
Lipid Membrane Mimetics in Functional and Structural Studies of Integral Membrane Proteins

Abstract
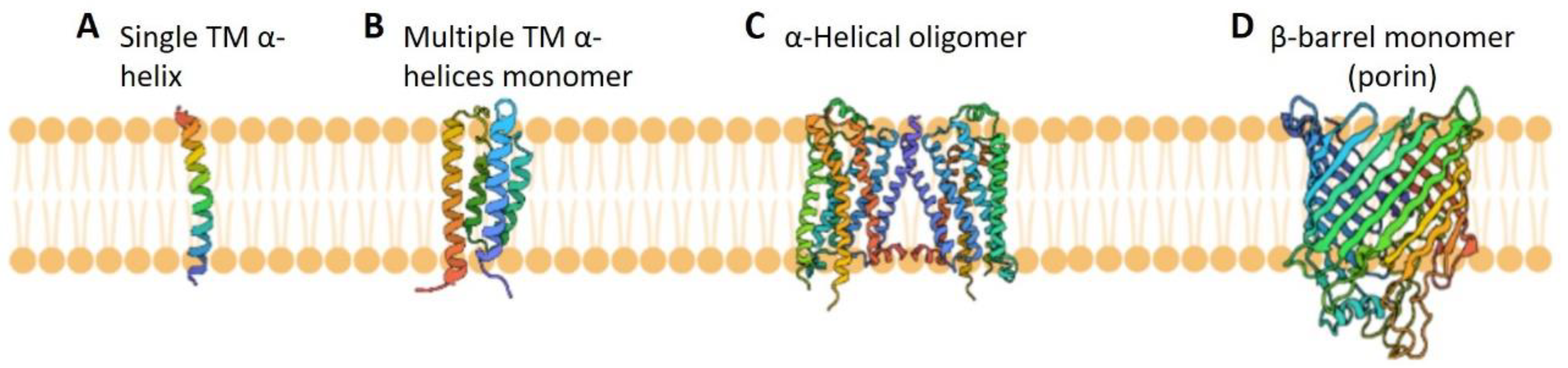
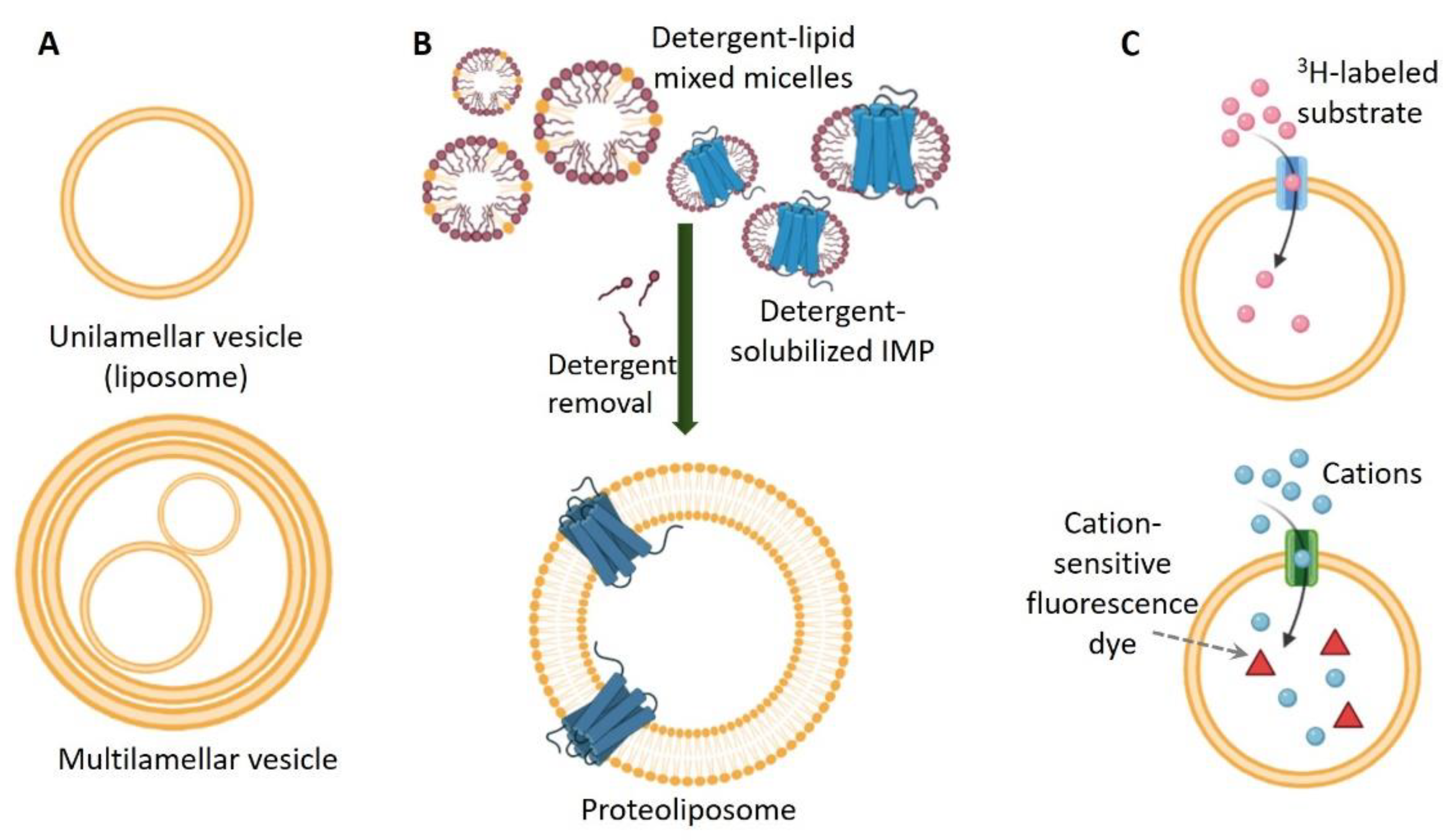
:1. Introduction
2. An Overview of the Most Widely Used Lipid Membrane Mimetics and Their Applications in Functional and Structural Studies of Integral Membrane Proteins
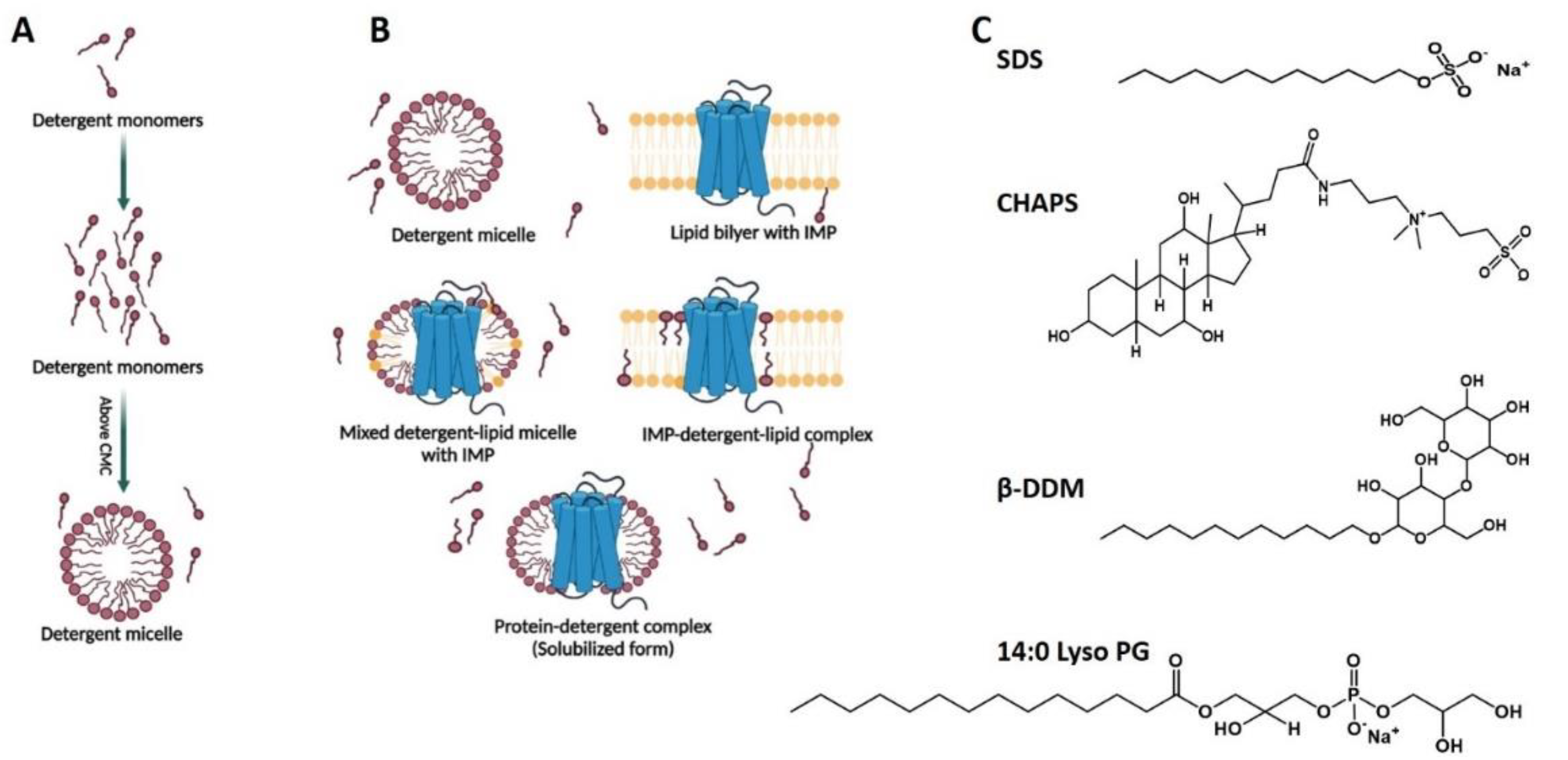
2.1. Detergents and Detergent Micelles in Studies of Integral Membrane Proteins
2.1.1. General Properties of Detergents and Detergent Micelles
2.1.2. Detergent Applications in Integral Membrane Proteins Solubilization, Purification, and Stabilization
2.1.3. Applications of Detergents in Functional Studies of Integral Membrane Proteins
2.1.4. Detergent Applications in Studies of Integral Membrane Proteins Using Biophysical and Structural Biology Methods
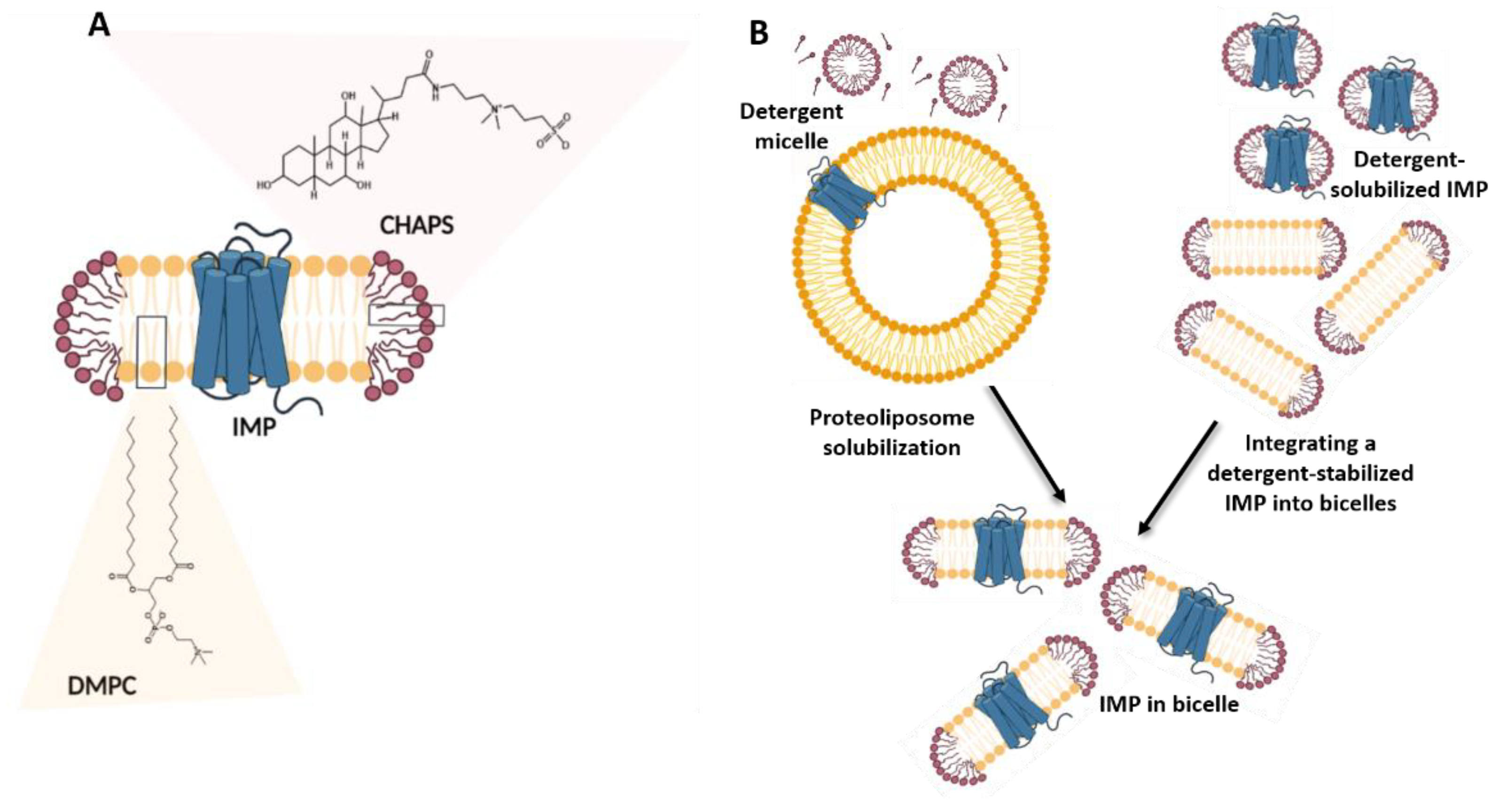
2.2. Bicelles in Studies of Integral Membrane Proteins
2.2.1. General Properties of Bicelles
2.2.2. Applications of Bicelles in Solubilizing and Stabilizing Integral Membrane Proteins
2.2.3. Applications of Bicelles in Studies on Integral Membrane Proteins Using Biophysical and Structural Biology Methods
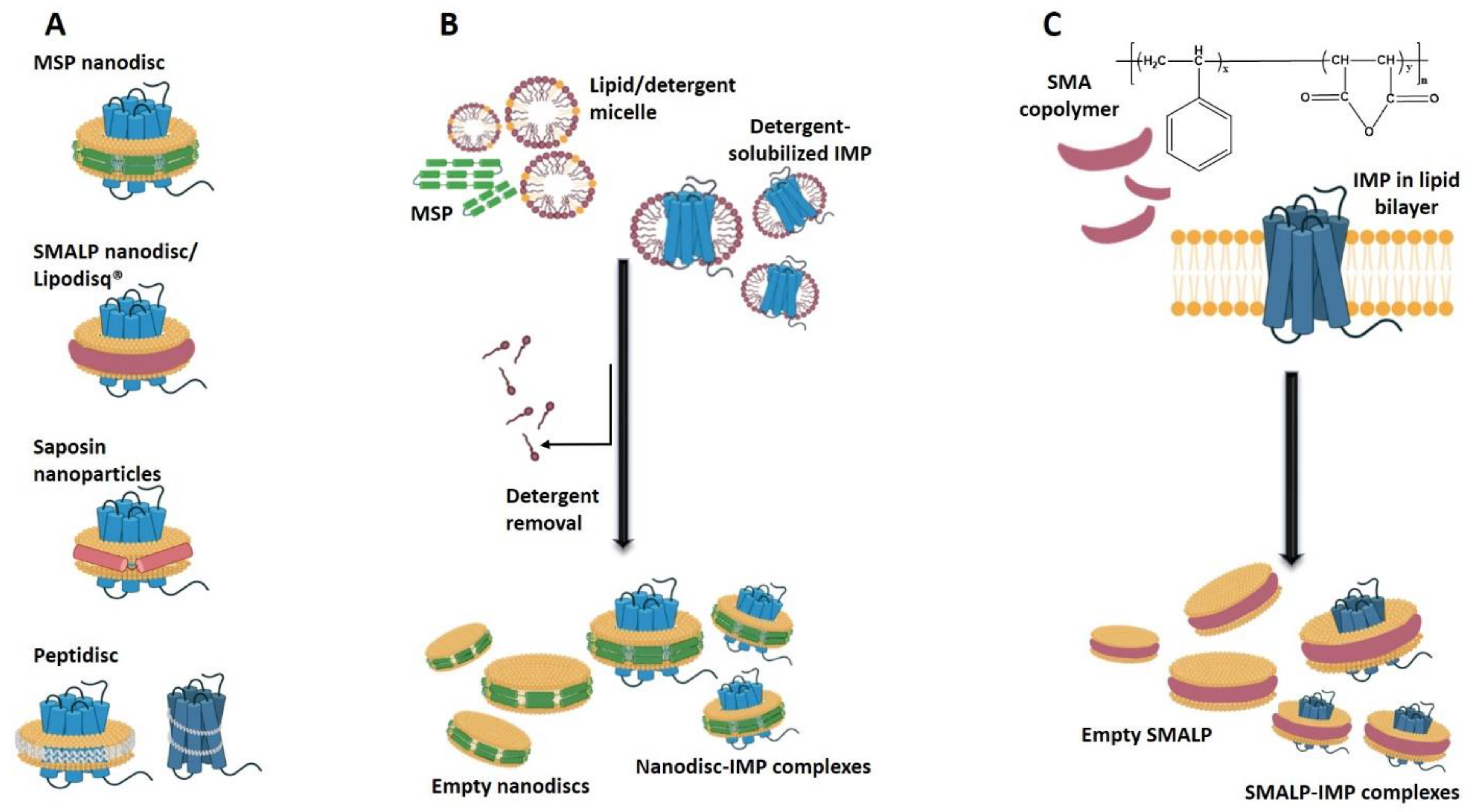
2.3. Nanodiscs in Studies of Integral Membrane Proteins
2.3.1. General Properties of Nanodiscs
2.3.2. Applications of Nanodiscs in Integral Membrane Protein Solubilization and Stabilization
2.3.3. Applications of Nanodiscs in Functional Studies of Integral Membrane Proteins
2.3.4. Applications of Nanodiscs in Studies of Integral Membrane Proteins Using Biophysical and Structural Biology Methods
2.4. Liposomes in Studies of Integral Membrane Proteins
2.4.1. General Properties of Liposomes
2.4.2. Reconstitution of Integral Membrane Proteins in Liposomes
2.4.3. Applications of Liposomes in Functional Studies of Integral Membrane Proteins
2.4.4. Applications of Liposomes in Studies of Integral Membrane Proteins Using Biophysical and Structural Biology Methods
2.5. Other Membrane Mimetics in Studies of Integral Membrane Proteins
2.5.1. Amphipols
2.5.2. Lipid Cubic Phases
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Fagerberg, L.; Jonasson, K.; von Heijne, G.; Uhlén, M.; Berglund, L. Prediction of the human membrane proteome. Proteomics 2010, 10, 1141–1149. [Google Scholar] [CrossRef]
- Wallin, E.; von Heijne, G. Genome-wide analysis of integral membrane proteins from eubacterial, archaean, and eukaryotic organisms. Protein Sci. 1998, 7, 1029–1038. [Google Scholar] [CrossRef]
- Cournia, Z.; Allen, T.W.; Andricioaei, I.; Antonny, B.; Baum, D.; Brannigan, G.; Buchete, N.V.; Deckman, J.T.; Delemotte, L.; Del Val, C.; et al. Membrane Protein Structure, Function, and Dynamics: A Perspective from Experiments and Theory. J. Membr. Biol. 2015, 248, 611–640. [Google Scholar] [CrossRef] [Green Version]
- Whitelegge, J.P. Integral membrane proteins and bilayer proteomics. Anal. Chem. 2013, 85, 2558–2568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, J.L. The lipid bilayer membrane and its protein constituents. J. Gen. Physiol. 2018, 150, 1472–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, C.R.; Myers, J.K. Disease-related misassembly of membrane proteins. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 25–51. [Google Scholar] [CrossRef] [PubMed]
- Marinko, J.T.; Huang, H.; Penn, W.D.; Capra, J.A.; Schlebach, J.P.; Sanders, C.R. Folding and Misfolding of Human Membrane Proteins in Health and Disease: From Single Molecules to Cellular Proteostasis. Chem. Rev. 2019, 119, 5537–5606. [Google Scholar] [CrossRef] [PubMed]
- Drews, J. Drug discovery: A historical perspective. Science 2000, 287, 1960–1964. [Google Scholar] [CrossRef] [Green Version]
- Overington, J.P.; Al-Lazikani, B.; Hopkins, A.L. How many drug targets are there? Nat. Rev. Drug Discov. 2006, 5, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Xie, H. Activity assay of membrane transport proteins. Acta Biochim. Biophys. Sin. 2008, 40, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Obergrussberger, A.; Stolzle-Feix, S.; Becker, N.; Bruggemann, A.; Fertig, N.; Moller, C. Novel screening techniques for ion channel targeting drugs. Channels 2015, 9, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Nozeret, K.; Boucharlat, A.; Agou, F.; Buddelmeijer, N. A sensitive fluorescence-based assay to monitor enzymatic activity of the essential integral membrane protein Apolipoprotein N-acyltransferase (Lnt). Sci. Rep. 2019, 9, 15978. [Google Scholar] [CrossRef] [PubMed]
- Quick, M.; Javitch, J.A. Monitoring the function of membrane transport proteins in detergent-solubilized form. Proc. Natl. Acad. Sci. USA 2007, 104, 3603–3608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loll, P.J. Membrane proteins, detergents and crystals: What is the state of the art? Acta Crystallogr. F Struct. Biol. Commun. 2014, 70, 1576–1583. [Google Scholar] [CrossRef] [Green Version]
- Bonnete, F.; Loll, P.J. Characterization of New Detergents and Detergent Mimetics by Scattering Techniques for Membrane Protein Crystallization. Methods Mol. Biol. 2017, 1635, 169–193. [Google Scholar] [CrossRef]
- Nikolaev, M.; Round, E.; Gushchin, I.; Polovinkin, V.; Balandin, T.; Kuzmichev, P.; Shevchenko, V.; Borshchevskiy, V.; Kuklin, A.; Round, A.; et al. Integral Membrane Proteins Can Be Crystallized Directly from Nanodiscs. Cryst. Growth Des. 2017, 17, 945–948. [Google Scholar] [CrossRef] [Green Version]
- Broecker, J.; Eger, B.T.; Ernst, O.P. Crystallogenesis of Membrane Proteins Mediated by Polymer-Bounded Lipid Nanodiscs. Structure 2017, 25, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Landau, E.M.; Rosenbusch, J.P. Lipidic cubic phases: A novel concept for the crystallization of membrane proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 14532–14535. [Google Scholar] [CrossRef] [Green Version]
- Cherezov, V. Lipidic cubic phase technologies for membrane protein structural studies. Curr. Opin. Struct. Biol. 2011, 21, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agah, S.; Faham, S. Crystallization of membrane proteins in bicelles. Methods Mol. Biol. 2012, 914, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Zorzi, R.D.; Mi, W.; Liao, M.; Walz, T. Single-particle electron microscopy in the study of membrane protein structure. Microscopy 2016, 65, 81–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinothkumar, K.R. Membrane protein structures without crystals, by single particle electron cryomicroscopy. Curr. Opin. Struct. Biol. 2015, 33, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Matthies, D.; Bae, C.; Toombes, G.E.; Fox, T.; Bartesaghi, A.; Subramaniam, S.; Swartz, K.J. Single-particle cryo-EM structure of a voltage-activated potassium channel in lipid nanodiscs. Elife 2018, 7, e37558. [Google Scholar] [CrossRef]
- Wang, S.; Brettmann, J.B.; Nichols, C.G. Studying Structural Dynamics of Potassium Channels by Single-Molecule FRET. Methods Mol. Biol. 2018, 1684, 163–180. [Google Scholar] [CrossRef]
- Gregorio, G.G.; Masureel, M.; Hilger, D.; Terry, D.S.; Juette, M.; Zhao, H.; Zhou, Z.; Perez-Aguilar, J.M.; Hauge, M.; Mathiasen, S.; et al. Single-molecule analysis of ligand efficacy in beta2AR-G-protein activation. Nature 2017, 547, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Asher, W.B.; Geggier, P.; Holsey, M.D.; Gilmore, G.T.; Pati, A.K.; Meszaros, J.; Terry, D.S.; Mathiasen, S.; Kaliszewski, M.J.; McCauley, M.D.; et al. Single-molecule FRET imaging of GPCR dimers in living cells. Nat. Methods 2021, 18, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Claxton, D.P.; Kazmier, K.; Mishra, S.; McHaourab, H.S. Navigating Membrane Protein Structure, Dynamics, and Energy Landscapes Using Spin Labeling and EPR Spectroscopy. Methods Enzymol. 2015, 564, 349–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgieva, E.R.; Borbat, P.P.; Ginter, C.; Freed, J.H.; Boudker, O. Conformational ensemble of the sodium-coupled aspartate transporter. Nat. Struct. Mol. Biol. 2013, 20, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Georgieva, E.R.; Borbat, P.P.; Norman, H.D.; Freed, J.H. Mechanism of influenza A M2 transmembrane domain assembly in lipid membranes. Sci. Rep. 2015, 5, 11757. [Google Scholar] [CrossRef] [Green Version]
- Georgieva, E.R. Nanoscale lipid membrane mimetics in spin-labeling and electron paramagnetic resonance spectroscopy studies of protein structure and function. Nanotechnol. Rev. 2017, 6, 75–92. [Google Scholar] [CrossRef]
- Jeschke, G. DEER distance measurements on proteins. Annu. Rev. Phys. Chem. 2012, 63, 419–446. [Google Scholar] [CrossRef] [Green Version]
- Borbat, P.P.; Surendhran, K.; Bortolus, M.; Zou, P.; Freed, J.H.; McHaourab, H.S. Conformational motion of the ABC transporter MsbA induced by ATP hydrolysis. PLoS Biol. 2007, 5, e271. [Google Scholar] [CrossRef]
- Altenbach, C.; Marti, T.; Khorana, H.G.; Hubbell, W.L. Transmembrane protein structure: Spin labeling of bacteriorhodopsin mutants. Science 1990, 248, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Gluck, J.M.; Wittlich, M.; Feuerstein, S.; Hoffmann, S.; Willbold, D.; Koenig, B.W. Integral membrane proteins in nanodiscs can be studied by solution NMR spectroscopy. J. Am. Chem. Soc. 2009, 131, 12060–12061. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Li, Q. Solution NMR study of integral membrane proteins. Curr. Opin. Chem. Biol. 2011, 15, 560–569. [Google Scholar] [CrossRef]
- Danmaliki, G.I.; Hwang, P.M. Solution NMR spectroscopy of membrane proteins. Biochim. Biophys. Acta BioMembr. 2020, 1862, 183356. [Google Scholar] [CrossRef]
- Patching, S.G. Solid-state NMR structures of integral membrane proteins. Mol. Membr. Biol. 2015, 32, 156–178. [Google Scholar] [CrossRef]
- Liang, B.; Tamm, L.K. NMR as a tool to investigate the structure, dynamics and function of membrane proteins. Nat. Struct. Mol. Biol. 2016, 23, 468–474. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.S.; Ladizhansky, V. Membrane proteins in their native habitat as seen by solid-state NMR spectroscopy. Protein Sci. 2015, 24, 1333–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balana, B.; Taylor, N.; Slesinger, P.A. Mutagenesis and functional analysis of ion channels heterologously expressed in mammalian cells. J. Vis. Exp. 2010, e2189. [Google Scholar] [CrossRef] [PubMed]
- Lueck, J.D.; Mackey, A.L.; Infield, D.T.; Galpin, J.D.; Li, J.; Roux, B.; Ahern, C.A. Atomic mutagenesis in ion channels with engineered stoichiometry. Elife 2016, 5, e18976. [Google Scholar] [CrossRef] [PubMed]
- Bezanilla, F. The voltage sensor in voltage-dependent ion channels. Physiol. Rev. 2000, 80, 555–592. [Google Scholar] [CrossRef] [PubMed]
- Chavent, M.; Duncan, A.L.; Sansom, M.S. Molecular dynamics simulations of membrane proteins and their interactions: From nanoscale to mesoscale. Curr. Opin. Struct. Biol. 2016, 40, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, T.; Han, W.; Maduke, M.; Tajkhorshid, E. Molecular Basis for Differential Anion Binding and Proton Coupling in the Cl(-)/H(+) Exchanger ClC-ec1. J. Am. Chem. Soc. 2016, 138, 3066–3075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, M.P.; Jiang, T.; Sun, C.; Lihan, M.; Pant, S.; Mahinthichaichan, P.; Trifan, A.; Tajkhorshid, E. Characterization of Lipid-Protein Interactions and Lipid-Mediated Modulation of Membrane Protein Function through Molecular Simulation. Chem. Rev. 2019, 119, 6086–6161. [Google Scholar] [CrossRef]
- Ahmad, I.; Nawaz, N.; Darwesh, N.M.; Ur Rahman, S.; Mustafa, M.Z.; Khan, S.B.; Patching, S.G. Overcoming challenges for amplified expression of recombinant proteins using Escherichia coli. Protein Expr. Purif. 2018, 144, 12–18. [Google Scholar] [CrossRef]
- Jensen, H.M.; Eng, T.; Chubukov, V.; Herbert, R.A.; Mukhopadhyay, A. Improving membrane protein expression and function using genomic edits. Sci. Rep. 2017, 7, 13030. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, K.; Javed, W.; Vallet, S.; Lesterlin, C.; Candusso, M.P.; Ding, F.; Xu, X.N.; Ebel, C.; Jault, J.M.; Orelle, C. Functionality of membrane proteins overexpressed and purified from E. coli is highly dependent upon the strain. Sci. Rep. 2019, 9, 2654. [Google Scholar] [CrossRef] [PubMed]
- Heimpel, S.; Basset, G.; Odoy, S.; Klingenberg, M. Expression of the mitochondrial ADP/ATP carrier in Escherichia coli. Renaturation, reconstitution, and the effect of mutations on 10 positive residues. J. Biol. Chem. 2001, 276, 11499–11506. [Google Scholar] [CrossRef] [Green Version]
- Angiulli, G.; Dhupar, H.S.; Suzuki, H.; Wason, I.S.; Duong Van Hoa, F.; Walz, T. New approach for membrane protein reconstitution into peptidiscs and basis for their adaptability to different proteins. Elife 2020, 9, e53530. [Google Scholar] [CrossRef]
- Chien, C.H.; Helfinger, L.R.; Bostock, M.J.; Solt, A.; Tan, Y.L.; Nietlispach, D. An Adaptable Phospholipid Membrane Mimetic System for Solution NMR Studies of Membrane Proteins. J. Am. Chem. Soc. 2017, 139, 14829–14832. [Google Scholar] [CrossRef] [PubMed]
- Schmidpeter, P.A.M.; Sukomon, N.; Nimigean, C.M. Reconstitution of Membrane Proteins into Platforms Suitable for Biophysical and Structural Analyses. Methods Mol. Biol. 2020, 2127, 191–205. [Google Scholar] [CrossRef]
- Georgieva, E.R.; Fanouraki, C.; Borbat, P. Purification and Biophysical Characterization Of LysE Membrane Exporter from Mycobacterium tuberculosis in Lipodiscs Made of Native E. coli Membranes and Detergent. Biophys. J. 2020, 118, 503a–504a. [Google Scholar] [CrossRef]
- Thoma, J.; Burmann, B.M. Fake It ‘Till You Make It—The Pursuit of Suitable Membrane Mimetics for Membrane Protein Biophysics. Int. J. Mol. Sci. 2021, 22, 50. [Google Scholar] [CrossRef]
- Lorsch, J. Laboratory Methods in Enzymology: Protein Part C; Elsevier Science: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Garavito, R.M.; Ferguson-Miller, S. Detergents as tools in membrane biochemistry. J. Biol. Chem. 2001, 276, 32403–32406. [Google Scholar] [CrossRef] [Green Version]
- Popot, J.-L. Alternatives to Detergents for Handling Membrane Proteins in Aqueous Solutions. In Membrane Proteins in Aqueous Solutions; Springer: Berlin/Heidelberg, Germany, 2018; pp. 97–149. [Google Scholar]
- Anson, M. The denaturation of proteins by detergents and bile salts. Science 1939, 90, 256–257. [Google Scholar] [CrossRef]
- Brooks, M.M. Comparative studies on respiration: XV. The Effect of Bile Salts and of Saponin upon Respiration. J. Gen. Physiol. 1921, 3, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.J.; Gabriel, F.; Tandale, A.; Nietlispach, D. Structure and Dynamics of GPCRs in Lipid Membranes: Physical Principles and Experimental Approaches. Molecules 2020, 25, 4729. [Google Scholar] [CrossRef] [PubMed]
- Seddon, A.M.; Curnow, P.; Booth, P.J. Membrane proteins, lipids and detergents: Not just a soap opera. Biochim. Biophys. Acta (BBA)-Biomembr. 2004, 1666, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Koehler, J.; Sulistijo, E.S.; Sakakura, M.; Kim, H.J.; Ellis, C.D.; Sanders, C.R. Lysophospholipid micelles sustain the stability and catalytic activity of diacylglycerol kinase in the absence of lipids. Biochemistry 2010, 49, 7089–7099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chipot, C.; Dehez, F.; Schnell, J.R.; Zitzmann, N.; Pebay-Peyroula, E.; Catoire, L.J.; Miroux, B.; Kunji, E.R.S.; Veglia, G.; Cross, T.A.; et al. Perturbations of Native Membrane Protein Structure in Alkyl Phosphocholine Detergents: A Critical Assessment of NMR and Biophysical Studies. Chem. Rev. 2018, 118, 3559–3607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almgren, M. Mixed micelles and other structures in the solubilization of bilayer lipid membranes by surfactants. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1508, 146–163. [Google Scholar] [CrossRef] [Green Version]
- Ilgü, H.; Jeckelmann, J.-M.; Gachet, M.S.; Boggavarapu, R.; Ucurum, Z.; Gertsch, J.; Fotiadis, D. Variation of the detergent-binding capacity and phospholipid content of membrane proteins when purified in different detergents. Biophys. J. 2014, 106, 1660–1670. [Google Scholar] [CrossRef] [Green Version]
- Rigaud, J.-L.; Levy, D.; Mosser, G.; Lambert, O. Detergent removal by non-polar polystyrene beads. Eur. Biophys. J. 1998, 27, 305–319. [Google Scholar] [CrossRef]
- Tulumello, D.V.; Deber, C.M. Efficiency of detergents at maintaining membrane protein structures in their biologically relevant forms. Biochim. Biophys. Acta (BBA)-Biomembr. 2012, 1818, 1351–1358. [Google Scholar] [CrossRef] [Green Version]
- Wiener, M.C. A pedestrian guide to membrane protein crystallization. Methods 2004, 34, 364–372. [Google Scholar] [CrossRef]
- Klöpfer, K.; Hagn, F. Beyond detergent micelles: The advantages and applications of non-micellar and lipid-based membrane mimetics for solution-state NMR. Prog. Nucl. Magn. Reson. Spectrosc. 2019, 114, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.-X.; Cross, T.A. Influences of membrane mimetic environments on membrane protein structures. Annu. Rev. Biophys. 2013, 42, 361–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Mao, A.; Bhattacharya, S.; Robertson, N.; Grisshammer, R.; Tate, C.G.; Vaidehi, N. How do short chain nonionic detergents destabilize G-protein-coupled receptors? J. Am. Chem. Soc. 2016, 138, 15425–15433. [Google Scholar] [CrossRef] [Green Version]
- Rajarathnam, K.; Rosgen, J. Isothermal titration calorimetry of membrane proteins—Progress and challenges. Biochim. Biophys. Acta 2014, 1838, 69–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draczkowski, P.; Matosiuk, D.; Jozwiak, K. Isothermal titration calorimetry in membrane protein research. J. Pharm Biomed. Anal. 2014, 87, 313–325. [Google Scholar] [CrossRef]
- Hariharan, P.; Guan, L. Thermodynamic cooperativity of cosubstrate binding and cation selectivity of Salmonella typhimurium MelB. J. Gen. Physiol. 2017, 149, 1029–1039. [Google Scholar] [CrossRef] [Green Version]
- Amin, A.; Hariharan, P.; Chae, P.S.; Guan, L. Effect of Detergents on Galactoside Binding by Melibiose Permeases. Biochemistry 2015, 54, 5849–5855. [Google Scholar] [CrossRef] [Green Version]
- Reich-Slotky, R.; Panagiotidis, C.; Reyes, M.; Shuman, H.A. The detergent-soluble maltose transporter is activated by maltose binding protein and verapamil. J. Bacteriol. 2000, 182, 993–1000. [Google Scholar] [CrossRef] [Green Version]
- Schneider, E.; Hunke, S. ATP-binding-cassette (ABC) transport systems: Functional and structural aspects of the ATP-hydrolyzing subunits/domains. FEMS MicroBiol. Rev. 1998, 22, 1–20. [Google Scholar] [CrossRef]
- Infed, N.; Hanekop, N.; Driessen, A.J.; Smits, S.H.; Schmitt, L. Influence of detergents on the activity of the ABC transporter LmrA. Biochim. Biophys. Acta 2011, 1808, 2313–2321. [Google Scholar] [CrossRef] [Green Version]
- Slotboom, D.J.; Duurkens, R.H.; Olieman, K.; Erkens, G.B. Static light scattering to characterize membrane proteins in detergent solution. Methods 2008, 46, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Kawate, T.; Gouaux, E. Fluorescence-detection size-exclusion chromatography for precrystallization screening of integral membrane proteins. Structure 2006, 14, 673–681. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, A.I.; Mileni, M.; Chien, E.Y.; Hanson, M.A.; Stevens, R.C. Microscale fluorescent thermal stability assay for membrane proteins. Structure 2008, 16, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Kotov, V.; Bartels, K.; Veith, K.; Josts, I.; Subhramanyam, U.K.T.; Gunther, C.; Labahn, J.; Marlovits, T.C.; Moraes, I.; Tidow, H.; et al. High-throughput stability screening for detergent-solubilized membrane proteins. Sci. Rep. 2019, 9, 10379. [Google Scholar] [CrossRef] [Green Version]
- Miles, A.J.; Wallace, B.A. Circular dichroism spectroscopy of membrane proteins. Chem. Soc. Rev. 2016, 45, 4859–4872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ireland, S.M.; Sula, A.; Wallace, B.A. Thermal melt circular dichroism spectroscopic studies for identifying stabilising amphipathic molecules for the voltage-gated sodium channel NavMs. Biopolymers 2018, 109, e23067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancusso, R.; Karpowich, N.K.; Czyzewski, B.K.; Wang, D.N. Simple screening method for improving membrane protein thermostability. Methods 2011, 55, 324–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunji, E.R.; Harding, M.; Butler, P.J.; Akamine, P. Determination of the molecular mass and dimensions of membrane proteins by size exclusion chromatography. Methods 2008, 46, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.L.; Newstead, S. Membrane Protein Crystallisation: Current Trends and Future Perspectives. Adv. Exp. Med. Biol. 2016, 922, 61–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newstead, S.; Ferrandon, S.; Iwata, S. Rationalizing alpha-helical membrane protein crystallization. Protein Sci. 2008, 17, 466–472. [Google Scholar] [CrossRef] [Green Version]
- Newstead, S.; Hobbs, J.; Jordan, D.; Carpenter, E.P.; Iwata, S. Insights into outer membrane protein crystallization. Mol. Membr. Biol. 2008, 25, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Sonoda, Y.; Cameron, A.; Newstead, S.; Omote, H.; Moriyama, Y.; Kasahara, M.; Iwata, S.; Drew, D. Tricks of the trade used to accelerate high-resolution structure determination of membrane proteins. FEBS Lett. 2010, 584, 2539–2547. [Google Scholar] [CrossRef] [Green Version]
- Durr, K.L.; Chen, L.; Stein, R.A.; De Zorzi, R.; Folea, I.M.; Walz, T.; McHaourab, H.S.; Gouaux, E. Structure and dynamics of AMPA receptor GluA2 in resting, pre-open, and desensitized states. Cell 2014, 158, 778–792. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, H.; Gouaux, E. X-ray structures of LeuT in substrate-free outward-open and apo inward-open states. Nature 2012, 481, 469–474. [Google Scholar] [CrossRef] [Green Version]
- Quick, M.; Winther, A.M.; Shi, L.; Nissen, P.; Weinstein, H.; Javitch, J.A. Binding of an octylglucoside detergent molecule in the second substrate (S2) site of LeuT establishes an inhibitor-bound conformation. Proc. Natl. Acad. Sci. USA 2009, 106, 5563–5568. [Google Scholar] [CrossRef] [Green Version]
- Perez, C.; Koshy, C.; Yildiz, O.; Ziegler, C. Alternating-access mechanism in conformationally asymmetric trimers of the betaine transporter BetP. Nature 2012, 490, 126–130. [Google Scholar] [CrossRef]
- Birch, J.; Cheruvara, H.; Gamage, N.; Harrison, P.J.; Lithgo, R.; Quigley, A. Changes in Membrane Protein Structural Biology. Biology 2020, 9, 401. [Google Scholar] [CrossRef] [PubMed]
- Gewering, T.; Januliene, D.; Ries, A.B.; Moeller, A. Know your detergents: A case study on detergent background in negative stain electron microscopy. J. Struct. Biol. 2018, 203, 242–246. [Google Scholar] [CrossRef]
- Hauer, F.; Gerle, C.; Fischer, N.; Oshima, A.; Shinzawa-Itoh, K.; Shimada, S.; Yokoyama, K.; Fujiyoshi, Y.; Stark, H. GraDeR: Membrane Protein Complex Preparation for Single-Particle Cryo-EM. Structure 2015, 23, 1769–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autzen, H.E.; Julius, D.; Cheng, Y. Membrane mimetic systems in CryoEM: Keeping membrane proteins in their native environment. Curr. Opin. Struct. Biol. 2019, 58, 259–268. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, J. Atomic Structure of the Cystic Fibrosis Transmembrane Conductance Regulator. Cell 2016, 167, 1586–1597.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basak, S.; Gicheru, Y.; Samanta, A.; Molugu, S.K.; Huang, W.; Fuente, M.; Hughes, T.; Taylor, D.J.; Nieman, M.T.; Moiseenkova-Bell, V.; et al. Cryo-EM structure of 5-HT3A receptor in its resting conformation. Nat. Commun. 2018, 9, 514. [Google Scholar] [CrossRef]
- Basak, S.; Gicheru, Y.; Rao, S.; Sansom, M.S.P.; Chakrapani, S. Cryo-EM reveals two distinct serotonin-bound conformations of full-length 5-HT3A receptor. Nature 2018, 563, 270–274. [Google Scholar] [CrossRef]
- Basak, S.; Kumar, A.; Ramsey, S.; Gibbs, E.; Kapoor, A.; Filizola, M.; Chakrapani, S. High-resolution structures of multiple 5-HT3AR-setron complexes reveal a novel mechanism of competitive inhibition. Elife 2020, 9, e57870. [Google Scholar] [CrossRef]
- Mineev, K.S.; Nadezhdin, K.D. Membrane mimetics for solution NMR studies of membrane proteins. Nanotechnol. Rev. 2017, 6, 15–32. [Google Scholar] [CrossRef]
- Meissner, A.; Sorensen, O.W. Optimization of three-dimensional TROSY-type HCCH NMR correlation of aromatic (1)H-(13)C groups in proteins. J. Magn. Reson. 1999, 139, 447–450. [Google Scholar] [CrossRef]
- Tugarinov, V.; Hwang, P.M.; Kay, L.E. Nuclear magnetic resonance spectroscopy of high-molecular-weight proteins. Annu. Rev. BioChem. 2004, 73, 107–146. [Google Scholar] [CrossRef] [Green Version]
- Columbus, L.; Lipfert, J.; Jambunathan, K.; Fox, D.A.; Sim, A.Y.; Doniach, S.; Lesley, S.A. Mixing and matching detergents for membrane protein NMR structure determination. J. Am. Chem. Soc. 2009, 131, 7320–7326. [Google Scholar] [CrossRef]
- Hiller, S.; Garces, R.G.; Malia, T.J.; Orekhov, V.Y.; Colombini, M.; Wagner, G. Solution structure of the integral human membrane protein VDAC-1 in detergent micelles. Science 2008, 321, 1206–1210. [Google Scholar] [CrossRef] [Green Version]
- Bayrhuber, M.; Meins, T.; Habeck, M.; Becker, S.; Giller, K.; Villinger, S.; Vonrhein, C.; Griesinger, C.; Zweckstetter, M.; Zeth, K. Structure of the human voltage-dependent anion channel. Proc. Natl. Acad. Sci. USA 2008, 105, 15370–15375. [Google Scholar] [CrossRef] [Green Version]
- Liang, B.; Tamm, L.K. Structure of outer membrane protein G by solution NMR spectroscopy. Proc. Natl. Acad. Sci. USA 2007, 104, 16140–16145. [Google Scholar] [CrossRef] [Green Version]
- Etzkorn, M.; Raschle, T.; Hagn, F.; Gelev, V.; Rice, A.J.; Walz, T.; Wagner, G. Cell-free expressed bacteriorhodopsin in different soluble membrane mimetics: Biophysical properties and NMR accessibility. Structure 2013, 21, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Isogai, S.; Deupi, X.; Opitz, C.; Heydenreich, F.M.; Tsai, C.J.; Brueckner, F.; Schertler, G.F.; Veprintsev, D.B.; Grzesiek, S. Backbone NMR reveals allosteric signal transduction networks in the beta1-adrenergic receptor. Nature 2016, 530, 237–241. [Google Scholar] [CrossRef]
- Nygaard, R.; Zou, Y.; Dror, R.O.; Mildorf, T.J.; Arlow, D.H.; Manglik, A.; Pan, A.C.; Liu, C.W.; Fung, J.J.; Bokoch, M.P.; et al. The dynamic process of beta(2)-adrenergic receptor activation. Cell 2013, 152, 532–542. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, S.; Kimura, T.; Nozaki, T.; Harada, H.; Shimada, I.; Osawa, M. Structural basis for the inhibition of voltage-dependent K+ channel by gating modifier toxin. Sci. Rep. 2015, 5, 14226. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Horst, R.; Geralt, M.; Ma, X.; Hong, W.X.; Finn, M.G.; Stevens, R.C.; Wuthrich, K. Microscale NMR screening of new detergents for membrane protein structural biology. J. Am. Chem. Soc. 2008, 130, 7357–7363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horst, R.; Stanczak, P.; Wuthrich, K. NMR polypeptide backbone conformation of the E. coli outer membrane protein W. Structure 2014, 22, 1204–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maslennikov, I.; Klammt, C.; Hwang, E.; Kefala, G.; Okamura, M.; Esquivies, L.; Mors, K.; Glaubitz, C.; Kwiatkowski, W.; Jeon, Y.H.; et al. Membrane domain structures of three classes of histidine kinase receptors by cell-free expression and rapid NMR analysis. Proc. Natl. Acad. Sci. USA 2010, 107, 10902–10907. [Google Scholar] [CrossRef] [Green Version]
- Kucharska, I.; Seelheim, P.; Edrington, T.; Liang, B.; Tamm, L.K. OprG Harnesses the Dynamics of its Extracellular Loops to Transport Small Amino Acids across the Outer Membrane of Pseudomonas aeruginosa. Structure 2015, 23, 2234–2245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altenbach, C.; Greenhalgh, D.A.; Khorana, H.G.; Hubbell, W.L. A collision gradient method to determine the immersion depth of nitroxides in lipid bilayers: Application to spin-labeled mutants of bacteriorhodopsin. Proc. Natl. Acad. Sci. USA 1994, 91, 1667–1671. [Google Scholar] [CrossRef] [Green Version]
- Bordignon, E. Site-directed spin labeling of membrane proteins. Top. Curr. Chem. 2012, 321, 121–157. [Google Scholar] [CrossRef] [PubMed]
- Hubbell, W.L.; McHaourab, H.S.; Altenbach, C.; Lietzow, M.A. Watching proteins move using site-directed spin labeling. Structure 1996, 4, 779–783. [Google Scholar] [CrossRef] [Green Version]
- Klug, C.S.; Feix, J.B. Methods and applications of site-directed spin labeling EPR spectroscopy. Methods Cell Biol. 2008, 84, 617–658. [Google Scholar] [CrossRef] [PubMed]
- McHaourab, H.S.; Steed, P.R.; Kazmier, K. Toward the fourth dimension of membrane protein structure: Insight into dynamics from spin-labeling EPR spectroscopy. Structure 2011, 19, 1549–1561. [Google Scholar] [CrossRef] [Green Version]
- Sahu, I.D.; Lorigan, G.A. Site-Directed Spin Labeling EPR for Studying Membrane Proteins. BioMed Res. Int. 2018, 2018, 3248289. [Google Scholar] [CrossRef]
- Georgieva, E.R.; Fanouraki, C.; Borbat, P.P. Expression, purification and initial characterization of LysE membrane exporter from Mycobacterium tuberculosis: Towards comprehensive functional and structural study. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Hanelt, I.; Wunnicke, D.; Bordignon, E.; Steinhoff, H.J.; Slotboom, D.J. Conformational heterogeneity of the aspartate transporter Glt(Ph). Nat. Struct. Mol. Biol. 2013, 20, 210–214. [Google Scholar] [CrossRef] [Green Version]
- Hilger, D.; Polyhach, Y.; Padan, E.; Jung, H.; Jeschke, G. High-resolution structure of a Na+/H+ antiporter dimer obtained by pulsed electron paramagnetic resonance distance measurements. Biophys. J. 2007, 93, 3675–3683. [Google Scholar] [CrossRef] [Green Version]
- Joseph, B.; Jeschke, G.; Goetz, B.A.; Locher, K.P.; Bordignon, E. Transmembrane gate movements in the type II ATP-binding cassette (ABC) importer BtuCD-F during nucleotide cycle. J. Biol. Chem. 2011, 286, 41008–41017. [Google Scholar] [CrossRef] [Green Version]
- Nicklisch, S.C.; Wunnicke, D.; Borovykh, I.V.; Morbach, S.; Klare, J.P.; Steinhoff, H.J.; Kramer, R. Conformational changes of the betaine transporter BetP from Corynebacterium glutamicum studied by pulse EPR spectroscopy. Biochim. Biophys. Acta 2012, 1818, 359–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnova, I.; Kasho, V.; Choe, J.Y.; Altenbach, C.; Hubbell, W.L.; Kaback, H.R. Sugar binding induces an outward facing conformation of LacY. Proc. Natl. Acad. Sci. USA 2007, 104, 16504–16509. [Google Scholar] [CrossRef] [Green Version]
- Yardeni, E.H.; Bahrenberg, T.; Stein, R.A.; Mishra, S.; Zomot, E.; Graham, B.; Tuck, K.L.; Huber, T.; Bibi, E.; McHaourab, H.S.; et al. Probing the solution structure of the E. coli multidrug transporter MdfA using DEER distance measurements with nitroxide and Gd(III) spin labels. Sci. Rep. 2019, 9, 12528. [Google Scholar] [CrossRef] [Green Version]
- Dastvan, R.; Mishra, S.; Peskova, Y.B.; Nakamoto, R.K.; McHaourab, H.S. Mechanism of allosteric modulation of P-glycoprotein by transport substrates and inhibitors. Science 2019, 364, 689–692. [Google Scholar] [CrossRef]
- Georgieva, E.R.; Borbat, P.P.; Grushin, K.; Stoilova-McPhie, S.; Kulkarni, N.J.; Liang, Z.; Freed, J.H. Conformational Response of Influenza A M2 Transmembrane Domain to Amantadine Drug Binding at Low pH (pH 5.5). Front. Physiol. 2016, 7, 317. [Google Scholar] [CrossRef] [Green Version]
- Cuello, L.G.; Jogini, V.; Cortes, D.M.; Sompornpisut, A.; Purdy, M.D.; Wiener, M.C.; Perozo, E. Design and characterization of a constitutively open KcsA. FEBS Lett. 2010, 584, 1133–1138. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Stein, R.A.; Yoshioka, C.; Lee, C.H.; Goehring, A.; McHaourab, H.S.; Gouaux, E. Mechanism of NMDA Receptor Inhibition and Activation. Cell 2016, 165, 704–714. [Google Scholar] [CrossRef] [Green Version]
- Wingler, L.M.; Elgeti, M.; Hilger, D.; Latorraca, N.R.; Lerch, M.T.; Staus, D.P.; Dror, R.O.; Kobilka, B.K.; Hubbell, W.L.; Lefkowitz, R.J. Angiotensin Analogs with Divergent Bias Stabilize Distinct Receptor Conformations. Cell 2019, 176, 468–478.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castell, O.K.; Dijkman, P.M.; Wiseman, D.N.; Goddard, A.D. Single molecule fluorescence for membrane proteins. Methods 2018, 147, 221–228. [Google Scholar] [CrossRef]
- Bartels, K.; Lasitza-Male, T.; Hofmann, H.; Low, C. Single-Molecule FRET of Membrane Transport Proteins. ChembioChem 2021. [Google Scholar] [CrossRef]
- Zhao, Y.; Terry, D.; Shi, L.; Weinstein, H.; Blanchard, S.C.; Javitch, J.A. Single-molecule dynamics of gating in a neurotransmitter transporter homologue. Nature 2010, 465, 188–193. [Google Scholar] [CrossRef] [Green Version]
- Raghuraman, H.; Chatterjee, S.; Das, A. Site-Directed Fluorescence Approaches for Dynamic Structural Biology of Membrane Peptides and Proteins. Front. Mol. BioSci. 2019, 6, 96. [Google Scholar] [CrossRef] [PubMed]
- Ram, P.; Prestegard, J.H. Magnetic field induced ordering of bile salt/phospholipid micelles: New media for NMR structural investigations. Biochim. Biophys. Acta 1988, 940, 289–294. [Google Scholar] [CrossRef]
- Sanders, C.R.; Landis, G.C. Reconstitution of membrane proteins into lipid-rich bilayered mixed micelles for NMR studies. Biochemistry 1995, 34, 4030–4040. [Google Scholar] [CrossRef]
- Sanders, C.R.; Prosser, R.S. Bicelles: A model membrane system for all seasons? Structure 1998, 6, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Lucyanna, B.B.; Gelen, R.; Merce, C.; Laia, R.; Carmen, L.I.; de Alfons, L.M.; Olga, L. Structural Versatility of Bicellar Systems and Their Possibilities as Colloidal Carriers. Pharmaceutics 2011, 3, 636–664. [Google Scholar] [CrossRef]
- Nusair, N.A.; Tiburu, E.K.; Dave, P.C.; Lorigan, G.A. Investigating fatty acids inserted into magnetically aligned phospholipid bilayers using EPR and solid-state NMR spectroscopy. J. Magn. Reson. 2004, 168, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.X.; Caporini, M.A.; Lorigan, G.A. The effects of cholesterol on magnetically aligned phospholipid bilayers: A solid-state NMR and EPR spectroscopy study. J. Magn. Reson. 2004, 168, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Piai, A.; Fu, Q.; Dev, J.; Chou, J.J. Optimal bicelle q for solution NMR studies of protein transmembrane partition. Chemistry 2017, 23, 1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poget, S.F.; Girvin, M.E. Solution NMR of membrane proteins in bilayer mimics: Small is beautiful, but sometimes bigger is better. Biochim. Biophys. Acta (BBA)-Biomembr. 2007, 1768, 3098–3106. [Google Scholar] [CrossRef] [Green Version]
- Puthenveetil, R.; Vinogradova, O. Solution NMR: A powerful tool for structural and functional studies of membrane proteins in reconstituted environments. J. Biol. Chem. 2019, 294, 15914–15931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgieva, E.R.; Ramlall, T.F.; Borbat, P.P.; Freed, J.H.; Eliezer, D. Membrane-bound alpha-synuclein forms an extended helix: Long-distance pulsed ESR measurements using vesicles, bicelles, and rodlike micelles. J. Am. Chem. Soc. 2008, 130, 12856–12857. [Google Scholar] [CrossRef] [Green Version]
- Glover, K.J.; Whiles, J.A.; Wu, G.; Yu, N.; Deems, R.; Struppe, J.O.; Stark, R.E.; Komives, E.A.; Vold, R.R. Structural evaluation of phospholipid bicelles for solution-state studies of membrane-associated biomolecules. Biophys. J. 2001, 81, 2163–2171. [Google Scholar] [CrossRef] [Green Version]
- Yeh, V.; Goode, A.; Bonev, B.B. Membrane protein structure determination and characterisation by solution and solid-state nmr. Biology 2020, 9, 396. [Google Scholar] [CrossRef]
- Prosser, R.S.; Evanics, F.; Kitevski, J.L.; Al-Abdul-Wahid, M.S. Current applications of bicelles in NMR studies of membrane-associated amphiphiles and proteins. Biochemistry 2006, 45, 8453–8465. [Google Scholar] [CrossRef]
- Vold, R.R.; Prosser, R.S. Magnetically oriented phospholipid bilayered micelles for structural studies of polypeptides. Does the ideal bicelle exist? J. Magn. Reson. 1996, 113, 267–271. [Google Scholar] [CrossRef]
- Smrt, S.T.; Draney, A.W.; Singaram, I.; Lorieau, J.L. Structure and Dynamics of Membrane Proteins and Membrane Associated Proteins with Native Bicelles from Eukaryotic Tissues. Biochemistry 2017, 56, 5318–5327. [Google Scholar] [CrossRef]
- Cardon, T.B.; Tiburu, E.K.; Padmanabhan, A.; Howard, K.P.; Lorigan, G.A. Magnetically aligned phospholipid bilayers at the parallel and perpendicular orientations for X-band spin-label EPR studies. J. Am. Chem. Soc. 2001, 123, 2913–2914. [Google Scholar] [CrossRef]
- Tiburu, E.K.; Moton, D.M.; Lorigan, G.A. Development of magnetically aligned phospholipid bilayers in mixtures of palmitoylstearoylphosphatidylcholine and dihexanoylphosphatidylcholine by solid-state NMR spectroscopy. Biochim. Biophys. Acta 2001, 1512, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Fanucci, G.E.; Lee, J.Y.; Cafiso, D.S. Membrane mimetic environments alter the conformation of the outer membrane protein BtuB. J. Am. Chem. Soc. 2003, 125, 13932–13933. [Google Scholar] [CrossRef]
- Uchida, N.; Nishizawa Horimoto, N.; Yamada, K.; Hikima, T.; Ishida, Y. Kinetically Stable Bicelles with Dilution Tolerance, Size Tunability, and Thermoresponsiveness for Drug Delivery Applications. ChembioChem 2018, 19, 1922–1926. [Google Scholar] [CrossRef] [PubMed]
- Dürr, U.H.; Gildenberg, M.; Ramamoorthy, A. The magic of bicelles lights up membrane protein structure. Chem. Rev. 2012, 112, 6054–6074. [Google Scholar] [CrossRef]
- Morrison, E.A.; Henzler-Wildman, K.A. Reconstitution of integral membrane proteins into isotropic bicelles with improved sample stability and expanded lipid composition profile. Biochim. Biophys. Acta 2012, 1818, 814–820. [Google Scholar] [CrossRef] [Green Version]
- Graslund, A.; Maler, L. Testing membrane interactions of CPPs. Methods Mol. Biol. 2011, 683, 33–40. [Google Scholar] [CrossRef]
- Maler, L. Solution NMR studies of peptide-lipid interactions in model membranes. Mol. Membr. Biol. 2012, 29, 155–176. [Google Scholar] [CrossRef]
- Gautier, A.; Mott, H.R.; Bostock, M.J.; Kirkpatrick, J.P.; Nietlispach, D. Structure determination of the seven-helix transmembrane receptor sensory rhodopsin II by solution NMR spectroscopy. Nat. Struct. Mol. Biol. 2010, 17, 768–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, E.A.; DeKoster, G.T.; Dutta, S.; Vafabakhsh, R.; Clarkson, M.W.; Bahl, A.; Kern, D.; Ha, T.; Henzler-Wildman, K.A. Antiparallel EmrE exports drugs by exchanging between asymmetric structures. Nature 2012, 481, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bocharov, E.V.; Mayzel, M.L.; Volynsky, P.E.; Goncharuk, M.V.; Ermolyuk, Y.S.; Schulga, A.A.; Artemenko, E.O.; Efremov, R.G.; Arseniev, A.S. Spatial structure and pH-dependent conformational diversity of dimeric transmembrane domain of the receptor tyrosine kinase EphA1. J. Biol. Chem. 2008, 283, 29385–29395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opella, S.J.; Marassi, F.M. Structure determination of membrane proteins by NMR spectroscopy. Chem. Rev. 2004, 104, 3587–3606. [Google Scholar] [CrossRef] [Green Version]
- Carlotti, C.; Aussenac, F.; Dufourc, E.J. Towards high-resolution 1H-NMR in biological membranes: Magic angle spinning of bicelles. Biochim. Biophys. Acta 2002, 1564, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Ujwal, R.; Bowie, J.U. Crystallizing membrane proteins using lipidic bicelles. Methods 2011, 55, 337–341. [Google Scholar] [CrossRef] [Green Version]
- Ujwal, R.; Abramson, J. High-throughput crystallization of membrane proteins using the lipidic bicelle method. J. Vis. Exp. 2012, e3383. [Google Scholar] [CrossRef]
- Faham, S.; Bowie, J.U. Bicelle crystallization: A new method for crystallizing membrane proteins yields a monomeric bacteriorhodopsin structure. J. Mol. Biol. 2002, 316, 1–6. [Google Scholar] [CrossRef]
- Rasmussen, S.G.; Choi, H.J.; Rosenbaum, D.M.; Kobilka, T.S.; Thian, F.S.; Edwards, P.C.; Burghammer, M.; Ratnala, V.R.; Sanishvili, R.; Fischetti, R.F.; et al. Crystal structure of the human beta2 adrenergic G-protein-coupled receptor. Nature 2007, 450, 383–387. [Google Scholar] [CrossRef]
- Vinothkumar, K.R. Structure of rhomboid protease in a lipid environment. J. Mol. Biol. 2011, 407, 232–247. [Google Scholar] [CrossRef] [Green Version]
- Ujwal, R.; Cascio, D.; Colletier, J.P.; Faham, S.; Zhang, J.; Toro, L.; Ping, P.; Abramson, J. The crystal structure of mouse VDAC1 at 2.3 A resolution reveals mechanistic insights into metabolite gating. Proc. Natl. Acad. Sci. USA 2008, 105, 17742–17747. [Google Scholar] [CrossRef] [Green Version]
- Sahu, I.D.; Mayo, D.J.; Subbaraman, N.; Inbaraj, J.J.; McCarrick, R.M.; Lorigan, G.A. Probing topology and dynamics of the second transmembrane domain (M2delta) of the acetyl choline receptor using magnetically aligned lipid bilayers (bicelles) and EPR spectroscopy. Chem. Phys. Lipids 2017, 206, 9–15. [Google Scholar] [CrossRef]
- Nusair, N.A.; Mayo, D.J.; Dorozenski, T.D.; Cardon, T.B.; Inbaraj, J.J.; Karp, E.S.; Newstadt, J.P.; Grosser, S.M.; Lorigan, G.A. Time-resolved EPR immersion depth studies of a transmembrane peptide incorporated into bicelles. Biochim. Biophys. Acta 2012, 1818, 821–828. [Google Scholar] [CrossRef] [Green Version]
- Bountra, K.; Hagelueken, G.; Choudhury, H.G.; Corradi, V.; El Omari, K.; Wagner, A.; Mathavan, I.; Zirah, S.; Yuan Wahlgren, W.; Tieleman, D.P.; et al. Structural basis for antibacterial peptide self-immunity by the bacterial ABC transporter McjD. EMBO J. 2017, 36, 3062–3079. [Google Scholar] [CrossRef]
- Bayburt, T.H.; Grinkova, Y.V.; Sligar, S.G. Self-assembly of discoidal phospholipid bilayer nanoparticles with membrane scaffold proteins. Nano Lett. 2002, 2, 853–856. [Google Scholar] [CrossRef]
- Bayburt, T.H.; Carlson, J.W.; Sligar, S.G. Reconstitution and imaging of a membrane protein in a nanometer-size phospholipid bilayer. J. Struct. Biol. 1998, 123, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denisov, I.G.; Sligar, S.G. Nanodiscs in membrane biochemistry and biophysics. Chem. Rev. 2017, 117, 4669–4713. [Google Scholar] [CrossRef] [PubMed]
- Civjan, N.R.; Bayburt, T.H.; Schuler, M.A.; Sligar, S.G. Direct solubilization of heterologously expressed membrane proteins by incorporation into nanoscale lipid bilayers. Biotechniques 2003, 35, 556–563. [Google Scholar] [CrossRef]
- Frauenfeld, J.; Löving, R.; Armache, J.-P.; Sonnen, A.F.; Guettou, F.; Moberg, P.; Zhu, L.; Jegerschöld, C.; Flayhan, A.; Briggs, J.A. A saposin-lipoprotein nanoparticle system for membrane proteins. Nat. Methods 2016, 13, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Carlson, M.L.; Young, J.W.; Zhao, Z.; Fabre, L.; Jun, D.; Li, J.; Li, J.; Dhupar, H.S.; Wason, I.; Mills, A.T. The Peptidisc, a simple method for stabilizing membrane proteins in detergent-free solution. Elife 2018, 7, e34085. [Google Scholar] [CrossRef]
- Juarez, J.F.B.; Harper, A.J.; Judge, P.J.; Tonge, S.R.; Watts, A. From polymer chemistry to structural biology: The development of SMA and related amphipathic polymers for membrane protein extraction and solubilisation. Chem. Phys. Lipids 2019, 221, 167–175. [Google Scholar] [CrossRef]
- Hagn, F.; Etzkorn, M.; Raschle, T.; Wagner, G. Optimized phospholipid bilayer nanodiscs facilitate high-resolution structure determination of membrane proteins. J. Am. Chem. Soc. 2013, 135, 1919–1925. [Google Scholar] [CrossRef] [Green Version]
- Hiller, S.; Wagner, G. Solution NMR spectroscopy of integral membrane proteins. In Comprehensive Biophysics; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Boldog, T.; Grimme, S.; Li, M.; Sligar, S.G.; Hazelbauer, G.L. Nanodiscs separate chemoreceptor oligomeric states and reveal their signaling properties. Proc. Natl. Acad. Sci. USA 2006, 103, 11509–11514. [Google Scholar] [CrossRef] [Green Version]
- Bibow, S.; Polyhach, Y.; Eichmann, C.; Chi, C.N.; Kowal, J.; Albiez, S.; McLeod, R.A.; Stahlberg, H.; Jeschke, G.; Güntert, P. Solution structure of discoidal high-density lipoprotein particles with a shortened apolipoprotein AI. Nat. Struct. Mol. Biol. 2017, 24, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Denisov, I.G.; Grinkova, Y.V.; Lazarides, A.A.; Sligar, S.G. Directed self-assembly of monodisperse phospholipid bilayer Nanodiscs with controlled size. J. Am. Chem. Soc. 2004, 126, 3477–3487. [Google Scholar] [CrossRef]
- Grinkova, Y.V.; Denisov, I.G.; Sligar, S.G. Engineering extended membrane scaffold proteins for self-assembly of soluble nanoscale lipid bilayers. Protein Eng. Des. Sel. 2010, 23, 843–848. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, T.K.; Grinkova, Y.V.; Bayburt, T.H.; Denisov, I.G.; Zolnerciks, J.K.; Atkins, W.M.; Sligar, S.G. Chapter 11—Reconstitution of membrane proteins in phospholipid bilayer nanodiscs. Methods Enzymol. 2009, 464, 211–231. [Google Scholar] [CrossRef] [Green Version]
- Nasr, M.L.; Baptista, D.; Strauss, M.; Sun, Z.-Y.J.; Grigoriu, S.; Huser, S.; Plückthun, A.; Hagn, F.; Walz, T.; Hogle, J.M. Covalently circularized nanodiscs for studying membrane proteins and viral entry. Nat. Methods 2017, 14, 49–52. [Google Scholar] [CrossRef]
- Knowles, T.J.; Finka, R.; Smith, C.; Lin, Y.-P.; Dafforn, T.; Overduin, M. Membrane proteins solubilized intact in lipid containing nanoparticles bounded by styrene maleic acid copolymer. J. Am. Chem. Soc. 2009, 131, 7484–7485. [Google Scholar] [CrossRef] [PubMed]
- Hazell, G.; Arnold, T.; Barker, R.D.; Clifton, L.A.; Steinke, N.-J.; Tognoloni, C.; Edler, K.J. Evidence of lipid exchange in styrene maleic acid lipid particle (SMALP) nanodisc systems. Langmuir 2016, 32, 11845–11853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamshad, M.; Grimard, V.; Idini, I.; Knowles, T.J.; Dowle, M.R.; Schofield, N.; Sridhar, P.; Lin, Y.; Finka, R.; Wheatley, M. Structural analysis of a nanoparticle containing a lipid bilayer used for detergent-free extraction of membrane proteins. Nano Res. 2015, 8, 774–789. [Google Scholar] [CrossRef]
- Vargas, C.; Arenas, R.C.; Frotscher, E.; Keller, S. Nanoparticle self-assembly in mixtures of phospholipids with styrene/maleic acid copolymers or fluorinated surfactants. Nanoscale 2015, 7, 20685–20696. [Google Scholar] [CrossRef] [Green Version]
- Orwick, M.C.; Judge, P.J.; Procek, J.; Lindholm, L.; Graziadei, A.; Engel, A.; Gröbner, G.; Watts, A. Detergent-Free Formation and Physicochemical Characterization of Nanosized Lipid–Polymer Complexes: Lipodisq. Angew. Chem. Int. Ed. 2012, 51, 4653–4657. [Google Scholar] [CrossRef] [PubMed]
- Ravula, T.; Hardin, N.Z.; Ramadugu, S.K.; Cox, S.J.; Ramamoorthy, A. Formation of pH-Resistant Monodispersed Polymer–Lipid Nanodiscs. Angew. Chem. Int. Ed. 2018, 57, 1342–1345. [Google Scholar] [CrossRef] [PubMed]
- Ravula, T.; Ramadugu, S.K.; Di Mauro, G.; Ramamoorthy, A. Bioinspired, Size-Tunable Self-Assembly of Polymer–Lipid Bilayer Nanodiscs. Angew. Chem. 2017, 129, 11624–11628. [Google Scholar] [CrossRef]
- Oluwole, A.O.; Danielczak, B.; Meister, A.; Babalola, J.O.; Vargas, C.; Keller, S. Solubilization of membrane proteins into functional lipid-bilayer nanodiscs using a diisobutylene/maleic acid copolymer. Angew. Chem. Int. Ed. 2017, 56, 1919–1924. [Google Scholar] [CrossRef] [Green Version]
- Anantharamaiah, G.; Jones, J.; Brouillette, C.; Schmidt, C.; Chung, B.H.; Hughes, T.A.; Bhown, A.; Segrest, J. Studies of synthetic peptide analogs of the amphipathic helix. Structure of complexes with dimyristoyl phosphatidylcholine. J. Biol. Chem. 1985, 260, 10248–10255. [Google Scholar] [CrossRef]
- Mishra, V.K.; Anantharamaiah, G.; Segrest, J.P.; Palgunachari, M.N.; Chaddha, M.; Sham, S.S.; Krishna, N.R. Association of a model class A (apolipoprotein) amphipathic α helical peptide with lipid: High resolution NMR studies of peptide· lipid discoidal complexes. J. Biol. Chem. 2006, 281, 6511–6519. [Google Scholar] [CrossRef] [Green Version]
- Epand, R.; Gawish, A.; Iqbal, M.; Gupta, K.; Chen, C.H.; Segrest, J.; Anantharamaiah, G. Studies of synthetic peptide analogs of the amphipathic helix. Effect of charge distribution, hydrophobicity, and secondary structure on lipid association and lecithin: Cholesterol acyltransferase activation. J. Biol. Chem. 1987, 262, 9389–9396. [Google Scholar] [CrossRef]
- Midtgaard, S.R.; Pedersen, M.C.; Kirkensgaard, J.J.K.; Sørensen, K.K.; Mortensen, K.; Jensen, K.J.; Arleth, L. Self-assembling peptides form nanodiscs that stabilize membrane proteins. Soft Matter 2014, 10, 738–752. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Berkamp, S.; Cook, G.A.; Chan, M.K.; Viadiu, H.; Opella, S.J. Nanodiscs versus macrodiscs for NMR of membrane proteins. Biochemistry 2011, 50, 8983–8985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, M. Development of Methods for Study of Membrane Proteins in the Absence of Detergents. Doctoral Dissertation, University of British Columbia, Vancouver, AB, Canada, 2018. [Google Scholar]
- Kondo, H.; Ikeda, K.; Nakano, M. Formation of size-controlled, denaturation-resistant lipid nanodiscs by an amphiphilic self-polymerizing peptide. Colloids Surf. B Biointerfaces 2016, 146, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Lyons, J.A.; Boggild, A.; Nissen, P.; Frauenfeld, J. Saposin-Lipoprotein Scaffolds for Structure Determination of Membrane Transporters. Methods Enzymol. 2017, 594, 85–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flayhan, A.; Mertens, H.D.; Ural-Blimke, Y.; Molledo, M.M.; Svergun, D.I.; Löw, C. Saposin lipid nanoparticles: A highly versatile and modular tool for membrane protein research. Structure 2018, 26, 345–355.e345. [Google Scholar] [CrossRef] [Green Version]
- Kolter, T.; Sandhoff, K. Lysosomal degradation of membrane lipids. FEBS Lett. 2010, 584, 1700–1712. [Google Scholar] [CrossRef] [Green Version]
- Hausler, E.; Fredriksson, K.; Goba, I.; Peters, C.; Raltchev, K.; Sperl, L.; Steiner, A.; Weinkauf, S.; Hagn, F. Quantifying the insertion of membrane proteins into lipid bilayer nanodiscs using a fusion protein strategy. Biochim. Biophys. Acta BioMembr. 2020, 1862, 183190. [Google Scholar] [CrossRef] [PubMed]
- Shirzad-Wasei, N.; van Oostrum, J.; Bovee-Geurts, P.H.; Kusters, L.J.; Bosman, G.J.; DeGrip, W.J. Rapid transfer of overexpressed integral membrane protein from the host membrane into soluble lipid nanodiscs without previous purification. Biol. Chem. 2015, 396, 903–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overduin, M.; Esmaili, M. Native Nanodiscs and the Convergence of Lipidomics, Metabolomics, Interactomics and Proteomics. Appl. Sci. 2019, 9, 1230. [Google Scholar] [CrossRef]
- Fiori, M.C.; Zheng, W.; Kamilar, E.; Simiyu, G.; Altenberg, G.A.; Liang, H. Extraction and reconstitution of membrane proteins into lipid nanodiscs encased by zwitterionic styrene-maleic amide copolymers. Sci. Rep. 2020, 10, 9940. [Google Scholar] [CrossRef]
- Malhotra, K.; Alder, N.N. Reconstitution of Mitochondrial Membrane Proteins into Nanodiscs by Cell-Free Expression. Methods Mol. Biol. 2017, 1567, 155–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyukmanova, E.N.; Shenkarev, Z.O.; Khabibullina, N.F.; Kopeina, G.S.; Shulepko, M.A.; Paramonov, A.S.; Mineev, K.S.; Tikhonov, R.V.; Shingarova, L.N.; Petrovskaya, L.E.; et al. Lipid-protein nanodiscs for cell-free production of integral membrane proteins in a soluble and folded state: Comparison with detergent micelles, bicelles and liposomes. Biochim. Biophys. Acta 2012, 1818, 349–358. [Google Scholar] [CrossRef] [Green Version]
- Roos, C.; Kai, L.; Proverbio, D.; Ghoshdastider, U.; Filipek, S.; Dotsch, V.; Bernhard, F. Co-translational association of cell-free expressed membrane proteins with supplied lipid bilayers. Mol. Membr. Biol. 2013, 30, 75–89. [Google Scholar] [CrossRef]
- Ly, S.; Bourguet, F.; Fischer, N.O.; Lau, E.Y.; Coleman, M.A.; Laurence, T.A. Quantifying interactions of a membrane protein embedded in a lipid nanodisc using fluorescence correlation spectroscopy. Biophys. J. 2014, 106, L05–L08. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, K.; Alder, N.N. Advances in the use of nanoscale bilayers to study membrane protein structure and function. Biotechnol. Genet. Eng. Rev. 2014, 30, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Nasr, M.L.; Singh, S.K. Radioligand binding to nanodisc-reconstituted membrane transporters assessed by the scintillation proximity assay. Biochemistry 2014, 53, 4–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Caaveiro, J.M.; Abe, R.; Katagiri, T.; Tsumoto, K. Catalytic activity of MsbA reconstituted in nanodisc particles is modulated by remote interactions with the bilayer. FEBS Lett. 2011, 585, 3533–3537. [Google Scholar] [CrossRef] [Green Version]
- Gardill, B.; Huang, J.; Tu, L.; Van Petegem, F.; Oxenoid, K.; Thomson, C.A. Nanodisc technology facilitates identification of monoclonal antibodies targeting multi-pass membrane proteins. Sci. Rep. 2020, 10, 1130. [Google Scholar] [CrossRef]
- Swainsbury, D.J.; Scheidelaar, S.; van Grondelle, R.; Killian, J.A.; Jones, M.R. Bacterial reaction centers purified with styrene maleic acid copolymer retain native membrane functional properties and display enhanced stability. Angew. Chem. Int. Ed. Engl. 2014, 53, 11803–11807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swainsbury, D.J.K.; Scheidelaar, S.; Foster, N.; van Grondelle, R.; Killian, J.A.; Jones, M.R. The effectiveness of styrene-maleic acid (SMA) copolymers for solubilisation of integral membrane proteins from SMA-accessible and SMA-resistant membranes. Biochim. Biophys. Acta BioMembr. 2017, 1859, 2133–2143. [Google Scholar] [CrossRef]
- Kalienkova, V.; Alvadia, C.; Clerico Mosina, V.; Paulino, C. Single-Particle Cryo-EM of Membrane Proteins in Lipid Nanodiscs. Methods Mol. Biol. 2020, 2127, 245–273. [Google Scholar] [CrossRef]
- Efremov, R.G.; Gatsogiannis, C.; Raunser, S. Lipid Nanodiscs as a Tool for High-Resolution Structure Determination of Membrane Proteins by Single-Particle Cryo-EM. Methods Enzymol. 2017, 594, 1–30. [Google Scholar] [CrossRef]
- Qiu, W.; Fu, Z.; Xu, G.G.; Grassucci, R.A.; Zhang, Y.; Frank, J.; Hendrickson, W.A.; Guo, Y. Structure and activity of lipid bilayer within a membrane-protein transporter. Proc. Natl. Acad. Sci. USA 2018, 115, 12985–12990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Basak, S.; Rao, S.; Gicheru, Y.; Mayer, M.L.; Sansom, M.S.P.; Chakrapani, S. Mechanisms of activation and desensitization of full-length glycine receptor in lipid nanodiscs. Nat. Commun. 2020, 11, 3752. [Google Scholar] [CrossRef]
- Rheinberger, J.; Gao, X.; Schmidpeter, P.A.; Nimigean, C.M. Ligand discrimination and gating in cyclic nucleotide-gated ion channels from apo and partial agonist-bound cryo-EM structures. Elife 2018, 7, e39775. [Google Scholar] [CrossRef]
- Nygaard, R.; Kim, J.; Mancia, F. Cryo-electron microscopy analysis of small membrane proteins. Curr. Opin. Struct. Biol. 2020, 64, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Tan, Y.Z.; Wicht, K.J.; Erramilli, S.K.; Dhingra, S.K.; Okombo, J.; Vendome, J.; Hagenah, L.M.; Giacometti, S.I.; Warren, A.L.; et al. Structure and drug resistance of the Plasmodium falciparum transporter PfCRT. Nature 2019, 576, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Puthenveetil, R.; Vinogradova, O. Optimization of the design and preparation of nanoscale phospholipid bilayers for its application to solution NMR. Proteins Struct. Funct. Bioinform. 2013, 81, 1222–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kijac, A.Z.; Li, Y.; Sligar, S.G.; Rienstra, C.M. Magic-angle spinning solid-state NMR spectroscopy of nanodisc-embedded human CYP3A4. Biochemistry 2007, 46, 13696–13703. [Google Scholar] [CrossRef] [Green Version]
- Lavington, S.; Watts, A. Lipid nanoparticle technologies for the study of G protein-coupled receptors in lipid environments. Biophys. Rev. 2020, 12, 1287–1302. [Google Scholar] [CrossRef]
- Viennet, T.; Bungert-Plumke, S.; Elter, S.; Viegas, A.; Fahlke, C.; Etzkorn, M. Reconstitution and NMR Characterization of the Ion-Channel Accessory Subunit Barttin in Detergents and Lipid-Bilayer Nanodiscs. Front. Mol. BioSci. 2019, 6, 13. [Google Scholar] [CrossRef]
- Martens, C.; Stein, R.A.; Masureel, M.; Roth, A.; Mishra, S.; Dawaliby, R.; Konijnenberg, A.; Sobott, F.; Govaerts, C.; McHaourab, H.S. Lipids modulate the conformational dynamics of a secondary multidrug transporter. Nat. Struct. Mol. Biol. 2016, 23, 744–751. [Google Scholar] [CrossRef]
- Sahu, I.D.; Dixit, G.; Reynolds, W.D.; Kaplevatsky, R.; Harding, B.D.; Jaycox, C.K.; McCarrick, R.M.; Lorigan, G.A. Characterization of the Human KCNQ1 Voltage Sensing Domain (VSD) in Lipodisq Nanoparticles for Electron Paramagnetic Resonance (EPR) Spectroscopic Studies of Membrane Proteins. J. Phys. Chem. B 2020, 124, 2331–2342. [Google Scholar] [CrossRef] [PubMed]
- Samanta, D.; Widom, J.; Borbat, P.P.; Freed, J.H.; Crane, B.R. Bacterial Energy Sensor Aer Modulates the Activity of the Chemotaxis Kinase CheA Based on the Redox State of the Flavin Cofactor. J. Biol. Chem. 2016, 291, 25809–25814. [Google Scholar] [CrossRef] [Green Version]
- Nath, A.; Trexler, A.J.; Koo, P.; Miranker, A.D.; Atkins, W.M.; Rhoades, E. Single-molecule fluorescence spectroscopy using phospholipid bilayer nanodiscs. Methods Enzymol. 2010, 472, 89–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nath, A.; Koo, P.K.; Rhoades, E.; Atkins, W.M. Allosteric effects on substrate dissociation from cytochrome P450 3A4 in nanodiscs observed by ensemble and single-molecule fluorescence spectroscopy. J. Am. Chem. Soc. 2008, 130, 15746–15747. [Google Scholar] [CrossRef] [Green Version]
- Sadler, E.E.; Kapanidis, A.N.; Tucker, S.J. Solution-Based Single-Molecule FRET Studies of K(+) Channel Gating in a Lipid Bilayer. Biophys. J. 2016, 110, 2663–2670. [Google Scholar] [CrossRef] [Green Version]
- Overduin, M.; Esmaili, M. Structures and Interactions of Transmembrane Targets in Native Nanodiscs. SLAS Discov. 2019, 24, 943–952. [Google Scholar] [CrossRef]
- Bangham, A.D.; Horne, R.W. Negative Staining of Phospholipids and Their Structural Modification by Surface-Active Agents as Observed in the Electron Microscope. J. Mol. Biol. 1964, 8, 660–668. [Google Scholar] [CrossRef]
- Akbarzadeh, A.; Rezaei-Sadabady, R.; Davaran, S.; Joo, S.W.; Zarghami, N.; Hanifehpour, Y.; Samiei, M.; Kouhi, M.; Nejati-Koshki, K. Liposome: Classification, preparation, and applications. Nanoscale Res. Lett. 2013, 8, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sessa, G.; Weissmann, G. Phospholipid spherules (liposomes) as a model for biological membranes. J. Lipid Res. 1968, 9, 310–318. [Google Scholar] [CrossRef]
- Hickey, K.D.; Buhr, M.M. Lipid bilayer composition affects transmembrane protein orientation and function. J. Lipids 2011, 2011, 208457. [Google Scholar] [CrossRef] [Green Version]
- Daraee, H.; Etemadi, A.; Kouhi, M.; Alimirzalu, S.; Akbarzadeh, A. Application of liposomes in medicine and drug delivery. Artif. Cells Nanomed. Biotechnol. 2016, 44, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Österlund, N.; Luo, J.; Wärmländer, S.K.; Gräslund, A. Membrane-mimetic systems for biophysical studies of the amyloid-β peptide. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2019, 1867, 492–501. [Google Scholar] [CrossRef]
- Smirnova, I.A.; Adelroth, P.; Brzezinski, P. Extraction and liposome reconstitution of membrane proteins with their native lipids without the use of detergents. Sci. Rep. 2018, 8, 14950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skrzypek, R.; Iqbal, S.; Callaghan, R. Methods of reconstitution to investigate membrane protein function. Methods 2018, 147, 126–141. [Google Scholar] [CrossRef] [Green Version]
- Johnston, M.J.; Semple, S.C.; Klimuk, S.K.; Ansell, S.; Maurer, N.; Cullis, P.R. Characterization of the drug retention and pharmacokinetic properties of liposomal nanoparticles containing dihydrosphingomyelin. Biochim. Biophys. Acta 2007, 1768, 1121–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, T.M. Liposomes. Opportunities in drug delivery. Drugs 1997, 54 (Suppl. 4), 8–14. [Google Scholar] [CrossRef]
- Parmar, M.M.; Edwards, K.; Madden, T.D. Incorporation of bacterial membrane proteins into liposomes: Factors influencing protein reconstitution. Biochim. Biophys. Acta 1999, 1421, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Winterstein, L.M.; Kukovetz, K.; Rauh, O.; Turman, D.L.; Braun, C.; Moroni, A.; Schroeder, I.; Thiel, G. Reconstitution and functional characterization of ion channels from nanodiscs in lipid bilayers. J. Gen. Physiol. 2018, 150, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.J.; Blaza, J.N.; Bridges, H.R.; May, B.; Moore, A.L.; Hirst, J. A Self-Assembled Respiratory Chain that Catalyzes NADH Oxidation by Ubiquinone-10 Cycling between Complex I and the Alternative Oxidase. Angew. Chem. Int. Ed. 2016, 55, 728–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouritsen, O.G. Model answers to lipid membrane questions. Cold Spring Harb. Perspect. Biol. 2011, 3, a004622. [Google Scholar] [CrossRef]
- Plummer, A.M.; Fleming, K.G. BamA alone accelerates outer membrane protein folding in vitro through a catalytic mechanism. Biochemistry 2015, 54, 6009–6011. [Google Scholar] [CrossRef]
- Teucher, M.; Zhang, H.; Bader, V.; Winklhofer, K.F.; Garcia-Saez, A.J.; Rajca, A.; Bleicken, S.; Bordignon, E. A new perspective on membrane-embedded Bax oligomers using DEER and bioresistant orthogonal spin labels. Sci. Rep. 2019, 9, 13013. [Google Scholar] [CrossRef]
- Sejwal, K.; Chami, M.; Baumgartner, P.; Kowal, J.; Müller, S.A.; Stahlberg, H. Proteoliposomes—A system to study membrane proteins under buffer gradients by cryo-EM. Nanotechnol. Rev. 2017, 6, 57–74. [Google Scholar] [CrossRef]
- Rideau, E.; Dimova, R.; Schwille, P.; Wurm, F.R.; Landfester, K. Liposomes and polymersomes: A comparative review towards cell mimicking. Chem. Soc. Rev. 2018, 47, 8572–8610. [Google Scholar] [CrossRef] [Green Version]
- Angelova, M.I.; Dimitrov, D.S. Liposome electroformation. Faraday Discuss. Chem. Soc. 1986, 81, 303–311. [Google Scholar] [CrossRef]
- Girard, P.; Pecreaux, J.; Lenoir, G.; Falson, P.; Rigaud, J.L.; Bassereau, P. A new method for the reconstitution of membrane proteins into giant unilamellar vesicles. Biophys. J. 2004, 87, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Dorr, J.M.; Koorengevel, M.C.; Schafer, M.; Prokofyev, A.V.; Scheidelaar, S.; van der Cruijsen, E.A.; Dafforn, T.R.; Baldus, M.; Killian, J.A. Detergent-free isolation, characterization, and functional reconstitution of a tetrameric K+ channel: The power of native nanodiscs. Proc. Natl. Acad. Sci. USA 2014, 111, 18607–18612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalise, M.; Pochini, L.; Giangregorio, N.; Tonazzi, A.; Indiveri, C. Proteoliposomes as tool for assaying membrane transporter functions and interactions with xenobiotics. Pharmaceutics 2013, 5, 472–497. [Google Scholar] [CrossRef] [Green Version]
- Zollmann, T.; Moiset, G.; Tumulka, F.; Tampe, R.; Poolman, B.; Abele, R. Single liposome analysis of peptide translocation by the ABC transporter TAPL. Proc. Natl. Acad. Sci. USA 2015, 112, 2046–2051. [Google Scholar] [CrossRef] [Green Version]
- Hresko, R.C.; Kraft, T.E.; Quigley, A.; Carpenter, E.P.; Hruz, P.W. Mammalian Glucose Transporter Activity Is Dependent upon Anionic and Conical Phospholipids. J. Biol. Chem. 2016, 291, 17271–17282. [Google Scholar] [CrossRef] [Green Version]
- Peterson, E.; Ryser, T.; Funk, S.; Inouye, D.; Sharma, M.; Qin, H.; Cross, T.A.; Busath, D.D. Functional reconstitution of influenza A M2(22-62). Biochim. Biophys. Acta 2011, 1808, 516–521. [Google Scholar] [CrossRef] [Green Version]
- Apell, H.J.; Marcus, M.M.; Anner, B.M.; Oetliker, H.; Lauger, P. Optical study of active ion transport in lipid vesicles containing reconstituted Na,K-ATPase. J. Membr. Biol. 1985, 85, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Nimigean, C.M. A radioactive uptake assay to measure ion transport across ion channel-containing liposomes. Nat. Protoc. 2006, 1, 1207–1212. [Google Scholar] [CrossRef]
- Johnson, Z.L.; Lee, S.Y. Liposome reconstitution and transport assay for recombinant transporters. Methods Enzymol. 2015, 556, 373–383. [Google Scholar] [CrossRef]
- Fiori, M.C.; Figueroa, V.; Zoghbi, M.E.; Saez, J.C.; Reuss, L.; Altenberg, G.A. Permeation of calcium through purified connexin 26 hemichannels. J. Biol. Chem. 2012, 287, 40826–40834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreiter, J.; Beitz, E.; Pohl, E.E. A Fluorescence-Based Method to Measure ADP/ATP Exchange of Recombinant Adenine Nucleotide Translocase in Liposomes. Biomolecules 2020, 10, 685. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, G.A.; Terry, D.S.; Warren, A.L.; Quick, M.; Javitch, J.A.; Blanchard, S.C. Quantifying secondary transport at single-molecule resolution. Nature 2019, 575, 528–534. [Google Scholar] [CrossRef]
- Mulligan, C.; Fitzgerald, G.A.; Wang, D.N.; Mindell, J.A. Functional characterization of a Na+-dependent dicarboxylate transporter from Vibrio cholerae. J. Gen. Physiol. 2014, 143, 745–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, R.M.; Compton, E.L.; Mindell, J.A. Functional characterization of a Na+-dependent aspartate transporter from Pyrococcus horikoshii. J. Biol. Chem. 2009, 284, 17540–17548. [Google Scholar] [CrossRef] [Green Version]
- Stokes, D.L.; Rice, W.J.; Hu, M.; Kim, C.; Ubarretxena-Belandia, I. Two-dimensional crystallization of integral membrane proteins for electron crystallography. Methods Mol. Biol. 2010, 654, 187–205. [Google Scholar] [CrossRef] [Green Version]
- Jiko, C.; Davies, K.M.; Shinzawa-Itoh, K.; Tani, K.; Maeda, S.; Mills, D.J.; Tsukihara, T.; Fujiyoshi, Y.; Kuhlbrandt, W.; Gerle, C. Bovine F1Fo ATP synthase monomers bend the lipid bilayer in 2D membrane crystals. Elife 2015, 4, e06119. [Google Scholar] [CrossRef]
- Chadda, R.; Robertson, J.L. Measuring Membrane Protein Dimerization Equilibrium in Lipid Bilayers by Single-Molecule Fluorescence Microscopy. Methods Enzymol. 2016, 581, 53–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, M.; Li, E.; Wimley, W.C.; Hristova, K. Forster resonance energy transfer in liposomes: Measurements of transmembrane helix dimerization in the native bilayer environment. Anal. BioChem. 2005, 340, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, Y.; Deberg, H.A.; Nomura, T.; Hoffman, M.T.; Rohde, P.R.; Schulten, K.; Martinac, B.; Selvin, P.R. Single molecule FRET reveals pore size and opening mechanism of a mechano-sensitive ion channel. Elife 2014, 3, e01834. [Google Scholar] [CrossRef]
- Tribet, C.; Audebert, R.; Popot, J.-L. Amphipols: Polymers that keep membrane proteins soluble in aqueous solutions. Proc. Natl. Acad. Sci. USA 1996, 93, 15047–15050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popot, J.-L.; Althoff, T.; Bagnard, D.; Banères, J.-L.; Bazzacco, P.; Billon-Denis, E.; Catoire, L.; Champeil, P.; Charvolin, D.; Cocco, M. Amphipols from A to Z. Annu. Rev. Biophys. 2011, 40, 379–408. [Google Scholar] [CrossRef] [PubMed]
- Zoonens, M.; Catoire, L.J.; Giusti, F.; Popot, J.-L. NMR study of a membrane protein in detergent-free aqueous solution. Proc. Natl. Acad. Sci. USA 2005, 102, 8893–8898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazzacco, P.; Billon-Denis, E.; Sharma, K.S.; Catoire, L.J.; Mary, S.; Le Bon, C.; Point, E.; Banères, J.-L.; Durand, G.g.; Zito, F. Nonionic homopolymeric amphipols: Application to membrane protein folding, cell-free synthesis, and solution nuclear magnetic resonance. Biochemistry 2012, 51, 1416–1430. [Google Scholar] [CrossRef] [PubMed]
- Zoonens, M.; Popot, J.-L. Amphipols for each season. J. Membr. Biol. 2014, 247, 759–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gohon, Y.; Giusti, F.; Prata, C.; Charvolin, D.; Timmins, P.; Ebel, C.; Tribet, C.; Popot, J.L. Well-defined nanoparticles formed by hydrophobic assembly of a short and polydisperse random terpolymer, amphipol A8-35. Langmuir 2006, 22, 1281–1290. [Google Scholar] [CrossRef] [PubMed]
- Gaffrey, M. A comprehensive review of the lipid cubic phase or in meso method for crystallizing membrane and soluble proteins and complexes. Acta Crystallogr. Sect. F 2015, F71, 3–18. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://cherezov.usc.edu/resources.htm (accessed on 30 August 2021).
- Zhang, H.; Luginina, A.; Mishin, A.; Baidya, M.; Shukla, A.K.; Cherezov, V. Structural insights into ligand recognition and activation of angiotensin receptors. Trends Pharmacol. Sci. 2021, 42, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, D.M.; Cherezov, V.; Hanson, M.A.; Rasmussen, S.G.; Thian, F.S.; Kobilka, T.S.; Choi, H.J.; Yao, X.J.; Weis, W.I.; Stevens, R.C.; et al. GPCR engineering yields high-resolution structural insights into beta2-adrenergic receptor function. Science 2007, 318, 1266–1273. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Caffrey, M. Lipid cubic phase as a membrane mimetic for integral membrane protein enzymes. Proc. Natl. Acad. Sci. USA 2011, 108, 8639–8644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Caffrey, M. Structure and Functional Characterization of Membrane Integral Proteins in the Lipid Cubic Phase. J. Mol. Biol. 2020, 432, 5104–5123. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System/Type | Applicable Techniques to Study IMPs | Advantages | Disadvantages |
| Detergent micelles Ionic detergents Zwitterionic detergents Non-ionic detergents | X-ray crystallography Single-particle cryoEM Solution NMR EPR spectroscopy Fluorescence spectroscopy smFRET Isothermal titration calorimetry (ITC) for ligand binding/protein interactions Functional assays | Easy handling Starting point for downstream applications Availability of large variety of detergents | Propensity of IMP denaturation Chances of non-physiological IMP conformations due to mismatched ‘IMP-micelle’ hydrophobic thicknesses CMC of the detergent must be considered |
| Bicelles | Solution NMR Solid-state NMR X-ray crystallography EPR spectroscopy | Easy preparation Homogeneous and translucent suspensions Provide true lipid environment physiological conditions Diverse types of lipids can be incorporated to match Bicelles of different sizes can be prepared | Total lipid concentration can affect size and geometry of bicelle Risk of IMP perturbation in case of insufficient bilayer size |
| Nanodisc MSP nanodiscs SMALP/Lipodisq® Synthetic peptide-based nanodiscs Saposin nanoparticles | Single particle cryoEM Solution NMR Fluorescence spectroscopy and microscopy smFRET EPR spectroscopy ITC for ligand binding/protein interactions Functional assays | Maintain integrity and shape even upon dilution Easy accessibility of soluble domains in IMPs Possibility of size adjustment to accommodate a monomeric IMP or larger IMP complex | Optimization of assembly conditions can be time consuming Not suitable for large MP oligomers Dynamics of lipids affected by protein ‘belt’ Limited size range |
| Liposomes Small unilamellar vesicles (SUVs) Large unilamellar vesicles (LUVs) Giant unilamellar vesicles (GUVs) Multilamellar vesicles (MLVs) | Electron crystallography Solid-state NMR EPR spectroscopy smFRET Functional assays/substrate uptake Electrophysiology | Large size can accommodate large and multicomponent systems Represent continuous membrane providing closer to native environment for IMPs Diffusion behavior similar to native phospholipid membrane Broad range of possible lipid compositions | The orientation of IMP is often non-native Expensive compared to the traditional systems Low solubility |
| Amphipols | Single-particle cryoEM Solid-state NMR | Assist IMPs study in aqueous environment Stability of IMP-amphipol complex stable on dilution Provides better IMP stability compared to micelle Facilitate refolding of denatured IMPs | Commercially evaluability of only one amphipol type Too difficult to maintain the IMP-amphipol complex sometimes Multivalent cations- and pH-dependent solubility |
| Lipidic cubic phase | X-ray crystallography Functional studies | More native-like environment for IMPs facilitating their crystallization | Relatively expensive |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majeed, S.; Ahmad, A.B.; Sehar, U.; Georgieva, E.R. Lipid Membrane Mimetics in Functional and Structural Studies of Integral Membrane Proteins. Membranes 2021, 11, 685. https://doi.org/10.3390/membranes11090685
Majeed S, Ahmad AB, Sehar U, Georgieva ER. Lipid Membrane Mimetics in Functional and Structural Studies of Integral Membrane Proteins. Membranes. 2021; 11(9):685. https://doi.org/10.3390/membranes11090685
Chicago/Turabian StyleMajeed, Saman, Akram Bani Ahmad, Ujala Sehar, and Elka R. Georgieva. 2021. "Lipid Membrane Mimetics in Functional and Structural Studies of Integral Membrane Proteins" Membranes 11, no. 9: 685. https://doi.org/10.3390/membranes11090685