
Expression of Calbindin, a Marker of Gamma-Aminobutyric Acid Neurons, Is Reduced in the Amygdala of Oestrogen Receptor β-Deficient Female Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tissue Preparation
2.3. Immunohistochemistry
2.4. Controls
2.5. Counts and Measurements
2.6. Statistics
3. Results
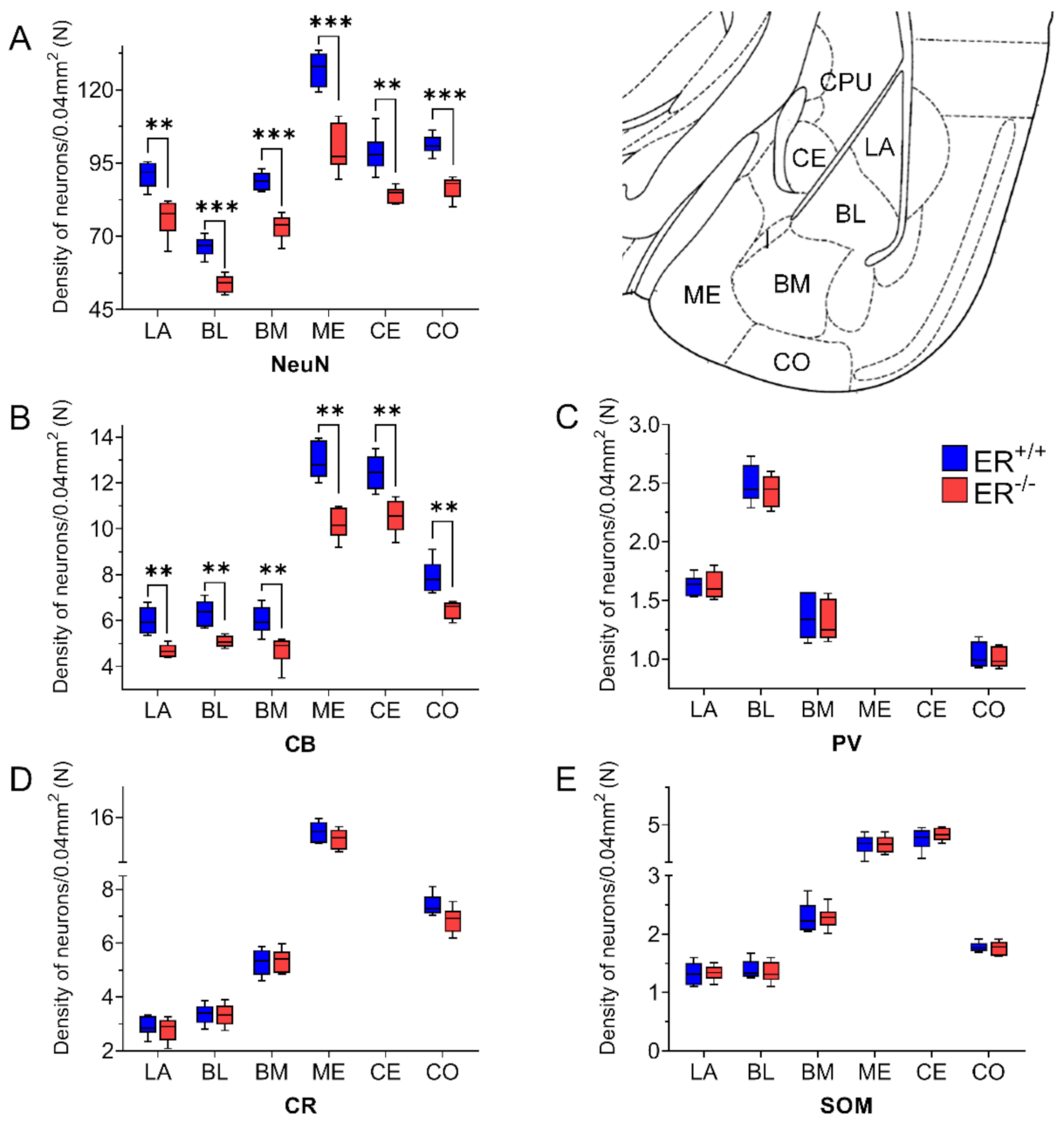
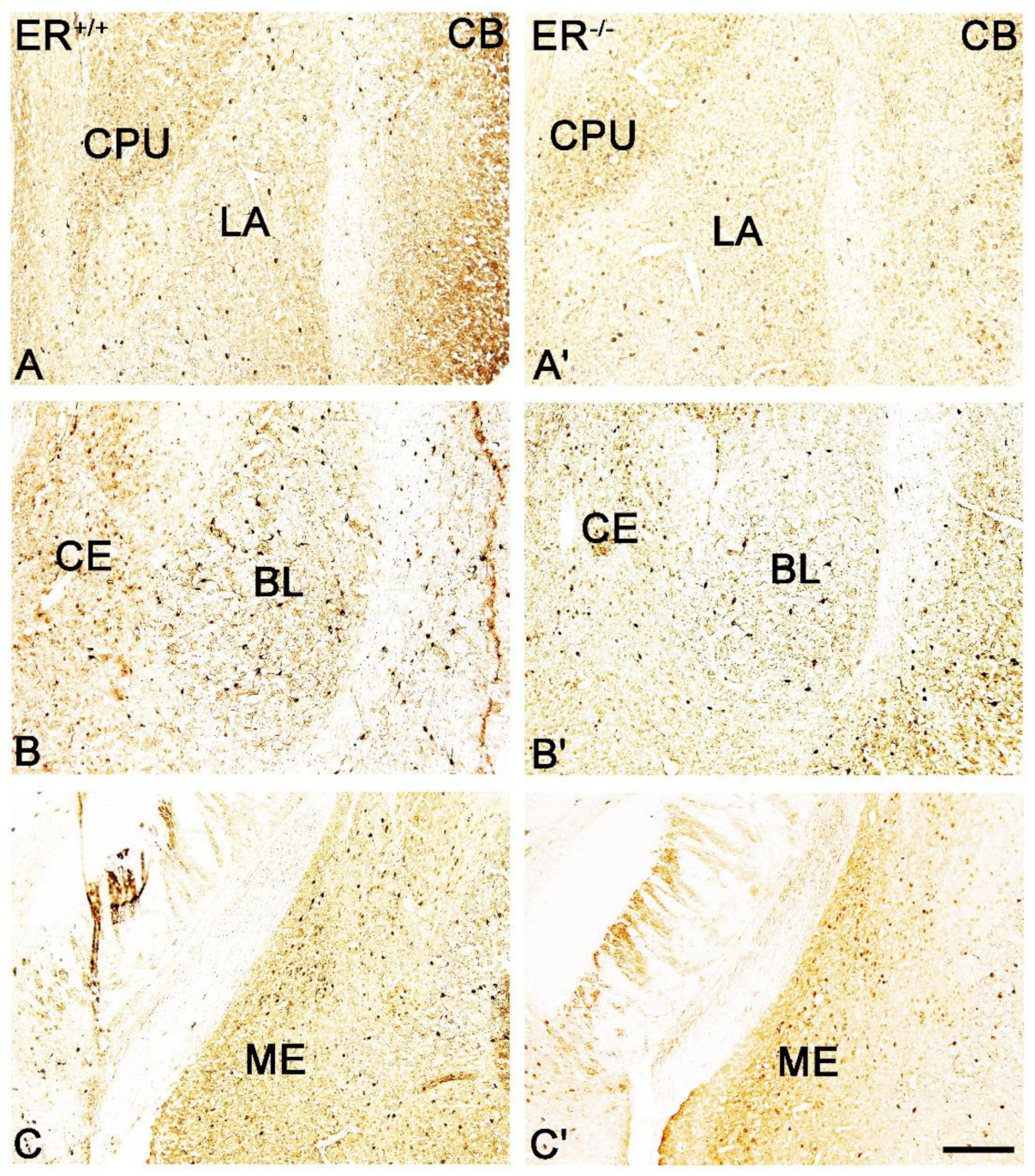
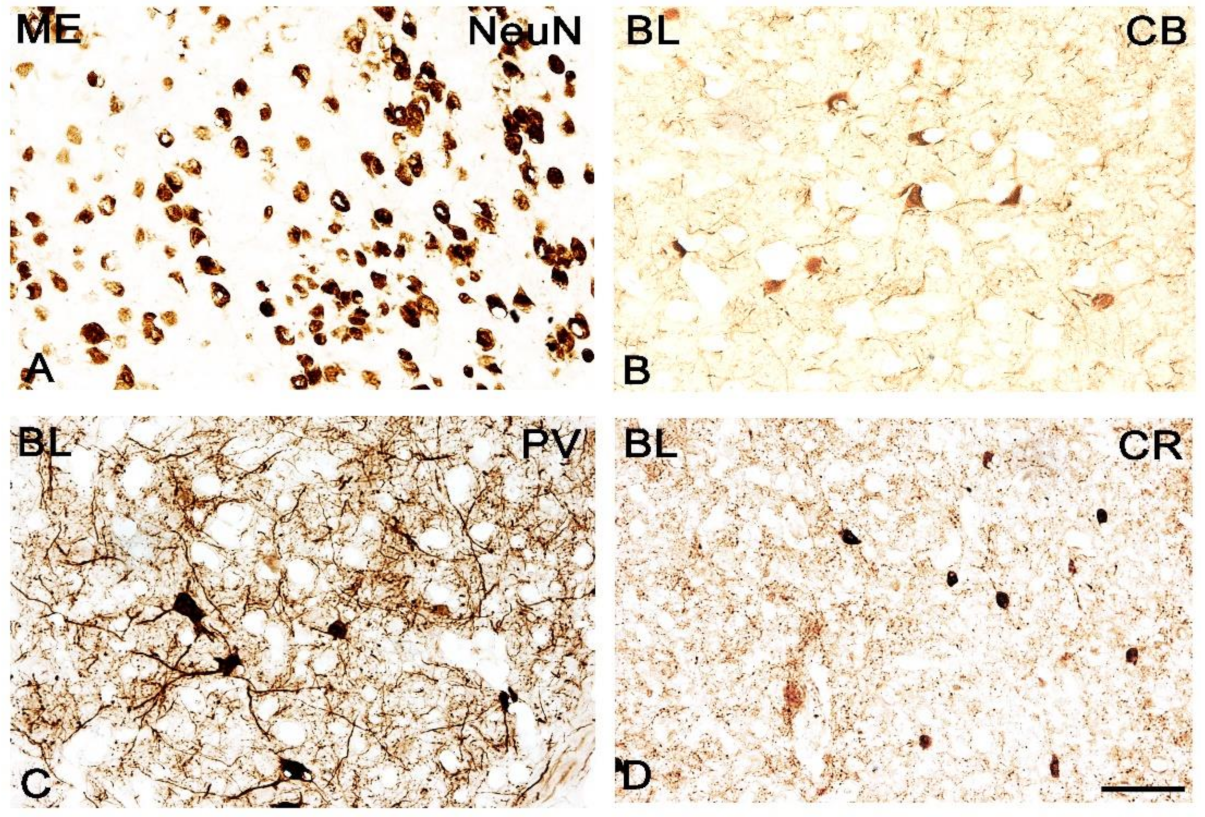
3.1. Neuronal Deficits in the Amygdala of ERβ−/− Mice
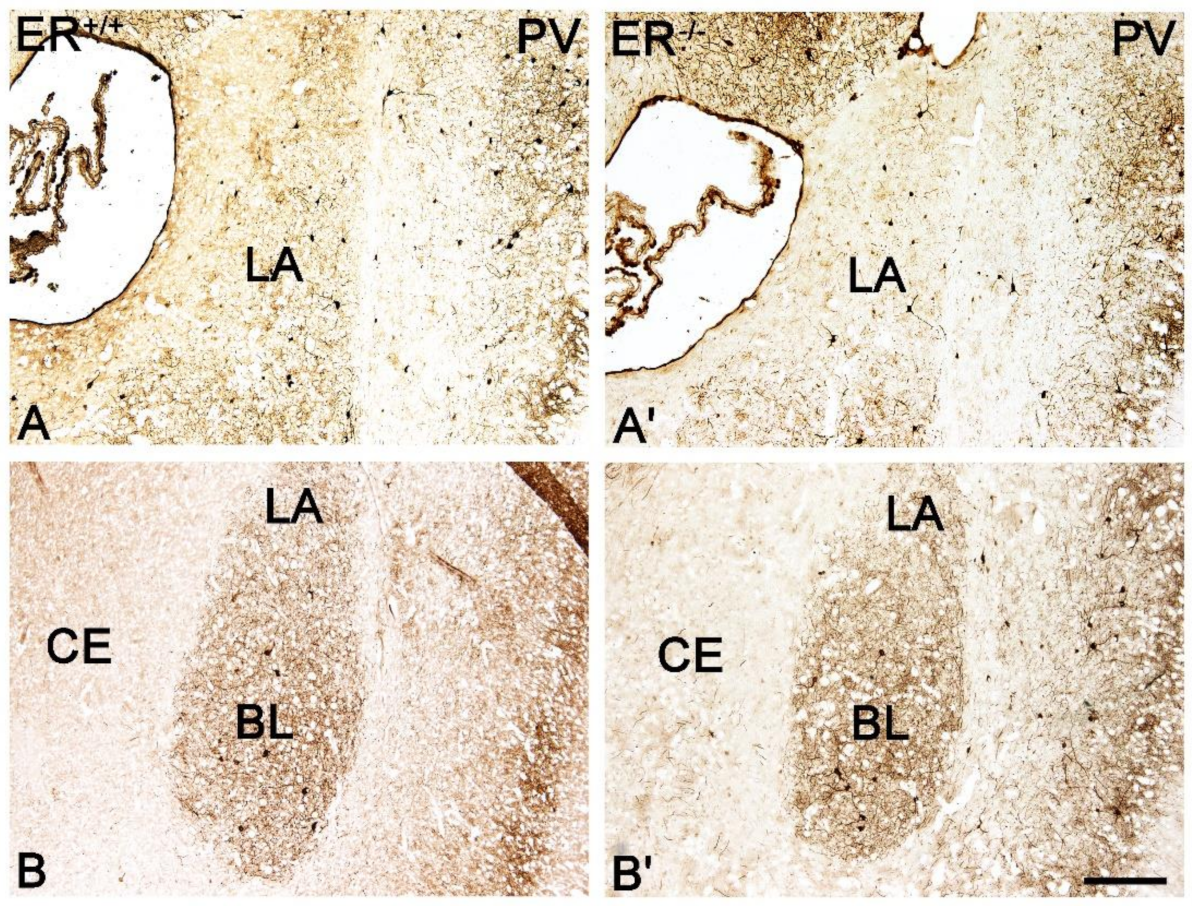
3.2. Calcium-Binding Proteins in the Amygdala of ERβ−/− Mice
3.3. Somatostatin in the Amygdala of ERβ−/− Mice
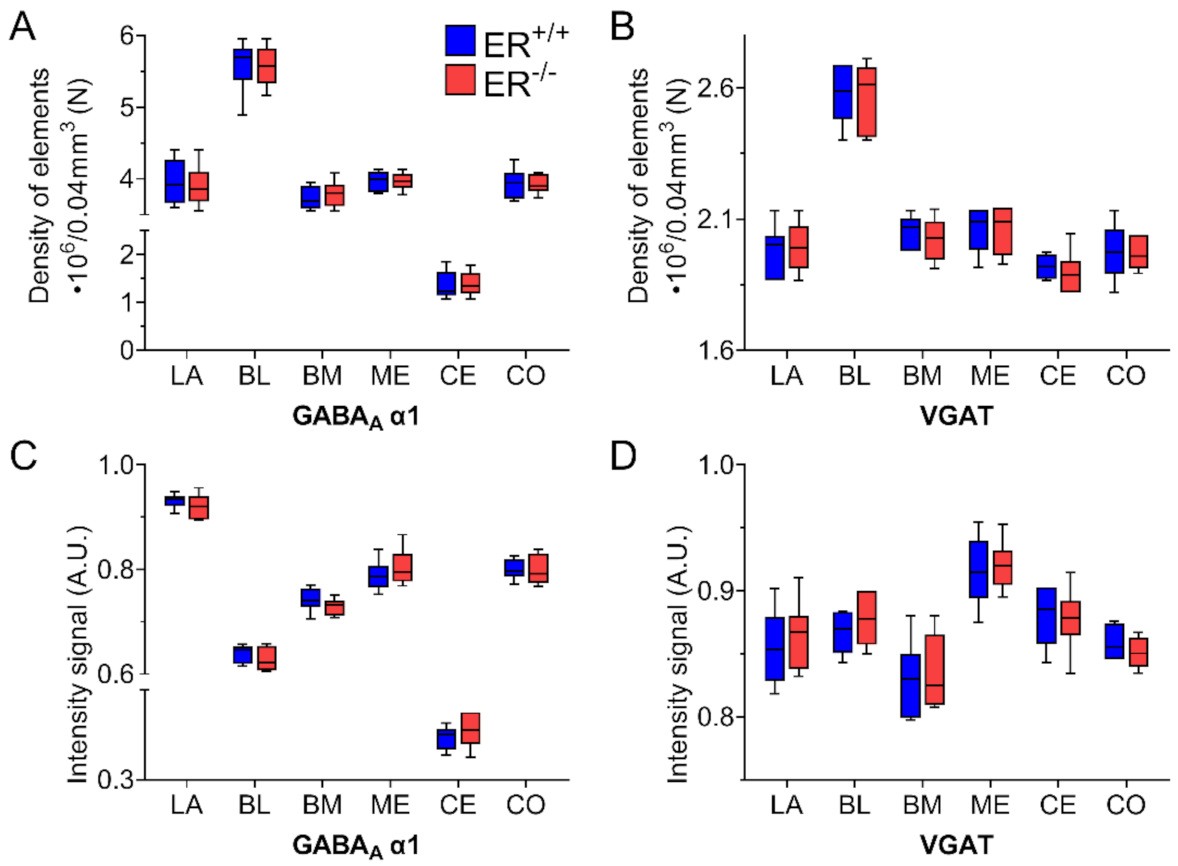
3.4. GABA Type A Receptor with α1 Subunit in the Amygdala of ERβ−/− Mice
3.5. Vesicular GABA Transporter in the Amygdala of ERβ−/− Mice
3.6. Overexpression of Glial Fibrillary Acidic Protein in the Amygdala of ERβ−/− Mice
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kessler, R.C.; Angermeyer, M.; Anthony, J.C.; DE Graaf, R.; Demyttenaere, K.; Gasquet, I.; DE Girolamo, G.; Gluzman, S.; Gureje, O.; Haro, J.M.; et al. Lifetime Prevalence and Age-of-Onset Distributions of Mental Disorders in the World Health Organization’s World Mental Health Survey Initiative. World Psychiatry 2007, 6, 168–176. [Google Scholar] [PubMed]
- Steel, Z.; Marnane, C.; Iranpour, C.; Chey, T.; Jackson, J.W.; Patel, V.; Silove, D. The Global Prevalence of Common Mental Disorders: A Systematic Review and Meta-Analysis 1980–2013. Int. J. Epidemiol. 2014, 43, 476–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craske, M.G.; Stein, M.B. Anxiety. Lancet 2016, 388, 3048–3059. [Google Scholar] [CrossRef]
- Nomura, M.; Durbak, L.; Chan, J.; Smithies, O.; Gustafsson, J.-Å.; Korach, K.S.; Pfaff, D.W.; Ogawa, S. Genotype/Age Interactions on Aggressive Behavior in Gonadally Intact Estrogen Receptor β Knockout (ΒERKO) Male Mice. Horm. Behav. 2002, 41, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Baxter, A.J.; Scott, K.M.; Vos, T.; Whiteford, H.A. Global Prevalence of Anxiety Disorders: A Systematic Review and Meta-Regression. Psychol. Med. 2013, 43, 897–910. [Google Scholar] [CrossRef] [PubMed]
- Bandelow, B.; Michaelis, S. Epidemiology of Anxiety Disorders in the 21st Century. Dialogues Clin. Neurosci. 2015, 17, 327–335. [Google Scholar]
- Ritchie, H.; Roser, M. Mental Health. Our World Data. 2018. Available online: https://ourworldindata.org/mental-health (accessed on 17 March 2021).
- Locke, A.; Kirst, N.; Shultz, C.G. Diagnosis and Management of Generalized Anxiety Disorder and Panic Disorder in Adults. AFP 2015, 91, 617–624. [Google Scholar]
- LeDoux, J. The Emotional Brain, Fear, and the Amygdala. Cell. Mol. Neurobiol. 2003, 23, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Etkin, A.; Prater, K.E.; Schatzberg, A.F.; Menon, V.; Greicius, M.D. Disrupted Amygdalar Subregion Functional Connectivity and Evidence of a Compensatory Network in Generalized Anxiety Disorder. Arch. Gen. Psychiatry 2009, 66, 1361–1372. [Google Scholar] [CrossRef] [Green Version]
- Ramasubbu, R.; Konduru, N.; Cortese, F.; Bray, S.; Gaxiola, I.; Goodyear, B. Reduced Intrinsic Connectivity of Amygdala in Adults with Major Depressive Disorder. Front. Psychiatry 2014, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Cullen, K.R.; Vizueta, N.; Thomas, K.M.; Han, G.J.; Lim, K.O.; Camchong, J.; Mueller, B.A.; Bell, C.H.; Heller, M.D.; Schulz, S.C. Amygdala Functional Connectivity in Young Women with Borderline Personality Disorder. Brain Connect. 2011, 1, 61. [Google Scholar] [CrossRef] [Green Version]
- Buffalari, D.M.; See, R.E. Amygdala Mechanisms of Pavlovian Psychostimulant Conditioning and Relapse. Curr. Top. Behav. Neurosci. 2010, 3, 73–99. [Google Scholar] [CrossRef]
- Swaab, D.F.; Chung, W.C.J.; Kruijver, F.P.M.; Hofman, M.A.; Hestiantoro, A. Sex Differences in the Hypothalamus in the Different Stages of Human Life. Neurobiol. Aging 2003, 24 (Suppl. 1), S1–S16; discussion S17–S19. [Google Scholar] [CrossRef]
- Riecher-Rössler, A. Prospects for the Classification of Mental Disorders in Women. Eur. Psychiatry 2010, 25, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalal, P.K.; Agarwal, M. Postmenopausal Syndrome. Indian J. Psychiatry 2015, 57, S222–S232. [Google Scholar] [CrossRef] [PubMed]
- Marsh, W.K.; Bromberger, J.T.; Crawford, S.L.; Leung, K.; Kravitz, H.M.; Randolph, J.F.; Joffe, H.; Soares, C.N. Lifelong Estradiol Exposure and Risk of Depressive Symptoms during the Transition to Menopause and Postmenopause. Menopause 2017, 24, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Joffe, H.; Cohen, L.S. Estrogen, Serotonin, and Mood Disturbance: Where Is the Therapeutic Bridge? Biol.. Psychiatry 1998, 44, 798–811. [Google Scholar] [CrossRef]
- O’Bryant, S.E.; Palav, A.; McCaffrey, R.J. A Review of Symptoms Commonly Associated with Menopause: Implications for Clinical Neuropsychologists and Other Health Care Providers. Neuropsychol. Rev. 2003, 13, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.N.; Almeida, O.P.; Joffe, H.; Cohen, L.S. Efficacy of Estradiol for the Treatment of Depressive Disorders in Perimenopausal Women: A Double-Blind, Randomized, Placebo-Controlled Trial. Arch. Gen. Psychiatry 2001, 58, 529–534. [Google Scholar] [CrossRef] [Green Version]
- Bromberger, J.T.; Kravitz, H.M.; Chang, Y.; Randolph, J.F.; Avis, N.E.; Gold, E.B.; Matthews, K.A. Does Risk for Anxiety Increase during the Menopausal Transition? Study of Women’s Health Across the Nation. Menopause J. N. Am. Menopause Soc. 2013, 20, 488. [Google Scholar] [CrossRef] [Green Version]
- Hoyt, L.T.; Falconi, A. Puberty and Perimenopause: Reproductive Transitions and Their Implications for Women’s Health. Soc. Sci. Med. 2015, 132, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.R.; Sen, S.; Diepenhorst, L.L.; Rudick, C.N.; Maren, S. Estrogen Modulates Sexually Dimorphic Contextual Fear Conditioning and Hippocampal Long-Term Potentiation (LTP) in Rats(1). Brain Res. 2001, 888, 356–365. [Google Scholar] [CrossRef]
- Krȩżel, W.; Dupont, S.; Krust, A.; Chambon, P.; Chapman, P.F. Increased Anxiety and Synaptic Plasticity in Estrogen Receptor β-Deficient Mice. Proc. Natl. Acad. Sci. USA 2001, 98, 12278–12282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lund, T.D.; Rovis, T.; Chung, W.C.J.; Handa, R.J. Novel Actions of Estrogen Receptor-β on Anxiety-Related Behaviors. Endocrinology 2005, 146, 797–807. [Google Scholar] [CrossRef]
- Oyola, M.G.; Portillo, W.; Reyna, A.; Foradori, C.D.; Kudwa, A.; Hinds, L.; Handa, R.J.; Mani, S.K. Anxiolytic Effects and Neuroanatomical Targets of Estrogen Receptor-β (ERβ) Activation by a Selective ERβ Agonist in Female Mice. Endocrinology 2012, 153, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Andersson, S.; Warner, M.; Gustafsson, J.-A. Morphological Abnormalities in the Brains of Estrogen Receptor Knockout Mice. Proc. Natl. Acad. Sci. USA 2001, 98, 2792–2796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, Z.; Tan, L.; Wang, Y.; Lu, C.; Chen, R.; Zhang, S.; Gao, Y.; Liu, Y.; Yin, Y.; et al. Correcting MiR92a-VGAT-Mediated GABAergic Dysfunctions Rescues Human Tau-Induced Anxiety in Mice. Mol. Ther. 2017, 25, 140–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, S.; Kubo, C.; Rhee, J.-S.; Akaike, N. Presynaptic Serotonergic Inhibition of GABAergic Synaptic Transmission in Mechanically Dissociated Rat Basolateral Amygdala Neurons. J. Physiol. 1999, 518, 525–538. [Google Scholar] [CrossRef]
- Sibille, E.; Pavlides, C.; Benke, D.; Toth, M. Genetic Inactivation of the Serotonin 1A Receptor in Mice Results in Downregulation of Major GABA A Receptor α Subunits, Reduction of GABA A Receptor Binding, and Benzodiazepine-Resistant Anxiety. J. Neurosci. 2000, 20, 2758–2765. [Google Scholar] [CrossRef] [Green Version]
- Stutzmann, G.E.; LeDoux, J.E. GABAergic Antagonists Block the Inhibitory Effects of Serotonin in the Lateral Amygdala: A Mechanism for Modulation of Sensory Inputs Related to Fear Conditioning. J. Neurosci. 1999, 19, RC8. [Google Scholar] [CrossRef] [PubMed]
- Gundlah, C.; Pecins-Thompson, M.; Schutzer, W.E.; Bethea, C.L. Ovarian Steroid Effects on Serotonin 1A, 2A and 2C Receptor MRNA in Macaque Hypothalamus. Mol. Brain Res. 1999, 63, 325–339. [Google Scholar] [CrossRef]
- Osterlund, M.K.; Halldin, C.; Hurd, Y.L. Effects of Chronic 17beta-Estradiol Treatment on the Serotonin 5-HT(1A) Receptor MRNA and Binding Levels in the Rat Brain. Synapse 2000, 35, 39–44. [Google Scholar] [CrossRef]
- Pitkänen, A.; Amaral, D.G. The Distribution of GABAergic Cells, Fibers, and Terminals in the Monkey Amygdaloid Complex: An Immunohistochemical and in Situ Hybridization Study. J. Neurosci. 1994, 14, 2200–2224. [Google Scholar] [CrossRef] [PubMed]
- Capogna, M. GABAergic Cell Type Diversity in the Basolateral Amygdala. Curr. Opin. Neurobiol. 2014, 26, 110–116. [Google Scholar] [CrossRef]
- Prager, E.M.; Bergstrom, H.C.; Wynn, G.H.; Braga, M.F.M. The Basolateral Amygdala GABAergic System in Health and Disease. J. Neurosci. Res. 2016, 94, 548–567. [Google Scholar] [CrossRef] [Green Version]
- Kemppainen, S.; Pitkänen, A. Distribution of Parvalbumin, Calretinin, and Calbindin-D(28k) Immunoreactivity in the Rat Amygdaloid Complex and Colocalization with Gamma-Aminobutyric Acid. J. Comp. Neurol. 2000, 426, 441–467. [Google Scholar] [CrossRef]
- McDonald, A.J.; Mascagni, F. Colocalization of Calcium-Binding Proteins and GABA in Neurons of the Rat Basolateral Amygdala. Neuroscience 2001, 105, 681–693. [Google Scholar] [CrossRef]
- Mascagni, F.; Muly, E.C.; Rainnie, D.G.; McDonald, A.J. Immunohistochemical Characterization of Parvalbumin-Containing Interneurons in the Monkey Basolateral Amygdala. Neuroscience 2009, 158, 1541–1550. [Google Scholar] [CrossRef] [Green Version]
- Sorvari, H.; Soininen, H.; Paljärvi, L.; Karkola, K.; Pitkänen, A. Distribution of Parvalbumin-Immunoreactive Cells and Fibers in the Human Amygdaloid Complex. J. Comp. Neurol. 1995, 360, 185–212. [Google Scholar] [CrossRef] [PubMed]
- Sorvari, H.; Miettinen, R.; Soininen, H.; Pitkänen, A. Parvalbumin-Immunoreactive Neurons Make Inhibitory Synapses on Pyramidal Cells in the Human Amygdala: A Light and Electron Microscopic Study. Neurosci. Lett. 1996, 217, 93–96. [Google Scholar] [CrossRef]
- Woodruff, A.R.; Sah, P. Inhibition and Synchronization of Basal Amygdala Principal Neuron Spiking by Parvalbumin-Positive Interneurons. J. Neurophysiol. 2007, 98, 2956–2961. [Google Scholar] [CrossRef] [Green Version]
- Muller, J.F.; Mascagni, F.; McDonald, A.J. Postsynaptic Targets of Somatostatin-Containing Interneurons in the Rat Basolateral Amygdala. J. Comp. Neurol. 2007, 500, 513–529. [Google Scholar] [CrossRef]
- Gulyás, A.I.; Hájos, N.; Freund, T.F. Interneurons Containing Calretinin Are Specialized to Control Other Interneurons in the Rat Hippocampus. J. Neurosci. 1996, 16, 3397–3411. [Google Scholar] [CrossRef]
- Melchitzky, D.S.; Lewis, D.A. Dendritic-Targeting GABA Neurons in Monkey Prefrontal Cortex: Comparison of Somatostatin- and Calretinin-Immunoreactive Axon Terminals. Synapse 2008, 62, 456–465. [Google Scholar] [CrossRef] [Green Version]
- Meskenaite, V. Calretinin-Immunoreactive Local Circuit Neurons in Area 17 of the Cynomolgus Monkey, Macaca Fascicularis. J. Comp. Neurol. 1997, 379, 113–132. [Google Scholar] [CrossRef]
- Gonchar, Y.; Burkhalter, A. Connectivity of GABAergic Calretinin-Immunoreactive Neurons in Rat Primary Visual Cortex. Cereb. Cortex 1999, 9, 683–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kritzer, M.F. Regional, Laminar and Cellular Distribution of Immunoreactivity for ER in the Cerebral Cortex of Hormonally Intact, Postnatally Developing Male and Female Rats. Cereb. Cortex 2005, 16, 1181–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Równiak, M. The Neurons Expressing Calcium-Binding Proteins in the Amygdala of the Guinea Pig: Precisely Designed Interface for Sex Hormones. Brain Struct. Funct. 2017, 222, 3775–3793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.-Y.; Zeng, P.; Qu, N.; Ning, L.-N.; Chu, J.; Zhang, T.; Zhou, X.-W.; Tian, Q. Evidence of Altered Depression and Dementia-Related Proteins in the Brains of Young Rats after Ovariectomy. J. Neurochem. 2018, 146, 703–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walf, A.A.; Koonce, C.J.; Frye, C.A. Estradiol or Diarylpropionitrile Decrease Anxiety-like Behavior of Wildtype, but Not Estrogen Receptor Beta Knockout, Mice. Behav. Neurosci. 2008, 122, 974–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, A.J.; Mascagni, F. Immunohistochemical Characterization of Somatostatin Containing Interneurons in the Rat Basolateral Amygdala. Brain Res. 2002, 943, 237–244. [Google Scholar] [CrossRef]
- Dombret, C.; Naulé, L.; Trouillet, A.-C.; Parmentier, C.; Hardin-Pouzet, H.; Mhaouty-Kodja, S. Effects of Neural Estrogen Receptor Beta Deletion on Social and Mood-Related Behaviors and Underlying Mechanisms in Male Mice. Sci. Rep. 2020, 10, 6242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, M.E.; Vandenput, L.; Tivesten, Å.; Norlén, A.-K.; Lagerquist, M.K.; Windahl, S.H.; Börjesson, A.E.; Farman, H.H.; Poutanen, M.; Benrick, A.; et al. Measurement of a Comprehensive Sex Steroid Profile in Rodent Serum by High-Sensitive Gas Chromatography-Tandem Mass Spectrometry. Endocrinology 2015, 156, 2492–2502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Równiak, M.; Bogus-Nowakowska, K.; Robak, A. The Densities of Calbindin and Parvalbumin, but Not Calretinin Neurons, Are Sexually Dimorphic in the Amygdala of the Guinea Pig. Brain Res. 2015, 1604, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Bentea, E.; Van der Perren, A.; Van Liefferinge, J.; El Arfani, A.; Albertini, G.; Demuyser, T.; Merckx, E.; Michotte, Y.; Smolders, I.; Baekelandt, V.; et al. Nigral Proteasome Inhibition in Mice Leads to Motor and Non-Motor Deficits and Increased Expression of Ser129 Phosphorylated α-Synuclein. Front. Behav. Neurosci. 2015, 9, 68. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Chen, Y.; Shi, L.; Liu, D.; Liu, Y.; Yuan, T.; Zhang, X.; Zhu, G.; Zhang, J. Deep Brain Stimulation of the Anterior Nuclei of the Thalamus Relieves Basal Ganglia Dysfunction in Monkeys with Temporal Lobe Epilepsy. CNS Neurosci. Ther. 2021, 27, 341–351. [Google Scholar] [CrossRef]
- Deng, J.V.; Wan, Y.; Wang, X.; Cohen, S.; Wetsel, W.C.; Greenberg, M.E.; Kenny, P.J.; Calakos, N.; West, A.E. MeCP2 Phosphorylation Limits Psychostimulant-Induced Behavioral and Neuronal Plasticity. J. Neurosci. 2014, 34, 4519–4527. [Google Scholar] [CrossRef] [Green Version]
- Molgaard, S.; Ulrichsen, M.; Boggild, S.; Holm, M.-L.; Vaegter, C.; Nyengaard, J.; Glerup, S. Immunofluorescent Visualization of Mouse Interneuron Subtypes. F1000Res 2014, 3, 242. [Google Scholar] [CrossRef] [Green Version]
- Równiak, M.; Kolenkiewicz, M.; Kozłowska, A. Parvalbumin, but Not Calretinin, Neurons Express High Levels of A1-Containing GABAA Receptors, A7-Containing Nicotinic Acetylcholine Receptors and D2-Dopamine Receptors in the Basolateral Amygdala of the Rat. J. Chem. Neuroanat. 2017, 86, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Ren, S.; Gao, P.; Wan, D.; Rong, S.; Li, X.; Liu, S.; Xu, S.; Sun, K.; Guo, B.; et al. ALG13 Participates in Epileptogenesis via Regulation of GABAA Receptors in Mouse Models. Cell Death Discov. 2020, 6, 87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Jiao, Y.-Y.; Sun, Q.-Q. Developmental Maturation of Excitation and Inhibition Balance in Principal Neurons across Four Layers of Somatosensory Cortex. Neuroscience 2011, 174, 10–25. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates; Gulf Professional Publishing: Oxford, UK, 2004; ISBN 978-0-12-547640-9. [Google Scholar]
- Sathyanesan, A.; Ogura, T.; Lin, W. Automated Measurement of Nerve Fiber Density Using Line Intensity Scan Analysis. J. Neurosci. Methods 2012, 206, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijering, E. FeatureJ: A Java Package for Image Feature Extraction. Available online: https://imagescience.org/meijering/software/featurej/ (accessed on 17 March 2021).
- McDonald, A.J.; Mascagni, F. Parvalbumin-Containing Interneurons in the Basolateral Amygdala Express High Levels of the ?1 Subunit of the GABAA Receptor. J. Comp. Neurol. 2004, 473, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, W.A.; Humpel, C.; Alheid, G.F.; Marksteiner, J. Compartmentation of Alpha 1 and Alpha 2 GABAA Receptor Subunits within Rat Extended Amygdala: Implications for Benzodiazepine Action. Brain Res. 2003, 964, 91–99. [Google Scholar] [CrossRef]
- Hörtnagl, H.; Tasan, R.O.; Wieselthaler, A.; Kirchmair, E.; Sieghart, W.; Sperk, G. Patterns of MRNA and Protein Expression for 12 GABAA Receptor Subunits in the Mouse Brain. Neuroscience 2013, 236, 345–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyles, D.W.; McGrath, J.J.; Reynolds, G.P. Neuronal Calcium-Binding Proteins and Schizophrenia. Schizophr. Res. 2002, 57, 27–34. [Google Scholar] [CrossRef]
- Köhr, G.; Lambert, C.E.; Mody, I. Calbindin-D28K (CaBP) Levels and Calcium Currents in Acutely Dissociated Epileptic Neurons. Exp. Brain Res. 1991, 85, 543–551. [Google Scholar] [CrossRef]
- Chard, P.S.; Jordan, J.; Marcuccilli, C.J.; Miller, R.J.; Leiden, J.M.; Roos, R.P.; Ghadge, G.D. Regulation of Excitatory Transmission at Hippocampal Synapses by Calbindin D28k. Proc. Natl. Acad. Sci. USA 1995, 92, 5144–5148. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.P.; Abel, J.M.; Tejada, L.D.; Rissman, E.F. Calbindin Knockout Alters Sex-Specific Regulation of Behavior and Gene Expression in Amygdala and Prefrontal Cortex. Endocrinology 2016, 157, 1967–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benes, F.M.; Kwok, E.W.; Vincent, S.L.; Todtenkopf, M.S. A Reduction of Nonpyramidal Cells in Sector CA2 of Schizophrenics and Manic Depressives. Biol. Psychiatry 1998, 44, 88–97. [Google Scholar] [CrossRef]
- Kaalund, S.S.; Riise, J.; Broberg, B.V.; Fabricius, K.; Karlsen, A.S.; Secher, T.; Plath, N.; Pakkenberg, B. Differential Expression of Parvalbumin in Neonatal Phencyclidine-Treated Rats and Socially Isolated Rats. J. Neurochem. 2013, 124, 548–557. [Google Scholar] [CrossRef]
- Schwaller, B. The Continuing Disappearance of “Pure” Ca2+ Buffers. Cell. Mol. Life Sci. 2009, 66, 275–300. [Google Scholar] [CrossRef] [PubMed]
- Kreiner, L.; Christel, C.J.; Benveniste, M.; Schwaller, B.; Lee, A. Compensatory Regulation of Cav2.1 Ca2+ Channels in Cerebellar Purkinje Neurons Lacking Parvalbumin and Calbindin D-28k. J. Neurophysiol 2010, 103, 371–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwaller, B. Emerging Functions of the “Ca2+ Buffers” Parvalbumin, Calbindin D-28k and Calretinin in the Brain. In Handbook of Neurochemistry and Molecular Neurobiology: Neural Protein Metabolism and Function; Lajtha, A., Banik, N., Eds.; Springer: Boston, MA, USA, 2007; pp. 197–221. ISBN 978-0-387-30379-6. [Google Scholar]
- Rozov, A.; Burnashev, N.; Sakmann, B.; Neher, E. Transmitter Release Modulation by Intracellular Ca2+ Buffers in Facilitating and Depressing Nerve Terminals of Pyramidal Cells in Layer 2/3 of the Rat Neocortex Indicates a Target Cell-Specific Difference in Presynaptic Calcium Dynamics. J. Physiol. 2001, 531, 807–826. [Google Scholar] [CrossRef] [PubMed]
- Blatow, M.; Caputi, A.; Burnashev, N.; Monyer, H.; Rozov, A. Ca2+ Buffer Saturation Underlies Paired Pulse Facilitation in Calbindin-D28k-Containing Terminals. Neuron 2003, 38, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Tasan, R.O.; Bukovac, A.; Peterschmitt, Y.N.; Sartori, S.B.; Landgraf, R.; Singewald, N.; Sperk, G. Altered GABA Transmission in a Mouse Model of Increased Trait Anxiety. Neuroscience 2011, 183, 71–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zink, M.; Vollmayr, B.; Gebicke-Haerter, P.J.; Henn, F.A. Reduced Expression of GABA Transporter GAT3 in Helpless Rats, an Animal Model of Depression. Neurochem. Res. 2009, 34, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-L.; Sun, Y.-X.; Liu, X.; Wang, H.; Ma, Y.-N.; Su, Y.-A.; Li, J.-T.; Si, T.-M. Adolescent Stress Increases Depression-like Behaviors and Alters the Excitatory-Inhibitory Balance in Aged Mice. Chin. Med. J. 2019, 132, 1689–1699. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Xu, A.; Cui, S.; Sun, M.-R.; Xue, Y.-C.; Wang, J.-H. Impaired GABA Synthesis, Uptake and Release Are Associated with Depression-like Behaviors Induced by Chronic Mild Stress. Transl. Psychiatry 2016, 6, e910. [Google Scholar] [CrossRef] [PubMed]
- Tye, K.M.; Prakash, R.; Kim, S.-Y.; Fenno, L.E.; Grosenick, L.; Zarabi, H.; Thompson, K.R.; Gradinaru, V.; Ramakrishnan, C.; Deisseroth, K. Amygdala Circuitry Mediating Reversible and Bidirectional Control of Anxiety. Nature 2011, 471, 358–362. [Google Scholar] [CrossRef]
- Zhang, W.; Guo, Y.; Wang, K.; Chen, L.; Jiang, P. Neuroprotective Effects of Vitamin D and 17ß-Estradiol against Ovariectomy-Induced Neuroinflammation and Depressive-like State: Role of the AMPK/NF-ΚB Pathway. Int. Immunopharmacol. 2020, 86, 106734. [Google Scholar] [CrossRef]
- Gadea, A.; López-Colomé, A.M. Glial Transporters for Glutamate, Glycine, and GABA: II. GABA Transporters. J. Neurosci. Res. 2001, 63, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Chazalon, M.; Paredes-Rodriguez, E.; Morin, S.; Martinez, A.; Cristóvão-Ferreira, S.; Vaz, S.; Sebastiao, A.; Panatier, A.; Boué-Grabot, E.; Miguelez, C.; et al. GAT-3 Dysfunction Generates Tonic Inhibition in External Globus Pallidus Neurons in Parkinsonian Rodents. Cell Rep. 2018, 23, 1678–1690. [Google Scholar] [CrossRef] [Green Version]
- Patrone, C.; Andersson, S.; Korhonen, L.; Lindholm, D. Estrogen Receptor-Dependent Regulation of Sensory Neuron Survival in Developing Dorsal Root Ganglion. Proc. Natl. Acad. Sci. USA 1999, 96, 10905–10910. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Andersson, S.; Warner, M.; Gustafsson, J.-A. Estrogen Receptor (ER) Knockout Mice Reveal a Role for ER in Migration of Cortical Neurons in the Developing Brain. Proc. Natl. Acad. Sci. 2003, 100, 703–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, K.G.; Milic, J.; Zaciragic, A.; Wen, K.; Jaspers, L.; Nano, J.; Dhana, K.; Bramer, W.M.; Kraja, B.; van Beeck, E.; et al. The Functions of Estrogen Receptor Beta in the Female Brain: A Systematic Review. Maturitas 2016, 93, 41–57. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.K.; Christakos, S. Regulation by Estrogen through the 5’-Flanking Region of the Mouse Calbindin-D28k Gene. Mol. Endocrinol. 1995, 9, 319–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shortall, S.E.; Brown, A.M.; Newton-Mann, E.; Dawe-Lane, E.; Evans, C.; Fowler, M.; King, M.V. Calbindin Deficits May Underlie Dissociable Effects of 5-HT6 and MGlu7 Antagonists on Glutamate and Cognition in a Dual-Hit Neurodevelopmental Model for Schizophrenia. Mol. Neurobiol. 2020, 57, 3439–3457. [Google Scholar] [CrossRef] [PubMed]
- Helmeke, C.; Ovtscharoff, W.; Poeggel, G.; Braun, K. Imbalance of Immunohistochemically Characterized Interneuron Populations in the Adolescent and Adult Rodent Medial Prefrontal Cortex after Repeated Exposure to Neonatal Separation Stress. Neuroscience 2008, 152, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Giachino, C.; Canalia, N.; Capone, F.; Fasolo, A.; Alleva, E.; Riva, M.A.; Cirulli, F.; Peretto, P. Maternal Deprivation and Early Handling Affect Density of Calcium Binding Protein-Containing Neurons in Selected Brain Regions and Emotional Behavior in Periadolescent Rats. Neuroscience 2007, 145, 568–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iritani, S.; Kuroki, N.; Ikeda, K.; Kazamatsuri, H. Calbindin Immunoreactivity in the Hippocampal Formation and Neocortex of Schizophrenics. Prog. Neuropsychopharmacol. Biol. Psychiatry 1999, 23, 409–421. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Code | Clonality | Host Species | Dilution | Supplier | Location |
|---|---|---|---|---|---|---|
| Primary antibodies | ||||||
| NeuN | ABN78 | polyclonal | Rabbit | 1:1000 | Millipore | Temecula, CA, USA |
| CB | CB-38 | polyclonal | Rabbit | 1:4000 | SWANT | Bellinzona, Switzerland |
| PV | PV27 | polyclonal | Rabbit | 1:1000 | SWANT | Bellinzona, Switzerland |
| CR | 7697 | polyclonal | Rabbit | 1:1000 | SWANT | Bellinzona, Switzerland |
| SOM | MAB354 | monoclonal | Rat | 1:1000 | Millipore | Temecula, CA, USA |
| GABAA α1 | AB33299 | polyclonal | Rabbit | 1:2000 | Abcam | Cambridge, UK |
| VGAT | AB5062P | polyclonal | Rabbit | 1:3000 | Millipore | Temecula, CA, USA |
| GFAP | G9269 | polyclonal | Rabbit | 1:200 | Millipore | Temecula, CA, USA |
| Secondary antibodies | ||||||
| ImmPRESS HRP Universal Antibody (anti-rabbit Ig, Peroxidase) | 1:1 | Vector Laboratories | Burlingame, CA, USA | |||
| ALEXA Fluor 555 | A-31572 | polyclonal | Donkey anti-rabbit | 1:1000 | Thermo Fisher | Rockford, IL, USA |
| CY3 | 712-165-153 | polyclonal | Donkey anti-rat | 1:1000 | Jackson ImmunoResearch Laboratories | West Grove, PA, USA |
| Other reagents | ||||||
| 3,3-diaminobenzidine substrate chromogen | 3% | Dako Cytomation | Glostrup, Denmark | |||
| LA | BL | BM | ME | CE | CO | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p | p | p | p | p | p | |||||||
| NeuN | t8.88 = 4.77 | 0.001057 | t9.70 = 6.88 | 0.000051 | t9.02 = 7.25 | 0.000048 | t8.98 = 6.74 | 0.000085 | t9.78 = 7.05 | 0.000039 | t6.66 = 4.86 | 0.002112 |
| CB | t7.27 = 4.74 | 0.001913 | t7.27 = 4.74 | 0.002045 | t9.55 = 4.16 | 0.004259 | t9.58 = 6.33 | 0.001030 | t9.99 = 4.46 | 0.001229 | t7.80 = 4.40 | 0.002435 |
| PV | t10 = 0.054 | 0.9581 | t10 = 0.62 | 0.5482 | t10 = 0.48 | 0.6642 | − | − | − | − | t10 = 0.46 | 0.6581 |
| CR | t10 = 0.53 | 0.6113 | t10 = 0.16 | 0.8787 | t10 = −0.24 | 0.8119 | t10 = 1.24 | 0.2426 | − | − | t10 = 2.13 | 0.0594 |
| SOM | t10 = −0.22 | 0.8274 | t10 = 0.08 | 0.9344 | t10 = −0.26 | 0.8036 | t10 = 0.17 | 0.8662 | t10 = 0.63 | 0.5441 | t10 = 0.16 | 0.8732 |
| GABAAα1 | t10 = 0.35 | 0.7337 | t10 = 0.11 | 0.9178 | t10 = −0.62 | 0.5478 | t10 = 0.08 | 0.9357 | t10 = −0.19 | 0.8513 | t10 = 0.11 | 0.9106 |
| Intensity of GABAAα1 | t10 = 0.94 | 0.3674 | t10 = 0.97 | 0.3536 | t10 = 1.24 | 0.2445 | t10 = −0.82 | 0.4334 | t10 = −0.85 | 0.4133 | t10 = −0.08 | 0.9379 |
| VGAT | t10 = −0.22 | 0.8274 | t10 = 0.08 | 0.9344 | t10 = −0.26 | 0.8036 | t10 = 0.17 | 0.8662 | t10 = 0.63 | p = 0.5441 | t10 = 0.16 | 0.8732 |
| Intensity of GABAAα1 | t10 = −0.57 | 0.5829 | t10 = −1.00 | 0.3432 | t10 = −0.29 | 0.7796 | t10 = −0.31 | 0.7607 | t10 = 0.19 | p = 0.8495 | t10 = 1.07 | 0.3091 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalinowski, D.; Bogus-Nowakowska, K.; Kozłowska, A.; Równiak, M. Expression of Calbindin, a Marker of Gamma-Aminobutyric Acid Neurons, Is Reduced in the Amygdala of Oestrogen Receptor β-Deficient Female Mice. J. Clin. Med. 2022, 11, 1760. https://doi.org/10.3390/jcm11071760
Kalinowski D, Bogus-Nowakowska K, Kozłowska A, Równiak M. Expression of Calbindin, a Marker of Gamma-Aminobutyric Acid Neurons, Is Reduced in the Amygdala of Oestrogen Receptor β-Deficient Female Mice. Journal of Clinical Medicine. 2022; 11(7):1760. https://doi.org/10.3390/jcm11071760
Chicago/Turabian StyleKalinowski, Daniel, Krystyna Bogus-Nowakowska, Anna Kozłowska, and Maciej Równiak. 2022. "Expression of Calbindin, a Marker of Gamma-Aminobutyric Acid Neurons, Is Reduced in the Amygdala of Oestrogen Receptor β-Deficient Female Mice" Journal of Clinical Medicine 11, no. 7: 1760. https://doi.org/10.3390/jcm11071760