

Methylene Blue Inhibits Cromakalim-Activated K+ Currents in Follicle-Enclosed Oocytes


 , and
, and 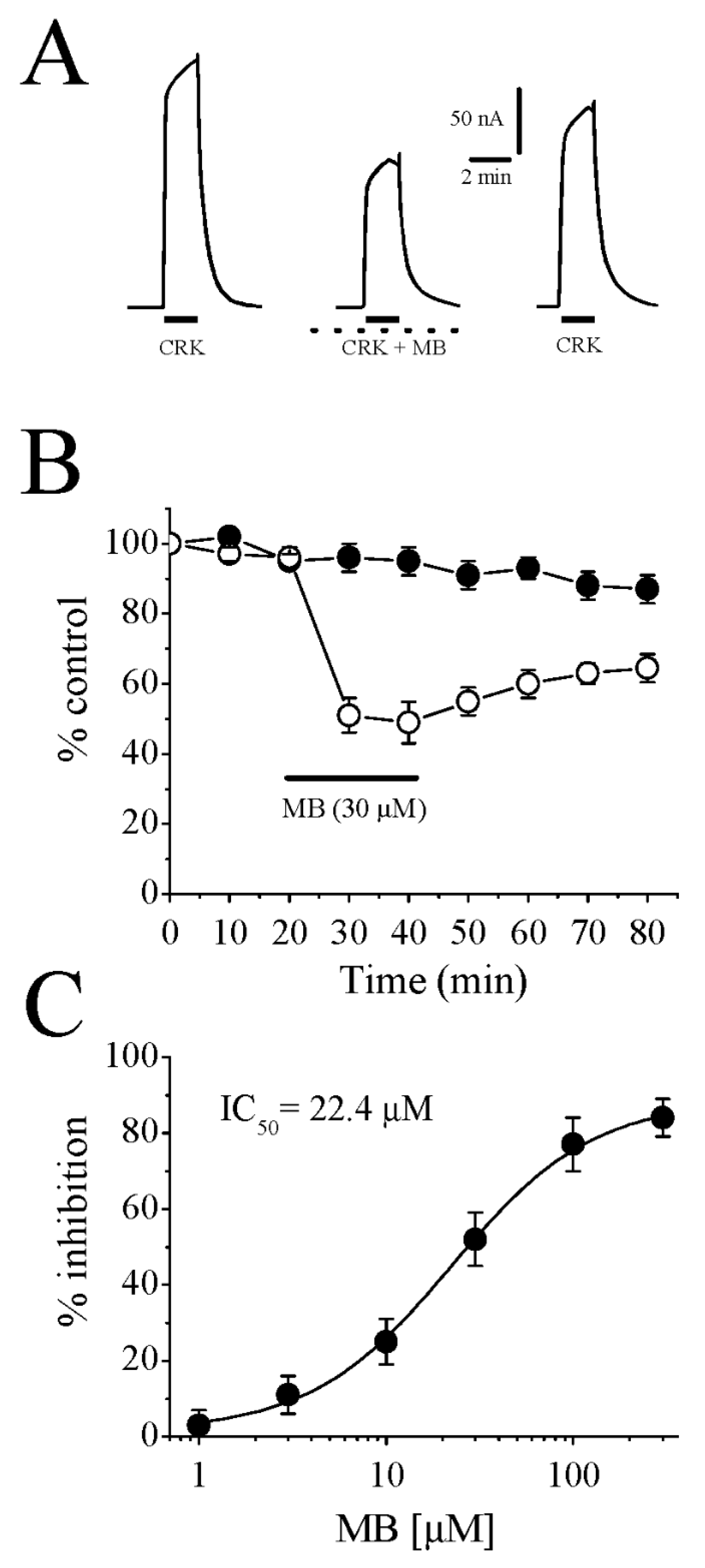{kind=link}
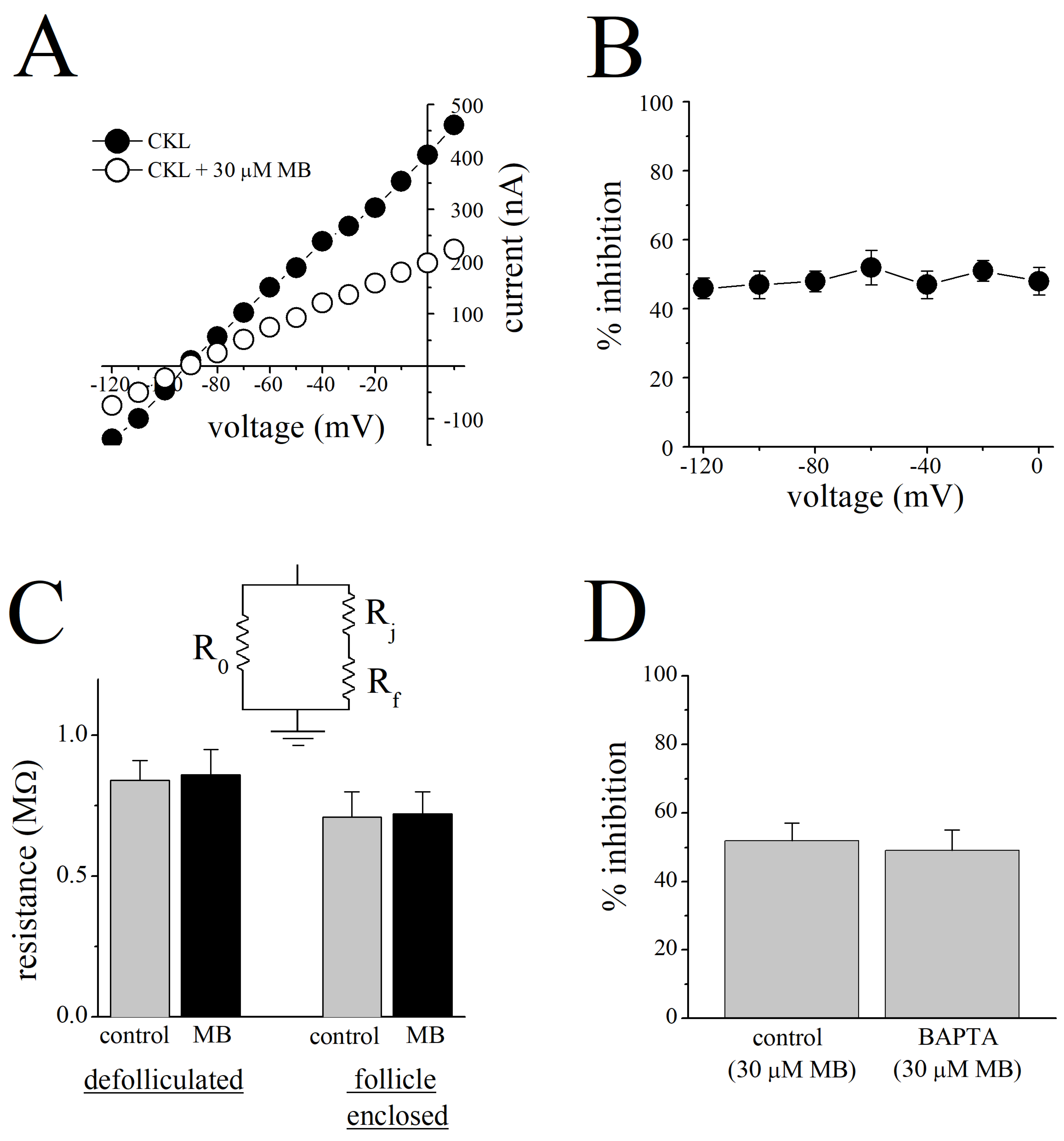{kind=link}
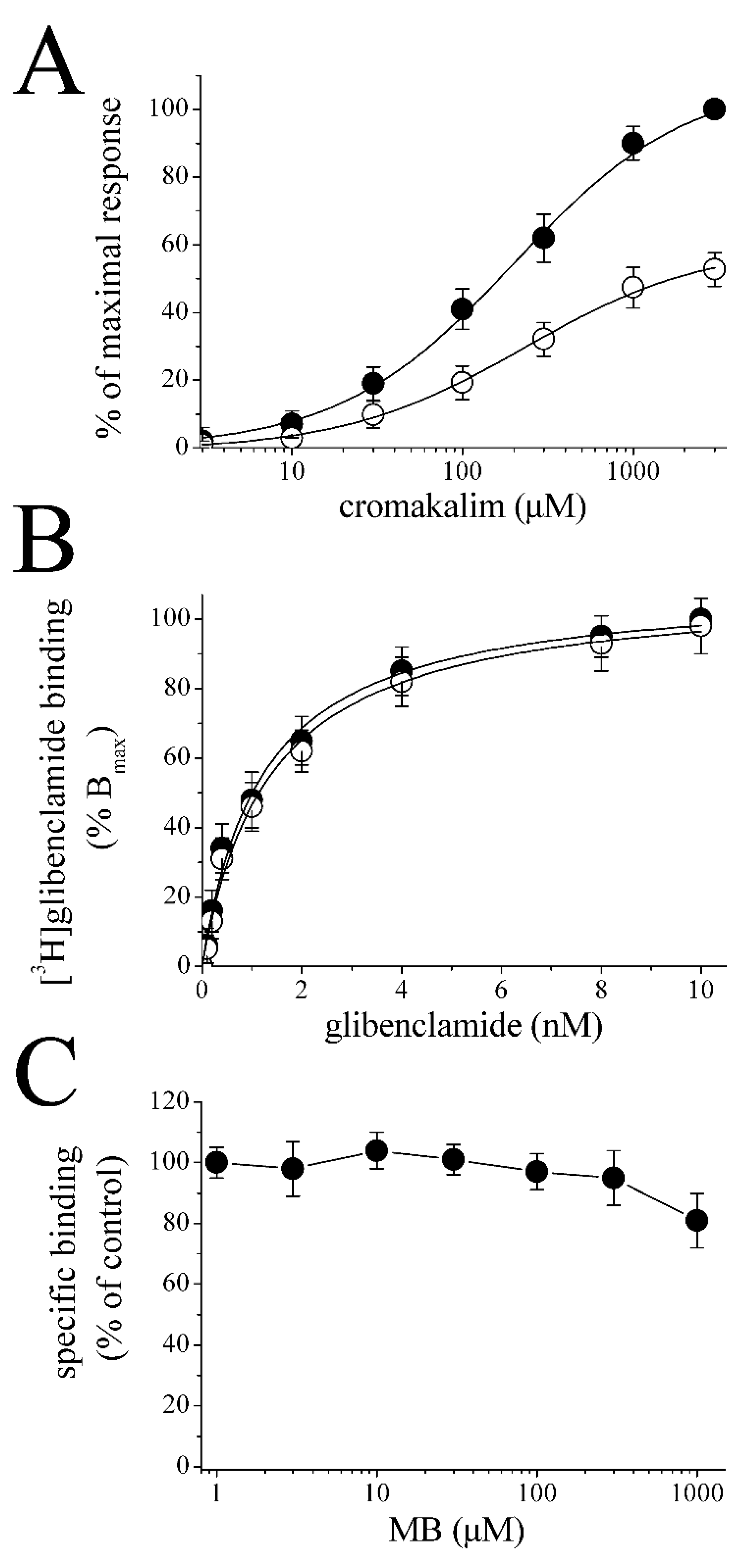{kind=link}
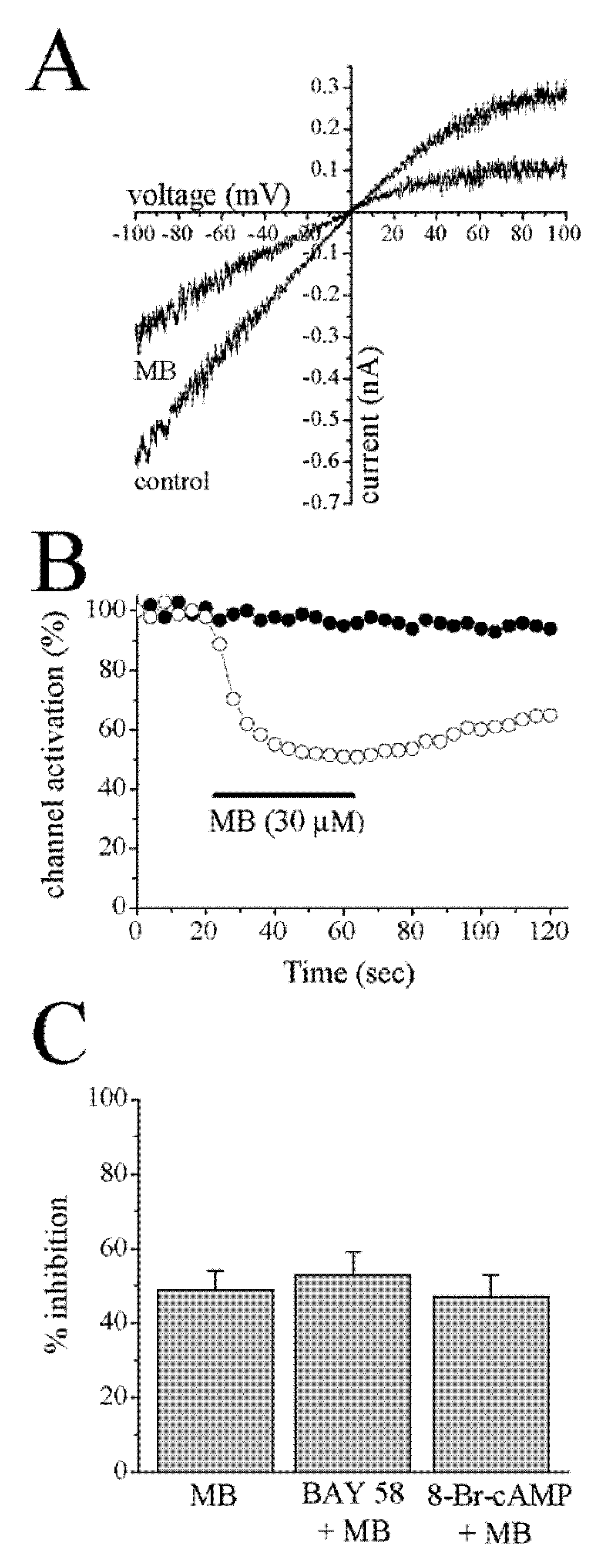{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wainwright, M.; Crossley, K. Methylene Blue—A Therapeutic Dye for All Seasons? J. Chemother. 2002, 14, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Oz, M.; Lorke, D.E.; Petroianu, G.A. Methylene blue and Alzheimer’s disease. Biochem. Pharmacol. 2009, 78, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Oz, M.; Lorke, D.E.; Hasan, M.; Petroianu, G.A. Cellular and molecular actions of Methylene Blue in the nervous system. Med. Res. Rev. 2011, 31, 93–117. [Google Scholar] [CrossRef] [Green Version]
- Oktay, S.; Onat, F.; Karahan, F.; Alican, I.; Ozkutlu, U.; Yegen, B. Effect of Methylene Blue on Blood Pressure in Rats. Pharmacology 1993, 46, 206–210. [Google Scholar] [CrossRef]
- Cheng, X.; Pang, C.C.Y. Pressor and vasoconstrictor effects of methylene blue in endotoxaemic rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1998, 357, 648–653. [Google Scholar] [CrossRef]
- Levin, R.L.; Degrange, M.A.; Bruno, G.F.; Del Mazo, C.D.; Taborda, D.J.; Griotti, J.J.; Boullon, F.J. Methylene blue reduces mortality and morbidity in vasoplegic patients after cardiac surgery. Ann. Thorac. Surg. 2004, 77, 496–499. [Google Scholar] [CrossRef]
- Özal, E.; Kuralay, E.; Yildirim, V.; Kilic, S.; Bolcal, C.; Kücükarslan, N.; Günay, C.; Demirkilic, U.; Tatar, H. Preoperative Methylene Blue Administration in Patients at High Risk for Vasoplegic Syndrome During Cardiac Surgery. Ann. Thorac. Surg. 2005, 79, 1615–1619. [Google Scholar] [CrossRef]
- Lavigne, D. Vasopressin and Methylene Blue: Alternate Therapies in Vasodilatory Shock. Semin. Cardiothorac. Vasc. Anesthesia 2010, 14, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Starkus, J.; Heggeness, S.; Rayner, M. Kinetic analysis of sodium channel block by internal methylene blue in pronased crayfish giant axons. Biophys. J. 1984, 46, 205–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thuneberg, L. Methylene blue as a pharmacological probe of intestinal pacemaker activity. Am. J. Physiol. Liver Physiol. 1990, 258, G992–G994. [Google Scholar] [CrossRef]
- Stockand, J.D.; Sansom, S.C. Activation by methylene blue of large Ca2+-activated K+ channels. Biochim. Biophys. Acta (BBA) Biomembr. 1996, 1285, 123–126. [Google Scholar] [CrossRef] [Green Version]
- Abi-Gerges, N.; Eschenhagen, T.; Hove-Madsen, L.; Fischmeister, R.; Mery, P.-F. Methylene Blue Is a Muscarinic Antagonist in Cardiac Myocytes. Mol. Pharmacol. 1997, 52, 482–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Mansouri, A.S.; Lorke, D.E.; Nurulain, S.M.; Ashoor, A.; Keun-Hang, S.Y.; Petroianu, G.; Isaev, D.; Oz, M. Methylene blue inhibits the function of α7-nicotinic acetylcholine receptors. CNS Neurol. Disord. Drug Targets 2012, 11, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Oz, M.; Isaev, D.; E Lorke, D.; Hasan, M.; Petroianu, G.; Shippenberg, T.S. Methylene blue inhibits function of the 5-HT transporter. Br. J. Pharmacol. 2012, 166, 168–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Küçükkılınç, T.; Özer, I. Multi-site inhibition of human plasma cholinesterase by cationic phenoxazine and phenothiazine dyes. Arch. Biochem. Biophys. 2007, 461, 294–298. [Google Scholar] [CrossRef]
- Ramsay, R.; Dunford, C.; Gillman, K. Methylene blue and serotonin toxicity: Inhibition of monoamine oxidase A (MAO A) confirms a theoretical prediction. Br. J. Pharmacol. 2007, 152, 946–951. [Google Scholar] [CrossRef]
- Quast, U.; Guillon, J.-M.; Cavero, I. Cellular pharmacology of potassium channel openers in vascular smooth muscle. Cardiovasc. Res. 1994, 28, 805–810. [Google Scholar] [CrossRef]
- Mannhold, R. KATP channel openers: Structure-activity relationships and therapeutic potential. Med. Res. Rev. 2004, 24, 213–266. [Google Scholar] [CrossRef]
- Edwards, G.; Weston, A.H. The Pharmacology of ATP-Sensitive Potassium Channels. Annu. Rev. Pharmacol. Toxicol. 1993, 33, 597–637. [Google Scholar] [CrossRef]
- Gribble, F.; Reimann, F. Pharmacological modulation of KATP channels. Biochem. Soc. Trans. 2002, 30, 333–339. [Google Scholar] [CrossRef]
- Rubaiy, H.N. The therapeutic agents that target ATP-sensitive potassium channels. Acta Pharm. 2016, 66, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoshiki, H.; Sunagawa, M.; Seki, T.; Sperelakis, N. ATP-sensitive K+channels in pancreatic, cardiac, and vascular smooth muscle cells. Am. J. Physiol. 1998, 274, C25–C37. [Google Scholar] [CrossRef] [PubMed]
- Nichols, C.G. Adenosine Triphosphate-Sensitive Potassium Currents in Heart Disease and Cardioprotection. Card. Electrophysiol. Clin. 2016, 8, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Tinker, A.; Aziz, Q.; Li, Y.; Specterman, M. ATP-Sensitive Potassium Channels and Their Physiological and Pathophysiological Roles. Compr. Physiol. 2018, 8, 1463–1511. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, W.I. ATP-Sensitive Potassium Channels in the Cerebral Circulation. Stroke 2003, 34, 1547–1552. [Google Scholar] [CrossRef] [Green Version]
- Honoré, E.; Lazdunski, M. Hormone-regulated K+ channels in follicle-enclosed oocytes are activated by vasorelaxing K+ channel openers and blocked by antidiabetic sulfonylureas. Proc. Natl. Acad. Sci. USA 1991, 88, 5438–5442. [Google Scholar] [CrossRef] [Green Version]
- Sakuta, H.; Sekiguchi, M.; Okamoto, K.; Sakai, Y. Inactivation of glibenclamide-sensitive K+ channels in Xenopus oocytes by various calmodulin antagonists. Eur. J. Pharmacol. Mol. Pharmacol. 1992, 226, 199–207. [Google Scholar] [CrossRef]
- Arellano, R.O.; Woodward, R.M.; Miledi, R. ION Channels and Membrane Receptors in Follicle-Enclosed Xenopus Oocytes. Ion Channels 1996, 4, 203–259. [Google Scholar] [CrossRef]
- Zhang, L.; Oz, M.; Weight, F.F. Potentiation of 5-HT3 receptor-mediated responses by protein kinase C activation. Neuroreport 1995, 6, 1464–1468. [Google Scholar] [CrossRef]
- Oz, M.; Zhang, L.; Spivak, C.E. Direct noncompetitive inhibition of 5-HT3 receptor-mediated responses by forskolin and steroids. Arch. Biochem. Biophys. 2002, 404, 293–301. [Google Scholar] [CrossRef]
- Oz, M.; Melia, M.T.; Soldatov, N.M.; Abernethy, D.R.; Morad, M. Functional Coupling of Human L-Type Ca2+ Channels and Angiotensin AT1A Receptors Coexpressed in Xenopus laevis Oocytes: Involvement of the Carboxyl-Terminal Ca2+ Sensors. Mol. Pharmacol. 1998, 54, 1106–1112. [Google Scholar] [CrossRef] [Green Version]
- Oz, M.; Zakharova, I.; Dinc, M.; Shippenberg, T. Cocaine inhibits cromakalim-activated K+ currents in follicle-enclosed Xenopus oocytes. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2004, 369, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Oz, M.; Yang, K.-H.; Dinc, M.; Shippenberg, T.S. The Endogenous Cannabinoid Anandamide Inhibits Cromakalim-Activated K+ Currents in Follicle-Enclosed Xenopus Oocytes. J. Pharmacol. Exp. Ther. 2007, 323, 547–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillemare, E.; Honore, E.; De Weille, J.; Fosset, M.; Lazdunski, M.; Meisheri, K. Functional receptors in Xenopus oocytes for U-37883A, a novel ATP-sensitive K+ channel blocker: Comparison with rat insulinoma cells. Mol. Pharmacol. 1994, 46, 139–145. [Google Scholar] [PubMed]
- Dascal, N. The Use of Xenopus Oocytes for the Study of Ion Channel. CRC Crit. Rev. Biochem. 1987, 22, 317–387. [Google Scholar] [CrossRef]
- Oz, M.; Tchugunova, Y.; Dinc, M. Inhibition of cromakalim-activated K+ current by ethanol in follicle-enclosed Xenopus oocytes. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2003, 367, 80–85. [Google Scholar] [CrossRef]
- Lee, Y.S.; Wurster, R.D. Methylene blue induces cytotoxicity in human brain tumor cells. Cancer Lett. 1995, 88, 141–145. [Google Scholar] [CrossRef]
- Tretter, L.; Horvath, G.; Hölgyesi, A.; Essek, F.; Adam-Vizi, V. Enhanced hydrogen peroxide generation accompanies the beneficial bioenergetic effects of methylene blue in isolated brain mitochondria. Free Radic. Biol. Med. 2014, 77, 317–330. [Google Scholar] [CrossRef] [Green Version]
- Berrocal, M.; Corbacho, I.; Gutierrez-Merino, C.; Mata, A.M. Methylene blue activates the PMCA activity and cross-interacts with amyloid β-peptide, blocking Aβ-mediated PMCA inhibition. Neuropharmacology 2018, 139, 163–172. [Google Scholar] [CrossRef]
- Nemeth, P.R.; Daly, K.; Erde, S.; Wood, J.D. Effects of methylene blue on electrical behavior of myenteric neurons. Experientia 1985, 41, 259–261. [Google Scholar] [CrossRef]
- Saitow, F.; Nakaoka, Y. The photodynamic action of methylene blue on the ion channels of Paramecium causes cell damage. Photochem. Photobiol. 1997, 65, 902–907. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T. Acetylcholine Induces Ca-Dependent K Currents in Rabbit Endothelial Cells. Jpn. J. Pharmacol. 1990, 53, 235–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashcroft, F.M.; Gribble, F.M. New windows on the mechanism of action of KATP channel openers. Trends Pharmacol. Sci. 2000, 21, 439–445. [Google Scholar] [CrossRef]
- Tucker, S.J.; Gribble, F.M.; Zhao, C.; Trapp, S.; Ashcroft, F.M. Truncation of Kir6.2 produces ATP-sensitive K+ channels in the absence of the sulphonylurea receptor. Nature 1997, 387, 179–183. [Google Scholar] [CrossRef]
- Gruetter, C.A.; Kadowitz, P.J.; Ignarro, L.J. Methylene blue inhibits coronary arterial relaxation and guanylate cyclase activation by nitroglycerin, sodium nitrite, and amyl nitrite. Can. J. Physiol. Pharmacol. 1981, 59, 150–156. [Google Scholar] [CrossRef]
- Ignarro, L.J.; Harbison, R.G.; Wood, K.S.; Kadowitz, P.J. Dissimilarities between methylene blue and cyanide on relaxation and cyclic GMP formation in endothelium-intact intrapulmonary artery caused by nitrogen oxide-containing vasodilators and acetylcholine. J. Pharmacol. Exp. Ther. 1986, 236, 30–36. [Google Scholar]
- Sakuta, H.; Okamoto, K.; Watanabe, Y. Modification by cGMP of Glibenclamide-Sensitive K+ Currents in Xenopus Oocytes. Jpn. J. Pharmacol. 1993, 61, 259–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuta, H.; Okamoto, K.; Tandai, M. Atrial natriuretic factor potentiates glibenclamide-sensitive K+ currents via the activation of receptor guanylate cyclase in follicle-enclosed Xenopus oocytes. Eur. J. Pharmacol. Mol. Pharmacol. 1994, 267, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Stasch, J.-P.; Schmidt, P.; Alonso-Alija, C.; Apeler, H.; Dembowsky, K.; Haerter, M.; Heil, M.; Minuth, T.; Perzborn, E.; Pleiss, U.; et al. NO- and haem-independent activation of soluble guanylyl cyclase: Molecular basis and cardiovascular implications of a new pharmacological principle. Br. J. Pharmacol. 2002, 136, 773–783. [Google Scholar] [CrossRef]
- Miledi, R.; Parker, I. Chloride current induced by injection of calcium into Xenopus oocytes. J. Physiol. 1984, 357, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Sakuta, H. Inhibition by histamine H1 receptor antagonists of endogenous glibenclamide-sensitive K+ channels in follicle-enclosed Xenopus oocytes. Eur. J. Pharmacol. Mol. Pharmacol. 1994, 266, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; De Weille, J.R.; Lazdunski, M. Chlorpromazine and related phenothiazines inhibit the ATP-sensitive K+ channel. Eur. J. Pharmacol. 1991, 198, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Gribble, F.M.; E Davis, T.M.; E Higham, C.; Clark, A.; Ashcroft, F.M. The antimalarial agent mefloquine inhibits ATP-sensitive K-channels. Br. J. Pharmacol. 2000, 131, 756–760. [Google Scholar] [CrossRef] [Green Version]
- Sakuta, H.; Yoneda, I. Inhibition by SKF 525A and quinacrine of endogenous glibenclamide-sensitive K+ channels in follicle-enclosed Xenopus oocytes. Eur. J. Pharmacol. 1994, 252, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.M.; Burke, E.P.; Stevens, R.J. Effects of methylene blue on rhythmic activity and membrane potential in the canine proximal colon. Am. J. Physiol. Liver Physiol. 1989, 256, G779–G784. [Google Scholar] [CrossRef]
- Nánási, P.P.; Dely, M. Effects of methylene blue and ascorbate on transmembrane potential in frog skeletal muscle. Gen. Pharmacol. Vasc. Syst. 1995, 26, 1307–1311. [Google Scholar] [CrossRef]
- Kress, M.; Petersen, M.; Reeh, P.W. Methylene blue induces ongoing activity in rat cutaneous primary afferents and depolarization of DRG neurons via a photosensitive mechanism. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1997, 356, 619–625. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isaev, D.; Yang, K.-H.S.; Petroianu, G.; Lorke, D.E.; Oz, M. Methylene Blue Inhibits Cromakalim-Activated K+ Currents in Follicle-Enclosed Oocytes. Membranes 2023, 13, 121. https://doi.org/10.3390/membranes13020121
Isaev D, Yang K-HS, Petroianu G, Lorke DE, Oz M. Methylene Blue Inhibits Cromakalim-Activated K+ Currents in Follicle-Enclosed Oocytes. Membranes. 2023; 13(2):121. https://doi.org/10.3390/membranes13020121
Chicago/Turabian StyleIsaev, Dmytro, Keun-Hang Susan Yang, Georg Petroianu, Dietrich Ernst Lorke, and Murat Oz. 2023. "Methylene Blue Inhibits Cromakalim-Activated K+ Currents in Follicle-Enclosed Oocytes" Membranes 13, no. 2: 121. https://doi.org/10.3390/membranes13020121