

The Interaction of Anthracycline Based Quinone-Chelators with Model Lipid Membranes: 1H NMR and MD Study

Abstract
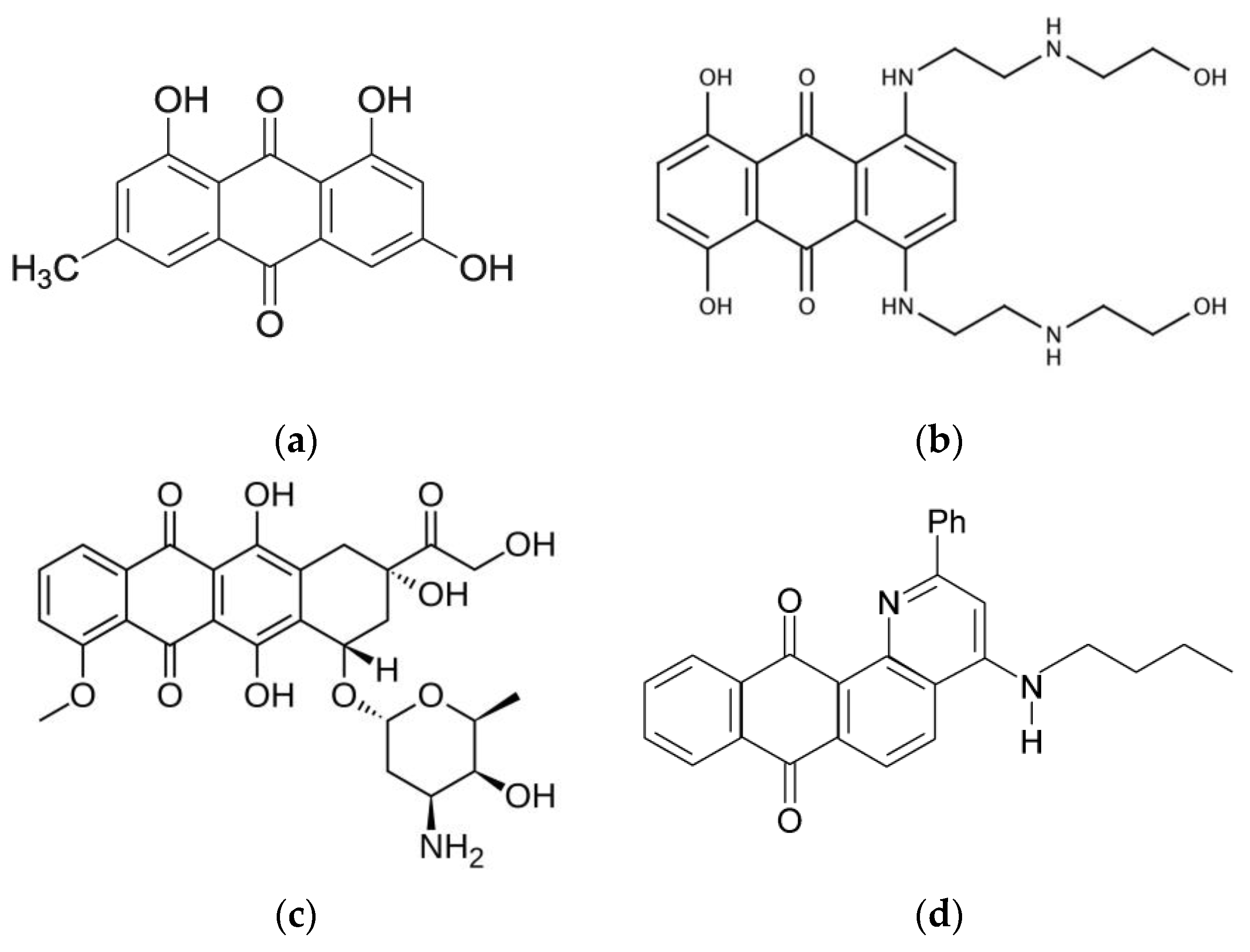
:1. Introduction
2. Materials and Methods
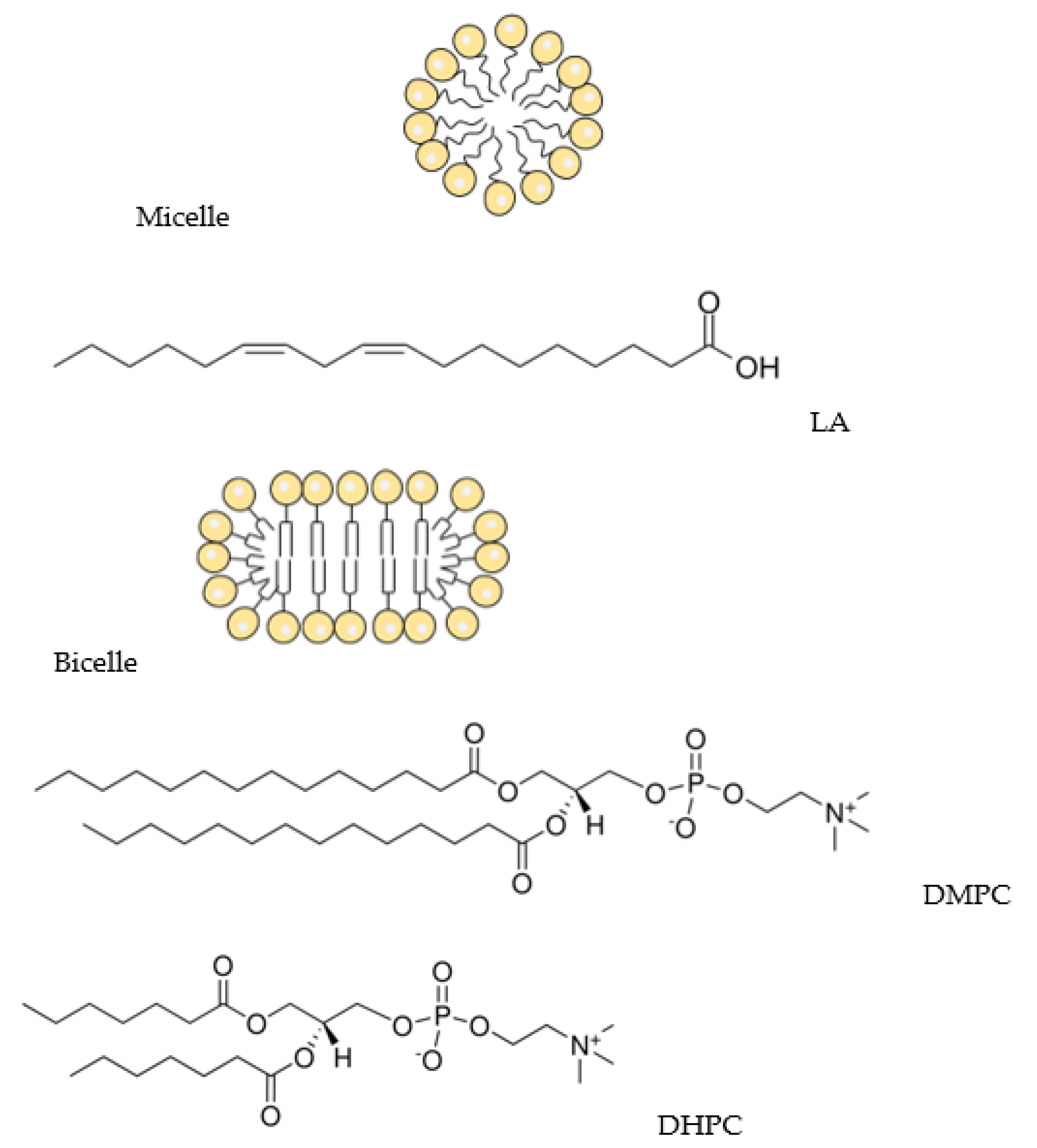
2.1. Materials
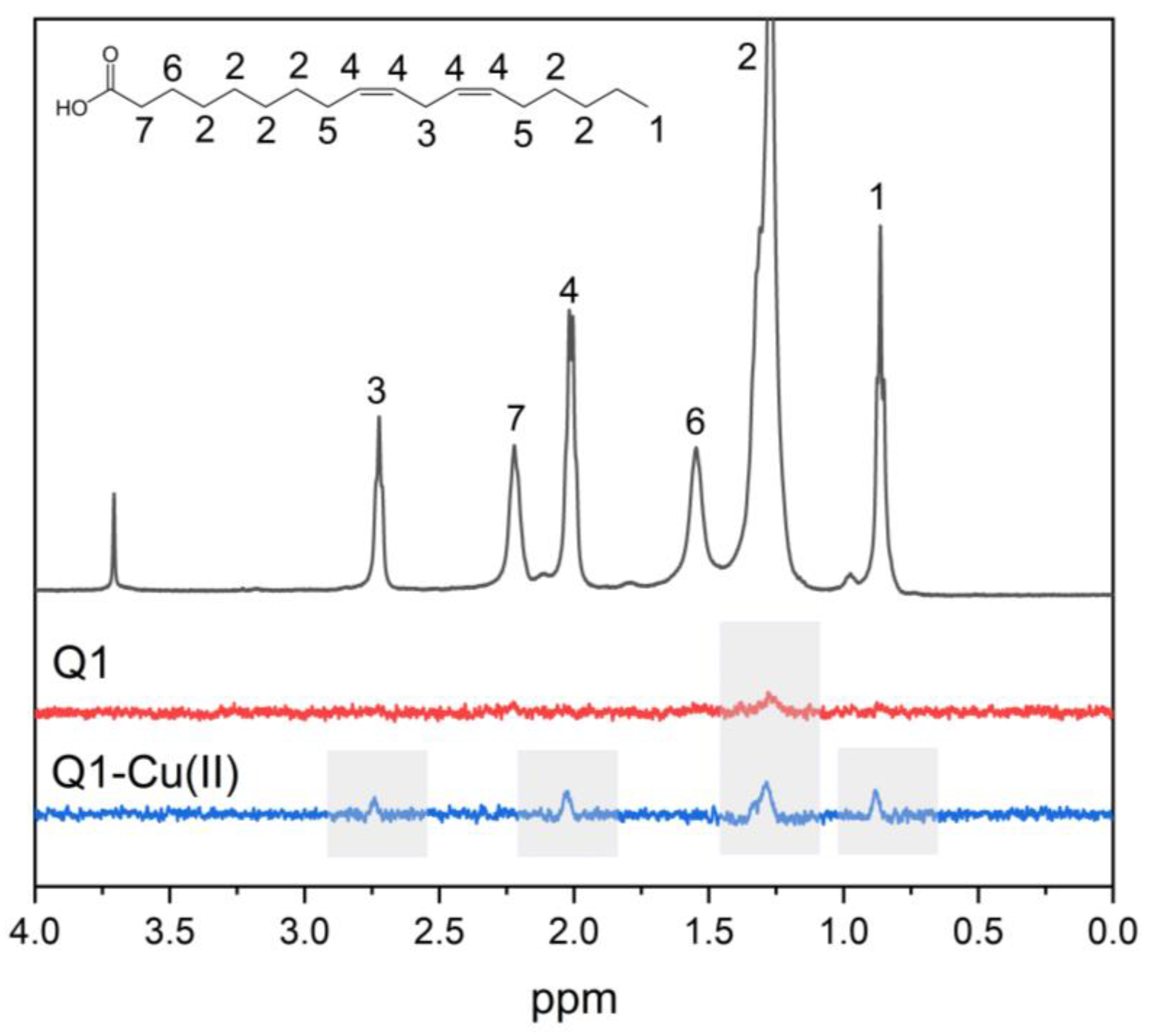
2.2. NMR Study
2.3. Molecular Dynamics Simulations
2.4. Relative Lipophilicity (log P) Determination
3. Results and Discussion
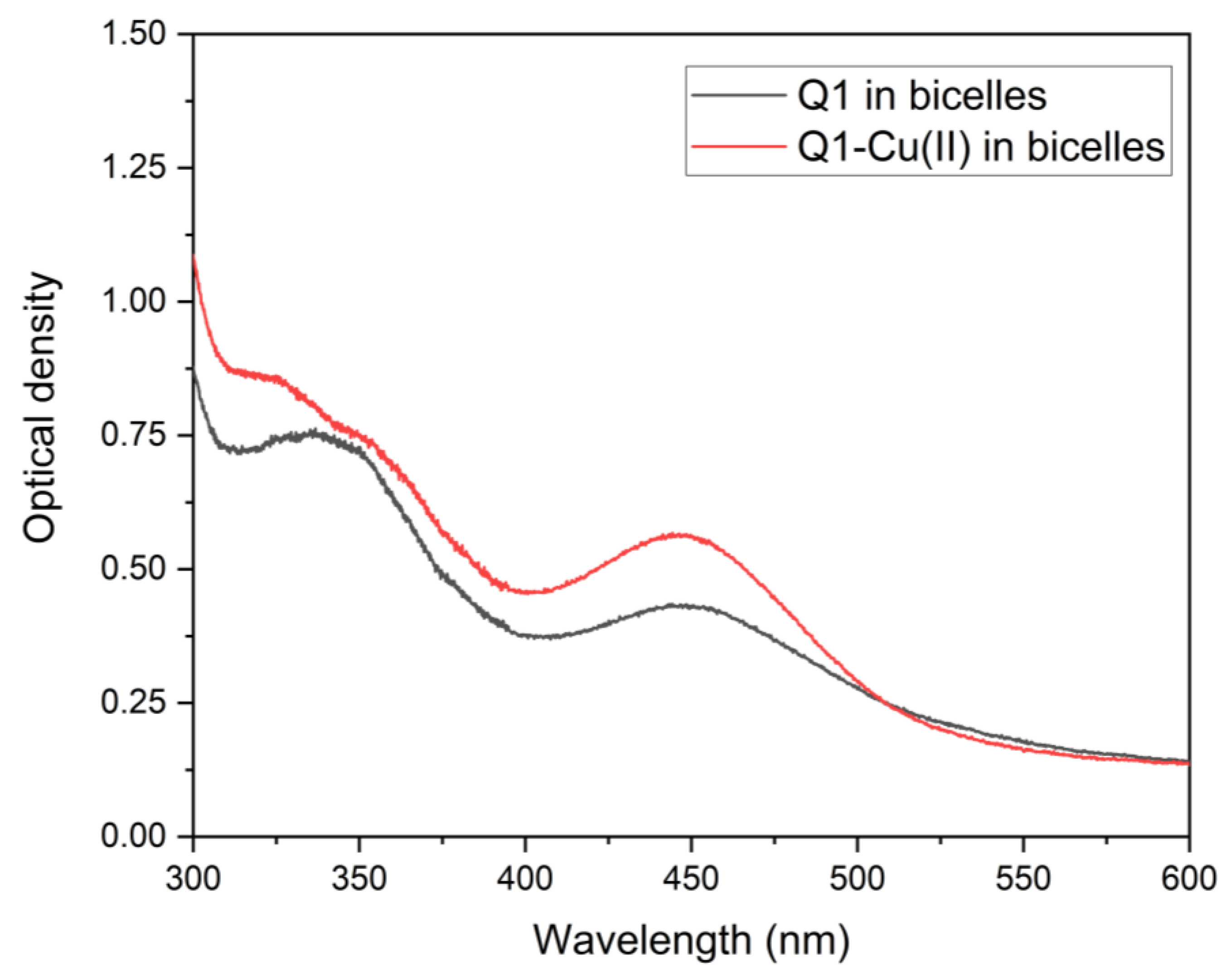
3.1. Q1 and Q1−Cu(II) Interaction with Linoleic Acid Micells
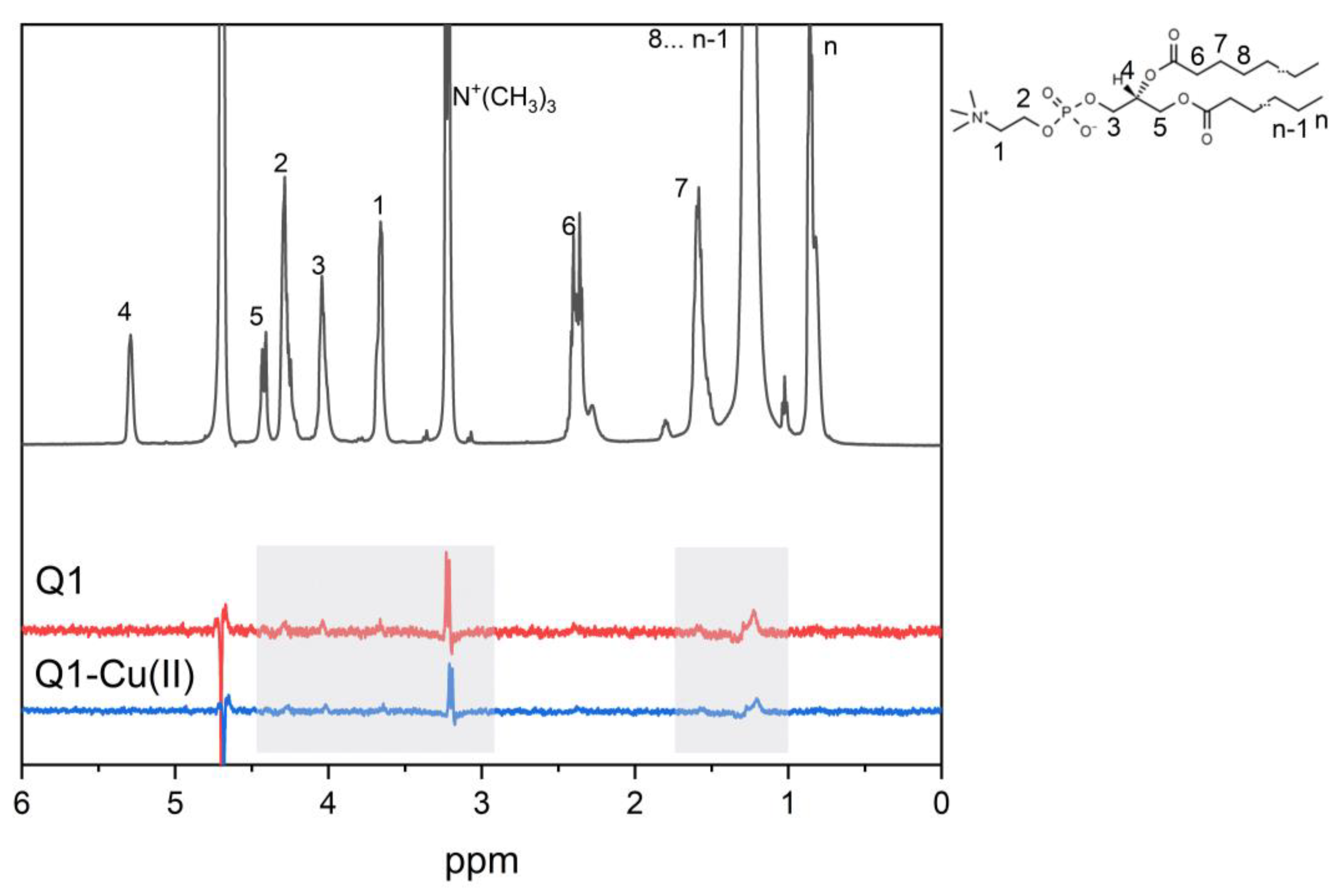
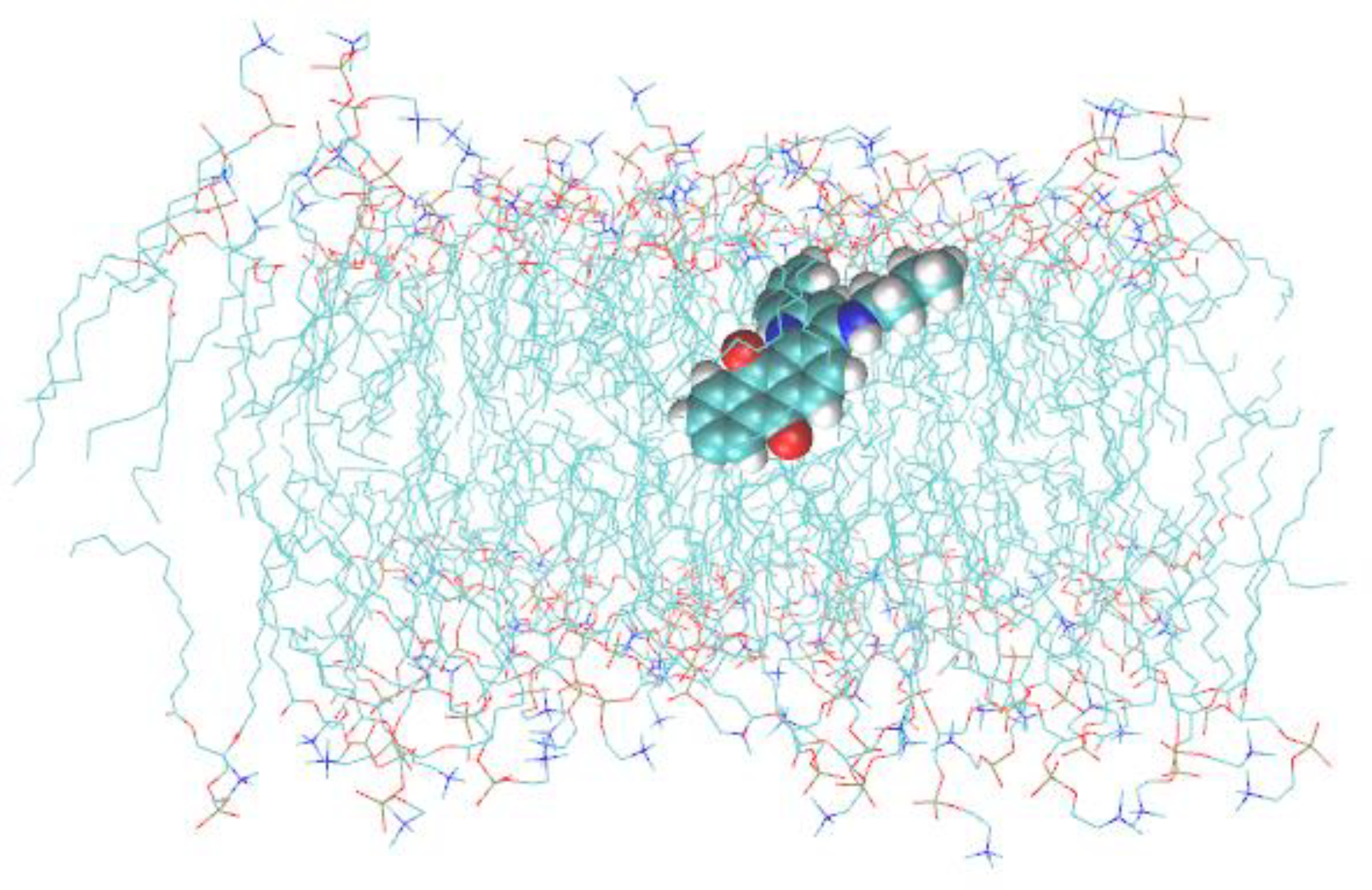
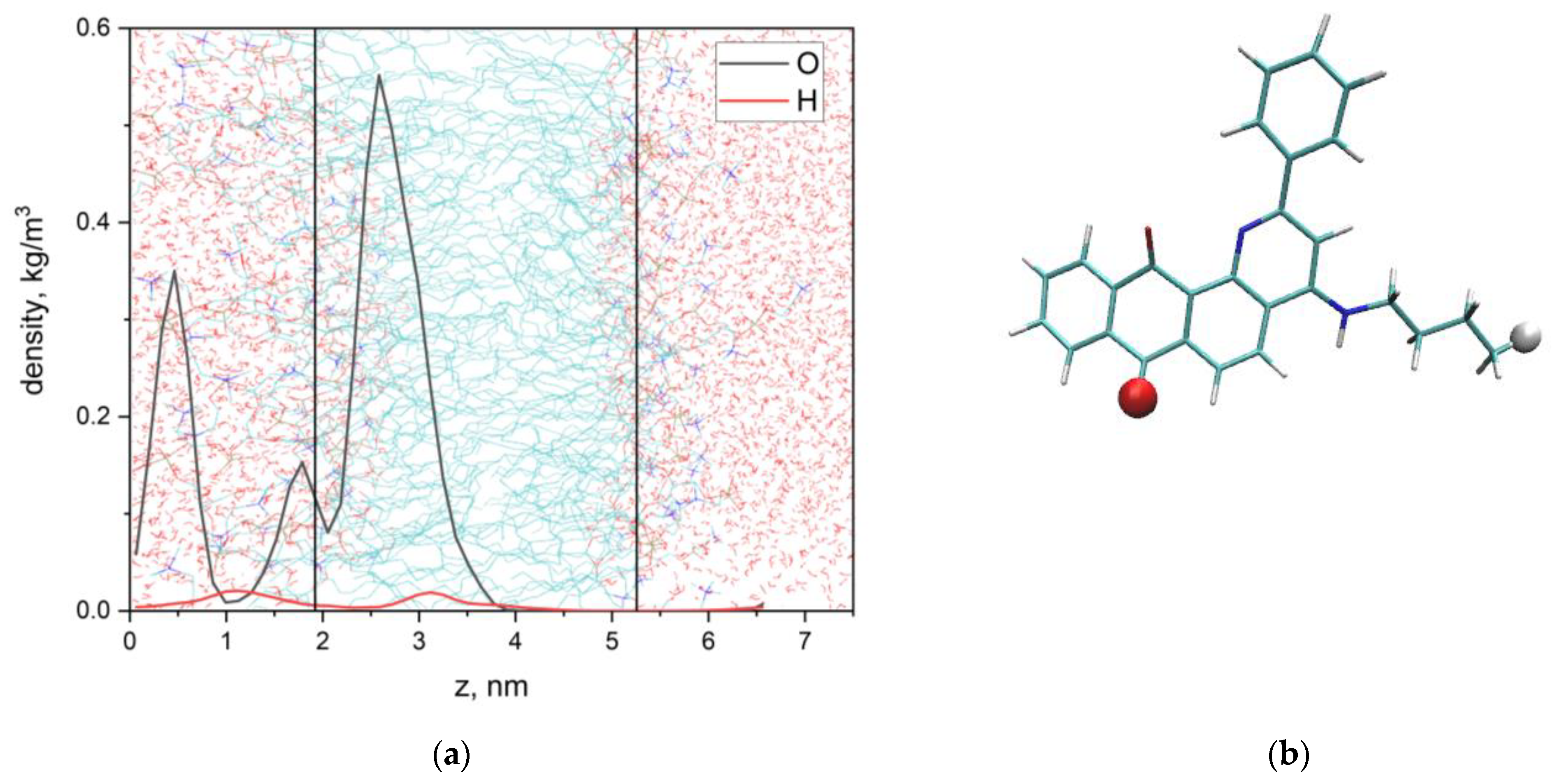
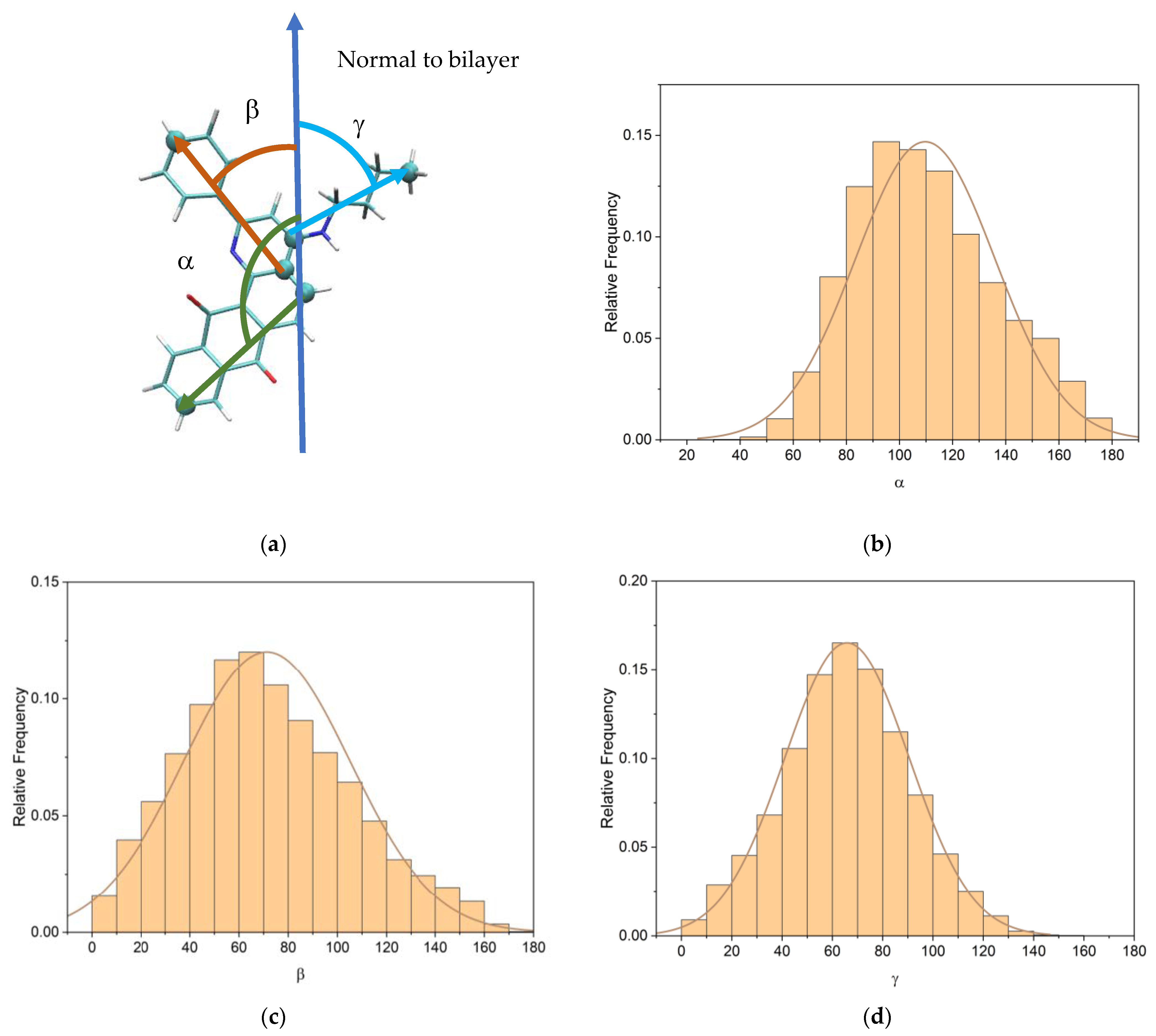
3.2. Q1 and Q1−Cu(II) Interaction with DMPC/DHPC Bicelles
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frézard, F.; Garnier-Suillerot, A. Permeability of lipid bilayer to anthracycline derivatives. Role of the bilayer composition and of the temperature. Biochim. Biophys. Acta—Lipids Lipid Metab. 1998, 1389, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Toroz, D.; Gould, I.R. A computational study of Anthracyclines interacting with lipid bilayers: Correlation of membrane insertion rates, orientation effects and localisation with cytotoxicity. Sci. Rep. 2019, 9, 2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powis, G. Free radical formation by antitumor quinones. Free Radic. Biol. Med. 1989, 6, 63–101. [Google Scholar] [CrossRef] [PubMed]
- Kankeu, C.; Clarke, K.; Passante, E.; Huber, H.J. Doxorubicin-induced chronic dilated cardiomyopathy—The apoptosis hypothesis revisited. J. Mol. Med. 2017, 95, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, B.; Mukherjee, S.; Bhattacharya, B.; Mukherjee, S. Cancer Therapy Using Antibiotics. J. Cancer Ther. 2015, 6, 849–858. [Google Scholar] [CrossRef] [Green Version]
- Hrelia, S.; Fiorentini, D.; Maraldi, T.; Angeloni, C.; Bordoni, A.; Biagi, P.L.; Hakim, G. Doxorubicin induces early lipid peroxidation associated with changes in glucose transport in cultured cardiomyocytes. Biochim. Biophys. Acta—Biomembr. 2002, 1567, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Guo, F.; Guan, Y.; Chen, T.; Ma, K.; Zhang, L.; Wang, Z.; Su, Q.; Feng, L.; Liu, Y.; et al. Novel Anthraquinone Compounds Inhibit Colon Cancer Cell Proliferation via the Reactive Oxygen Species/JNK Pathway. Molecules 2020, 25, 1672. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.S.; Alsantali, R.I.; Jassas, R.S.; Alsimaree, A.A.; Syed, R.; Alsharif, M.A.; Kalpana, K.; Morad, M.; Althagafi, I.I.; Ahmed, S.A. Journey of anthraquinones as anticancer agents—A systematic review of recent literature. RSC Adv. 2021, 11, 35806–35827. [Google Scholar] [CrossRef]
- Pang, M.J.; Yang, Z.; Zhang, X.L.; Liu, Z.F.; Fan, J.; Zhang, H.Y. Physcion, a naturally occurring anthraquinone derivative, induces apoptosis and autophagy in human nasopharyngeal carcinoma. Acta Pharmacol. Sin. 2016, 37, 1623–1640. [Google Scholar] [CrossRef] [Green Version]
- Abdella, B.R.; Fisher, J. A chemical perspective on the anthracycline antitumor antibiotics. Environ. Health Perspect. 1985, 64, 4. [Google Scholar] [CrossRef]
- Wheeler, C.; Rader, R.; Kessel, D. Membrane alterations associated with progressive adriamycin resistance. Biochem. Pharmacol. 1982, 31, 2691–2693. [Google Scholar] [CrossRef] [PubMed]
- Alves, D.S.; Pérez-Fons, L.; Estepa, A.; Micol, V. Membrane-related effects underlying the biological activity of the anthraquinones emodin and barbaloin. Biochem. Pharmacol. 2004, 68, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, J.M.; Burke, T.G.; Tritton, T.R. Cellular transport of anthracyclines by passive diffusion: Implications for drug resistance. Biochem. Pharmacol. 1985, 34, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, J.A.; Kennedy, K.A.; Sartorelli, A.C.; Tritton, T.R. The role of membranes in the mechanism of action of the antineoplastic agent adriamycin. Spin-labeling studies with chronically hypoxic and drug-resistant tumor cells. J. Biol. Chem. 1983, 258, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Escriba, P.V.; Ferrer-Montiel, A.V.; Ferragut, J.A.; Gonzalez-Ros, J.M. Role of membrane lipids in the interaction of daunomycin with plasma membranes from tumor cells: Implications in drug-resistance phenomena. Biochemistry 1990, 29, 7275–7282. [Google Scholar] [CrossRef] [PubMed]
- Mandal, B.; Singha, S.; Dey, S.K.; Mazumdar, S.; Kumar, S.; Karmakar, P.; Das, S. CuII complex of emodin with improved anticancer activity as demonstrated by its performance on HeLa and Hep G2 cells. RSC Adv. 2017, 7, 41403–41418. [Google Scholar] [CrossRef] [Green Version]
- Jabłońska-Trypuć, A.; Świderski, G.; Krętowski, R.; Lewandowski, W. Newly Synthesized Doxorubicin Complexes with Selected Metals-Synthesis, Structure and Anti-Breast Cancer Activity. Molecules 2017, 22, 1106. [Google Scholar] [CrossRef] [Green Version]
- Christidi, E.; Brunham, L.R. Regulated cell death pathways in doxorubicin-induced cardiotoxicity. Cell Death Dis. 2021, 12, 339. [Google Scholar] [CrossRef]
- Rahimipour, S.; Gescheidt, G.; Bilkis, I.; Fridkin, M.; Weiner, L. Towards the Efficiency of Pharmacologically Active Quinoid Compounds: Electron Transfer and Formation of Reactive Oxygen Species. Appl. Magn. Reson. 2009, 37, 629–648. [Google Scholar] [CrossRef]
- Polyakov, N.; Leshina, T.; Fedenok, L.; Slepneva, I.; Kirilyuk, I.; Furso, J.; Olchawa, M.; Sarna, T.; Elas, M.; Bilkis, I.; et al. Redox-Active Quinone Chelators: Properties, Mechanisms of Action, Cell Delivery, and Cell Toxicity. Antioxid. Redox Signal. 2018, 28, 1394–1403. [Google Scholar] [CrossRef]
- Denoyer, D.; Masaldan, S.; La Fontaine, S.; Cater, M.A. Targeting copper in cancer therapy: “Copper That Cancer”. Metallomics 2015, 7, 1459–1476. [Google Scholar] [CrossRef] [PubMed]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupte, A.; Mumper, R.J. Elevated copper and oxidative stress in cancer cells as a target for cancer treatment. Cancer Treat. Rev. 2009, 35, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, D.B.; Jansson, P.J.; Brunk, U.T.; Wong, J.; Ponka, P.; Richardson, D.R. Antitumor activity of metal-chelating compound Dp44mT is mediated by formation of a redox-active copper complex that accumulates in lysosomes. Cancer Res. 2011, 71, 5871–5880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiallo, M.M.L.; Garnier-Suillerot, A. Metal Anthracycline Complexes as a New Class of Anthracycline Derivatives. Pd(II)-Adriamycin and Pd(II)-Daunorubicin Complexes: Physicochemical Characteristics and Antitumor Activity. Biochemistry 1986, 25, 924–930. [Google Scholar] [CrossRef]
- Eliot, H.; Gianni, L.; Myers, C. Oxidative Destruction of DNA by the Adriamycin-iron Complex. Biochemistry 1984, 23, 928–936. [Google Scholar] [CrossRef]
- Myers, C.E.; Gianni, L.; Simone, C.B.; Klecker, R.; Greene, R. Oxidative destruction of erythrocyte ghost membranes catalyzed by the doxorubicin-iron complex. Biochemistry 1982, 21, 1707–1713. [Google Scholar] [CrossRef]
- Mizutani, H.; Oikawa, S.; Hiraku, Y.; Murata, M.; Kojima, M.; Kawanishi, S. Distinct mechanisms of site-specific oxidative DNA damage by doxorubicin in the presence of copper(II) and NADPH-cytochrome P450 reductase. Cancer Sci. 2003, 94, 686–691. [Google Scholar] [CrossRef]
- Mizutani, H.; Nishimoto, A.; Hotta, S.; Ikemura, K.; Imai, M.; Miyazawa, D.; Ohta, K.; Ikeda, Y.; Maeda, T.; Yoshikawa, M.; et al. Oxidative DNA Damage Induced by Pirarubicin, an Anthracycline Anticancer Agent, in the Presence of Copper(II). Anticancer Res. 2018, 38, 2643–2648. [Google Scholar] [CrossRef]
- Yang, P.; Wang, H.; Gao, F.; Yang, B. Antitumor activity of the Cu(II)-mitoxantrone complex and its interaction with deoxyribonucleic acid. J. Inorg. Biochem. 1996, 62, 137–145. [Google Scholar] [CrossRef]
- Rao, V.A. Iron Chelators with Topoisomerase-Inhibitory Activity and Their Anticancer Applications. Antioxid. Redox Signal. 2013, 18, 930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monti, E.; Paracchini, L.; Piccinini, F.; Malatesta, V.; Morazzoni, F.; Supino, R. Cardiotoxicity and antitumor activity of a copper(II)-doxorubicin chelate. Cancer Chemother. Pharmacol. 1990, 25, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Dikalov, S.; Alov, P.; Rangelova, D. Role of Iron Ion Chelation by Quinones in Their Reduction, OH-Radical Generation, and Lipid Peroxidation. Biochem. Biophys. Res. Commun. 1993, 195, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Selyutina, O.Y.; Kononova, P.A.; Koshman, V.E.; Fedenok, L.G.; Polyakov, N.E. The Interplay of Ascorbic Acid with Quinones-Chelators—Influence on Lipid Peroxidation: Insight into Anticancer Activity. Antioxidants 2022, 11, 376. [Google Scholar] [CrossRef] [PubMed]
- Sanders, C.R.; Prosser, R.S. Bicelles: A model membrane system for all seasons? Structure 1998, 6, 1227–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchison, J.M.; Shih, K.C.; Scheidt, H.A.; Fantin, S.M.; Parson, K.F.; Pantelopulos, G.A.; Harrington, H.R.; Mittendorf, K.F.; Qian, S.; Stein, R.A.; et al. Bicelles Rich in both Sphingolipids and Cholesterol and Their Use in Studies of Membrane Proteins. J. Am. Chem. Soc. 2020, 142, 12715–12729. [Google Scholar] [CrossRef] [PubMed]
- Dikalov, S.I.; Rumyantseva, G.V.; Piskunov, A.V.; Weiner, L.M. Role of Quinone-Iron(III) Interaction in NADPH-Dependent Enzymatic Generation of Hydroxyl Radicals. Biochemistry 1992, 31, 8947–8953. [Google Scholar] [CrossRef]
- Stroet, M.; Caron, B.; Visscher, K.M.; Geerke, D.P.; Malde, A.K.; Mark, A.E. Automated Topology Builder Version 3.0: Prediction of Solvation Free Enthalpies in Water and Hexane. J. Chem. Theory Comput. 2018, 14, 5834–5845. [Google Scholar] [CrossRef]
- Poger, D.; Mark, A.E. On the validation of molecular dynamics simulations of saturated and cis-monounsaturated phosphatidylcholine lipid bilayers: A comparison with experiment. J. Chem. Theory Comput. 2010, 6, 325–336. [Google Scholar] [CrossRef]
- Parrinello, M.; Rahman, A. Polymorphic transitions in single crystals: A new molecular dynamics method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Hoover, W.G. Canonical dynamics: Equilibrium phase-space distributions. Phys. Rev. A 1985, 31, 1695–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef] [Green Version]
- da Cunha, A.R.; Duarte, E.L.; Stassen, H.; Lamy, M.T.; Coutinho, K. Experimental and theoretical studies of emodin interacting with a lipid bilayer of DMPC. Biophys. Rev. 2017, 9, 729–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, D.R.; Wis Vitolo, L.M.; Hefter, G.T.; May, P.M.; Clare, B.W.; Webb, J.; Wilairat, P. Iron chelators of the pyridoxal isonicotinoyl hydrazone class Part I. Ionisation characteristics of the ligands and their relevance to biological properties. Inorg. Chim. Acta 1990, 170, 165–170. [Google Scholar] [CrossRef]
- Richardson, D.R.; Tran, E.H.; Ponka, P. The potential of iron chelators of the pyridoxal isonicotinoyl hydrazone class as effective antiproliferative agents. Blood 1995, 86, 4295–4306. [Google Scholar] [CrossRef] [Green Version]
- Alrushaid, S.; Sayre, C.L.; Yáñez, J.A.; Forrest, M.L.; Senadheera, S.N.; Burczynski, F.J.; Löbenberg, R.; Davies, N.M. Pharmacokinetic and Toxicodynamic Characterization of a Novel Doxorubicin Derivative. Pharmaceutics 2017, 9, 35. [Google Scholar] [CrossRef] [Green Version]
- Selyutina, O.Y.; Kononova, P.A.; Koshman, V.E.; Shelepova, E.A.; Azad, M.G.; Afroz, R.; Dharmasivam, M.; Bernhardt, P.V.; Polyakov, N.E.; Richardson, D.R. Ascorbate-and iron-driven redox activity of Dp44mT and Emodin facilitates peroxidation of micelles and bicelles. Biochim. Biophys. Acta—Gen. Subj. 2022, 1866, 130078. [Google Scholar] [CrossRef]
- Burns, C.P.; Haugstad, B.N.; Mossman, C.J.; North, J.A.; Ingraham, L.M. Membrane lipid alteration: Effect on cellular uptake of mitoxantrone. Lipids 1988, 23, 393–397. [Google Scholar] [CrossRef]
- Stefani, C.; Jansson, P.J.; Gutierrez, E.; Bernhardt, P.V.; Richardson, D.R.; Kalinowski, D.S. Alkyl substituted 2′-benzoylpyridine thiosemicarbazone chelators with potent and selective anti-neoplastic activity: Novel ligands that limit methemoglobin formation. J. Med. Chem. 2013, 56, 357–370. [Google Scholar] [CrossRef]
- Stefani, C.; Punnia-Moorthy, G.; Lovejoy, D.B.; Jansson, P.J.; Kalinowski, D.S.; Sharpe, P.C.; Bernhardt, P.V.; Richardson, D.R. Halogenated 2′-benzoylpyridine thiosemicarbazone (XBpT) chelators with potent and selective anti-neoplastic activity: Relationship to intracellular redox activity. J. Med. Chem. 2011, 54, 6936–6948. [Google Scholar] [CrossRef]
- Selyutina, O.Y.; Kononova, P.A.; Polyakov, N.E. Experimental and Theoretical Study of Emodin Interaction with Phospholipid Bilayer and Linoleic Acid. Appl. Magn. Reson. 2020, 51, 951–960. [Google Scholar] [CrossRef]
- Spiteller, G. Linoleic acid peroxidation—The dominant lipid peroxidation process in low density lipoprotein—And its relationship to chronic diseases. Chem. Phys. Lipids 1998, 95, 105–162. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C. An update on products and mechanisms of lipid peroxidation. Mol. Nutr. Food Res. 2009, 53, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotticelli, M.G.; Crabbe, A.M.; Wilson, R.B.; Shchepinov, M.S. Insights into the role of oxidative stress in the pathology of Friedreich ataxia using peroxidation resistant polyunsaturated fatty acids. Redox Biol. 2013, 1, 398–404. [Google Scholar] [CrossRef] [Green Version]
- Struppe, J.; Whiles, J.A.; Void, R.R. Acidic Phospholipid Bicelles: A Versatile Model Membrane System. Biophys. J. 2000, 78, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Mastova, A.V.; Selyutina, O.Y.; Evseenko, V.I.; Polyakov, N.E. Photoinduced Oxidation of Lipid Membranes in the Presence of the Nonsteroidal Anti-Inflammatory Drug Ketoprofen. Membranes 2022, 12, 251. [Google Scholar] [CrossRef]
- Selyutina, O.Y.; Kononova, P.A.; Polyakov, N.E. Effect of glycyrrhizic acid on phospholipid membranes in media with different pH. Russ. Chem. Bull. 2022, 70, 2434–2439. [Google Scholar] [CrossRef]
- Ellena, J.F.; Lepore, L.S.; Cafiso, D.S. Estimating lipid lateral diffusion in phospholipid vesicles from carbon-13 spin-spin relaxation. J. Phys. Chem. 1993, 97, 2952–2957. [Google Scholar] [CrossRef]
- Lepore, L.S.; Ellena, J.F.; Cafiso, D.S. Comparison of the lipid acyl chain dynamics between small and large unilamellar vesicles. Biophys. J. 1992, 61, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Richards, M.J.; Hsia, C.Y.; Singh, R.R.; Haider, H.; Kumpf, J.; Kawate, T.; Daniel, S. Membrane Protein Mobility and Orientation Preserved in Supported Bilayers Created Directly from Cell Plasma Membrane Blebs. Langmuir 2016, 32, 2963–2974. [Google Scholar] [CrossRef]
- Tian, J.; Sethi, A.; Swanson, B.I.; Goldstein, B.; Gnanakaran, S. Taste of Sugar at the Membrane: Thermodynamics and Kinetics of the Interaction of a Disaccharide with Lipid Bilayers. Biophys. J. 2013, 104, 622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdanov, M.; Mileykovskaya, E.; Dowhan, W. Lipids in the Assembly of Membrane Proteins and Organization of Protein Supercomplexes: Implications for Lipid-Linked Disorders. Subcell. Biochem. 2008, 49, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, A.C.; Magarkar, A.; Horta, M.; Lima, J.L.F.C.; Bunker, A.; Nunes, C.; Reis, S. Influence of doxorubicin on model cell membrane properties: Insights from in vitro and in silico studies. Sci. Rep. 2017, 7, 6343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hell, A.J.; Melo, M.N.; Van Blitterswijk, W.J.; Gueth, D.M.; Braumuller, T.M.; Pedrosa, L.R.C.; Song, J.Y.; Marrink, S.J.; Koning, G.A.; Jonkers, J.; et al. Defined lipid analogues induce transient channels to facilitate drug-membrane traversal and circumvent cancer therapy resistance. Sci. Rep. 2013, 3, 1949. [Google Scholar] [CrossRef] [Green Version]
- Tritton, T.R.; Yee, G. The Anticancer Agent Adriamycin Can Be Actively Cytotoxic Without Entering Cells. Science 1982, 217, 248–250. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N+(CH3)3 | CH2 | CH3 | |
|---|---|---|---|
| w/o Q1 | 0.81 ± 0.06 s | 1.120 ± 0.04 s | 1.370 ± 0.07 s |
| with Q1 | 0.7 ± 0.07 s | 0.8 ± 0.08 s | 1.170 ± 0.1 s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selyutina, O.Y.; Mastova, A.V.; Polyakov, N.E. The Interaction of Anthracycline Based Quinone-Chelators with Model Lipid Membranes: 1H NMR and MD Study. Membranes 2023, 13, 61. https://doi.org/10.3390/membranes13010061
Selyutina OY, Mastova AV, Polyakov NE. The Interaction of Anthracycline Based Quinone-Chelators with Model Lipid Membranes: 1H NMR and MD Study. Membranes. 2023; 13(1):61. https://doi.org/10.3390/membranes13010061
Chicago/Turabian StyleSelyutina, Olga Yu., Anna V. Mastova, and Nikolay E. Polyakov. 2023. "The Interaction of Anthracycline Based Quinone-Chelators with Model Lipid Membranes: 1H NMR and MD Study" Membranes 13, no. 1: 61. https://doi.org/10.3390/membranes13010061