
Protoplast Isolation, Fusion, Culture and Transformation in the Woody Plant Jasminum spp.

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials Sources and Culture Conditions
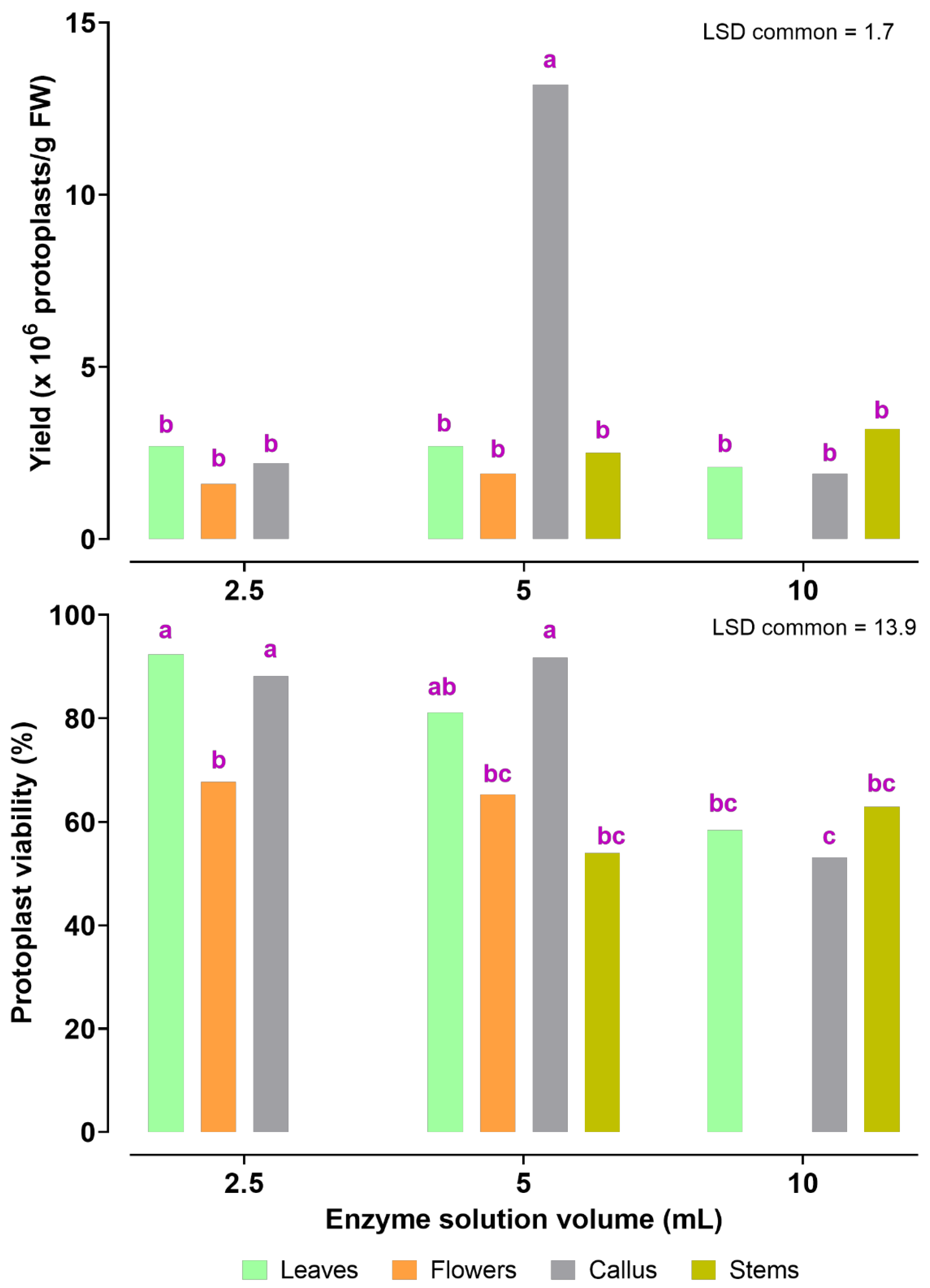
2.2. Factors Affecting the Yield and Viability of the Isolated Protoplasts
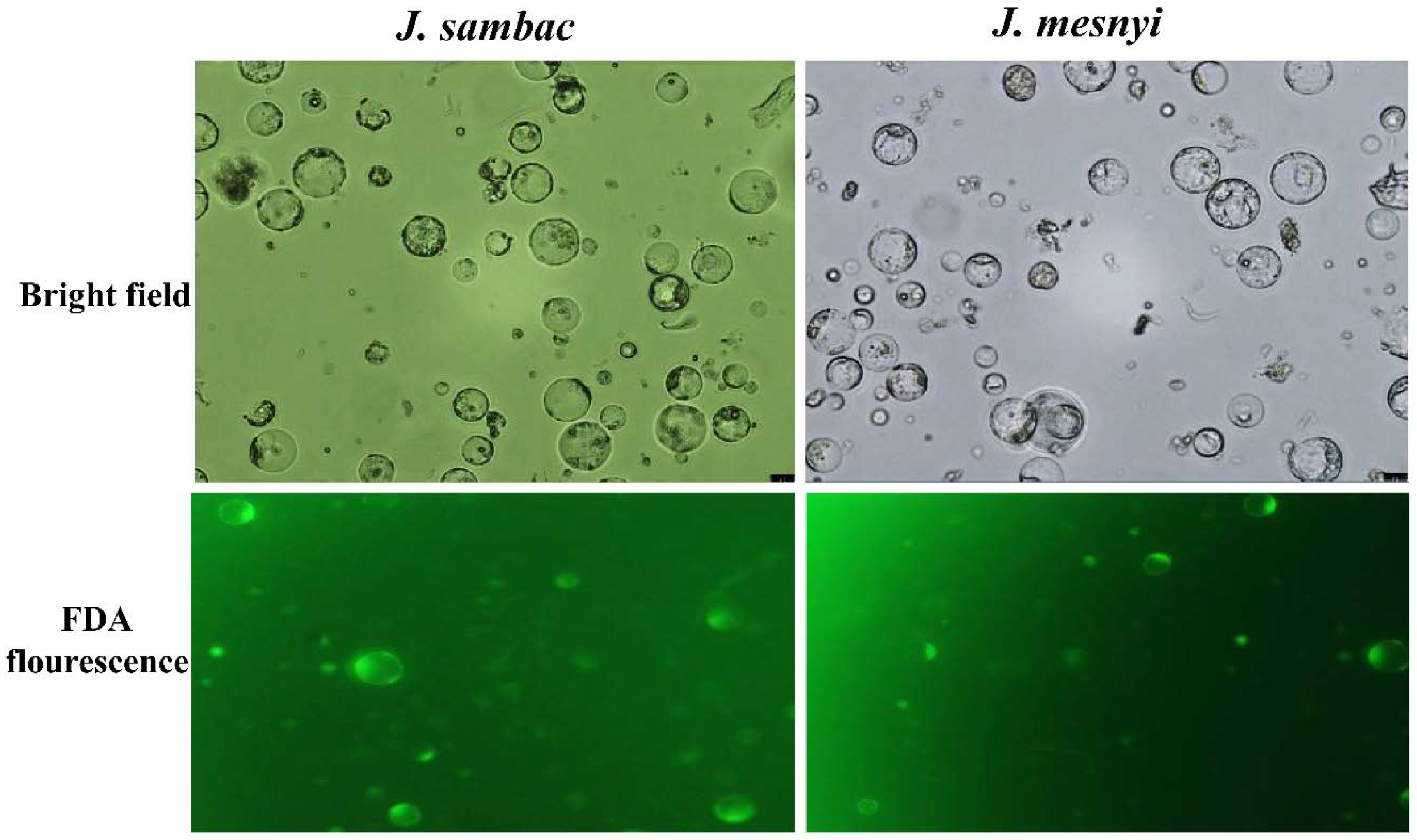
2.3. Isolation, Purification and Determination of the Protoplasts Yield and Viability
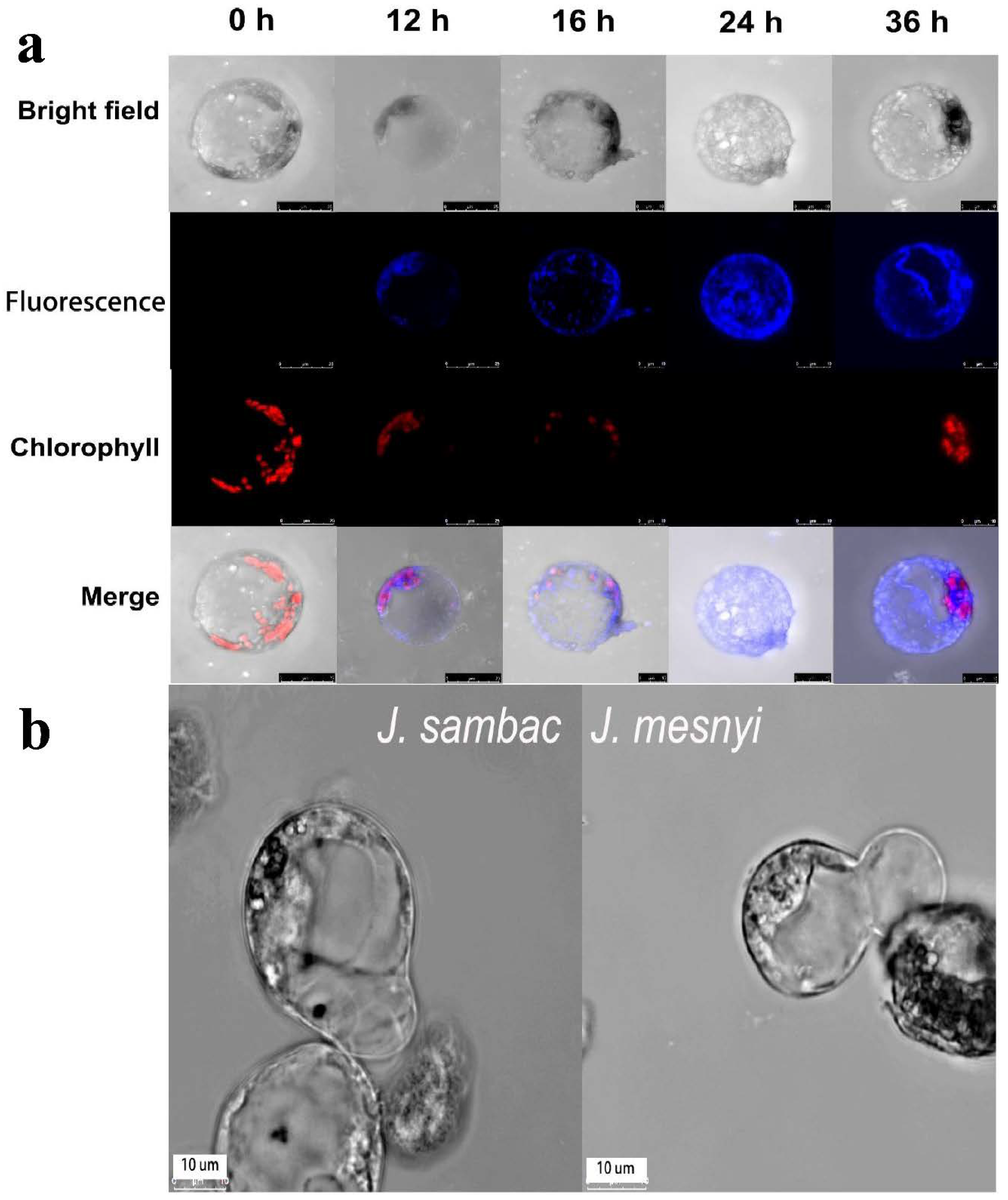
2.4. Protoplast Staining and Image Acquisition for Cellulose Fibrils
2.5. PEG-Induced Protoplast Fusion
2.6. Transient Transformation of Protoplasts
2.7. Protoplast Culture
2.8. Statistical Data Analysis
- ∙
- The experiment “the effect of plant growth regulators on callus induction from stem segment and shooting rate of J. mesnyi” has 29 treatments and each treatment has three replicates.
- ∙
- The experiment “protoplast yield and viability of isolated protoplast from leaves, flowers, stems and callus of J. sambac L. using different enzyme solutions” was designed as a split plot (main plot was the enzyme and buffer combination, with 2 levels, and the sub plot was the incubation time with 5 levels) and each treatment was replicated three times.
- ∙
- The experiment “effects of different fusion solution and fusion time on protoplast fusion frequency (%) between J. sambac. L and J. mesnyi” was designed as a split plot (main plot was the fusion solution with 6 levels, and the sub plot was the fusion time, with 3 levels) and each treatment was replicated three times.
3. Results
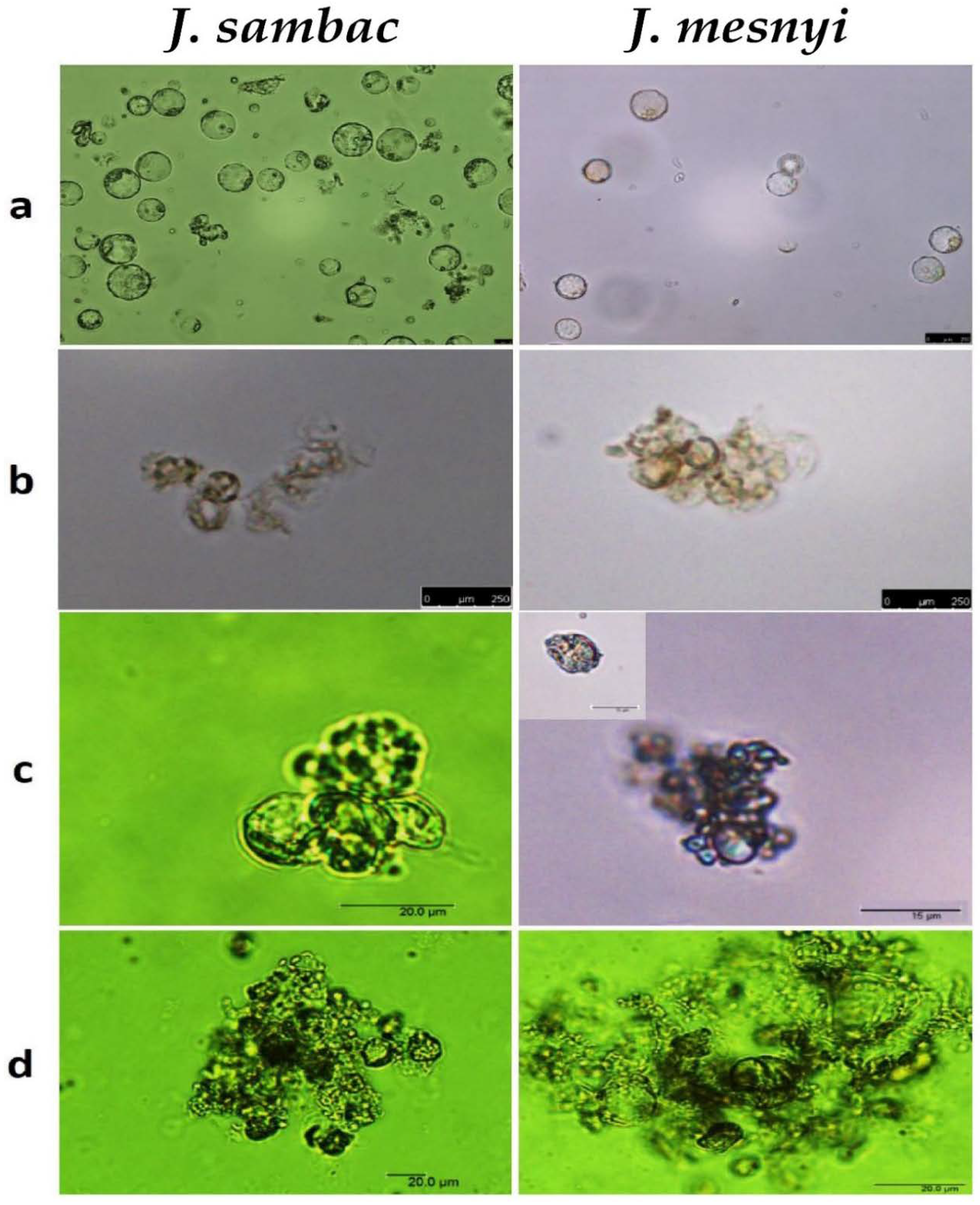
3.1. Establishment of J. sambac and J. mesnyi Callus Cultures for Protoplast Isolation
3.2. Optimization of Factors Affecting Protoplast Isolation and Transient Transformation System of Jasmine Protoplasts
3.3. Observation of the Cell Wall Regeneration and Optimization of PEG-Mediated Protoplast Fusion
3.4. Protoplasts Culture and Microcallus Formation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaitanya, H.S.; Nataraja, S.K.M. Review on Propagation Techniques of Jasmine (Jasminum sambac (L.)). J. Pharmacogn. Phytochem. 2018, 7, 593–596. [Google Scholar]
- Lu, Y.; Liu, Z.; Lyu, M.; Yuan, Y.; Wu, B. Characterization of JsWOX1 and JsWOX4 during callus and root induction in the shrub species Jasminum sambac. Plants 2019, 8, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.; Qarshi, I.A.; Nazir, H.; Ullah, I. Plant Tissue Culture: Current Status and Opportunities. In Recent Advances in Plant In Vitro Culture; Leva, A., Rinaldi, L., Eds.; IntechOpen: London, UK, 2012; pp. 1–28. [Google Scholar]
- Brown, D.C.W.; Thorpe, T.A. Crop improvement through tissue culture. World J. Microbiol. Biotechnol. 1995, 11, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Mccown, B. Special symposium: In vitro recalcitrance. Recalcitrance of woody and herbaceous perennial plants: Dealing with genetic predeterminism. Vitr. Cell. Dev. Biol. 2000, 36, 149–154. [Google Scholar] [CrossRef]
- Ruisheng, G.; Xiangning, J.; Zhongchen, G. Research progress on the mechanism of organogenesis in vitro culture of plants. Chin. J. Bot. 1999, 16, 238–244. [Google Scholar]
- Naibo, Y. Tissue culture and organogenesis of several woody Plants. Chin. J. Plant Physiol. 1982, 4, 25–29. [Google Scholar]
- Umate, P.; Rao, K.V.; Kiranmayee, K.; Sree, T.J.; Sadanandam, A. Plant regeneration of mulberry (Morus indica) from mesophyll-derived protoplasts. Plant Cell Tissue Organ Cult. 2005, 82, 289–293. [Google Scholar] [CrossRef]
- Duquenne, B.; Eeckhaut, T.; Werbrouck, S.; Van Huylenbroeck, J. Effect of enzyme concentrations on protoplast isolation and protoplast culture of Spathiphyllum and Anthurium. Plant Cell Tissue Organ Cult. 2007, 91, 165–173. [Google Scholar] [CrossRef]
- Prabavathy, V.R.; Mathivanan, N.; Sagadevan, E.; Murugesan, K.; Lalithakumari, D. Intra-strain protoplast fusion enhances carboxymethyl cellulase activity in Trichoderma reesei. Enzym. Microb. Technol. 2006, 38, 719–723. [Google Scholar] [CrossRef]
- Nassour, M.; Dorion, N. Plant regeneration from protoplasts of micropropagated Pelargonium x hortorum “Alain”: Effect of some environmental and medium factors on protoplast system efficiency. Plant Sci. 2002, 163, 169–176. [Google Scholar] [CrossRef]
- Makonkawkeyoon, S.; Smitamana, P.; Hirunpetcharat, C.; Maneekarn, N. Production of mouse immunoglobulin G by a hybrid plant derived from tobacco-mouse cell fusions. Experentia 1995, 51, 19–25. [Google Scholar]
- Hassanein, A.; Hamama, L.; Loridon, K.; Dorion, N. Direct gene transfer study and transgenic plant regeneration after electroporation into mesophyll protoplasts of Pelargonium x hortorum, “Panaché Sud”. Plant Cell Rep. 2009, 28, 1521–1530. [Google Scholar] [CrossRef]
- Shen, Y.; Meng, D.; McGrouther, K.; Zhang, J.; Cheng, L. Efficient isolation of Magnolia protoplasts and the application to subcellular localization of MdeHSF1. Plant Methods 2017, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Su, J.; Duan, S.; Ao, Y.; Dai, J.; Liu, J.; Wang, P.; Li, Y.; Liu, B.; Feng, D.; et al. A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes. Plant Methods 2011, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Loyd, G.; McGown, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Proc. Int. Plant Propagators Soc. 1980, 30, 421–427. [Google Scholar]
- Frey, D.D.; Wang, H. Adaptive one-factor-at-a-time experimentation and expected value of improvement. Technometrics 2006, 48, 418–431. [Google Scholar] [CrossRef]
- Ling, A.P.K.; Phua, G.A.T.; Tee, C.S.; Hussein, S. Optimization of protoplast isolation protocols from callus of Eurycoma longifolia. J. Med. Plants Res. 2010, 4, 1778–1785. [Google Scholar]
- Sangra, A.; Shahin, L.; Dhir, S.K. Optimization of isolation and culture of protoplasts in alfalfa (Medicago sativa) cultivar Regen-SY. Am. J. Plant Sci. 2019, 10, 1206–1219. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widholm, J.M. The use of fluorescein diacetate and phenosafranine for determining viability of cultured plant cells. Stain Technol. 1972, 47, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Larkin, P.J. Purification and viability determinations of plant protoplasts. Planta 1976, 128, 213–216. [Google Scholar] [CrossRef]
- Durieu, P.; Ochatt, S.J. Efficient intergeneric fusion of pea (Pisum sativum L.) and grass pea (Lathyrus sativus L.) protoplasts. J. Exp. Bot. 2000, 51, 1237–1242. [Google Scholar]
- Menczel, L.; Wolfe, K. High frequency of fusion induced in freely suspended protoplast mixtures by polyethylene glycol and dimethylsulfoxide at high pH. Plant Cell Rep. 1984, 3, 196–198. [Google Scholar] [CrossRef]
- Assani, A.; Chabane, D.; Haïcour, R.; Bakry, F.; Wenzel, G.; Foroughi-Wehr, B. Protoplast fusion in banana (Musa spp.): Comparison of chemical (PEG: Polyethylene glycol) and electrical procedure. Plant Cell Tissue Organ Cult. 2005, 83, 145–151. [Google Scholar] [CrossRef]
- Hayat, S.; Christias, C. Isolation and fusion of protoplasts from the phytopathogenic fungus Sclerotium rolfsii (Sacc.). Braz. J. Microbiol. 2010, 41, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Geerts, P.; Druart, P.; Ochatt, S.; Baudoin, J.P. Protoplast fusion technology for somatic hybridisation in Phaseolus. Biotechnol. Agron. Soc. Environ. 2008, 12, 371–377. [Google Scholar]
- Peng, Z.; Tong, H.R.; Liang, G.L.; Shi, Y.Q.; Yuan, L.Y. Protoplast isolation and fusion induced by PEG with leaves and roots of tea plant (Camellia sinensis L. O. Kuntze). Acta Agron. Sin. 2018, 44, 463–470. [Google Scholar] [CrossRef]
- McCown, B.H. From gene manipulation to forest establishment: Shoot cultures of woody plants can be a central tool. Tappi 1985, 68, 116–119. [Google Scholar]
- U-Kong, W.; Wongsawad, P.; Buddharak, P. Shoot bud and young leaf induction of Jasminum spp. in in vitro culture. Int. J. Appl. Agric. Res. 2012, 7, 17–26. [Google Scholar]
- Salim, S.A.A. Effect of plant growth regulators BA, 2,4-D, IBA and Kin on in vitro propagation of white jasmine (Jasminum azoricum L.). J. Univ. Babylon 2016, 24, 795–802. [Google Scholar]
- Kareem, A.; Radhakrishnan, D.; Sondhi, Y.; Aiyaz, M.; Roy, M.V.; Sugimoto, K.; Prasad, K. De novo assembly of plant body plan: A step ahead of Deadpool. Regeneration 2016, 3, 182–197. [Google Scholar] [CrossRef]
- Aqeel, R.; Zehra, M.; Kazmi, S.K.; Khan, S.; Kayani, H.A.; Mirbahar, A.A. A study on the isolation of protoplasts from mesophyll cells of Dendrobium Queen Pink. Pak. J. Bot. 2016, 48, 693–697. [Google Scholar]
- Rahmani, M.S.; Pijut, P.M.; Shabanian, N. Protoplast isolation and genetically true-to-type plant regeneration from leaf- and callus-derived protoplasts of Albizia julibrissin. Plant Cell Tissue Organ Cult. 2016, 127, 475–488. [Google Scholar] [CrossRef]
- Eeckhaut, T.; Lakshmanan, P.S.; Deryckere, D.; Van Bockstaele, E.; Van Huylenbroeck, J. Progress in plant protoplast research. Planta 2013, 238, 991–1003. [Google Scholar] [CrossRef]
- Chamani, E.; Tahami, S.K. Efficient protocol for protoplast isolation and plant regeneration of Fritillaria imperialis L. J. Agric. Sci. Technol. 2016, 18, 467–482. [Google Scholar]
- Mohan Jain, S. Tissue culture-derived variation in crop improvement. Euphytica 2001, 118, 153–166. [Google Scholar] [CrossRef]
- Hesemann, C.U.; Schröder, G. Loss of nuclear DNA in leaves of rye. Theor. Appl. Genet. 1982, 62, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Melchers, G. Microbial techniques in somatic hybridisation by fusion of protoplasts. In International Cell Biology; Brinkley, B.R., Porter, K.R., Eds.; Rockefeller University Press: Boston, MA, USA, 1977; pp. 207–215. [Google Scholar]
- Guan, Q.; Guo, Y.; Wei, Y.; Meng, F.; Zhang, Z. Regeneration of somatic hybrids of ginger via chemical protoplast fusion. Plant Cell Tissue Organ Cult. 2010, 102, 279–284. [Google Scholar] [CrossRef]
- Wanmei, J.; Zhenhui, G.; Guirong, L. Plant genetic transformation methods and identification of transgenic plants. Shaanxi Agric. Sci. 2000, 1, 24–29. [Google Scholar]
- Wu, F.H.; Shen, S.C.; Lee, L.Y.; Lee, S.H.; Chan, M.T.; Lin, C.S. Tape-arabidopsis sandwich—A simpler arabidopsis protoplast isolation method. Plant Methods 2009, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endress, R. Plant cells as producers of secondary compounds. In Plant Cell Biotechnology; Springer: Berlin/Heidelberg, Germany, 1994; pp. 121–255. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solution Name | Solution Composition | Storage | Function |
|---|---|---|---|
| Buffer solution | 0.4 M Mannitol, 0.2 M MES, 1 M CaCl2, 0.2 M KCL, 1 M NaH2PO4 | Freshly prepared | Digestion of cell wall material |
| Enzyme solution | 1.5% Cellulases Onozuka R10, 1.5% Cellulases Onozuka RS, 0.4% Macerozyme R-10, 0.8% Pectinase | Freshly prepared | Digestion of cell wall material |
| W5 | 2 mM MES, 154 mM NaCl, 125 mM CaCl2 and 5 mM KCl, (pH 5.7) | 4 °C | Release and washing protoplasts |
| W1 | 0.8 M Mannitol, 10 mM KCL, 20 mM MES (pH 5.7), ddH2O | Freshly prepared | Re-suspend protoplast after transfection |
| MMg | 0.8 M Mannitol, 10 mM CaCl2, 20 mM MES (pH 5.7), ddH2O | Freshly prepared | Res-uspend the protoplast for counting and transfection |
| PEG | 40% (w/v) PEG 4000, 0.8 M Mannitol and 0.1 M Ca(NO3)2 | Freshly prepared | Transform plasmids (10 ug is used in this study) into protoplasts |
| Treatments | Basal Media | Phytohormones (mg/L) Auxin Cytokinin | Ratio Auxin/Cytokinin | Total Number of Callus (30–45 Days) 1,2 | Frequency of Callus Induction (%) (1 Month) 2 | Frequency of Shooting 3 (%) (3 Months) | Extended Callus Culture (4–6 Months) | |
|---|---|---|---|---|---|---|---|---|
| 1 | WPM | 0.25 NAA | 0.50 6-BA | 33/67 | 96 c | 80 a,b | 0 | browning, dying |
| 2 | 0.50 NAA | 1.00 6-BA | 33/67 | 112 b | 60 d,e,f,g,h | 0 | browning, dying | |
| 3 | 0 | 1.50 6-BA | 0/100 | 162 a | 87 a | 0 | browning, dying | |
| 4 | 0.50 NAA | 0 | 100/0 | 100 b,c | 61 d,e,f,g,h | 0 | most browning | |
| 5 | 0.50 NAA | 1.50 6-BA | 25/75 | 112 b | 61 d,e,f,g,h | 0 | browning, dying | |
| 6 | 0.50 NAA | 2.00 6-BA | 20/80 | 162 a | 87 a | 0 | browning, dying | |
| 7 | 0.10 IBA | 2.00 6-BA | 5/95 | 112 b | 60 e,f,g,h | 0 | most browning | |
| 8 | 0 | 0.50 6-BA | 0/100 | 100 b,c | 54 g,h | 0 | browning, dying | |
| 9 | 0.25 NAA | 0 | 100/0 | 112 b | 60 e,f,g,h | 0 | browning, dying | |
| 10 | 0.10 NAA | 4.00 6-BA | 2/98 | 100 b,c | 54 g,h | 0 | browning, dying | |
| 11 | 0 | 2.00 KT | 0/100 | 96 c | 52 h | 0 | browning, dying | |
| 12 | 0 | 4.00 6-BA | 0/100 | 100 b,c | 61 d,e,f,g,h | 0 | browning, dying | |
| 13 | 0.10 2,4-D | 4.00 6-BA | 2/98 | 112 b | 60 e,f,g,h | 0 | browning, dying | |
| 14 | 0 | 2.00 6-BA | 0/100 | 100 b,c | 56 g,h | 0 | some browning | |
| 15 | 0 | 1.00 KT | 0/100 | 100 b,c | 54 g,h | 0 | some browning | |
| 16 | 5.25 NAA | 15.75 6-BA | 25/75 | 112 b | 70 c,d | 0.89 | browning, dying | |
| 17 | 0.50 IAA | 1.00 ZR | 33/67 | 100 b,c | 61 d,e,f,g,h | 0 | some browning | |
| 18 | 0.50 IBA | 2.00 ZR | 20/80 | 112 b | 69 c,d,e | 0 | most browning | |
| 19 | 0.50 IAA | 0.20 TDZ | 71/29 | 100 b,c | 56 g,h | 0 | some browning | |
| 20 | 0.50 IBA | 0.50 TDZ | 50/50 | 112 b | 60 e,f,g,h | 0 | some browning | |
| 21 | 0.50 IBA | 1.00 TDZ | 33/67 | 96 c | 59 f,g,h | 0 | most browning | |
| 22 | 0.50 IAA | 2.00 ZR | 20/80 | 100 b,c | 56 g,h | 0 | some browning | |
| 23 | 0.50 IAA | 5.00 ZR | 9/91 | 112 b | 62 d,e,f,g | 0 | some browning | |
| 24 | 0.50 IBA | 3.00 TDZ | 14/86 | 100 b,c | 68 c,d,e,f | 0 | some browning | |
| 25 | 0.60 NAA | 2.00 6-BA | 23/77 | 96 c | 62 d,e,f,g | 0.62 | browning, dying | |
| 26 | 0 | 5.00 6-BA | 0/100 | 100 b,c | 63 d,e,f,g | 0 | some browning | |
| 27 | MS | 0 | 2.00 6-BA | 0/100 | 112 b | 76 b,c | 0 | some browning |
| 28 | 0 | 1.00 KT | 0/100 | 162 a | 87 a | 0 | most browning | |
| 29 | 1.00 NAA | 3.00 6-BA | 25/75 | 100 b,c | 61 e,f,g,h | 0 | some browning | |
| Enzyme and Buffer Combinations | Time | Yield (×106 protoplasts/gFW) | Viability (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Leaves | Flowers | Stems | Callus | Leaves | Flowers | Stems | Callus | ||
| 1 | 1 | 2.5 ± 0.3 b,c,d | 2.4 ± 0.4 a | 0.4 ± 0.0 e | 1.2 ± 0.0 e | 63.7 ± 5.3 d | 54.4 ± 1.9 d | 72.3 ± 3.1 a | 80.3 ± 1.9 c,d |
| 2 | 3.3 ± 0.5 b | 2.8 ± 0.2 a | 0.2 ± 0.0 e | 1.6 ± 0.0 e | 63.2 ± 3.0 d | 63.5 ± 3.6 c | 61.1 ± 1.1 b,c | 78.8 ± 2.7 c,d | |
| 3 | 1.4 ± 0.3 c,d,e | 2.8 ± 0.2 a | 1.4 ± 0.3 d | 5.4 ± 2.0 d | 84.0 ± 1.6 a,b | 70.6 ± 1.3 b | 57.5 ±1.1 c | 72.8 ± 2.9 e | |
| 4 | 3.2 ± 0.5 b,c | 3.1 ± 0.2 a | 3.3 ± 0.5 a | 12.1 ± 3.8 b,c | 77.7 ± 2.8 b,c | 61.8 ± 6.2 c | 63.0 ± 0.8 b | 81.5 ± 1.2 b,c | |
| 5 | 0.18 ± 0.0 e | 3.3 ± 0.1 a | 2.8 ± 0.1 b | 1.3 ± 0.0 e | 46.5 ± 3.5 e | 61.5 ± 3.0 c | 64.2 ± 2.7 b | 75.7 ± 1.5 d,e | |
| 2 | 1 | 5.4 ± 0.1 a | 0.1 ± 0.0 a | 2.3 ± 0.1 b | 11.3 ± 0.4 c | 89.6 ± 8.0 a | 61.6 ± 1.2 c | 72.5 ± 2.0 a | 88.0 ± 2.7 a |
| 2 | 1.1 ± 0.0 d,e | 3.9 ± 5.0 a | 2.2 ± 0.1 c | 14.1 ± 1.5 b | 88.1 ± 4.6 a | 82.2 ± 1.6 a | 44.6 ± 3.0 e | 86.6 ± 5.8 a,b | |
| 3 | 2.5 ± 0.3 b,c,d | 2.0 ± 1.3 a | 0.1 ± 0.0 e | 23.8 ± 4.3 a | 78.7 ± 6.4 b,c | 83.0 ± 2.8 a | 53.3 ± 1.1 d | 76.7 ± 2.0 c,d,e | |
| 4 | 1.7 ± 0.4 b,c,d,e | 0.2 ± 0.0 a | 0.2 ± 0.0 e | 14.5 ± 3.6 b | 72.6 ± 5.6 c | 42.4 ± 2.0 e | 69.2 ± 2.8 a | 81.5 ± 4.3 b,c | |
| 5 | 5.6 ± 3.9 a | 0.1 ± 0.0 a | 3.1 ± 0.2 a | 12.6 ± 1.8 b,c | 49.2 ± 3.0 e | 53.6 ± 2.7 d | 58.6 ± 6.0 c | 77.2 ± 5.0 c,d,e | |
| Factor ANOVA | |||||||||
| Variables | Sources | Enzyme and buffer combinations | Time | Enzyme and buffer combinations X Time | R2 | LSD | |||
| Yield (×106 protoplasts/gFW) | Leaves | * | NS | ** | 0.777 | 1.9 | |||
| Flowers | * | NS | NS | 0.603 | - | ||||
| Stems | NS | ** | ** | 0.978 | 0.3 | ||||
| Callus | ** | ** | ** | 0.965 | 2.8 | ||||
| Viability (%) | Leaves | ** | ** | ** | 0.983 | 8.7 | |||
| Flowers | NS | ** | ** | 0.967 | 4.3 | ||||
| Stems | * | ** | ** | 0.943 | 4.1 | ||||
| Callus | ** | ** | * | 0.887 | 5.7 | ||||
| Fusion Solution | Fusion Time (min) | Fusion Solution X Fusion Time (min) | R2 | LSD + | |
|---|---|---|---|---|---|
| Fusion rate (binary fusion) (%) | ** | * | ** | 0.998 | 1.8 |
| Viability (%) | ** | ** | ** | 0.999 | 5.7 |
| Fusion Solution | Fusion Time (min) | Fusion Rate (Binary Fusion) (%) | Viability (%) |
|---|---|---|---|
| 1 | 15 | 0 ± 0 d | 0 ± 0 e |
| 20 | 0 ± 0 d | 0 ± 0 e | |
| 25 | 0 ± 0 d | 0 ± 0 e | |
| 2 | 15 | 1.34 ± 0.05 c,d | 25.06 ± 0.42 c |
| 20 | 2.04 ± 0.06 c | 33.33 ± 4.27 b | |
| 25 | 1.54 ± 0.05 c,d | 22.23 ± 0.21 c,d | |
| 3 | 15 | 0 ± 0 d | 0 ± 0 e |
| 20 | 0 ± 0 d | 0 ± 0 e | |
| 25 | 0 ± 0 d | 0 ± 0 e | |
| 4 | 15 | 1.23 ± 0.01c,d | 18.23 ± 0.35 d |
| 20 | 1.16 ± 0.02c,d | 22.27 ± 0.42 c,d | |
| 25 | 2.83 ± 0.04 b,c | 48.81 ± 0.21 a | |
| 5 | 15 | 2.67 ± 0.21 b,c | 36.07 ± 0.66 a,b |
| 20 | 5.61 ± 0.29 a | 54.45 ± 0.57 a | |
| 25 | 4.42 ± 0.16 a,b | 39.09 ± 0.36 a | |
| 6 | 15 | 0 ± 0 d | 0 ± 0 e |
| 20 | 0 ± 0 d | 0 ± 0 e | |
| 25 | 0 ± 0 d | 0 ± 0 e |
| PEG MW (% (w/v)) | Fusion Rate (%) | Viability (%) |
|---|---|---|
| 4000 | 2.08 ± 0.05 b | 25.72 ± 0.84 b |
| 6000 | 5.64 ± 0.19 a | 47.76 ± 0.92 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, M.A.A.; Miao, M.; Pratsinakis, E.D.; Zhang, H.; Wang, W.; Yuan, Y.; Lyu, M.; Iftikhar, J.; Yousef, A.F.; Madesis, P.; et al. Protoplast Isolation, Fusion, Culture and Transformation in the Woody Plant Jasminum spp. Agriculture 2021, 11, 699. https://doi.org/10.3390/agriculture11080699
Ahmed MAA, Miao M, Pratsinakis ED, Zhang H, Wang W, Yuan Y, Lyu M, Iftikhar J, Yousef AF, Madesis P, et al. Protoplast Isolation, Fusion, Culture and Transformation in the Woody Plant Jasminum spp. Agriculture. 2021; 11(8):699. https://doi.org/10.3390/agriculture11080699
Chicago/Turabian StyleAhmed, Mohamed A. A., Miao Miao, Emmanouil D. Pratsinakis, Hongliang Zhang, Wei Wang, Yuan Yuan, Meiling Lyu, Junaid Iftikhar, Ahmed F. Yousef, Panagiotis Madesis, and et al. 2021. "Protoplast Isolation, Fusion, Culture and Transformation in the Woody Plant Jasminum spp." Agriculture 11, no. 8: 699. https://doi.org/10.3390/agriculture11080699