

Unravelling Contributions of Astrocytic Connexin 43 to the Functional Activity of Brain Neuron–Glial Networks under Hypoxic State In Vitro
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Isolation and Characterization of the Primary Cerebral Cortex Cell Cultures
2.3. Experimental Design
2.4. RNA Extraction and RT-qPCR
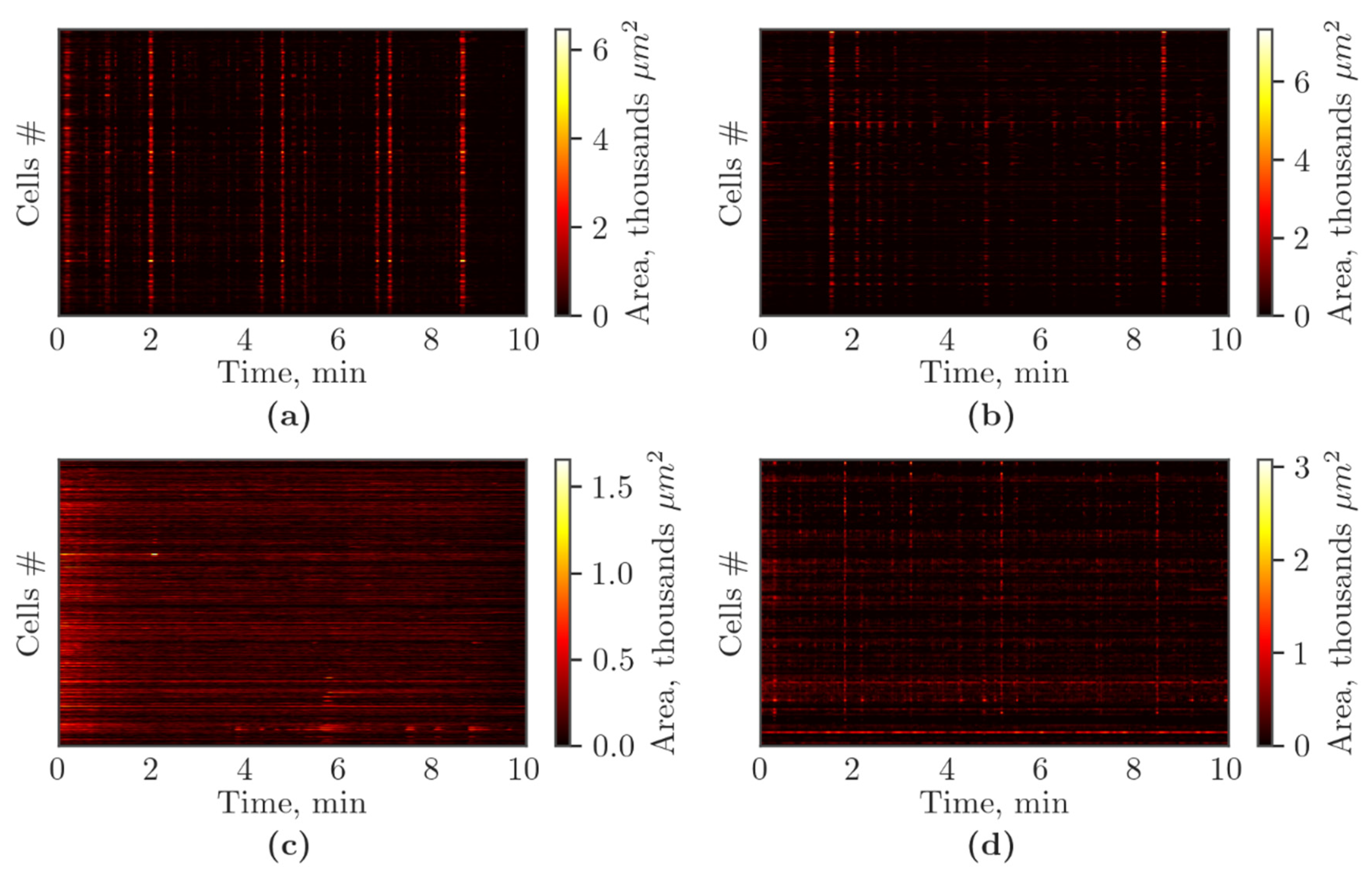
2.5. Functional Calcium Imaging
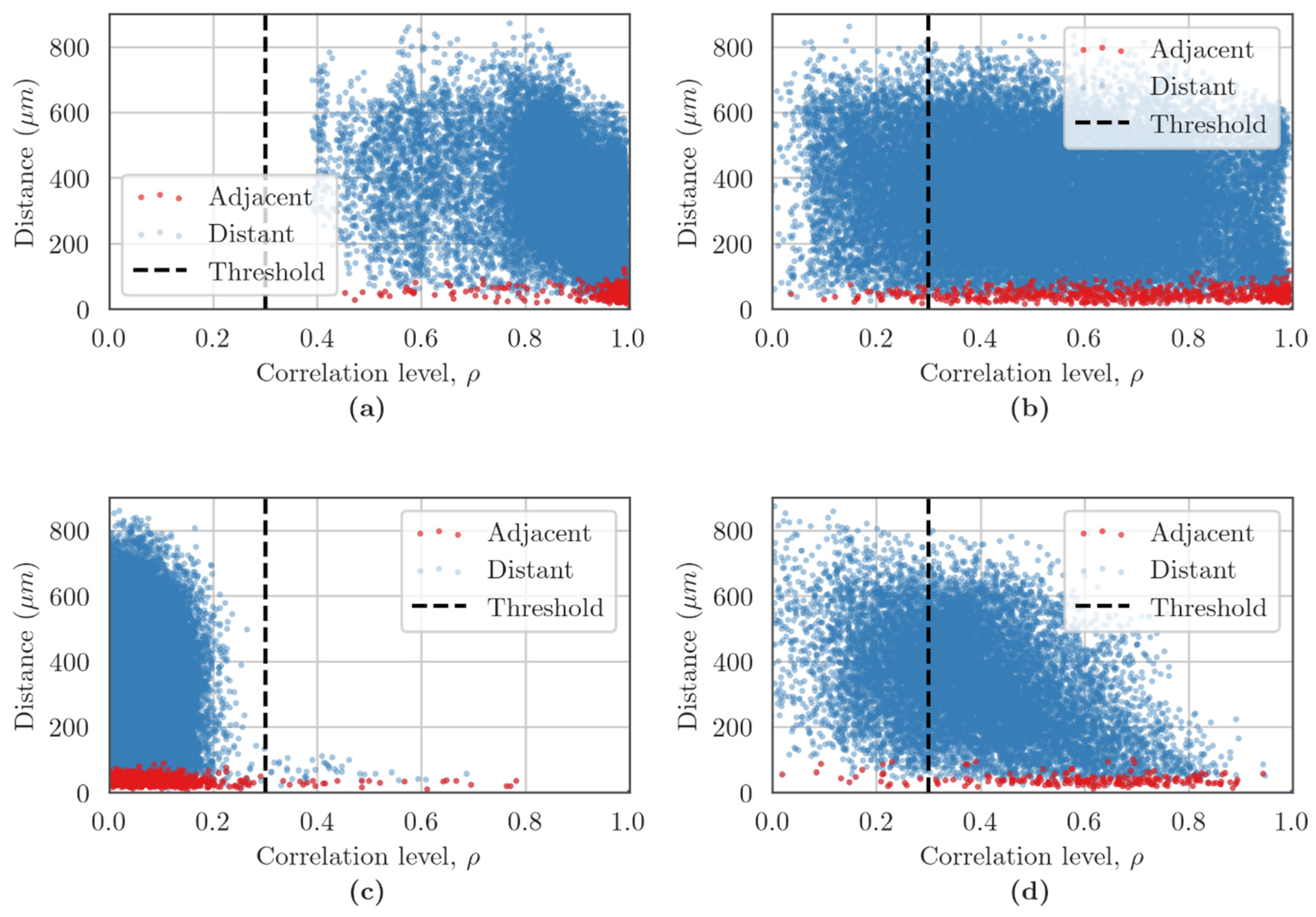
2.6. Network Characteristic Assessments in the Cerebral Cortex Cell Cultures
2.7. Statistical Analysis
3. Results
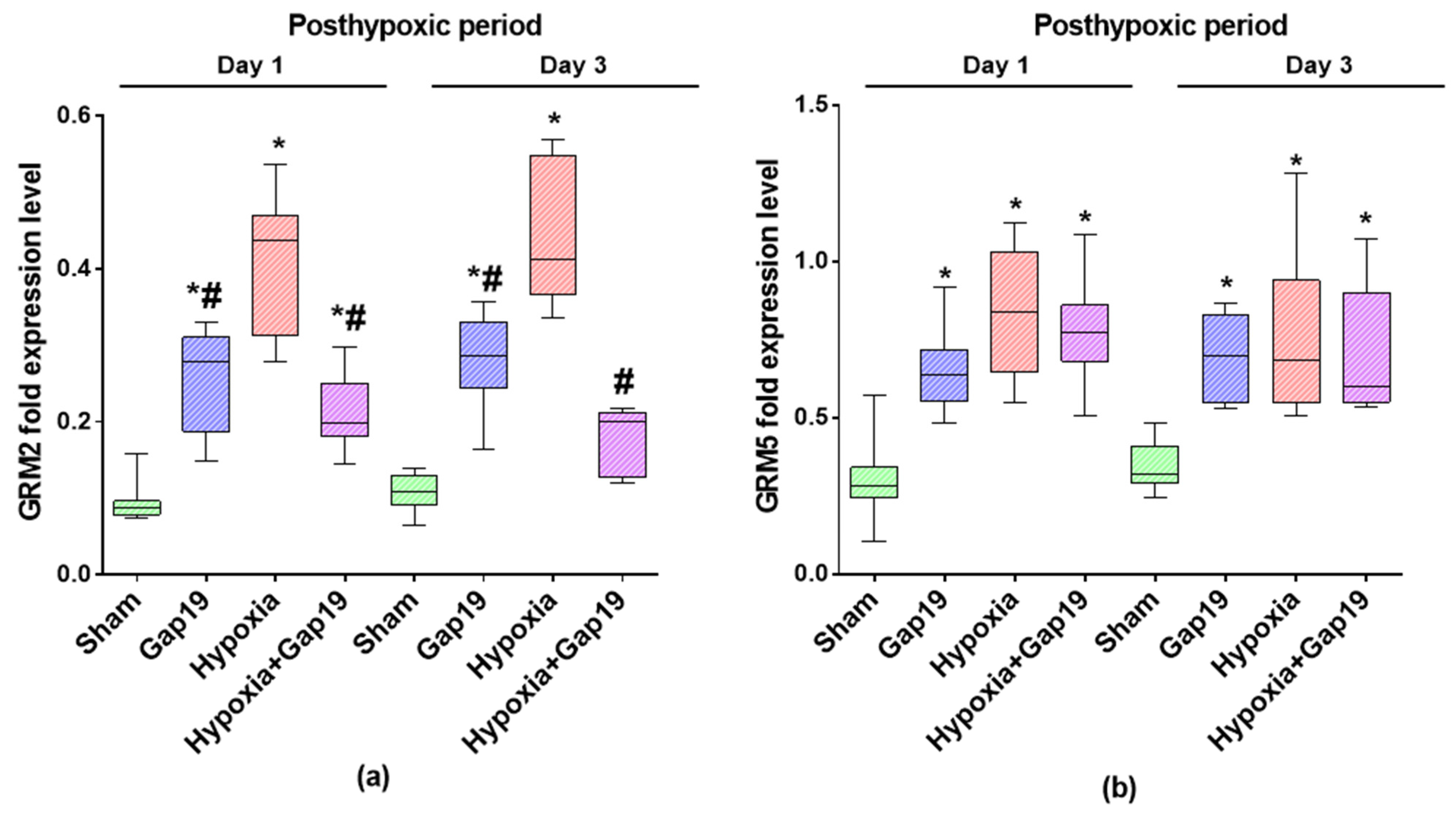
3.1. Assesment of the Expression Levels of Metabotropic Glutamate Receptors mGluR2 and mGluR5 in Primary Cerebral Cortex Cells under Blockade of Cx43 and Hypoxic Damage
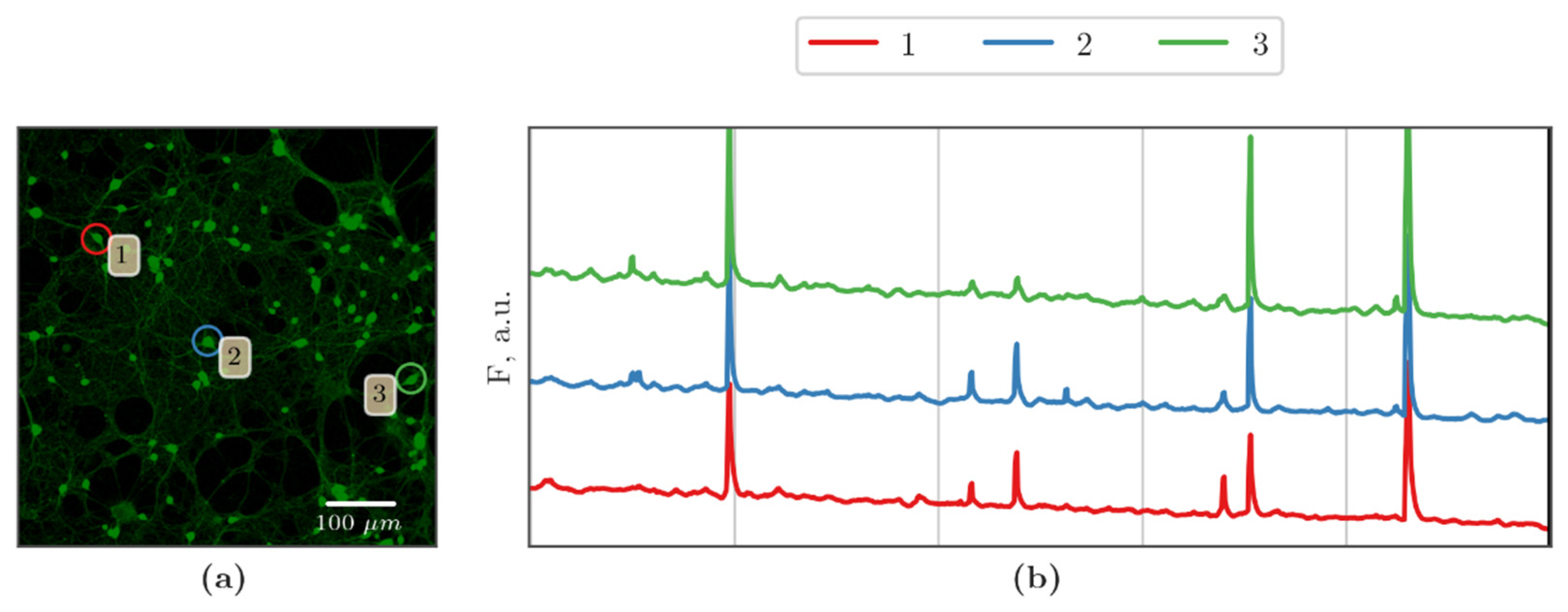
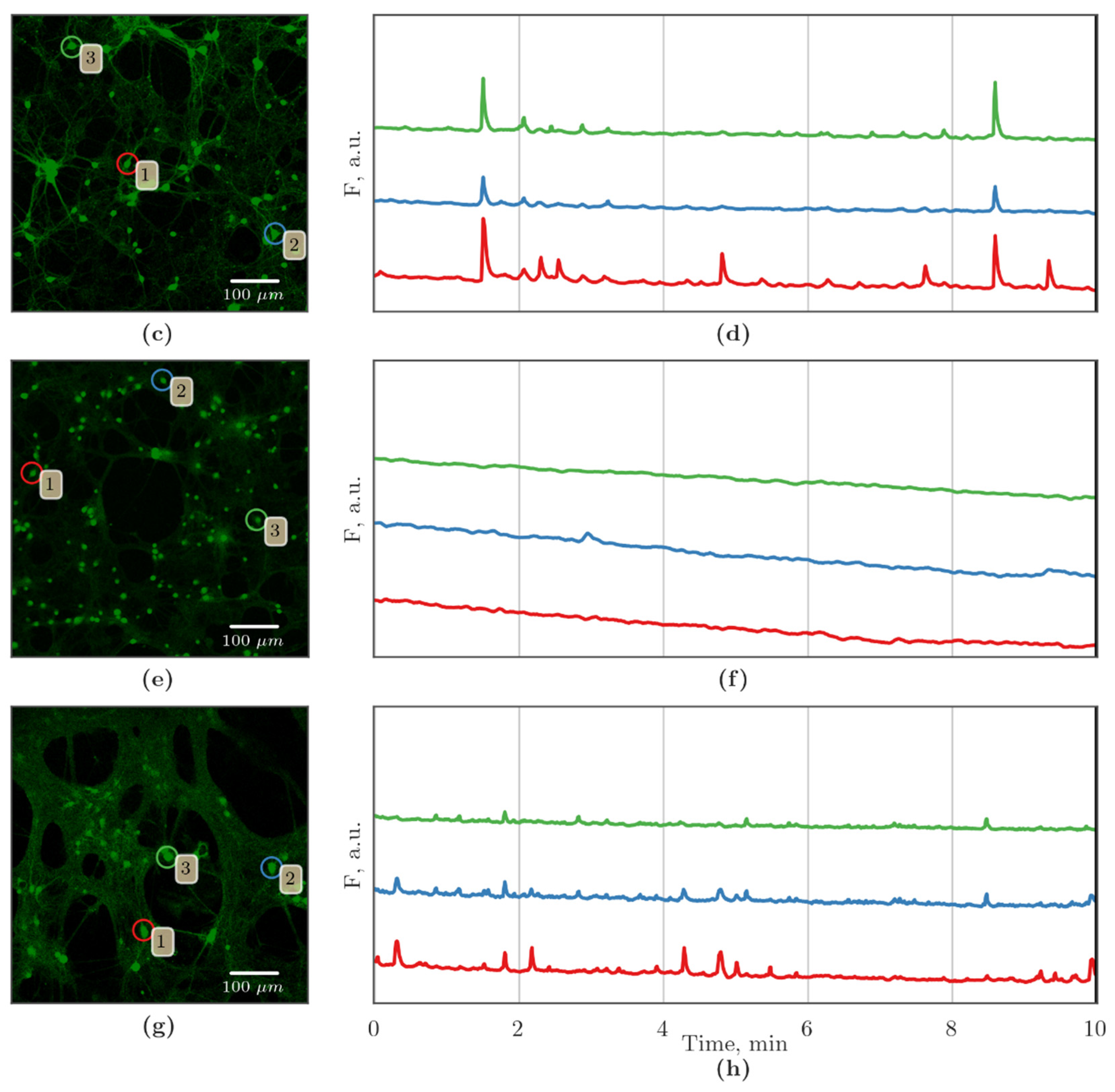
3.2. Features of Spontaneous Calcium Activity of Primary Cerebral Cortex Cell Cultures under Cx43 Blockade and Hypoxic Damage
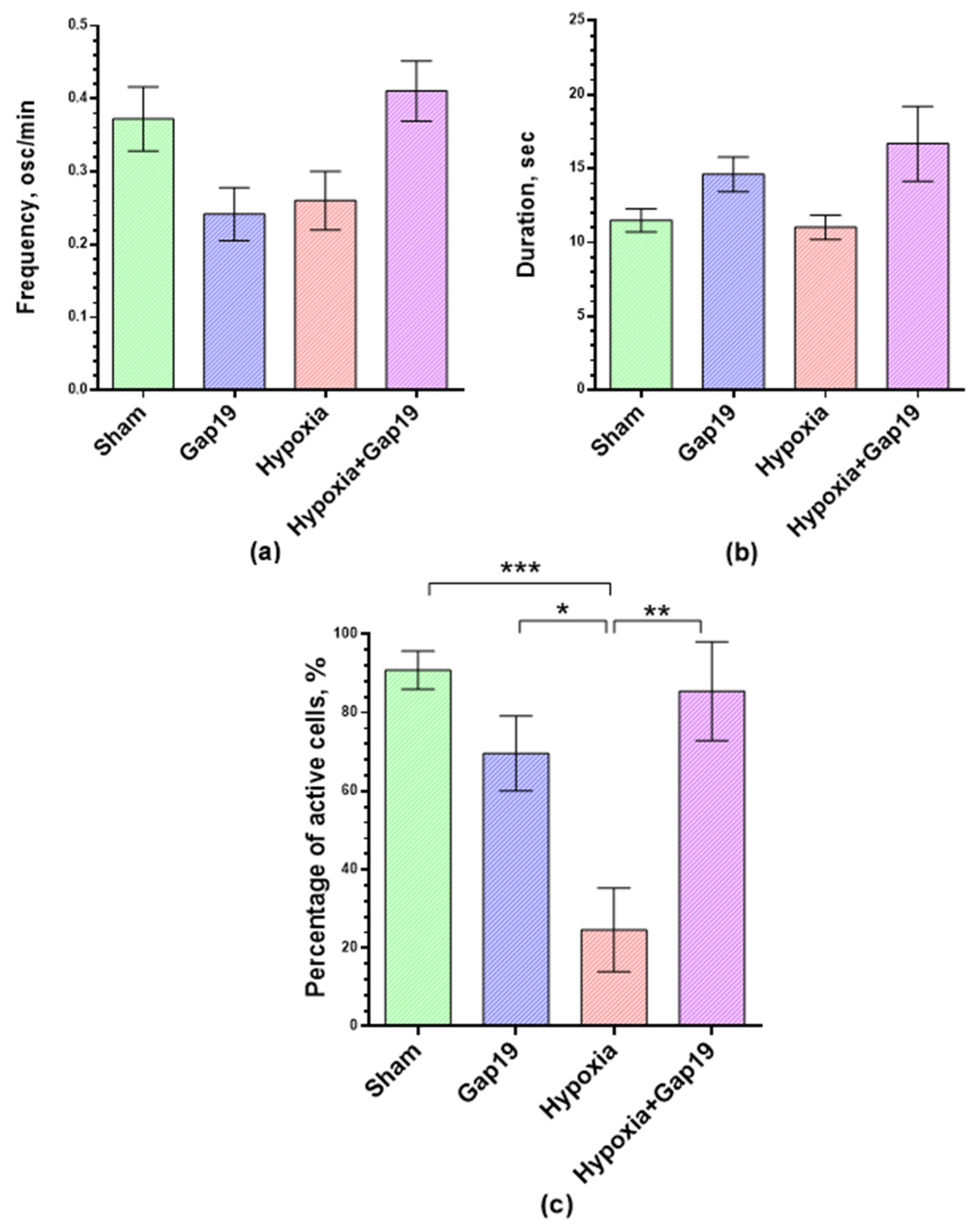
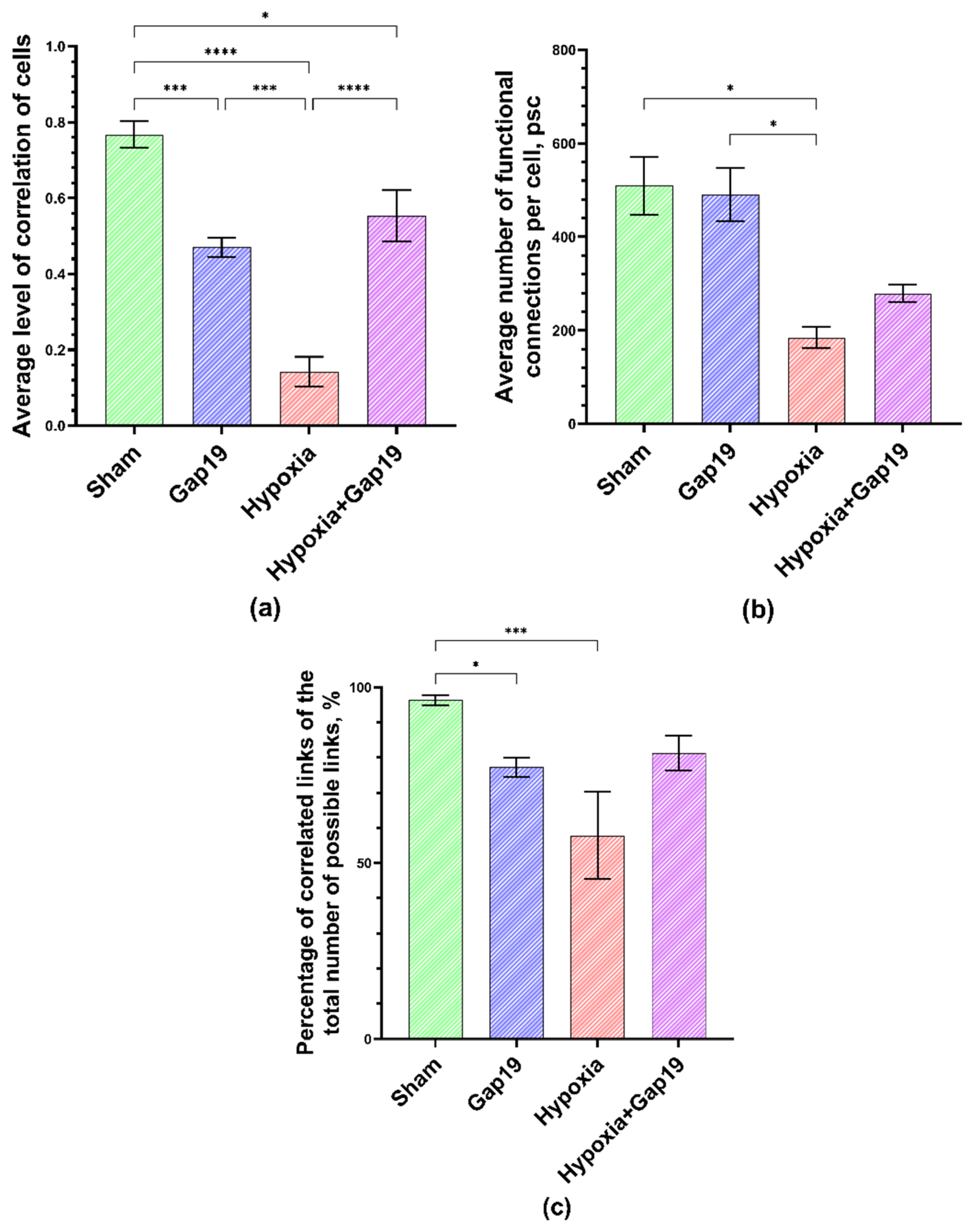
3.3. Features of Neuron–Glial Network Interactions in Primary Cerebral Cortex under Cx43 Blockade Hypoxic Damage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Radak, D.; Katsiki, N.; Resanovic, I.; Jovanovic, A.; Sudar-Milovanovic, E.; Zafirovic, S.; Mousad, S.A.; Isenovic, E.R. Apoptosis and Acute Brain Ischemia in Ischemic Stroke. Curr. Vasc. Pharmacol. 2017, 15, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Pathak, D.; Shields, L.Y.; Mendelsohn, B.A.; Haddad, D.; Lin, W.; Gerencser, A.A.; Kim, H.; Brand, M.D.; Edwards, R.H.; Nakamura, K. The Role of Mitochondrially Derived ATP in Synaptic Vesicle Recycling. J. Biol. Chem. 2015, 290, 22325–22336. [Google Scholar] [CrossRef]
- Rangaraju, V.; Calloway, N.; Ryan, T.A. Activity-Driven Local ATP Synthesis Is Required for Synaptic Function. Cell 2014, 156, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Hyder, F.; Rothman, D.L.; Bennett, M.R. Cortical Energy Demands of Signaling and Nonsignaling Components in Brain Are Conserved across Mammalian Species and Activity Levels. Proc. Natl. Acad. Sci. USA 2013, 110, 3549–3554. [Google Scholar] [CrossRef]
- Harris, J.J.; Jolivet, R.; Attwell, D. Synaptic Energy Use and Supply. Neuron 2012, 75, 762–777. [Google Scholar] [CrossRef] [PubMed]
- Mitroshina, E.V.; Savyuk, M.O.; Ponimaskin, E.; Vedunova, M.V. Hypoxia-Inducible Factor (HIF) in Ischemic Stroke and Neurodegenerative Disease. Front. Cell Dev. Biol. 2021, 9, 703084. [Google Scholar] [CrossRef]
- Domènech, M.; Hernández, A.; Plaja, A.; Martínez-Balibrea, E.; Balañà, C. Hypoxia: The Cornerstone of Glioblastoma. Int. J. Mol. Sci. 2021, 22, 12608. [Google Scholar] [CrossRef]
- Lestón Pinilla, L.; Ugun-Klusek, A.; Rutella, S.; de Girolamo, L.A. Hypoxia Signaling in Parkinson’s Disease: There Is Use in Asking “What HIF?”. Biology 2021, 10, 723. [Google Scholar] [CrossRef]
- Choi, D.W. Excitotoxicity: Still Hammering the Ischemic Brain in 2020. Front. Neurosci. 2020, 14, 579953. [Google Scholar] [CrossRef]
- Mukandala, G.; Tynan, R.; Lanigan, S.; O’Connor, J.J. The Effects of Hypoxia and Inflammation on Synaptic Signaling in the CNS. Brain Sci. 2016, 6, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirayama, Y.; Koizumi, S. Astrocytes and Ischemic Tolerance. Neurosci. Res. 2018, 126, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Le Feber, J.; Erkamp, N.; van Putten, M.J.A.M.; Hofmeijer, J. Loss and Recovery of Functional Connectivity in Cultured Cortical Networks Exposed to Hypoxia. J. Neurophysiol. 2017, 118, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Peña-Ortega, F. Neural Network Reconfigurations: Changes of the Respiratory Network by Hypoxia as an Example. In The Plastic Brain; Springer: Cham, Switzerland, 2017; pp. 217–237. [Google Scholar]
- Charvériat, M.; Naus, C.C.; Leybaert, L.; Sáez, J.C.; Giaume, C. Connexin-Dependent Neuroglial Networking as a New Therapeutic Target. Front. Cell Neurosci. 2017, 11, 174. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol. Rev. 2018, 98, 239–389. [Google Scholar] [CrossRef]
- Retamal, M.A.; Reyes, E.P.; García, I.E.; Pinto, B.; Martínez, A.D.; González, C. Diseases Associated with Leaky Hemichannels. Front. Cell Neurosci. 2015, 9, 267. [Google Scholar] [CrossRef]
- de Bock, M.; Leybaert, L.; Giaume, C. Connexin Channels at the Glio-Vascular Interface: Gatekeepers of the Brain. Neurochem. Res. 2017, 42, 2519–2536. [Google Scholar] [CrossRef]
- Vasile, F.; Dossi, E.; Rouach, N. Human Astrocytes: Structure and Functions in the Healthy Brain. Brain Struct. Funct. 2017, 222, 2017–2029. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Parpura, V.; Rodriguez-Arellano, J.J.; Zorec, R. Astroglia in Alzheimer’s Disease. Adv. Exp. Med. Biol. 2019, 1175, 273–324. [Google Scholar]
- Lapato, A.S.; Tiwari-Woodruff, S.K. Connexins and Pannexins: At the Junction of Neuro-Glial Homeostasis & Disease. J. Neurosci. Res. 2018, 96, 31–44. [Google Scholar] [CrossRef]
- Liang, Z.; Wang, X.; Hao, Y.; Qiu, L.; Lou, Y.; Zhang, Y.; Ma, D.; Feng, J. The Multifaceted Role of Astrocyte Connexin 43 in Ischemic Stroke through Forming Hemichannels and Gap Junctions. Front. Neurol. 2020, 11, 703. [Google Scholar] [CrossRef]
- Freitas-Andrade, M.; She, J.; Bechberger, J.; Naus, C.C.; Sin, W.C. Acute Connexin43 Temporal and Spatial Expression in Response to Ischemic Stroke. J. Cell Commun. Signal 2018, 12, 193–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ma, A.; Xi, J.; Wang, Y.; Zhao, B. Connexin 43 and Its Hemichannels Mediate Hypoxia–Ischemia-Induced Cell Death in Neonatal Rats. Child Neurol. Open 2014, 1, 2329048X1454495. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Yang, L.; Chen, J.; Chen, Y.; Zhang, L.; Wang, L.; Li, X.; Li, Y.; Yu, H. Inhibition of Connexin43 Hemichannels with Gap19 Protects Cerebral Ischemia/Reperfusion Injury via the JAK2/STAT3 Pathway in Mice. Brain Res. Bull. 2019, 146, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Feng, L.; Cheng, Y.; Xin, M.; You, J.; Yin, X.; Hao, Y.; Cui, L.; Feng, J. Astrocytic Gap Junction Inhibition by Carbenoxolone Enhances the Protective Effects of Ischemic Preconditioning Following Cerebral Ischemia. J. Neuroinflamm. 2018, 15, 198. [Google Scholar] [CrossRef]
- Kozoriz, M.G.; Bechberger, J.F.; Bechberger, G.R.; Suen, M.W.H.; Moreno, A.P.; Maass, K.; Willecke, K.; Naus, C.C. The Connexin43 C-Terminal Region Mediates Neuroprotection during Stroke. J. Neuropathol. Exp. Neurol. 2010, 69, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Orellana, J.A.; Hernández, D.E.; Ezan, P.; Velarde, V.; Bennett, M.V.L.; Giaume, C.; Sáez, J.C. Hypoxia in High Glucose Followed by Reoxygenation in Normal Glucose Reduces the Viability of Cortical Astrocytes through Increased Permeability of Connexin 43 Hemichannels. Glia 2010, 58, 329–343. [Google Scholar] [CrossRef]
- Froger, N.; Orellana, J.A.; Calvo, C.-F.; Amigou, E.; Kozoriz, M.G.; Naus, C.C.; Sáez, J.C.; Giaume, C. Inhibition of Cytokine-Induced Connexin43 Hemichannel Activity in Astrocytes Is Neuroprotective. Mol. Cell Neurosci. 2010, 45, 37–46. [Google Scholar] [CrossRef]
- Retamal, M.A.; Schalper, K.A.; Shoji, K.F.; Orellana, J.A.; Bennett, M.V.L.; Sáez, J.C. Possible Involvement of Different Connexin43 Domains in Plasma Membrane Permeabilization Induced by Ischemia-Reperfusion. J. Membr. Biol. 2007, 218, 49–63. [Google Scholar] [CrossRef]
- Nakase, T.; Söhl, G.; Theis, M.; Willecke, K.; Naus, C.C.G. Increased Apoptosis and Inflammation after Focal Brain Ischemia in Mice Lacking Connexin43 in Astrocytes. Am. J. Pathol. 2004, 164, 2067–2075. [Google Scholar] [CrossRef]
- Vedunova, M.; Sakharnova, T.; Mitroshina, E.; Perminova, M.; Pimashkin, A.; Zakharov, Y.; Dityatev, A.; Mukhina, I. Seizure-like Activity in Hyaluronidase-Treated Dissociated Hippocampal Cultures. Front. Cell Neurosci. 2013, 7, 149. [Google Scholar] [CrossRef]
- Loginova, M.; Mishchenko, T.; Savyuk, M.; Guseva, S.; Gavrish, M.; Krivonosov, M.; Ivanchenko, M.; Fedotova, J.; Vedunova, M. Double-Edged Sword of Vitamin D3 Effects on Primary Neuronal Cultures in Hypoxic States. Int. J. Mol. Sci. 2021, 22, 5417. [Google Scholar] [CrossRef] [PubMed]
- Mitroshina, E.V.; Loginova, M.M.; Savyuk, M.O.; Krivonosov, M.I.; Mishchenko, T.A.; Tarabykin, V.S.; Ivanchenko, M.V.; Vedunova, M.V. Neuroprotective Effect of Kinase Inhibition in Ischemic Factor Modeling In Vitro. Int. J. Mol. Sci. 2021, 22, 1885. [Google Scholar] [CrossRef] [PubMed]
- Mishchenko, T.A.; Mitroshina, E.V.; Usenko, A.V.; Voronova, N.V.; Astrakhanova, T.A.; Shirokova, O.M.; Kastalskiy, I.A.; Vedunova, M.V. Features of Neural Network Formation and Their Functions in Primary Hippocampal Cultures in the Context of Chronic TrkB Receptor System Influence. Front. Physiol. 2019, 9, 1925. [Google Scholar] [CrossRef] [PubMed]
- Shirokova, O.M.; Mukhina, I.V.; Frumkina, L.E.; Vedunova, M.V.; Mitroshina, E.V.; Zakharov, Y.N.; Khaspekov, L.G. Morphofunctional Patterns of Neuronal Network Developing in Dissociated Hippocampal Cell Cultures. Sovrem. Tehnol. Med. 2013, 5, 6–13. [Google Scholar]
- Vedunova, M.V.; Mishchenko, T.A.; Mitroshina, E.V.; Mukhina, I.V. TrkB-Mediated Neuroprotective and Antihypoxic Properties of Brain-Derived Neurotrophic Factor. Oxid. Med. Cell Longev. 2015, 2015, 453901. [Google Scholar] [CrossRef]
- Savyuk, M.O.; Turubanova, V.D.; Mishchenko, T.A.; Lermontova, S.A.; Klapshina, L.G.; Krysko, D.V.; Vedunova, M.V. Unraveling of Functional Activity of Primary Hippocampal Neuron-Glial Networks in Photodynamic Therapy Based on Tetracyanotetra(Aryl)Porphyrazines. Cells 2022, 11, 1212. [Google Scholar] [CrossRef]
- Savyuk, M.; Krivonosov, M.; Mishchenko, T.; Gazaryan, I.; Ivanchenko, M.; Khristichenko, A.; Poloznikov, A.; Hushpulian, D.; Nikulin, S.; Tonevitsky, E.; et al. Neuroprotective Effect of HIF Prolyl Hydroxylase Inhibition in an In Vitro Hypoxia Model. Antioxidants 2020, 9, 662. [Google Scholar] [CrossRef]
- Kustikova, V.; Krivonosov, M.; Pimashkin, A.; Denisov, P.; Zaikin, A.; Ivanchenko, M.; Meyerov, I.; Semyanov, A. CalciumCV: Computer Vision Software for Calcium Signaling in Astrocytes. In Proceedings of the International Conference on Analysis of Images, Social Networks and Texts, Moscow, Russia, 5–7 July 2018; Lecture Notes in Computer Science (Including Subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics). Springer: Cham, Switzerland, 2018; Volume 11179. [Google Scholar]
- Mishchenko, T.A.; Klimenko, M.O.; Kuznetsova, A.I.; Yarkov, R.S.; Savelyev, A.G.; Sochilina, A.V.; Mariyanats, A.O.; Popov, V.K.; Khaydukov, E.V.; Zvyagin, A.V.; et al. 3D-Printed Hyaluronic Acid Hydrogel Scaffolds Impregnated with Neurotrophic Factors (BDNF, GDNF) for Post-Traumatic Brain Tissue Reconstruction. Front. Bioeng. Biotechnol. 2022, 10, 1344. [Google Scholar] [CrossRef]
- Wang, X.; Hou, Y.; Li, Q.; Li, X.; Wang, W.; Ai, X.; Kuang, T.; Chen, X.; Zhang, Y.; Zhang, J.; et al. Rhodiola Crenulata Attenuates Apoptosis and Mitochondrial Energy Metabolism Disorder in Rats with Hypobaric Hypoxia-Induced Brain Injury by Regulating the HIF-1α/MicroRNA 210/ISCU1/2(COX10) Signaling Pathway. J. Ethnopharmacol. 2019, 241, 111801. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, Q.; Wang, J.; Liu, F.; Mi, M.; Xu, H.; Chen, F.; Zeng, K. Taurine Protects Transformed Rat Retinal Ganglion Cells from Hypoxia-Induced Apoptosis by Preventing Mitochondrial Dysfunction. Brain Res. 2009, 1279, 131–138. [Google Scholar] [CrossRef]
- Zhuravin, I.A.; Dubrovskaya, N.M.; Vasilev, D.S.; Postnikova, T.Y.; Zaitsev, A.V. Prenatal Hypoxia Produces Memory Deficits Associated with Impairment of Long-Term Synaptic Plasticity in Young Rats. Neurobiol. Learn Mem. 2019, 164, 107066. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Mehmet, H.; Penrice, J.; Cooper, C.; Cady, E.; Wyatt, J.S.; Reynolds, E.O.R.; Edwards, A.D.; Squier, M.V. Apoptosis and Necrosis in the Newborn Piglet Brain Following Transient Cerebral Hypoxia-Ischaemia. Neuropathol. Appl. Neurobiol. 1997, 23, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Yuste, R. From the Neuron Doctrine to Neural Networks. Nat. Rev. Neurosci. 2015, 16, 487–497. [Google Scholar] [CrossRef]
- Mitroshina, E.V.; Loginova, M.M.; Yarkov, R.S.; Urazov, M.D.; Novozhilova, M.O.; Krivonosov, M.I.; Ivanchenko, M.V.; Vedunova, M.V. Inhibition of Neuronal Necroptosis Mediated by RIPK1 Provides Neuroprotective Effects on Hypoxia and Ischemia In Vitro and In Vivo. Int. J. Mol. Sci. 2022, 23, 735. [Google Scholar] [CrossRef]
- Mitroshina, E.; Mishchenko, T.; Usenko, A.; Epifanova, E.; Yarkov, R.; Gavrish, M.; Babaev, A.; Vedunova, M. AAV-Syn-BDNF-EGFP Virus Construct Exerts Neuroprotective Action on the Hippocampal Neural Network during Hypoxia In Vitro. Int. J. Mol. Sci. 2018, 19, 2295. [Google Scholar] [CrossRef]
- Volterra, A.; Meldolesi, J. Astrocytes, from Brain Glue to Communication Elements: The Revolution Continues. Nat. Rev. Neurosci. 2005, 6, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Kol, A.; Adamsky, A.; Groysman, M.; Kreisel, T.; London, M.; Goshen, I. Astrocytes Contribute to Remote Memory Formation by Modulating Hippocampal-Cortical Communication during Learning. Nat. Neurosci. 2020, 23, 1229–1239. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Nedergaard, M. Astroglial Cradle in the Life of the Synapse. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130595. [Google Scholar] [CrossRef]
- Svenningsen, Å.F.; Shan, W.-S.; Colman, D.R.; Pedraza, L. Rapid Method for Culturing Embryonic Neuron-Glial Cell Cocultures. J. Neurosci. Res. 2003, 72, 565–573. [Google Scholar] [CrossRef]
- Sahores, M.; Mendoza-Naranjo, A. Gap Junctions as Therapeutic Targets in Brain Injury Following Hypoxia-Ischemia. Recent Pat. CNS Drug Discov. 2008, 3, 209–215. [Google Scholar] [CrossRef]
- Seifert, G.; Schilling, K.; Steinhäuser, C. Astrocyte Dysfunction in Neurological Disorders: A Molecular Perspective. Nat. Rev. Neurosci. 2006, 7, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Niswender, C.M.; Conn, P.J. Metabotropic Glutamate Receptors: Physiology, Pharmacology, and Disease. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 295–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bocchio, M.; Lukacs, I.P.; Stacey, R.; Plaha, P.; Apostolopoulos, V.; Livermore, L.; Sen, A.; Ansorge, O.; Gillies, M.J.; Somogyi, P.; et al. Group II Metabotropic Glutamate Receptors Mediate Presynaptic Inhibition of Excitatory Transmission in Pyramidal Neurons of the Human Cerebral Cortex. Front. Cell Neurosci. 2018, 12, 508. [Google Scholar] [CrossRef]
- Coutinho, V.; Knöpfel, T. Metabotropic Glutamate Receptors: Electrical and Chemical Signaling Properties. Neuroscientist 2002, 8, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Purgert, C.A.; Izumi, Y.; Jong, Y.-J.I.; Kumar, V.; Zorumski, C.F.; O’Malley, K.L. Intracellular MGluR5 Can Mediate Synaptic Plasticity in the Hippocampus. J. Neurosci. 2014, 34, 4589–4598. [Google Scholar] [CrossRef] [PubMed]
- Piers, T.M.; Kim, D.H.; Kim, B.C.; Regan, P.; Whitcomb, D.J.; Cho, K. Translational Concepts of MGluR5 in Synaptic Diseases of the Brain. Front. Pharmacol. 2012, 3, 199. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Wang, Y.; Qin, Z. Molecular Mechanisms of Excitotoxicity and Their Relevance to Pathogenesis of Neurodegenerative Diseases. Acta Pharmacol. Sin. 2009, 30, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Drouin-Ouellet, J.; Brownell, A.-L.; Saint-Pierre, M.; Fasano, C.; Emond, V.; Trudeau, L.-E.; Lévesque, D.; Cicchetti, F. Neuroinflammation Is Associated with Changes in Glial MGluR5 Expression and the Development of Neonatal Excitotoxic Lesions. Glia 2011, 59, 188–199. [Google Scholar] [CrossRef]
- Crupi, R.; Impellizzeri, D.; Cuzzocrea, S. Role of Metabotropic Glutamate Receptors in Neurological Disorders. Front. Mol. Neurosci. 2019, 12, 20. [Google Scholar] [CrossRef]
- Ribeiro, F.M.; Vieira, L.B.; Pires, R.G.W.; Olmo, R.P.; Ferguson, S.S.G. Metabotropic Glutamate Receptors and Neurodegenerative Diseases. Pharmacol. Res. 2017, 115, 179–191. [Google Scholar] [CrossRef]
- Panatier, A.; Robitaille, R. Astrocytic MGluR5 and the Tripartite Synapse. Neuroscience 2016, 323, 29–34. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishchenko, T.A.; Yarkov, R.S.; Saviuk, M.O.; Krivonosov, M.I.; Perenkov, A.D.; Gudkov, S.V.; Vedunova, M.V. Unravelling Contributions of Astrocytic Connexin 43 to the Functional Activity of Brain Neuron–Glial Networks under Hypoxic State In Vitro. Membranes 2022, 12, 948. https://doi.org/10.3390/membranes12100948
Mishchenko TA, Yarkov RS, Saviuk MO, Krivonosov MI, Perenkov AD, Gudkov SV, Vedunova MV. Unravelling Contributions of Astrocytic Connexin 43 to the Functional Activity of Brain Neuron–Glial Networks under Hypoxic State In Vitro. Membranes. 2022; 12(10):948. https://doi.org/10.3390/membranes12100948
Chicago/Turabian StyleMishchenko, Tatiana A., Roman S. Yarkov, Mariia O. Saviuk, Mikhail I. Krivonosov, Alexey D. Perenkov, Sergey V. Gudkov, and Maria V. Vedunova. 2022. "Unravelling Contributions of Astrocytic Connexin 43 to the Functional Activity of Brain Neuron–Glial Networks under Hypoxic State In Vitro" Membranes 12, no. 10: 948. https://doi.org/10.3390/membranes12100948