
Stressor-Specific Sex Differences in Amygdala–Frontal Cortex Networks
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample
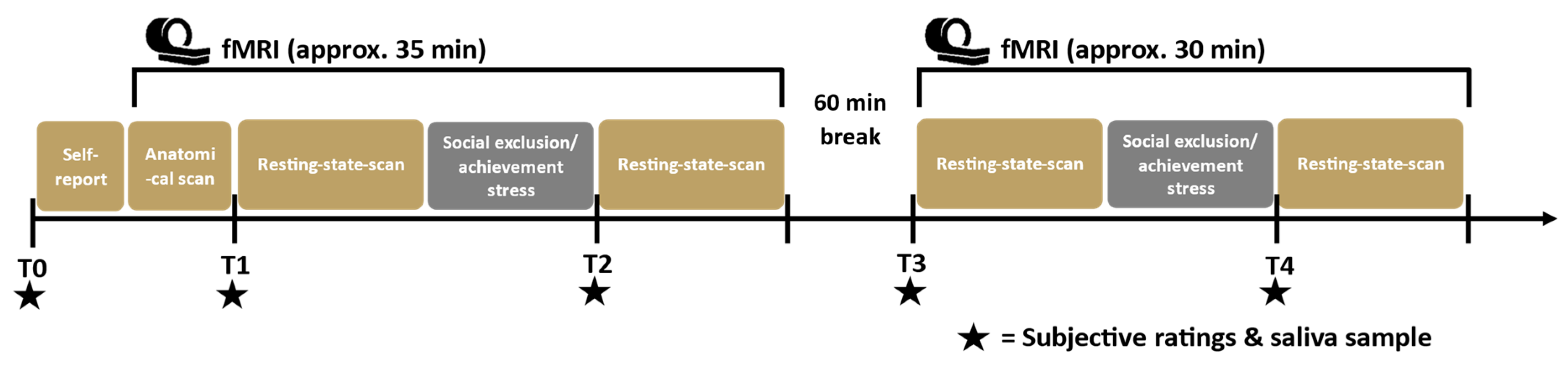
2.2. Procedure
2.3. Stress Tasks
2.3.1. Modified Montreal Imaging Stress Task—MIST
2.3.2. Cyberball
2.4. Saliva Samples
2.5. Data and Statistical Analysis of Behavioural and Hormonal Data
2.6. rsFC Analyses
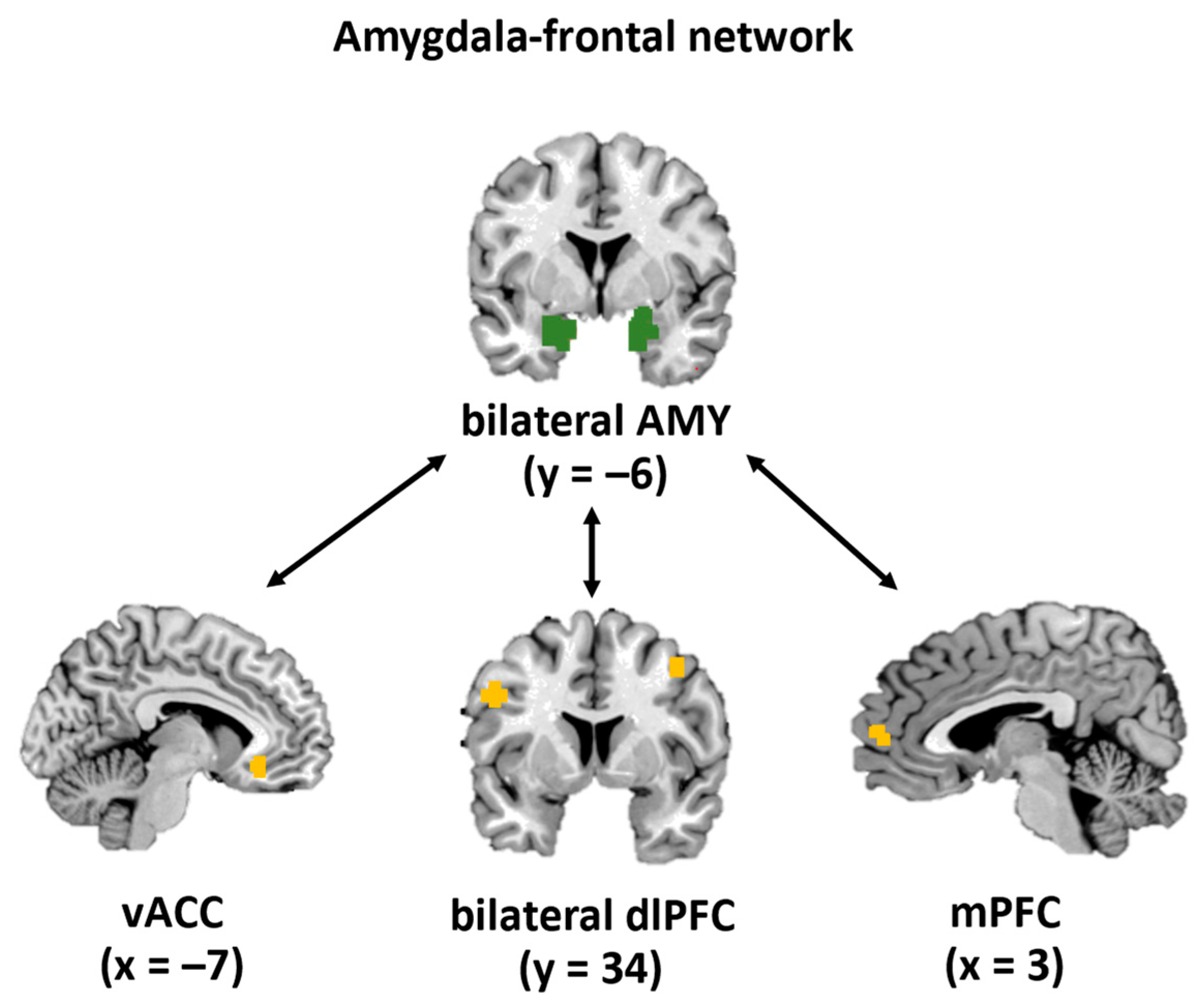
2.6.1. Definition of Regions of Interest
2.6.2. Acquisition, Pre-Processing, and Calculation of rsFC
2.6.3. Statistical Analyses of rsFCs
Exploratory Regression Analyses
3. Results
3.1. Sample Description
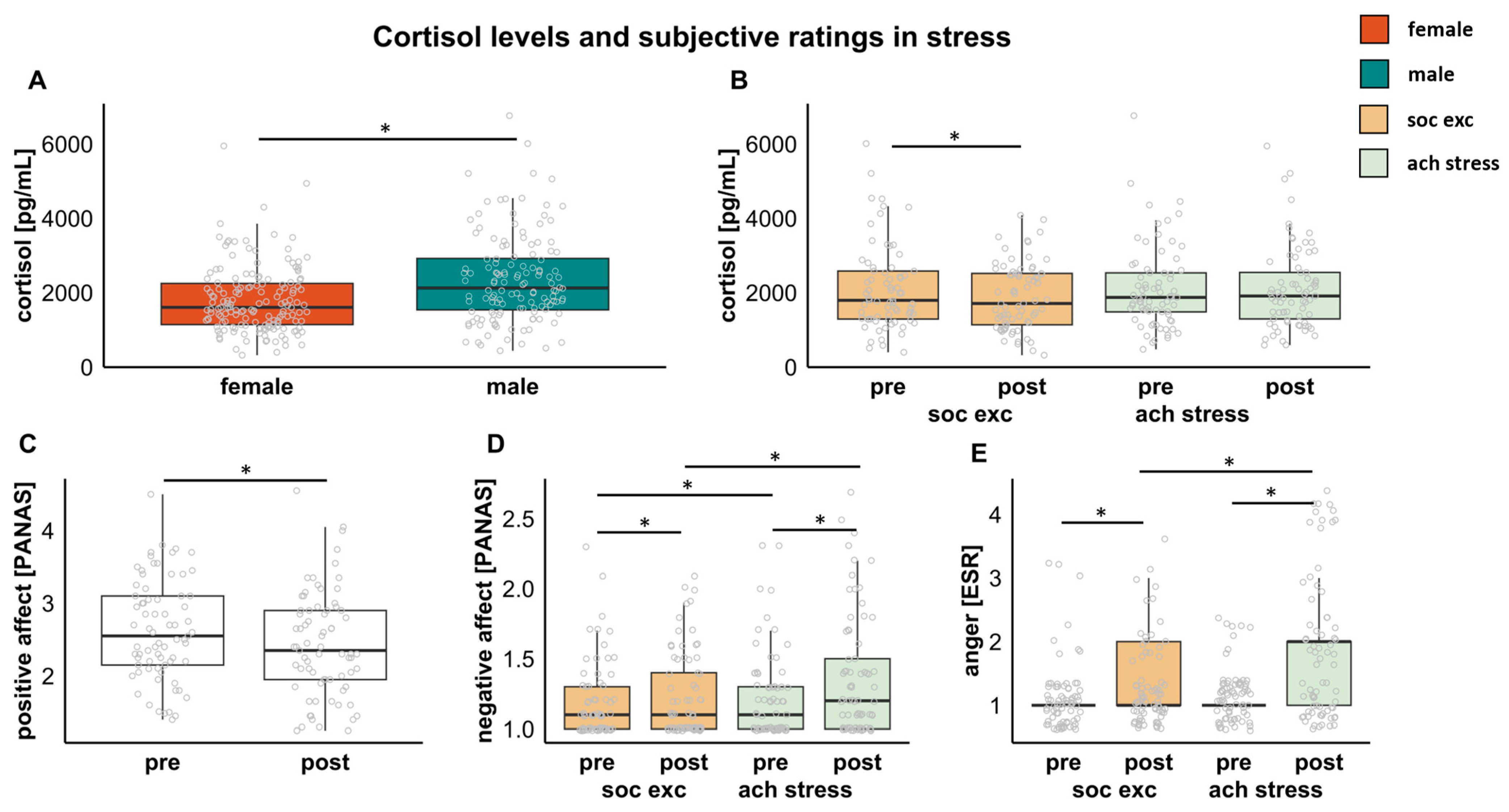
3.2. Associations between Stress and Cortisol
3.3. Associations between Stress and Subjective Rating
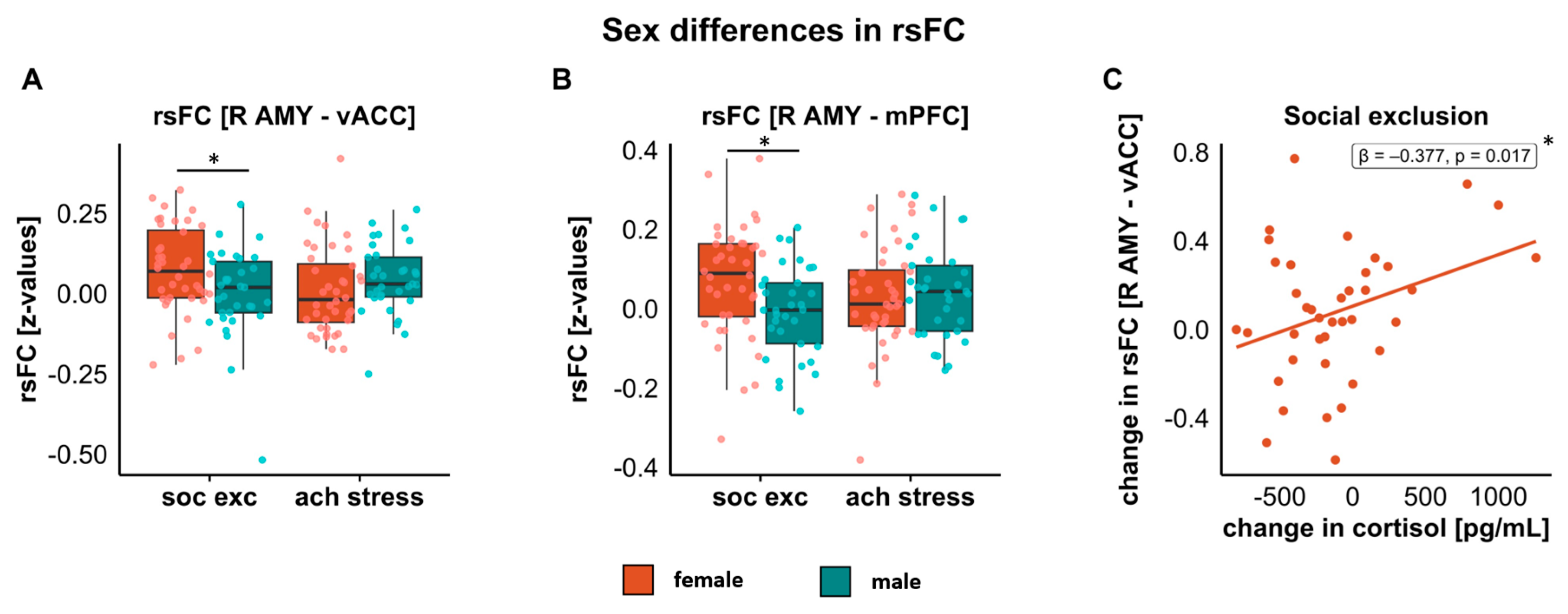
3.4. Associations between Stress and rsFC
3.4.1. Amygdala–vACC
3.4.2. Amygdala–mPFC
3.4.3. Amygdala–dlPFC
3.5. Cortisol and Subjective Ratings as Predictors of rsFC Changes
3.5.1. Predictor: Changes in Cortisol from Pre- to Post-Stress
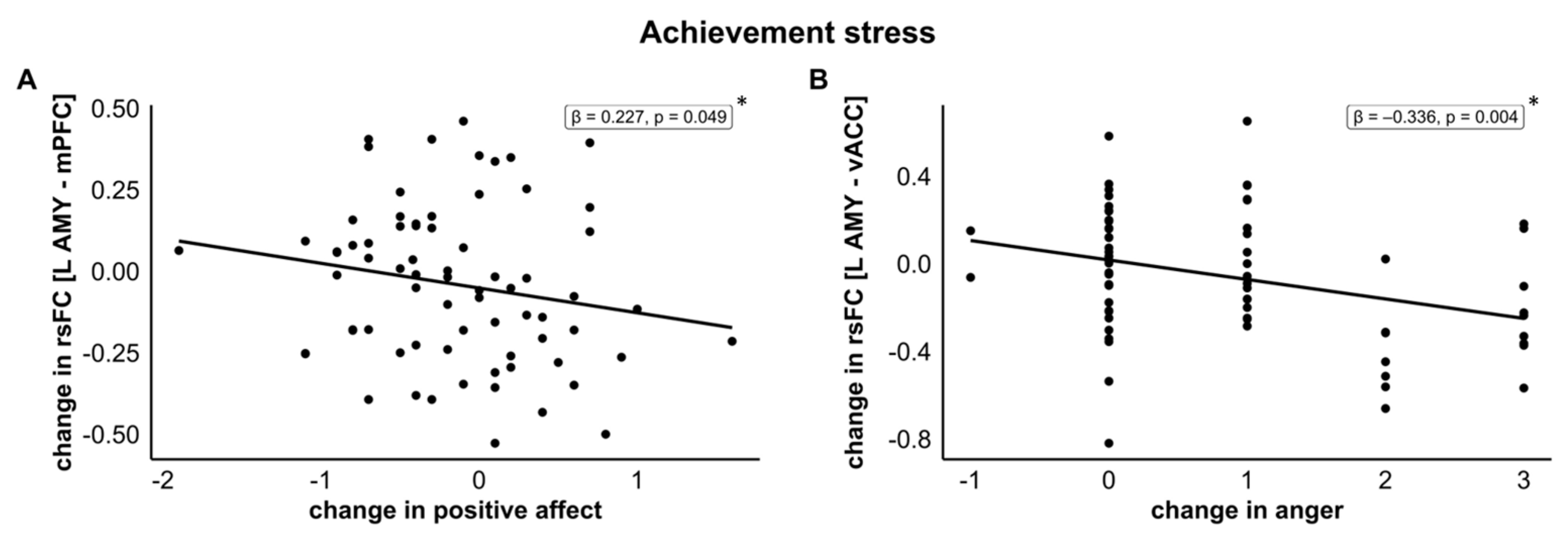
3.5.2. Predictor: Changes in Positive Affect from Pre- to Post-Stress
3.5.3. Predictor: Changes in Negative Affect from Pre- to Post-Stress
3.5.4. Predictor: Changes in Anger from Pre- to Post-Stress
4. Discussion
4.1. Sex Differences in Stress Reaction in Amygdala–Frontal Stress Networks
4.2. Sex-Independent Effects of Stress Reaction on Amygdala–Frontal Stress Networks
4.3. Strengths, Limitations, and Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Social Exclusion | Achievement Stress | |||||||
|---|---|---|---|---|---|---|---|---|
| Females (n = 40) | Males (n = 33) | Females (n = 40) | Males (n = 33) | |||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Cortisol [pg/mL] | ||||||||
| pre-stress | 1767.90 | 807.39 | 2518.82 | 1351.30 | 1826.48 | 922.10 | 2475.12 | 1249.39 |
| post-stress | 1672.45 | 806.46 | 2145.73 | 936.72 | 1950.62 | 1010.68 | 2322.97 | 1200.20 |
| Positive affect [PANAS] | ||||||||
| pre-stress | 2.62 | 0.78 | 2.66 | 0.77 | 2.56 | 0.80 | 2.71 | 0.72 |
| post-stress | 2.36 | 0.83 | 2.42 | 0.88 | 2.45 | 0.76 | 2.51 | 0.75 |
| Negative affect [PANAS] | ||||||||
| pre-stress | 1.19 | 0.29 | 1.17 | 0.25 | 1.22 | 0.35 | 1.21 | 0.25 |
| post-stress | 1.23 | 0.30 | 1.23 | 0.33 | 1.41 | 0.48 | 1.32 | 0.36 |
| Anger [ESR] | ||||||||
| pre-stress | 1.08 | 0.35 | 1.24 | 0.56 | 1.10 | 0.30 | 1.12 | 0.33 |
| post-stress | 1.25 | 0.54 | 1.48 | 0.80 | 1.90 | 1.13 | 1.88 | 1.05 |
References
- Dickerson, S.S.; Kemeny, M.E. Acute Stressors and Cortisol Responses: A Theoretical Integration and Synthesis of Laboratory Research. Psychol. Bull. 2004, 130, 355–391. [Google Scholar] [CrossRef] [Green Version]
- Williams, K.D.; Jarvis, B. Cyberball: A Program for Use in Research on Interpersonal Ostracism and Acceptance. Behav. Res. Methods 2006, 38, 174–180. [Google Scholar] [CrossRef] [Green Version]
- Dedovic, K.; Renwick, R.; Mahani, N.K.; Engert, V.; Lupien, S.J.; Pruessner, J.C. The Montreal Imaging Stress Task: Using Functional Imaging to Investigate the Effects of Perceiving and Processing Psychosocial Stress in the Human Brain. J. Psychiatry Neurosci. 2005, 30, 319–325. [Google Scholar]
- Clemens, B.; Wagels, L.; Bauchmüller, M.; Bergs, R.; Habel, U.; Kohn, N. Alerted Default Mode: Functional Connectivity Changes in the Aftermath of Social Stress. Sci. Rep. 2017, 7, 40180. [Google Scholar] [CrossRef] [Green Version]
- Kogler, L.; Müller, V.I.; Chang, A.; Eickhoff, S.B.; Fox, P.T.; Gur, R.C.; Derntl, B. Psychosocial versus Physiological Stress—Meta-Analyses on Deactivations and Activations of the Neural Correlates of Stress Reactions. NeuroImage 2015, 119, 235–251. [Google Scholar] [CrossRef] [Green Version]
- De Raedt, R.; Hooley, J.M. The Role of Expectancy and Proactive Control in Stress Regulation: A Neurocognitive Framework for Regulation Expectation. Clin. Psychol. Rev. 2016, 45, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Hermans, E.J.; Battaglia, F.P.; Atsak, P.; De Voogd, L.D.; Fernández, G.; Roozendaal, B. How the Amygdala Affects Emotional Memory by Altering Brain Network Properties. Neurobiol. Learn. Mem. 2014, 112, 2–16. [Google Scholar] [CrossRef]
- Koenigs, M.; Grafman, J. The Functional Neuroanatomy of Depression: Distinct Roles for Ventromedial and Dorsolateral Prefrontal Cortex. Behav. Brain Res. 2009, 201, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacioppo, S.; Frum, C.; Asp, E.; Weiss, R.M.; Lewis, J.W.; Cacioppo, J.T. A Quantitative Meta-Analysis of Functional Imaging Studies of Social Rejection. Sci. Rep. 2013, 3, 2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veer, I.M.; Oei, N.Y.L.; Spinhoven, P.; van Buchem, M.A.; Elzinga, B.M.; Rombouts, S.A.R.B. Beyond Acute Social Stress: Increased Functional Connectivity between Amygdala and Cortical Midline Structures. NeuroImage 2011, 57, 1534–1541. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, P.L.; Wittmann, M.K. Ventral Anterior Cingulate Cortex and Social Decision-Making. Neurosci. Biobehav. Rev. 2018, 92, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.X.; Elger, C.E.; Weber, B. Amygdala Tractography Predicts Functional Connectivity and Learning during Feedback-Guided Decision-Making. NeuroImage 2008, 39, 1396–1407. [Google Scholar] [CrossRef] [PubMed]
- Quaedflieg, C.W.E.M.; Van De Ven, V.; Meyer, T.; Siep, N.; Merckelbach, H.; Smeets, T. Temporal Dynamics of Stress-Induced Alternations of Intrinsic Amygdala Connectivity and Neuroendocrine Levels. PLoS ONE 2015, 10, e0124141. [Google Scholar] [CrossRef] [Green Version]
- Henckens, M.J.A.G.; Van Wingen, G.A.; Joëls, M.; Fernández, G. Time-Dependent Effects of Corticosteroids on Human Amygdala Processing. J. Neurosci. 2010, 30, 12725–12732. [Google Scholar] [CrossRef] [Green Version]
- Rose, A.J.; Rudolph, K.D. A Review of Sex Differences in Peer Relationship Processes: Potential Trade-Offs for the Emotional and Behavioral Development of Girls and Boys. Psychol. Bull. 2006, 132, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stroud, L.R.; Salovey, P.; Epel, E.S. Sex Differences in Stress Responses: Social Rejection versus Achievement Stress. Biol. Psychiatry 2002, 52, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Bangasser, D.A.; Valentino, R.J. Sex Differences in Stress-Related Psychiatric Disorders: Neurobiological Perspectives. Front. Neuroendocrinol. 2014, 35, 303–319. [Google Scholar] [CrossRef] [Green Version]
- Heck, A.L.; Handa, R.J. Sex Differences in the Hypothalamic–Pituitary–Adrenal Axis’ Response to Stress: An Important Role for Gonadal Hormones. Neuropsychopharmacology 2019, 44, 45. [Google Scholar] [CrossRef] [Green Version]
- Li, S.H.; Graham, B.M. Why Are Women so Vulnerable to Anxiety, Trauma-Related and Stress-Related Disorders? The Potential Role of Sex Hormones. Lancet Psychiatry 2017, 4, 73–82. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Bairey Merz, N.; Barnes, P.J.; Brinton, R.D.; Carrero, J.J.; DeMeo, D.L.; De Vries, G.J.; Epperson, C.N.; Govindan, R.; Klein, S.L.; et al. Sex and Gender: Modifiers of Health, Disease, and Medicine. Lancet 2020, 396, 565. [Google Scholar] [CrossRef]
- Kuehner, C. Why Is Depression More Common among Women than among Men? Lancet Psychiatry 2017, 4, 146–158. [Google Scholar] [CrossRef]
- Alarcón, G.; Cservenka, A.; Rudolph, M.D.; Fair, D.A.; Nagel, B.J. Developmental Sex Differences in Resting State Functional Connectivity of Amygdala Sub-Regions. NeuroImage 2015, 115, 235. [Google Scholar] [CrossRef] [Green Version]
- Engman, J.; Linnman, C.; Van Dijk, K.R.A.; Milad, M.R. Amygdala Subnuclei Resting-State Functional Connectivity Sex and Estrogen Differences. Psychoneuroendocrinology 2016, 63, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Kogler, L.; Müller, V.I.; Seidel, E.M.; Boubela, R.; Kalcher, K.; Moser, E.; Habel, U.; Gur, R.C.; Eickhoff, S.B.; Derntl, B. Sex Differences in the Functional Connectivity of the Amygdalae in Association with Cortisol. NeuroImage 2016, 134, 410–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G* Power 3: A Flexible Statistical Power Analysis Program for the Social, Behavioral, and Biomedical Sciences. Behav Res Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Wittchen, H.; Wunderlich, U.; Gruschwitz, S.; Zaudig, M. Skid i. Strukturiertes Klinisches Interview Für Dsm-Iv. Achse i: Psychische Störungen. Interviewheft und Beurteilungsheft; Verlag für Psychologie Hogrefe: Göttingen, Germany, 1997. [Google Scholar]
- Merz, J.; Lehrl, S.; Galster, V.; Erzigkeit, H. MWT-B—Ein Intelligenzkurztest [The Multiple Selection Vocabulary Test (MSVT-B)—An Accelerated Intelligence Test]. Psychiatr. Neurol. Med. Psychol. 1975, 27, 423–428. [Google Scholar]
- Berenson, K.R.; Gyurak, A.; Ayduk, Ö.; Downey, G.; Garner, M.J.; Mogg, K.; Bradley, B.P.; Pine, D.S. Rejection Sensitivity and Disruption of Attention by Social Threat Cues. J. Res. Personal. 2009, 43, 1064–1072. [Google Scholar] [CrossRef] [Green Version]
- Janke, W.; Erdmann, G.; Boucsein, W. Stressverarbeitungsfragebogen: (SVF); Verlag für Psychologie Hogrefe: Göttingen, Germany, 1985. [Google Scholar]
- Krohne, H.W.; Egloff, B.; Kohlmann, C.-W.; Tausch, A. Untersuchungen Mit Einer Deutschen Version der “Positive and Negative Affect Schedule“ (PANAS). Diagn. Gott. 1996, 42, 139–156. [Google Scholar] [CrossRef]
- Schneider, F.; Gur, R.C.; Gur, R.E.; Muenz, L.R. Standardized Mood Induction with Happy and Sad Facial Expressions. Psychiatry Res. 1994, 51, 19–31. [Google Scholar] [CrossRef]
- Kogler, L.; Seidel, E.M.; Metzler, H.; Thaler, H.; Boubela, R.N.; Pruessner, J.C.; Kryspin-Exner, I.; Gur, R.C.; Windischberger, C.; Moser, E.; et al. Impact of Self-Esteem and Sex on Stress Reactions. Sci. Rep. 2017, 7, e17210. [Google Scholar] [CrossRef] [Green Version]
- Radke, S.; Seidel, E.M.; Boubela, R.N.; Thaler, H.; Metzler, H.; Kryspin-Exner, I.; Moser, E.; Habel, U.; Derntl, B. Immediate and Delayed Neuroendocrine Responses to Social Exclusion in Males and Females. Psychoneuroendocrinology 2018, 93, 56–64. [Google Scholar] [CrossRef]
- Novembre, G.; Zanon, M.; Silani, G. Empathy for Social Exclusion Involves the Sensory-Discriminative Component of Pain: A within-Subject FMRI Study. Soc. Cogn. Affect. Neurosci. 2015, 10, 153–164. [Google Scholar] [CrossRef] [PubMed]
- van Marle, H.J.F.; Hermans, E.J.; Qin, S.; Fernández, G. Enhanced Resting-State Connectivity of Amygdala in the Immediate Aftermath of Acute Psychological Stress. NeuroImage 2010, 53, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Bolling, D.Z.; Pitskel, N.B.; Deen, B.; Crowley, M.J.; McPartland, J.C.; Mayes, L.C.; Pelphrey, K.A. Dissociable Brain Mechanisms for Processing Social Exclusion and Rule Violation. NeuroImage 2011, 54, 2462–2471. [Google Scholar] [CrossRef] [Green Version]
- Vaisvaser, S.; Lin, T.; Admon, R.; Podlipsky, I.; Greenman, Y.; Stern, N.; Fruchter, E.; Wald, I.; Pine, D.S.; Tarrasch, R.; et al. Neural Traces of Stress: Cortisol Related Sustained Enhancement of Amygdala-Hippocampal Functional Connectivity. Front. Hum. Neurosci. 2013, 7, 313. [Google Scholar] [CrossRef] [Green Version]
- Nostro, A.D.; Müller, V.I.; Varikuti, D.P.; Pläschke, R.N.; Hoffstaedter, F.; Langner, R.; Patil, K.R.; Eickhoff, S.B. Predicting Personality from Network-Based Resting-State Functional Connectivity. Brain Struct. Funct. 2018, 223, 2699–2719. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.; Pripfl, J.; Bauer, H.; Moser, E. The Impact of EPI Voxel Size on SNR and BOLD Sensitivity in the Anterior Medio-Temporal Lobe: A Comparative Group Study of Deactivation of the Default Mode. Magn. Reson. Mater. Phys. Biol. Med. 2008, 21, 279–290. [Google Scholar] [CrossRef]
- Robinson, S.; Moser, E.; Peper, M. FMRI of Emotion. Neuromethods 2009, 41, 411–456. [Google Scholar] [CrossRef]
- Holmes, C.J.; Hoge, R.; Collins, L.; Woods, R.; Toga, A.W.; Evans, A.C. Enhancement of MR Images Using Registration for Signal Averaging. J. Comput. Assist. Tomogr. 1998, 22, 324–333. [Google Scholar] [CrossRef]
- Ashburner, J.; Friston, K.J. Unified Segmentation. NeuroImage 2005, 26, 839–851. [Google Scholar] [CrossRef]
- Weissenbacher, A.; Kasess, C.; Gerstl, F.; Lanzenberger, R.; Moser, E.; Windischberger, C. Correlations and Anticorrelations in Resting-State Functional Connectivity MRI: A Quantitative Comparison of Preprocessing Strategies. NeuroImage 2009, 47, 1408–1416. [Google Scholar] [CrossRef]
- Satterthwaite, T.D.; Elliott, M.A.; Gerraty, R.T.; Ruparel, K.; Loughead, J.; Calkins, M.E.; Eickhoff, S.B.; Hakonarson, H.; Gur, R.C.; Gur, R.E.; et al. An Improved Framework for Confound Regression and Filtering for Control of Motion Artifact in the Preprocessing of Resting-State Functional Connectivity Data. NeuroImage 2013, 64, 240–256. [Google Scholar] [CrossRef] [PubMed]
- Correll, J.; Mellinger, C.; Pedersen, E.J. Flexible Approaches for Estimating Partial Eta Squared in Mixed-Effects Models with Crossed Random Factors. Behav. Res. 2022, 54, 1626–1642. [Google Scholar] [CrossRef]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but Systematic Correlations in Functional Connectivity MRI Networks Arise from Subject Motion. NeuroImage 2012, 59, 2142–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleshchova, O.; Rieder, J.K.; Grinband, J.; Weierich, M.R. Resting Amygdala Connectivity and Basal Sympathetic Tone as Markers of Chronic Hypervigilance. Psychoneuroendocrinology 2019, 102, 68–78. [Google Scholar] [CrossRef]
- Costafreda, S.G.; McCann, P.; Saker, P.; Cole, J.H.; Cohen-Woods, S.; Farmer, A.E.; Aitchison, K.J.; McGuffin, P.; Fu, C.H.Y. Modulation of Amygdala Response and Connectivity in Depression by Serotonin Transporter Polymorphism and Diagnosis. J. Affect. Disord. 2013, 150, 96–103. [Google Scholar] [CrossRef]
- Kanske, P.; Kotz, S.A. Emotion Speeds up Conflict Resolution: A New Role for the Ventral Anterior Cingulate Cortex? Cereb. Cortex 2011, 21, 911–919. [Google Scholar] [CrossRef] [Green Version]
- Stevens, F.L.; Hurley, R.A.; Taber, K.H.; Hurley, R.A.; Hayman, L.A.; Taber, K.H. Anterior Cingulate Cortex: Unique Role in Cognition and Emotion. JNP 2011, 23, 121–125. [Google Scholar] [CrossRef]
- Drevets, W.C.; Savitz, J.; Trimble, M. The Subgenual Anterior Cingulate Cortex in Mood Disorders. CNS Spectr. 2008, 13, 663–681. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.; Fadok, R.A.; Purcell, M.; Liu, S.; Stonnington, C.; Spetzler, R.F.; Baxter, L.C. Localizing Sadness Activation within the Subgenual Cingulate in Individuals: A Novel Functional MRI Paradigm for Detecting Individual Differences in the Neural Circuitry Underlying Depression. Brain Imaging Behav. 2011, 5, 229–239. [Google Scholar] [CrossRef]
- Liotti, M.; Mayberg, H.S.; Brannan, S.K.; McGinnis, S.; Jerabek, P.; Fox, P.T. Differential Limbic—Cortical Correlates of Sadness and Anxiety in Healthy Subjects: Implications for Affective Disorders. Biol. Psychiatry 2000, 48, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Coombs, G.; Loggia, M.L.; Greve, D.N.; Holt, D.J. Amygdala Perfusion Is Predicted by Its Functional Connectivity with the Ventromedial Prefrontal Cortex and Negative Affect. PLoS ONE 2014, 9, e9746. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Yang, J. Reward Anticipation Buffers Neuroendocrine and Cardiovascular Responses to Acute Psychosocial Stress in Healthy Young Adults. Stress 2021, 24, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov-Discher, A.; Wenzel, J.; Kabisch, N.; Hemmerling, J.; Bunz, M.; Schöndorf, J.; Walter, H.; Veer, I.M.; Adli, M. Residential Green Space and Air Pollution Are Associated with Brain Activation in a Social-Stress Paradigm. Sci Rep 2022, 12, 10614. [Google Scholar] [CrossRef]
- Noack, H.; Nolte, L.; Nieratschker, V.; Habel, U.; Derntl, B. Imaging Stress: An Overview of Stress Induction Methods in the MR Scanner. J. Neural. Transm. 2019, 126, 1187–1202. [Google Scholar] [CrossRef]
- Peters, S.; Cleare, A.J.; Papadopoulos, A.; Fu, C.H.Y. Cortisol Responses to Serial MRI Scans in Healthy Adults and in Depression. Psychoneuroendocrinology 2011, 36, 737–741. [Google Scholar] [CrossRef]
- Adam, E.K.; Kumari, M. Assessing Salivary Cortisol in Large-Scale, Epidemiological Research. Psychoneuroendocrinology 2009, 34, 1423–1436. [Google Scholar] [CrossRef]
- Hidalgo-Lopez, E.; Mueller, K.; Harris, T.A.; Aichhorn, M.; Sacher, J.; Pletzer, B. Human Menstrual Cycle Variation in Subcortical Functional Brain Connectivity: A Multimodal Analysis Approach. Brain Struct. Funct. 2020, 225, 591–605. [Google Scholar] [CrossRef] [Green Version]
- Kirschbaum, C.; Kudielka, B.M.; Gaab, J.; Schommer, N.C.; Hellhammer, D.H. Impact of Gender, Menstrual Cycle Phase, and Oral Contraceptives on the Activity of the Hypothalamus-Pituitary-Adrenal Axis. Psychosom. Med. 1999, 61, 154–162. [Google Scholar] [CrossRef]
- Zhang, W.; Llera, A.; Hashemi, M.M.; Kaldewaij, R.; Koch, S.B.J.; Beckmann, C.F.; Klumpers, F.; Roelofs, K. Discriminating Stress from Rest Based on Resting-State Connectivity of the Human Brain: A Supervised Machine Learning Study. Hum. Brain Mapp. 2020, 41, 3089–3099. [Google Scholar] [CrossRef] [Green Version]
- Heidari, S.; Babor, T.F.; De Castro, P.; Tort, S.; Curno, M. Sex and Gender Equity in Research: Rationale for the SAGER Guidelines and Recommended Use. Res. Integr. Peer Rev. 2016, 1, 2. [Google Scholar] [CrossRef] [PubMed]





| MNI Coordinates | X | Y | Z | |
|---|---|---|---|---|
| Amygdala (bilateral) | R | 26 | −6 | −14 |
| L | −24 | −6 | −14 | |
| vACC (unilateral) | −7 | 29 | −12 | |
| mPFC (unilateral) | 3 | 54 | 6 | |
| dlPFC unilateral | R | 37 | 34 | 35 |
| L | −37 | 44 | 37 |
| Females (n = 40) | Males (n = 33) | |||||
|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Statistical Parameters | p-Value | |
| Age (in years) | 24.7 | 3.8 | 24.0 | 3.0 | t(71) = 0.755 | 0.453 |
| Verbal intelligence | 26.7 | 4.0 | 27.4 | 3.5 | t(70 1) = 0.875 | 0.385 |
| Social support in preceding 10 days | 19.3 | 20.5 | 20.2 | 19.2 | t(71) = 0.201 | 0.841 |
| Rejection sensitivity | 8.4 | 3.3 | 8.1 | 2.9 | t(71) = 0.412 | 0.681 |
| Positive stress coping strategies | 2.2 | 0.4 | 2.3 | 0.3 | t(71) = 0.298 | 0.767 |
| Negative stress coping strategies | 2.0 | 0.6 | 1.7 | 0.6 | t(71) = 2.080 | 0.041 * |
| Estradiol levels on arrival [pg/mL] | 4.58 | 4.07 | n/a | n/a | n/a | n/a |
| Progesterone levels on arrival [pg/mL] | 71.97 | 57.61 | 42.65 | 29.48 | t(69 1) = 2.582 | 0.012 * |
| Testosterone levels on arrival [pg/mL] | 25.99 | 19.07 | 87.93 | 60.62 | t(71) = 6.114 | <0.001 * |
| Social Exclusion | Achievement Stress | |||||||
|---|---|---|---|---|---|---|---|---|
| Females (n = 40) | Males (n = 33) | Females (n = 40) | Males (n = 33) | |||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Change in cortisol [pg/mL] | −115.05 | 437.71 | −373.09 | 1021.32 | 124.15 | 630.81 | −152.91 | 909.76 |
| Change in positive affect (PANAS) | −0.26 | 0.55 | −0.24 | 0.57 | −0.11 | 0.61 | −0.21 | 0.58 |
| Change in negative affect (PANAS) | 0.03 | 0.25 | 0.05 | 0.25 | 0.19 | 0.51 | 0.11 | 0.30 |
| Change in anger (ESR) | 0.18 | 0.64 | 0.24 | 0.61 | 0.80 | 1.11 | 0.76 | 1.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bürger, Z.; Müller, V.I.; Hoffstaedter, F.; Habel, U.; Gur, R.C.; Windischberger, C.; Moser, E.; Derntl, B.; Kogler, L. Stressor-Specific Sex Differences in Amygdala–Frontal Cortex Networks. J. Clin. Med. 2023, 12, 865. https://doi.org/10.3390/jcm12030865
Bürger Z, Müller VI, Hoffstaedter F, Habel U, Gur RC, Windischberger C, Moser E, Derntl B, Kogler L. Stressor-Specific Sex Differences in Amygdala–Frontal Cortex Networks. Journal of Clinical Medicine. 2023; 12(3):865. https://doi.org/10.3390/jcm12030865
Chicago/Turabian StyleBürger, Zoé, Veronika I. Müller, Felix Hoffstaedter, Ute Habel, Ruben C. Gur, Christian Windischberger, Ewald Moser, Birgit Derntl, and Lydia Kogler. 2023. "Stressor-Specific Sex Differences in Amygdala–Frontal Cortex Networks" Journal of Clinical Medicine 12, no. 3: 865. https://doi.org/10.3390/jcm12030865