
Involvement of Multidrug Resistance Modulators in the Regulation of the Mitochondrial Permeability Transition Pore


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Preparation of Rat Liver Mitochondria
2.3. Determination of the Ca2+-Induced MPTP Opening
2.4. Determination of Swelling of Mitochondria
2.5. Statistical Analysis
3. Results
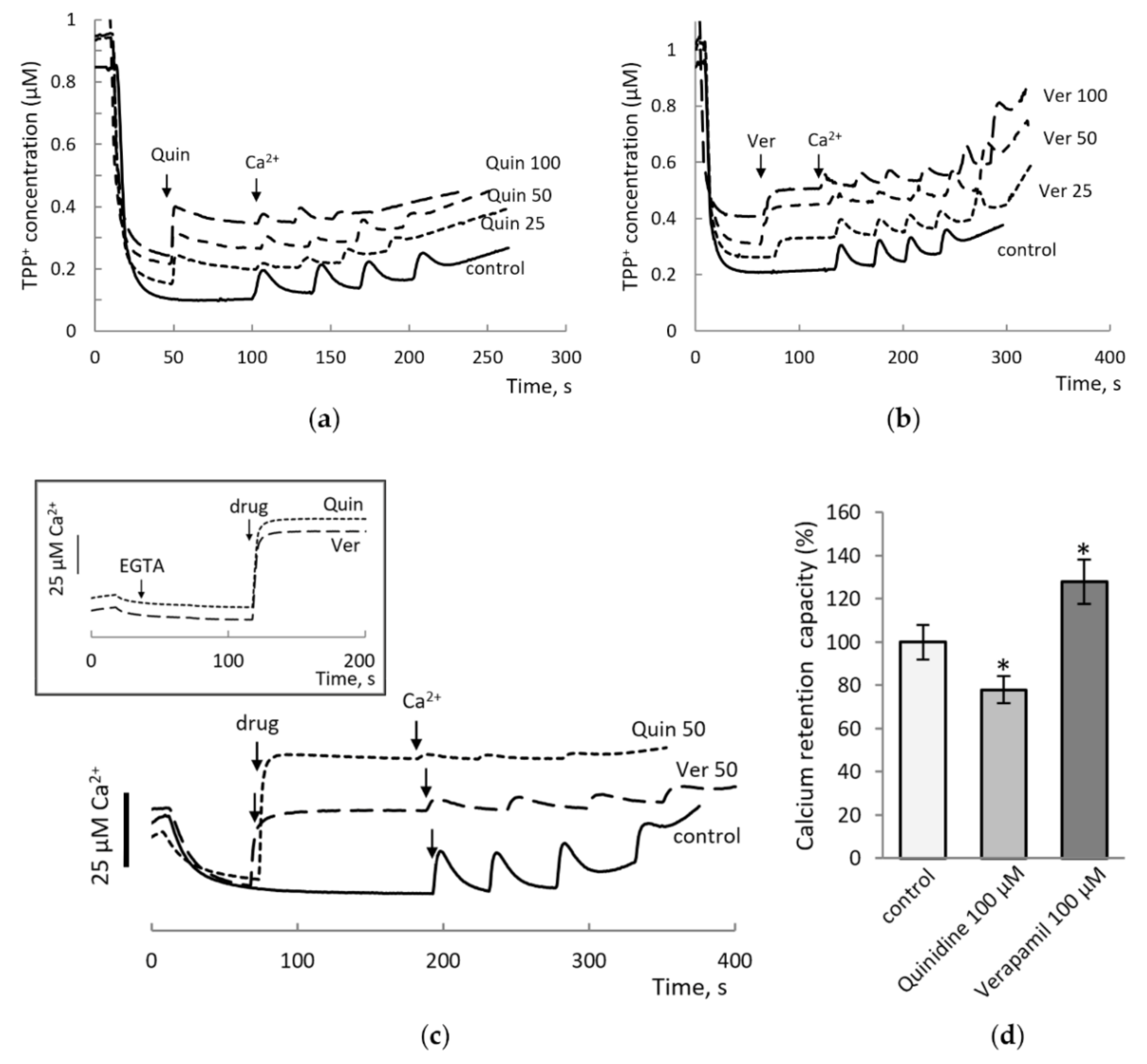
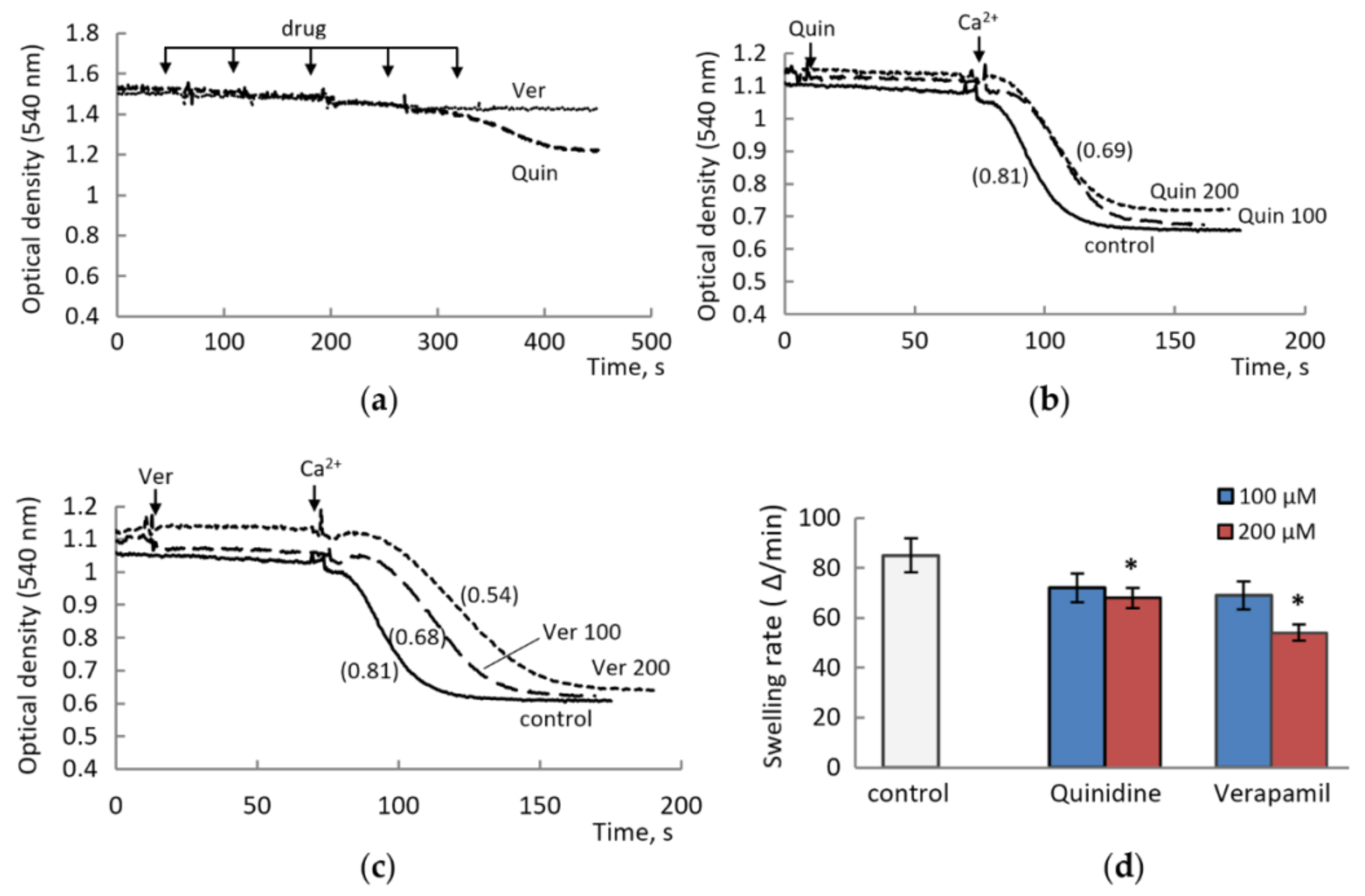
3.1. The Effect of Drugs Was Assessed by Changing the Threshold Concentrations of Calcium Ions Required to Open MPTP
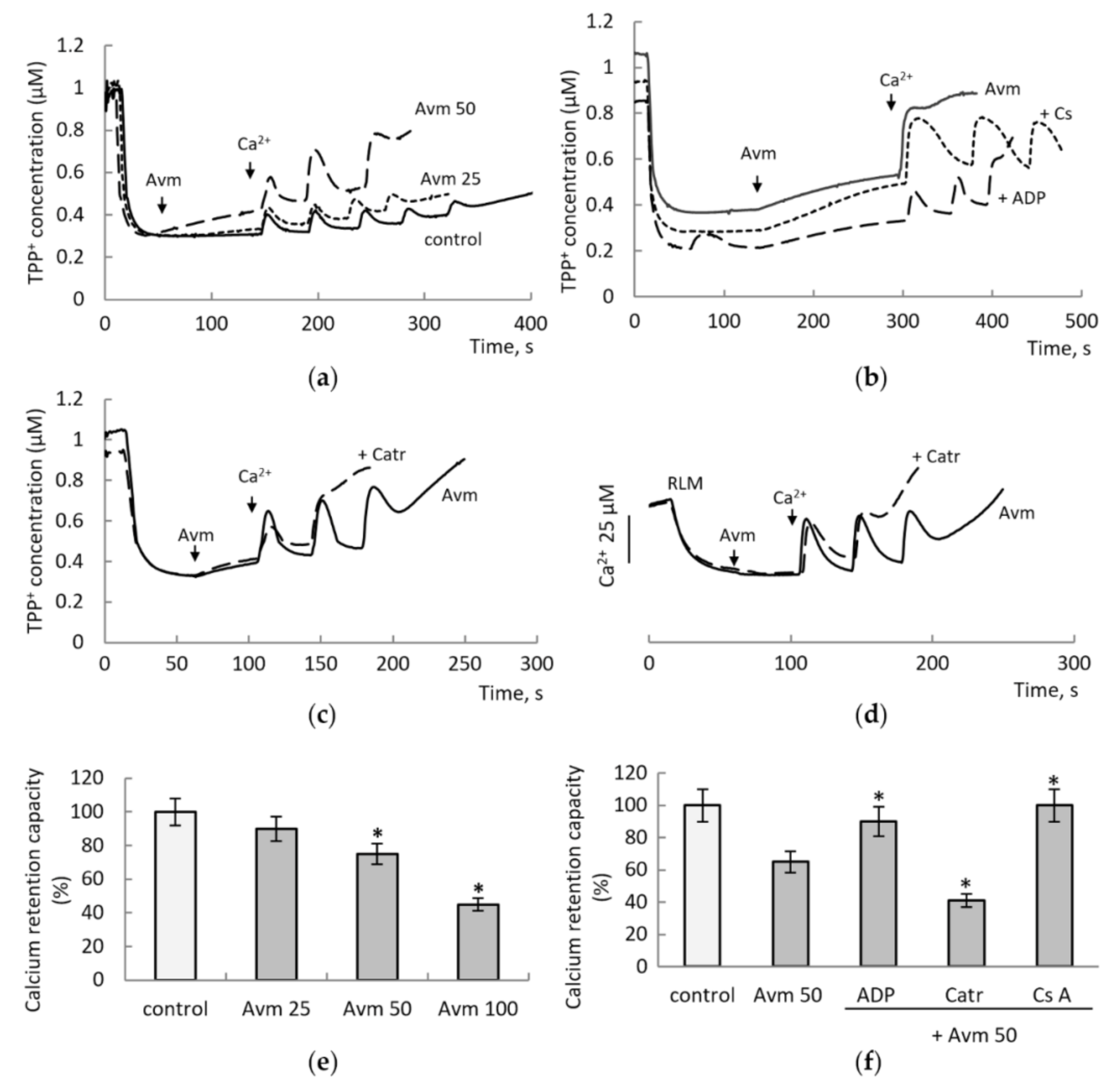
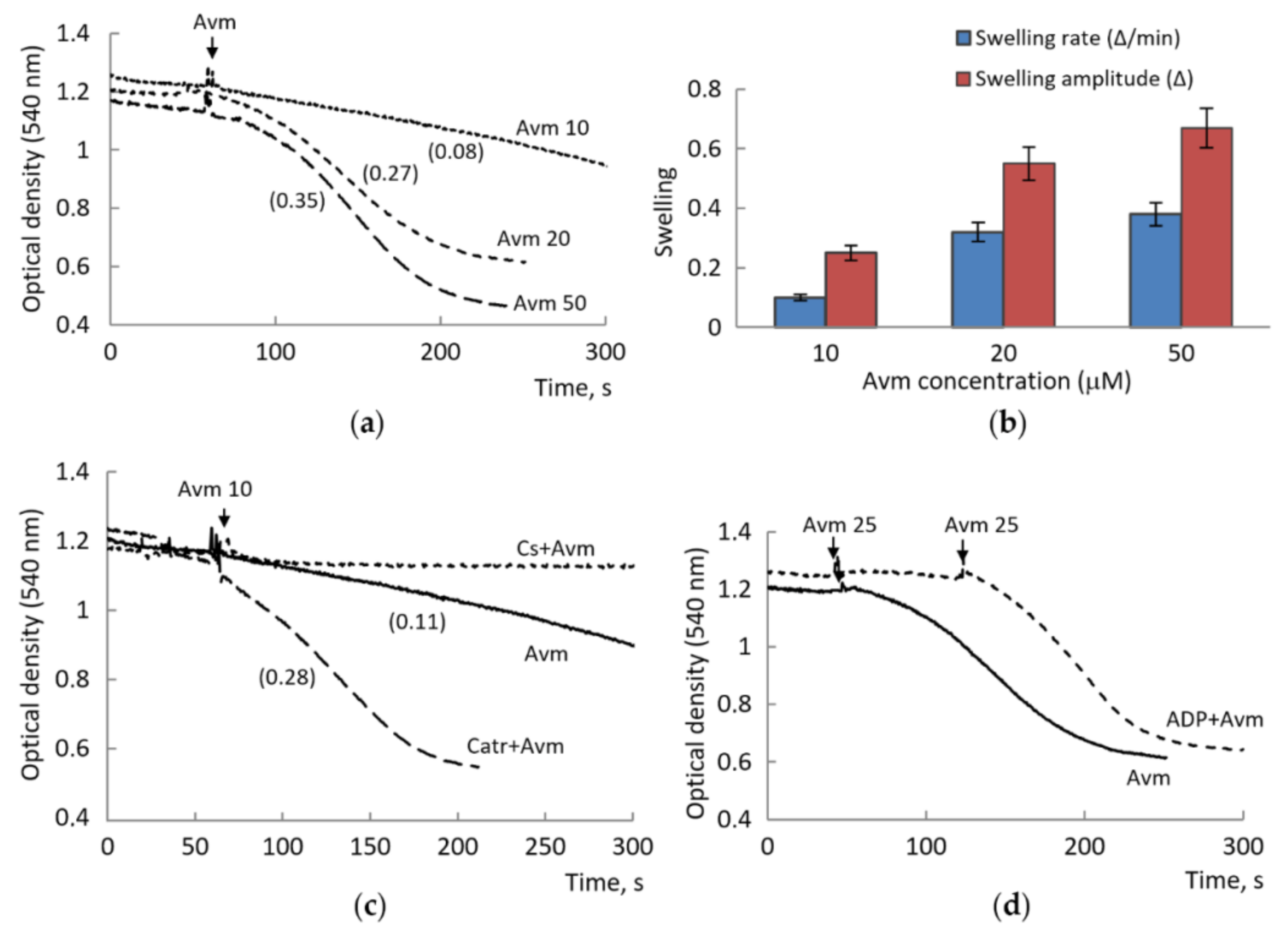
3.2. Influence of Avermectin on the MPTP Opening Induced by Calcium Ions
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Goebel, J.; Chmielewski, J.; Hrycyna, C.A. The roles of the human ATP-binding cassette transporters P-glycoprotein and ABCG2 in multidrug resistance in cancer and at endogenous sites: Future opportunities for structure-based drug design of inhibitors. Cancer Drug Resist 2021, 4, 784–804. [Google Scholar] [CrossRef] [PubMed]
- Seeger, M.A.; van Veen, H.W. Molecular basis of multidrug transport by ABC transporters. Biochim. Biophys. Acta-Proteins Proteom. 2009, 1794, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Zheng, Y.; Ma, L.; Tian, L.; Sun, Q. Clinically-Relevant ABC Transporter for Anti-Cancer Drug Resistance. Front Pharmacol. 2021, 12, 648407. [Google Scholar] [CrossRef]
- Robert, J. Approaches to multidrug resistance reversal. Expert Opin. Investig. Drugs 1998, 7, 929–939. [Google Scholar] [CrossRef]
- Werner, P.; Hilgeroth, A. Arise of MDR Inhibitors for Anticancer Therapy. Anti-Cancer Agents Med. Chem. 2021. [Google Scholar] [CrossRef]
- Ledwitch, K.V.; Roberts, A.G. Cardiovascular Ion Channel Inhibitor Drug-Drug Interactions with P-glycoprotein. AAPS J. 2016, 19, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Huang, G.; Wu, J.; Wu, Q.; Gao, S.; Yan, Z.; Lei, J.; Yan, N. Molecular Basis for Ligand Modulation of a Mammalian Voltage-Gated Ca2+ Channel. Cell 2019, 177, 1495–1506.e12. [Google Scholar] [CrossRef]
- Carraro, M.; Carrer, A.; Urbani, A.; Bernardi, P. Molecular nature and regulation of the mitochondrial permeability transition pore(s), drug target(s) in cardioprotection. J. Mol. Cell. Cardiol. 2020, 144, 76–86. [Google Scholar] [CrossRef]
- Bonora, M.; Giorgi, C.; Pinton, P. Molecular mechanisms and consequences of mitochondrial permeability transition. Nat. Rev. Mol. Cell Biol. 2021, 23, 266–285. [Google Scholar] [CrossRef]
- Rottenberg, H.; Hoek, J. The Mitochondrial Permeability Transition: Nexus of Aging, Disease and Longevity. Cells 2021, 10, 79. [Google Scholar] [CrossRef]
- Ponne, S.; Kumar, C.R.; Boopathy, R. Verapamil attenuates scopolamine induced cognitive deficits by averting oxidative stress and mitochondrial injury—A potential therapeutic agent for Alzheimer’s Disease. Metab. Brain Dis. 2019, 35, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Jangholi, E.; Sharifi, Z.N.; Hoseinian, M.; Zarrindast, M.-R.; Rahimi, H.R.; Mowla, A.; Aryan, H.; Javidi, M.A.; Parsa, Y.; Ghaffarpasand, F.; et al. Verapamil Inhibits Mitochondria-Induced Reactive Oxygen Species and Dependent Apoptosis Pathways in Cerebral Transient Global Ischemia/Reperfusion. Oxidative Med. Cell. Longev. 2020, 2020, 5872645. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Tarapcsák, S.; Gyöngy, Z.; Ritter, Z.; Batta, G.; Bosire, R.; Remenyik, J.; Goda, K. Effects of Polyphenols on P-Glycoprotein (ABCB1) Activity. Pharmaceutics 2021, 13, 2062. [Google Scholar] [CrossRef] [PubMed]
- Fedotcheva, T.A.; Sheichenko, O.P.; Fedotcheva, N.I. New Properties and Mitochondrial Targets of Polyphenol Agrimoniin as a Natural Anticancer and Preventive Agent. Pharmaceutics 2021, 13, 2089. [Google Scholar] [CrossRef]
- Fedotcheva, T.A.; Fedotcheva, N.I.; Shimanovsky, N.L. Progestins as Anticancer Drugs and Chemosensitizers, New Targets and Applications. Pharmaceutics 2021, 13, 1616. [Google Scholar] [CrossRef]
- Kudryavtsev, K.V.; Sokolov, M.N.; Varpetyan, E.E.; Kirsanova, A.A.; Fedotcheva, N.I.; Shimanovskii, N.L.; Fedotcheva, T.A. A Pregnane Steroid as the Chiral Auxiliary in 1,3-Dipolar Azomethine Ylide’s Cycloaddition: Asymmetric Synthesis and Anticancer Activity of Novel Hybrid Compounds. ChemistrySelect 2020, 5, 11467–11470. [Google Scholar] [CrossRef]
- Fedotcheva, N.I.; Teplova, V.V.; Fedotcheva, T.A.; Rzheznikov, V.M.; Shimanovskii, N.L. Effect of progesterone and its syn-thetic analogues on the activity of mitochondrial permeability ransition pore in isolated rat liver mitochondria. Biochem. Pharmacol. 2009, 78, 1060–1068. [Google Scholar] [CrossRef]
- Korystov, Y.N.; Ermakova, N.V.; Kublik, L.N.; Levitman, M.K.; Shaposhnikova, V.V.; Mosin, V.A.; Drinyaev, V.A.; Kruglyak, E.B.; Novik, T.S.; Sterlina, T.S. Avermectins inhibit multidrug resistance of tumor cells. Eur. J. Pharmacol. 2004, 493, 57–64. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, P.; Sun, Y.-J.; Wu, Y.-J. Ivermectin reverses the drug resistance in cancer cells through EGFR/ERK/Akt/NF-κB pathway. J. Exp. Clin. Cancer Res. 2019, 38, 1–18. [Google Scholar] [CrossRef]
- Zhang, Y.; Luo, M.; Xu, W.; Yang, M.; Wang, B.; Gao, J.; Li, Y.; Tao, L. Avermectin Confers Its Cytotoxic Effects by Inducing DNA Damage and Mitochondria-Associated Apoptosis. J. Agric. Food Chem. 2016, 64, 6895–6902. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, K.; Cheng, L.; Zhu, H.; Xu, T. Progress in Understanding the Molecular Mechanisms Underlying the Antitumour Effects of Ivermectin. Drug Des. Dev. Ther. 2020, 14, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Malik, C.; Ghosh, S. Quinidine partially blocks mitochondrial voltage-dependent anion channel (VDAC). Eur. Biophys. J. 2020, 49, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Mironova, G.D.; Grigoriev, S.M.; Yuyu, S.; E Negoda, A.; Kolomytkin, O.V. ATP-dependent potassium channel from rat liver mitochondria: Inhibitory analysis, channel clusterization. Membr. Cell Biol. 1997, 10. [Google Scholar]
- Agarwal, A.; Rao, S. Effect of quinidine on kidney biochemistry and function in male sprague-dawley rats. Food Chem. Toxicol. 1995, 33, 203–207. [Google Scholar] [CrossRef]
- Almotrefi, A.A. Effects of class I antiarrhythmic drugs on mitochondrial ATPase activity in guinea pig heart preparations. Gen. Pharmacol. Vasc. Syst. 1993, 24, 233–237. [Google Scholar] [CrossRef]
- Schild, L.; Bukowska, A.; Gardemann, A.; Polczyk, P.; Keilhoff, G.; Täger, M.; Dudley, S.C.; Klein, H.U.; Goette, A.; Lendeckel, U. Rapid pacing of embryoid bodies impairs mitochondrial ATP synthesis by a calcium-dependent mechanism—A model of in vitro differentiated cardiomyocytes to study molecular effects of tachycardia. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2006, 1762, 608–615. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, S.; Sun, Q.; Liu, B. Anthelmintic drug ivermectin inhibits angiogenesis, growth and survival of glioblastoma through inducing mitochondrial dysfunction and oxidative stress. Biochem. Biophys. Res. Commun. 2016, 480, 415–421. [Google Scholar] [CrossRef]
- Dalzell, A.M.; Mistry, P.; Wright, J.; Williams, F.M.; Brown, C.D. Characterization of multidrug transporter-mediated efflux of avermectins in human and mouse neuroblastoma cell lines. Toxicol. Lett. 2015, 235, 189–198. [Google Scholar] [CrossRef]
- Vezmar, M.; Georges, E. Reversal of MRP-mediated doxorubicin resistance with quinoline-based drugs. Biochem. Pharmacol. 2000, 59, 1245–1252. [Google Scholar] [CrossRef]
- Kawazu, K.; Yamada, K.; Nakamura, M.; Ota, A. Characterization of cyclosporin A transport in cultured rabbit corneal epi-thelial cells: P-glycoprotein transport activity and binding to cyclophilin. Invest. Ophthalmol. Vis. Sci. 1999, 40, 1738–1744. [Google Scholar]
- Meier, M.; Blatter, X.; Seelig, A.; Seelig, J. Interaction of Verapamil with Lipid Membranes and P-Glycoprotein: Connecting Thermodynamics and Membrane Structure with Functional Activity. Biophys. J. 2006, 91, 2943–2955. [Google Scholar] [CrossRef] [PubMed]
- Pohl, E.E.; Krylov, A.V.; Block, M.; Pohl, P. Changes of the membrane potential profile induced by verapamil and propranolol. Biochim. Biophys. Acta (BBA) Biomembr. 1998, 1373, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Tsushima, R.G.; Kelly, J.E.; Wasserstrom, J.A. Subconductance activity induced by quinidine and quinidinium in purified cardiac sarcoplasmic reticulum calcium release channels. J. Pharmacol. Exp. Ther. 2002, 301, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Tang, M.; Bode, A.M.; Liao, W.; Cao, Y. ANTs and cancer: Emerging pathogenesis, mechanisms, and perspectives. Biochim. Biophys. Acta 2020, 1875, 188485. [Google Scholar] [CrossRef] [PubMed]
- Zanoli, J.C.C.; Maioli, M.A.; Medeiros, H.; Mingatto, F.E. Abamectin affects the bioenergetics of liver mitochondria: A potential mechanism of hepatotoxicity. Toxicol. Vitr. 2012, 26, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Deng, X.; Li, Y.; Hu, J.; Xie, L.; Shi, F.; Tang, M.; Bode, A.M.; Zhang, X.; Liao, W.; et al. Conformational change of adenine nucleotide translocase-1 mediates cisplatin resistance induced by EBV-LMP1. EMBO Mol. Med. 2021, 13, e14072. [Google Scholar] [CrossRef]
- Zoghbi, M.; Mok, L.; Swartz, D.J.; Singh, A.; Fendley, G.A.; Urbatsch, I.L.; Altenberg, G.A. Substrate-induced conformational changes in the nucleotide-binding domains of lipid bilayer–associated P-glycoprotein during ATP hydrolysis. J. Biol. Chem. 2017, 292, 20412–20424. [Google Scholar] [CrossRef]
- Higgins, C.F.; Linton, K.J. The ATP switch model for ABC transporters. Nat. Struct. Mol. Biol. 2004, 11, 918–926. [Google Scholar] [CrossRef]
- Szöllősi, D.; Rose-Sperling, D.; Hellmich, U.; Stockner, T. Comparison of mechanistic transport cycle models of ABC exporters. Biochim. et Biophys. Acta (BBA) Biomembr. 2018, 1860, 818–832. [Google Scholar] [CrossRef]
- Palmeira, A.; Sousa, M.E.; Vasconcelos, M.H.; Pinto, M. Three Decades of P-gp Inhibitors: Skimming Through Several Generations and Scaffolds. Curr. Med. Chem. 2012, 19, 1946–2025. [Google Scholar] [CrossRef]
- Dartier, J.; Lemaitre, E.; Chourpa, I.; Goupille, C.; Servais, S.; Chevalier, S.; Mahéo, K.; Dumas, J.-F. ATP-dependent activity and mitochondrial localization of drug efflux pumps in doxorubicin-resistant breast cancer cells. Biochim. et Biophys. Acta (BBA) Gen. Subj. 2017, 1861, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Hendricks, A.L.; Wang, Y.; Villones, R.L.E.; Lindkvist-Petersson, K.; Meloni, G.; Cowan, J.A.; Wang, K.; Gourdon, P. Structures of Atm1 provide insight into [2Fe-2S] cluster export from mitochondria. Nat. Commun. 2022, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.; Fendley, G.A.; Saxberg, A.D.; Zoghbi, M.E. Stimulation of the human mitochondrial transporter ABCB10 by zinc-mesoporphrin. PLoS ONE 2020, 15, e0238754. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Kumar, T.R.S.; Kartha, C.C. Mitochondrial membrane transporters and metabolic switch in heart failure. Hear. Fail. Rev. 2018, 24, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Giddings, E.L.; Champagne, D.P.; Wu, M.-H.; Laffin, J.M.; Thornton, T.M.; Valenca-Pereira, F.; Culp-Hill, R.; Fortner, K.A.; Romero, N.; East, J.; et al. Mitochondrial ATP fuels ABC transporter-mediated drug efflux in cancer chemoresistance. Nat. Commun. 2021, 12, 1–19. [Google Scholar] [CrossRef]
- Lee, A.; Lau, P.; Kwan, Y.; Kong, S. Mitochondrial Fuel Dependence on Glutamine Drives Chemo-Resistance in the Cancer Stem Cells of Hepatocellular Carcinoma. Int. J. Mol. Sci. 2021, 22, 3315. [Google Scholar] [CrossRef]
- Bokil, A.; Sancho, P. Mitochondrial determinants of chemoresistance. Cancer Drug Resist 2019. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fedotcheva, T.; Shimanovsky, N.; Fedotcheva, N. Involvement of Multidrug Resistance Modulators in the Regulation of the Mitochondrial Permeability Transition Pore. Membranes 2022, 12, 890. https://doi.org/10.3390/membranes12090890
Fedotcheva T, Shimanovsky N, Fedotcheva N. Involvement of Multidrug Resistance Modulators in the Regulation of the Mitochondrial Permeability Transition Pore. Membranes. 2022; 12(9):890. https://doi.org/10.3390/membranes12090890
Chicago/Turabian StyleFedotcheva, Tatiana, Nikolai Shimanovsky, and Nadezhda Fedotcheva. 2022. "Involvement of Multidrug Resistance Modulators in the Regulation of the Mitochondrial Permeability Transition Pore" Membranes 12, no. 9: 890. https://doi.org/10.3390/membranes12090890