

Membrane Order Effect on the Photoresponse of an Organic Transducer
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
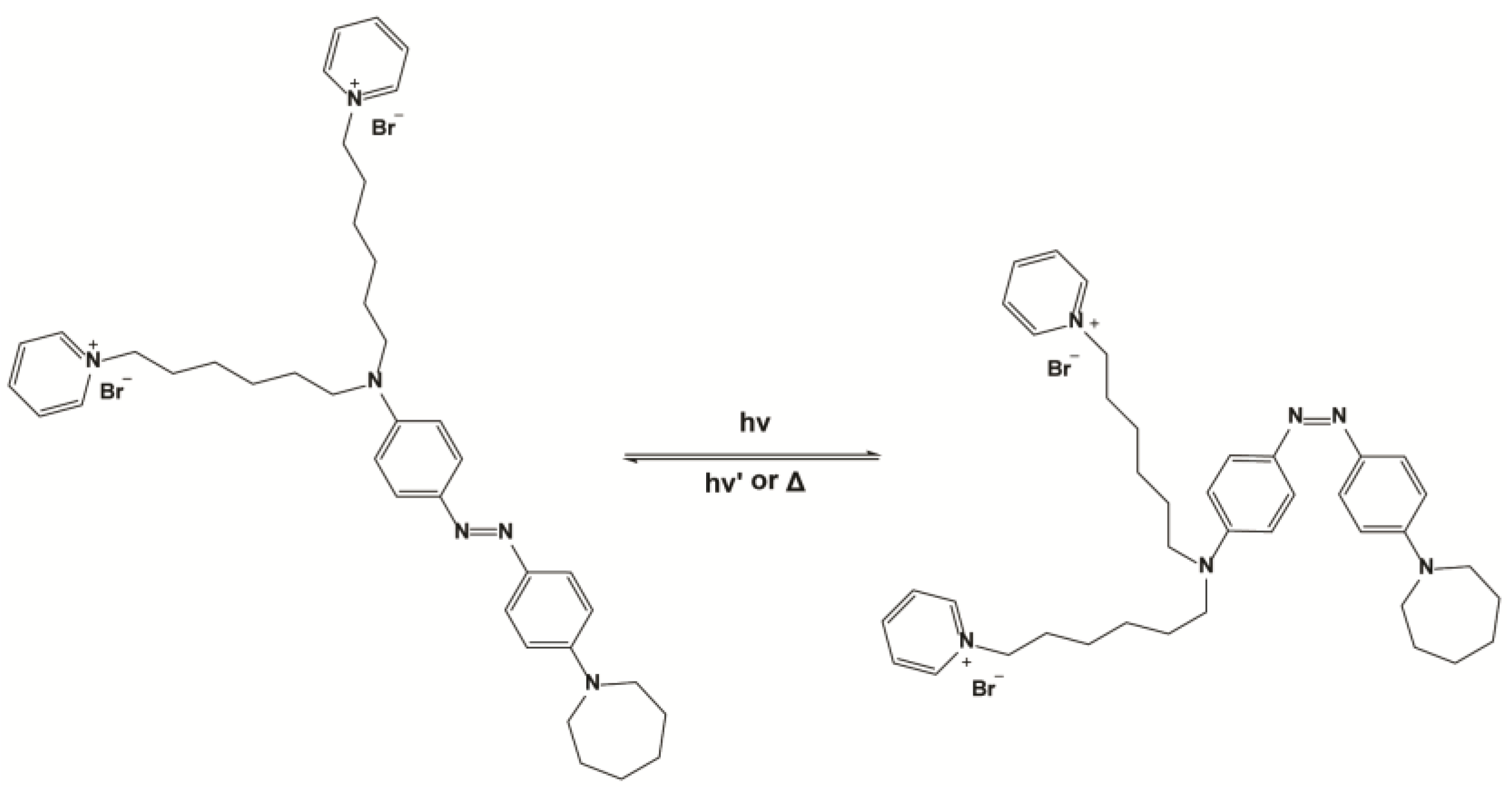
2. Materials and Methods
3. Results
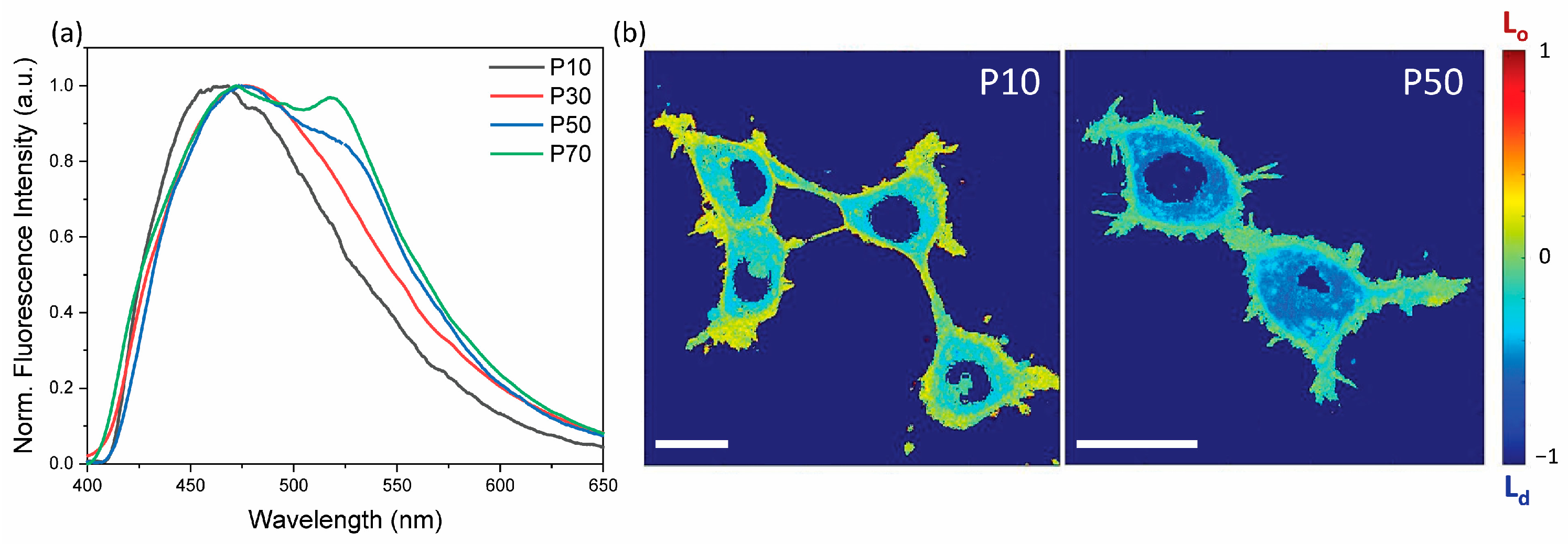
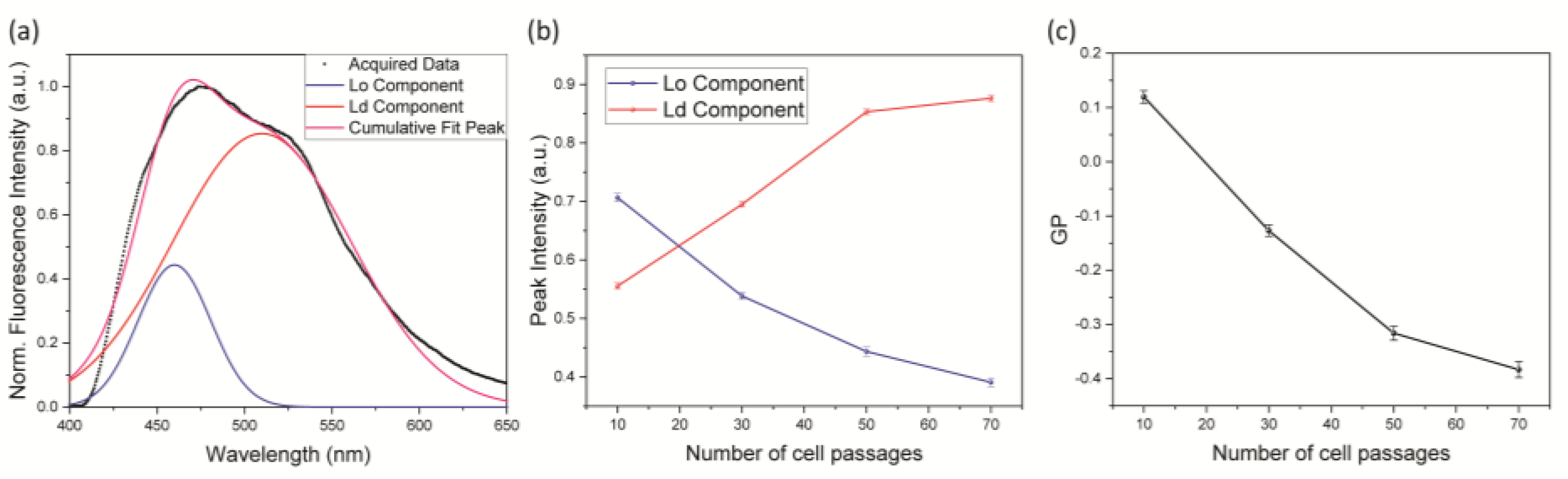
3.1. Membrane Disordering
3.2. Thermal Effect
3.3. Relationship between Photoactuation and Cell Passage
3.4. Photoactuation in Liposomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghezzi, D.; Antognazza, M.R.; Dal Maschio, M.; Lanzarini, E.; Benfenati, F.; Lanzani, G. A hybrid bioorganic interface for neuronal photoactivation. Nat. Commun. 2011, 2, 166. [Google Scholar] [CrossRef] [PubMed]
- Fenno, L.; Yizhar, O.; Deisseroth, K. The development and application of optogenetics. Annu. Rev. Neurosci. 2011, 34, 389–412. [Google Scholar] [CrossRef] [PubMed]
- Deisseroth, K. Optogenetics: 10 years of microbial opsins in neuroscience. Nat. Neurosci. 2015, 18, 1213–1225. [Google Scholar] [CrossRef]
- Antognazza, M.R.; Di Paolo, M.; Ghezzi, D.; Mete, M.; Di Marco, S.; Maya-Vetencourt, J.F.; Maccarone, R.; Desii, A.; Di Fonzo, F.; Bramini, M.; et al. Characterization of a Polymer-Based, Fully Organic Prosthesis for Implantation into the Subretinal Space of the Rat. Adv. Healthc. Mater. 2016, 5, 2271–2282. [Google Scholar] [CrossRef] [PubMed]
- Maya-Vetencourt, J.F.; Ghezzi, D.; Antognazza, M.R.; Colombo, E.; Mete, M.; Feyen, P.; Desii, A.; Buschiazzo, A.; Di Paolo, M.; Di Marco, S.; et al. A fully organic retinal prosthesis restores vision in a rat model of degenerative blindness. Nat. Mater. 2017, 16, 681–689. [Google Scholar] [CrossRef]
- Maya-Vetencourt, J.F.; Manfredi, G.; Mete, M.; Colombo, E.; Bramini, M.; Di Marco, S.; Shmal, D.; Mantero, G.; Dipalo, M.; Rocchi, A.; et al. Subretinally injected semiconducting polymer nanoparticles rescue vision in a rat model of retinal dystrophy. Nat. Nanotechnol. 2020, 15, 698–708. [Google Scholar] [CrossRef]
- Francia, S.; Shmal, D.; Di Marco, S.; Chiaravalli, G.; Maya-Vetencourt, J.F.; Mantero, G.; Michetti, C.; Cupini, S.; Manfredi, G.; DiFrancesco, M.L.; et al. Light-induced charge generation in polymeric nanoparticles restores vision in advanced-stage retinitis pigmentosa rats. Nat. Commun. 2022, 13, 3677. [Google Scholar] [CrossRef]
- Martino, N.; Ghezzi, D.; Benfenati, F.; Lanzani, G.; Antognazza, M.R. Organic semiconductors for artificial vision. J. Mater. Chem. B 2013, 1, 3768–3780. [Google Scholar] [CrossRef]
- Lanzani, G. Organic electronics meets biology. Nat. Mater. 2014, 13, 775–776. [Google Scholar] [CrossRef]
- Hu, W.; Li, Q.; Li, B.; Ma, K.; Zhang, C.; Fu, X. Optogenetics sheds new light on tissue engineering and regenerative medicine. Biomaterials 2020, 227, 119546. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; Tian, H. Taking orders from light: Progress in photochromic bio-materials. Mater. Horiz. 2014, 1, 169–184. [Google Scholar] [CrossRef]
- DiFrancesco, M.L.; Lodola, F.; Colombo, E.; Maragliano, L.; Bramini, M.; Paternò, G.M.; Baldelli, P.; Serra, M.D.; Lunelli, L.; Marchioretto, M.; et al. Neuronal firing modulation by a membrane-targeted photoswitch. Nat. Nanotechnol. 2020, 15, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Paternò, G.M.; Colombo, E.; Vurro, V.; Lodola, F.; Cimò, S.; Sesti, V.; Molotokaite, E.; Bramini, M.; Ganzer, L.; Fazzi, D.; et al. Membrane Environment Enables Ultrafast Isomerization of Amphiphilic Azobenzene. Adv. Sci. 2020, 7, 1903241. [Google Scholar] [CrossRef] [PubMed]
- Vurro, V.; Bondelli, G.; Sesti, V.; Lodola, F.; Paternò, G.M.; Lanzani, G.; Bertarelli, C. Molecular Design of Amphiphilic Plasma Membrane-Targeted Azobenzenes for Nongenetic Optical Stimulation. Front. Mater. 2021, 7, 472. [Google Scholar] [CrossRef]
- Paterno, G.M.; Lanzani, G.; Bondelli, G.; Sakai, V.G.; Sesti, V.; Bertarelli, C. The effect of an intramembrane light-actuator on the dynamics of phospholipids in model membranes and intact cells. Langmuir 2020, 36, 11517–11527. [Google Scholar] [CrossRef] [PubMed]
- Magni, A.; Bondelli, G.; Paternò, G.M.; Sardar, S.; Sesti, V.; D’Andrea, C.; Bertarelli, C.; Lanzani, G. Azobenzene photoisomerization probes cell membrane viscosity. Phys. Chem. Chem. Phys. 2022, 24, 8716–8723. [Google Scholar] [CrossRef]
- de Souza-Guerreiro, T.C.; Bondelli, G.; Grobas, I.; Donini, S.; Sesti, V.; Bertarelli, C.; Lanzani, G.; Asally, M.; Paternò, G.M. Membrane Targeted Azobenzene Drives Optical Modulation of Bacterial Membrane Potential. Adv. Sci. 2023, 10, 2205007. [Google Scholar] [CrossRef]
- Vurro, V.; Federici, B.; Ronchi, C.; Florindi, C.; Sesti, V.; Crasto, S.; Maniezzi, C.; Galli, C.; Antognazza, M.R.; Bertarelli, C.; et al. Optical modulation of excitation-contraction coupling in human-induced pluripotent stem cell-derived cardiomyocytes. iScience 2023, 26, 106121. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, G.; Lodola, F.; Paternó, G.M.; Vurro, V.; Baldelli, P.; Benfenati, F.; Lanzani, G. The physics of plasma membrane photostimulation. APL Mater. 2021, 9, 30901. [Google Scholar] [CrossRef]
- Bondelli, G.; Paternò, G.M.; Lanzani, G. Fluorescent probes for optical investigation of the plasma membrane. Opt. Mater. X 2021, 12, 100085. [Google Scholar] [CrossRef]
- Klymchenko, A.S. Fluorescent Probes for Lipid Membranes: From the Cell Surface to Organelles. Acc. Chem. Res. 2023, 56, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Garg, R.; Cohen-Karni, D.; Cohen-Karni, T. Neural modulation with photothermally active nanomaterials. Nat. Rev. Bioeng. 2023, 1, 193–207. [Google Scholar] [CrossRef]
- Kwist, K.; Bridges, W.C.; Burg, K.J.L. The effect of cell passage number on osteogenic and adipogenic characteristics of D1 cells. Cytotechnology 2016, 68, 1661–1667. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi Farsani, T.; Motevaseli, E.; Neyazi, N.; Khorramizadeh, M.R.; Zafarvahedian, E.; Ghahremani, M.H. Effect of passage number and culture time on the expression and activity of insulin-degrading enzyme in caco-2 cells. Iran. Biomed. J. 2018, 22, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, C.; Schildknecht, S. In vitro research reproducibility: Keeping up high standards. Front. Pharmacol. 2019, 10, 1484. [Google Scholar] [CrossRef]
- Cao, J.; Wu, X.; Qin, X.; Li, Z. Uncovering the Effect of Passage Number on HT29 Cell Line Based on the Cell Metabolomic Approach. J. Proteome Res. 2021, 20, 1582–1590. [Google Scholar] [CrossRef]
- Cantor, E.L.; Shen, F.; Jiang, G.; Tan, Z.; Cunningham, G.M.; Wu, X.; Philips, S.; Schneider, B.P. Passage number affects differentiation of sensory neurons from human induced pluripotent stem cells. Sci. Rep. 2022, 12, 15869. [Google Scholar] [CrossRef]
- Escribá, P.V. Membrane-lipid therapy: A new approach in molecular medicine. Trends Mol. Med. 2006, 12, 34–43. [Google Scholar] [CrossRef]
- Drolle, E.; Negoda, A.; Hammond, K.; Pavlov, E.; Leonenko, Z. Changes in lipid membranes may trigger amyloid toxicity in Alzheimer’s disease. PLoS ONE 2017, 12, e0182194. [Google Scholar] [CrossRef]
- Hardy, J. Membrane damage is at the core of Alzheimer’s disease. Lancet Neurol. 2017, 16, 342. [Google Scholar] [CrossRef]
- Tan, L.T.H.; Chan, K.G.; Pusparajah, P.; Lee, W.L.; Chuah, L.H.; Khan, T.M.; Lee, L.H.; Goh, B.H. Targeting membrane lipid a potential cancer cure? Front. Pharmacol. 2017, 8, 12. [Google Scholar] [CrossRef]
- Lakowicz, J.R. (Ed.) Instrumentation for Fluorescence Spectroscopy. In Principles of Fluorescence Spectroscopy; Springer: Boston, MA, USA, 2006; pp. 27–61. ISBN 978-0-387-46312-4. [Google Scholar]
- Parasassi, T.; Gratton, E. Membrane lipid domains and dynamics as detected by Laurdan fluorescence. J. Fluoresc. 1995, 5, 59–69. [Google Scholar] [CrossRef]
- Malacrida, L.; Jameson, D.M.; Gratton, E. A multidimensional phasor approach reveals LAURDAN photophysics in NIH-3T3 cell membranes. Sci. Rep. 2017, 7, 9215. [Google Scholar] [CrossRef] [PubMed]
- Bondelli, G.; Sardar, S.; Chiaravalli, G.; Vurro, V.; Paternò, G.M.; Lanzani, G.; D’Andrea, C. Shedding Light on Thermally Induced Optocapacitance at the Organic Biointerface. J. Phys. Chem. B 2021, 125, 10748–10758. [Google Scholar] [CrossRef]
- Golfetto, O.; Hinde, E.; Gratton, E. The laurdan spectral phasor method to explore membrane micro-heterogeneity and lipid domains in live cells. Methods Membr. Lipids Second Ed. 2014, 1232, 273–290. [Google Scholar] [CrossRef]
- Sankaran, J.; Wohland, T. Fluorescence strategies for mapping cell membrane dynamics and structures. APL Bioeng. 2020, 4, 20901. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.A.; Stinson, B.M.; Go, M.S.; Carmona, L.M.; Reminick, J.I.; Fang, X.; Baumgart, T. Temperature-dependent phase behavior and protein partitioning in giant plasma membrane vesicles. Biochim. Biophys. Acta Biomembr. 2010, 1798, 1427–1435. [Google Scholar] [CrossRef]
- Crocke, E.L. Cholesterol function in plasma membranes from ectotherms: Membrane-specific roles in adaptation to temperature. Am. Zool. 1998, 38, 291–304. [Google Scholar] [CrossRef]
- Simons, K.; Vaz, W.L.C. Model systems, lipid rafts, and cell membranes. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 269–295. [Google Scholar] [CrossRef]
- De Almeida, R.F.M.; Fedorov, A.; Prieto, M. Sphingomyelin/phosphatidylcholine/cholesterol phase diagram: Boundaries and composition of lipid rafts. Biophys. J. 2003, 85, 2406–2416. [Google Scholar] [CrossRef]
- Ovsepyan, L.M.; Kazaryan, G.S.; Akopdzhanyan, A.A.; Lvov, M.V. Age-dependent changes in phospholipid content and neutral lipid contents during aging. Adv. Gerontol. 2013, 3, 42–45. [Google Scholar] [CrossRef]
- Taciak, B.; Białasek, M.; Braniewska, A.; Sas, Z.; Sawicka, P.; Kiraga, Ł.; Rygiel, T.; Król, M. Evaluation of phenotypic and functional stability of RAW 264.7 cell line through serial passages. PLoS ONE 2018, 13, e0198943. [Google Scholar] [CrossRef]
- Capes-Davis, A.; Bairoch, A.; Barrett, T.; Burnett, E.C.; Dirks, W.G.; Hall, E.M.; Healy, L.; Kniss, D.A.; Korch, C.; Liu, Y.; et al. Cell Lines as Biological Models: Practical Steps for More Reliable Research. Chem. Res. Toxicol. 2019, 32, 1733–1736. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Free radical oxidation of proteins and its relationship with functional state of organisms. Biochemistry 2007, 72, 809–827. [Google Scholar] [CrossRef] [PubMed]
- Galatro, A.; González, P.M.; Malanga, G.; Robello, E.; Piloni, N.E.; Puntarulo, S. Nitric oxide and membrane lipid peroxidation in photosynthetic and non-photosynthetic organisms under several stress conditions. Front. Physiol. 2013, 4, 276. [Google Scholar] [CrossRef] [PubMed]
- Van Der Paal, J.; Neyts, E.C.; Verlackt, C.C.W.; Bogaerts, A. Effect of lipid peroxidation on membrane permeability of cancer and normal cells subjected to oxidative stress. Chem. Sci. 2016, 7, 489–498. [Google Scholar] [CrossRef]
- Ke, Y.; Karki, P.; Kim, J.; Son, S.; Berdyshev, E.; Bochkov, V.N.; Birukova, A.A.; Birukov, K.G. Elevated truncated oxidized phospholipids as a factor exacerbating ALI in the aging lungs. FASEB J. 2019, 33, 3887–3900. [Google Scholar] [CrossRef]
- Catalá, A.; Díaz, M. Editorial: Impact of lipid peroxidation on the physiology and pathophysiology of cell membranes. Front. Physiol. 2016, 7, 423. [Google Scholar] [CrossRef]
- Yang, H.; Zhou, M.; Li, H.; Wei, T.; Tang, C.; Zhou, Y.; Long, X. Effects of Low-level Lipid Peroxidation on the Permeability of Nitroaromatic Molecules across a Membrane: A Computational Study. ACS Omega 2020, 5, 4798–4806. [Google Scholar] [CrossRef]
- Iuchi, K.; Takai, T.; Hisatomi, H. Cell death via lipid peroxidation and protein aggregation diseases. Biology 2021, 10, 399. [Google Scholar] [CrossRef]
- Nishizawa, H.; Matsumoto, M.; Chen, G.; Ishii, Y.; Tada, K.; Onodera, M.; Kato, H.; Muto, A.; Tanaka, K.; Igarashi, K. Lipid peroxidation and the subsequent cell death transmitting from ferroptotic cells to neighboring cells. Cell Death Dis. 2021, 12, 332. [Google Scholar] [CrossRef] [PubMed]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vurro, V.; Moschetta, M.; Bondelli, G.; Sardar, S.; Magni, A.; Sesti, V.; Paternò, G.M.; Bertarelli, C.; D’Andrea, C.; Lanzani, G. Membrane Order Effect on the Photoresponse of an Organic Transducer. Membranes 2023, 13, 538. https://doi.org/10.3390/membranes13050538
Vurro V, Moschetta M, Bondelli G, Sardar S, Magni A, Sesti V, Paternò GM, Bertarelli C, D’Andrea C, Lanzani G. Membrane Order Effect on the Photoresponse of an Organic Transducer. Membranes. 2023; 13(5):538. https://doi.org/10.3390/membranes13050538
Chicago/Turabian StyleVurro, Vito, Matteo Moschetta, Gaia Bondelli, Samim Sardar, Arianna Magni, Valentina Sesti, Giuseppe Maria Paternò, Chiara Bertarelli, Cosimo D’Andrea, and Guglielmo Lanzani. 2023. "Membrane Order Effect on the Photoresponse of an Organic Transducer" Membranes 13, no. 5: 538. https://doi.org/10.3390/membranes13050538