Beauveria Bassiana and Isaria Sp. for the Management of Aphis Craccivora (Hemiptera: Aphididdae). J. Econ. Entomol. 2018, 111, 1587–1594. [Google Scholar] [CrossRef]
- Reingold, V.; Kottakota, C.; Birnbaum, N.; Goldenberg, M.; Lebedev, G.; Ghanim, M.; Ment, D. Intraspecies Variation of Metarhizium Brunneum against the Green Peach Aphid, Myzus Persicae, Provides Insight into the Complexity of Disease Progression. Pest Manag. Sci. 2021, 77, 2557–2567. [Google Scholar] [CrossRef]
- Branine, M.; Bazzicalupo, A.; Branco, S. Biology and Applications of Endophytic Insect-Pathogenic Fungi. PLoS Pathog. 2019, 15, e1007831. [Google Scholar] [CrossRef] [Green Version]
- Lovett, B.; St. Leger, R.J. Stress Is the Rule Rather than the Exception for Metarhizium. Curr. Genet. 2015, 61, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Finkel, O.M.; Castrillo, G.; Herrera Paredes, S.; Salas González, I.; Dangl, J.L. Understanding and Exploiting Plant Beneficial Microbes. Curr. Opin. Plant Biol. 2017, 38, 155–163. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.M.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Newman, M.A.; Pieterse, C.M.J.; Poinssot, B.; Pozo, M.J.; et al. Priming: Getting Ready for Battle. Mol. Plant-Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrath, U. Chapter 9 Priming of Induced Plant Defense Responses. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2009; Volume 51, pp. 361–395. [Google Scholar]
- Zamioudis, C.; Pieterse, C.M.J. Modulation of Host Immunity by Beneficial Microbes. Mol. Plant-Microbe Interact. 2012, 25, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Disi, J.; Simmons, J.; Zebelo, S. Plant Growth-Promoting Rhizobacteria-Induced Defense Against Insect Herbivores. In Field Crops: Sustainable Management by PGPR; Springer: Cham, Switzerland, 2019; pp. 385–410. [Google Scholar] [CrossRef]
- Ongena, M.; Thonart, P. Resistance Induced in Plants by Non-Pathogenic Microorganisms: Elicitation and Defense Responses. Floric. Ornam. Plant Biotechnol. 2006, 3, 447–463. [Google Scholar]
- Jourdan, E.; Ongena, M.; Thonart, P. Caractéristiques Moléculaires de l’immunité Des Plantes Induite Par Les Rhizobactéries Non Pathogènes. Biotechnol. Agron. Soc. Environ. 2008, 12, 437–449. [Google Scholar]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants 2022, 11, 2–19. [Google Scholar] [CrossRef]
- Rabbee, M.F.; Sarafat Ali, M.; Choi, J.; Hwang, B.S.; Jeong, S.C.; Baek, K. Bacillus Velezensis: A Valuable Member of Bioactive Molecules within Plant Microbiomes. Molecules 2019, 24, 1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karthiba, L.; Saveetha, K.; Suresh, S.; Raguchander, T.; Saravanakumar, D.; Samiyappan, R. PGPR and Entomopathogenic Fungus Bioformulation for the Synchronous Management of Leaffolder Pest and Sheath Blight Disease of Rice. Pest Manag. Sci. 2010, 66, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Cawoy, H.; Mariutto, M.; Henry, G.; Fisher, C.; Vasilyeva, N.; Thonart, P.; Dommes, J.; Ongena, M. Plant Defense Stimulation by Natural Isolates of Bacillus Depends on Efficient Surfactin Production. Mol. Plant-Microbe Interact. 2014, 27, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Debois, D.; Fernandez, O.; Franzil, L.; Jourdan, E.; de Brogniez, A.; Willems, L.; Clément, C.; Dorey, S.; De Pauw, E.; Ongena, M. Plant Polysaccharides Initiate Underground Crosstalk with Bacilli by Inducing Synthesis of the Immunogenic Lipopeptide Surfactin. Environ. Microbiol. Rep. 2015, 7, 570–582. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francis, F.; Then, C.; Francis, A.; Gbangbo, Y.A.C.; Iannello, L.; Ben Fekih, I. Complementary Strategies for Biological Control of Aphids and Related Virus Transmission in Sugar Beet to Replace Neonicotinoids. Agriculture 2022, 12, 1663. https://doi.org/10.3390/agriculture12101663
Francis F, Then C, Francis A, Gbangbo YAC, Iannello L, Ben Fekih I. Complementary Strategies for Biological Control of Aphids and Related Virus Transmission in Sugar Beet to Replace Neonicotinoids. Agriculture. 2022; 12(10):1663. https://doi.org/10.3390/agriculture12101663
Chicago/Turabian StyleFrancis, Frédéric, Christiane Then, Adrien Francis, Yao Aime Constant Gbangbo, Lisa Iannello, and Ibtissem Ben Fekih. 2022. "Complementary Strategies for Biological Control of Aphids and Related Virus Transmission in Sugar Beet to Replace Neonicotinoids" Agriculture 12, no. 10: 1663. https://doi.org/10.3390/agriculture12101663