Why Is Eradicating Typhoid Fever So Challenging: Implications for Vaccine and Therapeutic Design


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Why Is Eradicating Typhoid Fever So Challenging?
2.1. Epidemic of MDR and XDR S. Typhi
2.2. Animal Models for Studying S. Typhi Pathogenesis
2.2.1. Animal Models for Acute Typhoid
2.2.2. Animal Models for Chronic/Persistent Typhoid
2.2.3. Human Infection Studies
2.3. Other Important Factors
3. Pathogenesis of Chronic S. Typhi Infection
3.1. Difference between Typhoid Salmonella and NTS
3.2. S. Typhi Pathogenesis with Emphasis on Chronic/Persistent Infection
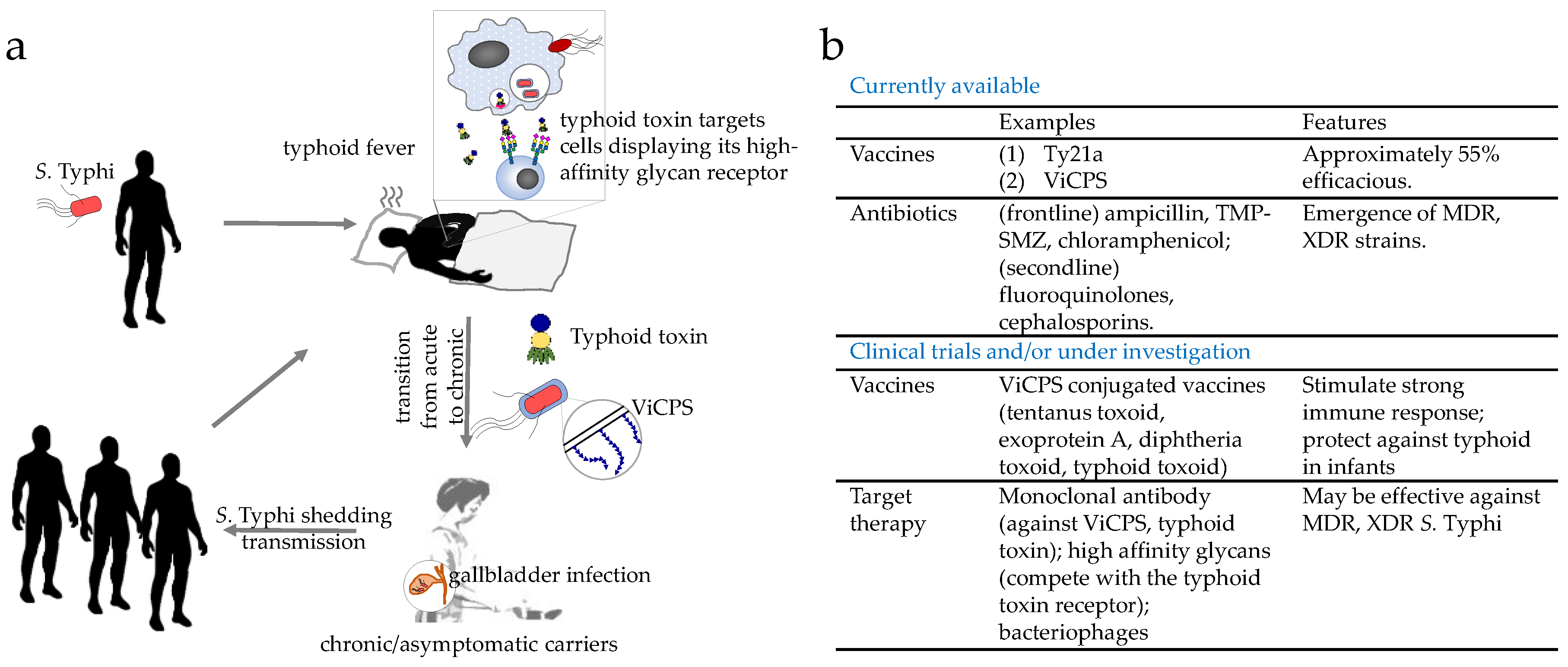
3.2.1. Importance of Understanding S. Typhi Persistence
3.2.2. ViCPS
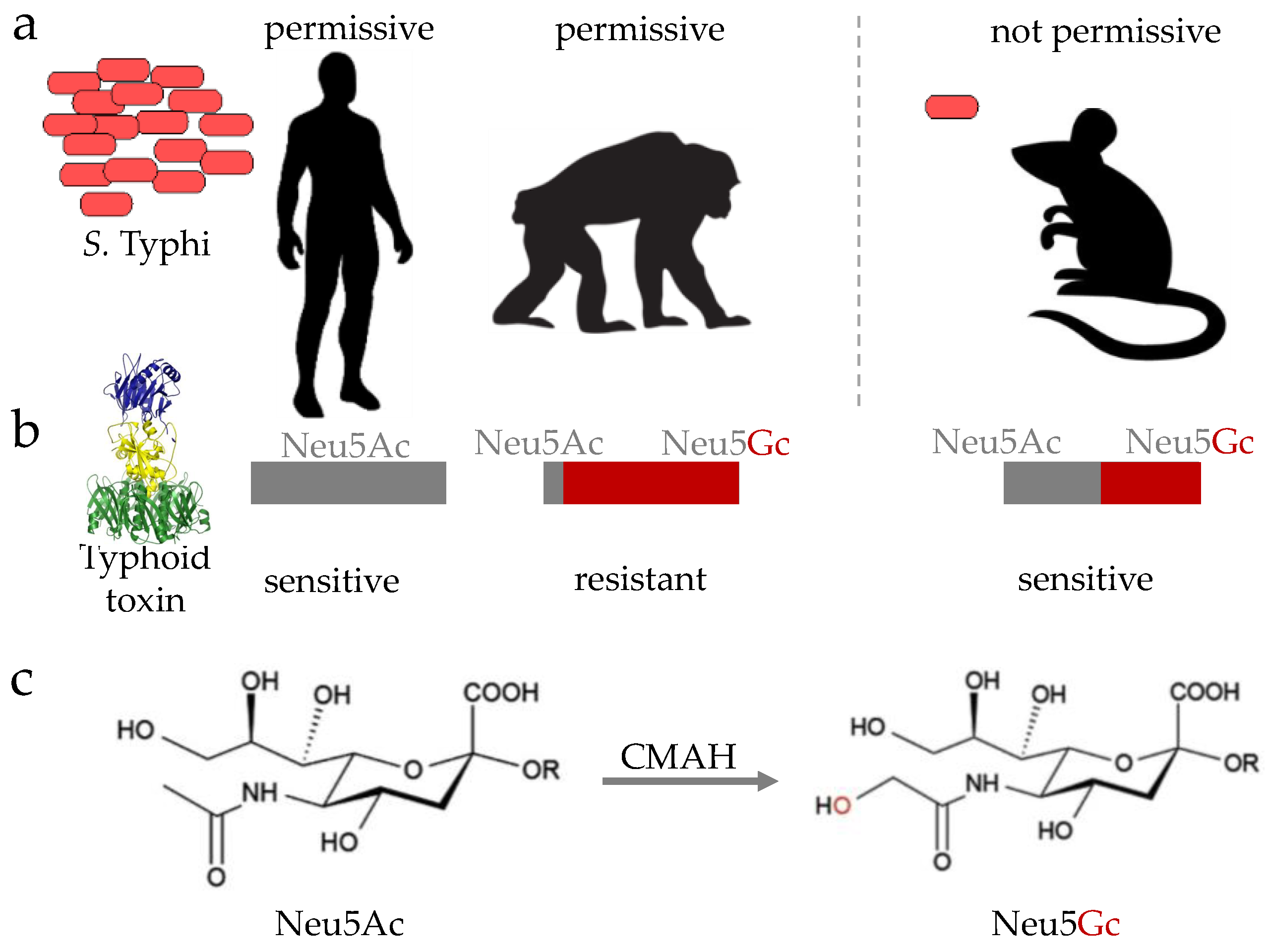
3.2.3. Typhoid Toxin
3.2.4. Gallbladder Infection
4. Vaccines and Therapeutics against S. Typhi Infection
4.1. Typhoid Vaccines
4.1.1. Typhoid Vaccines
4.1.2. Progress on Paratyphoid Vaccine Development
4.2. Limitations of Currently Available Typhoid Vaccines
4.3. Typhoid Treatment Strategies Alternative to Antibiotics
4.3.1. Strategies Targeting the Bacteria
4.3.2. Strategies Mitigating the Action of Secreted Virulence Factors
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brenner, F.W.; Villar, R.G.; Angulo, F.J.; Tauxe, R.; Swaminathan, B. Salmonella nomenclature. J. Clin. Microbiol. 2000, 38, 2465–2467. [Google Scholar] [PubMed]
- Sabbagh, S.C.; Forest, C.G.; Lepage, C.; Leclerc, J.M.; Daigle, F. So similar, yet so different: Uncovering distinctive features in the genomes of Salmonella enterica serovars Typhimurium and Typhi. FEMS Microbiol. Lett. 2010, 305, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gal-Mor, O.; Boyle, E.C.; Grassl, G.A. Same species, different diseases: How and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Front. Microbiol. 2014, 5, 391. [Google Scholar] [CrossRef] [PubMed]
- House, D.; Bishop, A.; Parry, C.; Dougan, G.; Wain, J. Typhoid fever: Pathogenesis and disease. Curr. Opin. Infect. Dis. 2001, 14, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Parry, C.M.; Hien, T.T.; Dougan, G.; White, N.J.; Farrar, J.J. Typhoid fever. N. Engl. J. Med. 2002, 347, 1770–1782. [Google Scholar] [CrossRef] [PubMed]
- Crump, J.A.; Mintz, E.D. Global trends in typhoid and paratyphoid Fever. Clin. Infect. Dis. 2010, 50, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Buckle, G.C.; Walker, C.L.; Black, R.E. Typhoid fever and paratyphoid fever: Systematic review to estimate global morbidity and mortality for 2010. J. Glob. Health 2012, 2. [Google Scholar] [CrossRef]
- Tsolis, R.M.; Xavier, M.N.; Santos, R.L.; Baumler, A.J. How to become a top model: Impact of animal experimentation on human Salmonella disease research. Infect. Immun. 2011, 79, 1806–1814. [Google Scholar] [CrossRef] [PubMed]
- Dougan, G.; Baker, S. Salmonella enterica serovar Typhi and the pathogenesis of typhoid fever. Annu. Rev. Microbiol. 2014, 68, 317–336. [Google Scholar] [CrossRef] [PubMed]
- Chong, A.; Lee, S.; Yang, Y.A.; Song, J. The Role of Typhoid Toxin in Salmonella typhi Virulence. Yale J. Biol. Med. 2017, 90, 283–290. [Google Scholar] [PubMed]
- Galan, J.E. Typhoid toxin provides a window into typhoid fever and the biology of Salmonella typhi. Proc. Natl. Acad. Sci. USA 2016, 113, 6338–6344. [Google Scholar] [CrossRef] [PubMed]
- Lutterloh, E.; Likaka, A.; Sejvar, J.; Manda, R.; Naiene, J.; Monroe, S.S.; Khaila, T.; Chilima, B.; Mallewa, M.; Kampondeni, S.D.; et al. Multidrug-resistant typhoid fever with neurologic findings on the Malawi-Mozambique border. Clin. Infect. Dis. 2012, 54, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Sejvar, J.; Lutterloh, E.; Naiene, J.; Likaka, A.; Manda, R.; Nygren, B.; Monroe, S.; Khaila, T.; Lowther, S.A.; Capewell, L.; et al. Neurologic manifestations associated with an outbreak of typhoid fever, Malawi—Mozambique, 2009: An epidemiologic investigation. PLoS ONE 2012, 7, e46099. [Google Scholar] [CrossRef] [PubMed]
- Crump, J.A.; Luby, S.P.; Mintz, E.D. The global burden of typhoid fever. Bull. World Health Organ. 2004, 82, 346–353. [Google Scholar] [PubMed]
- De Jong, H.K.; Parry, C.M.; van der Vaart, T.W.; Kager, L.M.; van den Ende, S.J.; Maude, R.R.; Wijedoru, L.; Ghose, A.; Hassan, M.U.; Hossain, M.A.; et al. Activation of coagulation and endothelium with concurrent impairment of anticoagulant mechanisms in patients with typhoid fever. J. Infect. 2018, 77, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Tran Vu Thieu, N.; Trinh Van, T.; Tran Tuan, A.; Klemm, E.J.; Nguyen Ngoc Minh, C.; Voong Vinh, P.; Pham Thanh, D.; Ho Ngoc Dan, T.; Pham Duc, T.; Langat, P.; et al. An evaluation of purified Salmonella typhi protein antigens for the serological diagnosis of acute typhoid fever. J. Infect. 2017, 75, 104–114. [Google Scholar] [CrossRef] [PubMed]
- MacFadden, D.R.; Bogoch, I.I.; Andrews, J.R. Advances in diagnosis, treatment, and prevention of invasive Salmonella infections. Curr. Opin. Infect. Dis. 2016, 29, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Monack, D.M. Helicobacter and Salmonella persistent infection strategies. Cold Spring Harb. Perspect. Med. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S.; Marshall, J.M.; Baker, S.; Dongol, S.; Charles, R.C.; Ryan, E.T. Salmonella chronic carriage: Epidemiology, diagnosis, and gallbladder persistence. Trends Microbiol. 2014, 22, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Lovane, L.; Martinez, M.J.; Massora, S.; Mandomando, I.; Ussene, E.; Jordao, D.; Castillo, P.; Ismail, M.R.; Lorenzoni, C.; Carrilho, C.; et al. Carriage prevalence of Salmonella enterica serotype Typhi in gallbladders of adult autopsy cases from Mozambique. J. Infect. Dev. Ctries. 2016, 10, 410–412. [Google Scholar] [CrossRef] [PubMed]
- Monack, D.M.; Mueller, A.; Falkow, S. Persistent bacterial infections: The interface of the pathogen and the host immune system. Nat. Rev. Microbiol. 2004, 2, 747–765. [Google Scholar] [CrossRef] [PubMed]
- Olarte, J.; Galindo, E. Salmonella typhi resistant to chloramphenicol, ampicillin, and other antimicrobial agents: Strains isolated during an extensive typhoid fever epidemic in Mexico. Antimicrob. Agents Chemother. 1973, 4, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Parry, C.M.; Threlfall, E.J. Antimicrobial resistance in typhoidal and nontyphoidal Salmonellae. Curr. Opin. Infect. Dis. 2008, 21, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Feasey, N.A.; Gaskell, K.; Wong, V.; Msefula, C.; Selemani, G.; Kumwenda, S.; Allain, T.J.; Mallewa, J.; Kennedy, N.; Bennett, A.; et al. Rapid emergence of multidrug resistant, H58-lineage Salmonella typhi in Blantyre, Malawi. PLoS Negl. Trop. Dis. 2015, 9, e0003748. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, R.L.; Acosta, C.J.; Danovaro-Holliday, M.C.; Baiqing, D.; Bhattacharya, S.K.; Agtini, M.D.; Bhutta, Z.A.; Canh, D.G.; Ali, M.; Shin, S.; et al. A study of typhoid fever in five Asian countries: Disease burden and implications for controls. Bull. World Health Organ. 2008, 86, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.D.; Serichantalergs, O.; Pitarangsi, C.; Chuanak, N.; Mason, C.J.; Regmi, L.R.; Pandey, P.; Laskar, R.; Shrestha, C.D.; Malla, S. Typhoid fever: A massive, single-point source, multidrug-resistant outbreak in Nepal. Clin. Infect. Dis. 2005, 40, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Chitnis, V.; Chitnis, D.; Verma, S.; Hemvani, N. Multidrug-resistant Salmonella typhi in India. Lancet 1999, 354, 514–515. [Google Scholar] [CrossRef]
- Yan, M.; Li, X.; Liao, Q.; Li, F.; Zhang, J.; Kan, B. The emergence and outbreak of multidrug-resistant typhoid fever in China. Emerg. Microbes Infect. 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Emary, K.; Moore, C.E.; Chanpheaktra, N.; An, K.P.; Chheng, K.; Sona, S.; Duy, P.T.; Nga, T.V.; Wuthiekanun, V.; Amornchai, P.; et al. Enteric fever in Cambodian children is dominated by multidrug-resistant H58 Salmonella enterica serovar Typhi with intermediate susceptibility to ciprofloxacin. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Cooke, F.J.; Wain, J.; Threlfall, E.J. Fluoroquinolone resistance in Salmonella typhi. BMJ 2006, 333, 353–354. [Google Scholar] [CrossRef] [PubMed]
- Klemm, E.J.; Shakoor, S.; Page, A.J.; Qamar, F.N.; Judge, K.; Saeed, D.K.; Wong, V.K.; Dallman, T.J.; Nair, S.; Baker, S.; et al. Emergence of an Extensively Drug-Resistant Salmonella enterica Serovar Typhi Clone Harboring a Promiscuous Plasmid Encoding Resistance to Fluoroquinolones and Third-Generation Cephalosporins. MBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Edsall, G.; Gaines, S.; Landy, M.; Tigertt, W.D.; Sprinz, H.; Trapani, R.J.; Mandel, A.D.; Benenson, A.S. Studies on infection and immunity in experimental typhoid fever. I. Typhoid fever in chimpanzees orally infected with Salmonella typhosa. J. Exp. Med. 1960, 112, 143–166. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Gao, X.; Galan, J.E. Structure and function of the Salmonella typhi chimaeric A(2)B(5) typhoid toxin. Nature 2013, 499, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.H.; Hayakawa, T.; Diaz, S.; Krings, M.; Indriati, E.; Leakey, M.; Paabo, S.; Satta, Y.; Takahata, N.; Varki, A. Inactivation of CMP-N-acetylneuraminic acid hydroxylase occurred prior to brain expansion during human evolution. Proc. Natl. Acad. Sci. USA 2002, 99, 11736–11741. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Song, J.; Gao, X.; Wang, J.; Yu, H.; Chen, X.; Varki, N.; Naito-Matsui, Y.; Galan, J.E.; Varki, A. Host adaptation of a bacterial toxin from the human pathogen Salmonella typhi. Cell 2014, 159, 1290–1299. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.A.; Lee, S.; Zhao, J.; Thompson, A.J.; McBride, R.; Tsogtbaatar, B.; Paulson, J.C.; Nussinov, R.; Deng, L.; Song, J. In vivo tropism of Salmonella typhi toxin to cells expressing a multiantennal glycan receptor. Nat. Microbiol. 2018, 3, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Charles, R.C.; Sheikh, A.; Krastins, B.; Harris, J.B.; Bhuiyan, M.S.; LaRocque, R.C.; Logvinenko, T.; Sarracino, D.A.; Kudva, I.T.; Eisenstein, J.; et al. Characterization of anti-Salmonella enterica serotype Typhi antibody responses in bacteremic Bangladeshi patients by an immunoaffinity proteomics-based technology. Clin. Vaccine Immunol. 2010, 17, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Juarez, S.; Nga, T.V.; Dunstan, S.; Nakajima-Sasaki, R.; Davies, D.H.; McSorley, S.; Baker, S.; Felgner, P.L. Immune profiling with a Salmonella typhi antigen microarray identifies new diagnostic biomarkers of human typhoid. Sci. Rep. 2013, 3, 1043. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, M.; Tangvoranuntakul, P.; Takematsu, H.; Long, J.M.; Housley, G.D.; Kozutsumi, Y.; Suzuki, A.; Wynshaw-Boris, A.; Ryan, A.F.; Gallo, R.L.; et al. N-glycolylneuraminic acid deficiency in mice: Implications for human biology and evolution. Mol. Cell. Biol. 2007, 27, 4340–4346. [Google Scholar] [CrossRef] [PubMed]
- Libby, S.J.; Brehm, M.A.; Greiner, D.L.; Shultz, L.D.; McClelland, M.; Smith, K.D.; Cookson, B.T.; Karlinsey, J.E.; Kinkel, T.L.; Porwollik, S.; et al. Humanized nonobese diabetic-scid IL2rgammanull mice are susceptible to lethal Salmonella typhi infection. Proc. Natl. Acad. Sci. USA 2010, 107, 15589–15594. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, A.D.; Rosenstreich, D.L.; Taylor, B.A. Control of natural resistance to Salmonella typhimurium and Leishmania donovani in mice by closely linked but distinct genetic loci. Nature 1980, 287, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.; Gros, P.; Skamene, E. Natural resistance to infection with intracellular parasites: Molecular genetics identifies Nramp1 as the Bcg/Ity/Lsh locus. J. Leukoc. Biol. 1995, 58, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Loomis, W.P.; Johnson, M.L.; Brasfield, A.; Blanc, M.P.; Yi, J.; Miller, S.I.; Cookson, B.T.; Hajjar, A.M. Temporal and anatomical host resistance to chronic Salmonella infection is quantitatively dictated by Nramp1 and influenced by host genetic background. PLoS ONE 2014, 9, e111763. [Google Scholar] [CrossRef] [PubMed]
- Zaharik, M.L.; Vallance, B.A.; Puente, J.L.; Gros, P.; Finlay, B.B. Host-pathogen interactions: Host resistance factor Nramp1 up-regulates the expression of Salmonella pathogenicity island-2 virulence genes. Proc. Natl. Acad. Sci. USA 2002, 99, 15705–15710. [Google Scholar] [CrossRef] [PubMed]
- Monack, D.M.; Bouley, D.M.; Falkow, S. Salmonella typhimurium persists within macrophages in the mesenteric lymph nodes of chronically infected Nramp1+/+ mice and can be reactivated by IFNgamma neutralization. J. Exp. Med. 2004, 199, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Silva-Herzog, E.; Detweiler, C.S. Intracellular microbes and haemophagocytosis. Cell Microbiol. 2008, 10, 2151–2158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, R.W.; Rosales-Reyes, R.; Ramirez-Aguilar Mde, L.; Chapa-Azuela, O.; Alpuche-Aranda, C.; Gunn, J.S. Gallstones play a significant role in Salmonella spp. gallbladder colonization and carriage. Proc. Natl. Acad. Sci. USA 2010, 107, 4353–4358. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Willinger, T.; Rongvaux, A.; Eynon, E.E.; Stevens, S.; Manz, M.G.; Flavell, R.A.; Galan, J.E. A mouse model for the human pathogen Salmonella typhi. Cell Host Microbe 2010, 8, 369–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darton, T.C.; Blohmke, C.J.; Pollard, A.J. Typhoid epidemiology, diagnostics and the human challenge model. Curr. Opin. Gastroenterol. 2014, 30, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Darton, T.C.; Jones, C.; Blohmke, C.J.; Waddington, C.S.; Zhou, L.; Peters, A.; Haworth, K.; Sie, R.; Green, C.A.; Jeppesen, C.A.; et al. Using a Human Challenge Model of Infection to Measure Vaccine Efficacy: A Randomised, Controlled Trial Comparing the Typhoid Vaccines M01ZH09 with Placebo and Ty21a. PLoS Negl. Trop. Dis. 2016, 10, e0004926. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Gibani, M.M.; Moore, M.; Juel, H.B.; Jones, E.; Meiring, J.; Harris, V.; Gardner, J.; Nebykova, A.; Kerridge, S.A.; et al. Efficacy and immunogenicity of a Vi-tetanus toxoid conjugate vaccine in the prevention of typhoid fever using a controlled human infection model of Salmonella typhi: A randomised controlled, phase 2b trial. Lancet 2017, 390, 2472–2480. [Google Scholar] [CrossRef]
- Feasey, N.A.; Levine, M.M. Typhoid vaccine development with a human challenge model. Lancet 2017, 390, 2419–2421. [Google Scholar] [CrossRef]
- World Health Organization. Typhoid vaccines: WHO position paper, March 2018—Recommendations. Vaccine 2018. [Google Scholar] [CrossRef]
- Dewan, A.M.; Corner, R.; Hashizume, M.; Ongee, E.T. Typhoid Fever and its association with environmental factors in the Dhaka Metropolitan Area of Bangladesh: A spatial and time-series approach. PLoS Negl. Trop. Dis. 2013, 7, e1998. [Google Scholar] [CrossRef] [PubMed]
- Sztein, M.B.; Salerno-Goncalves, R.; McArthur, M.A. Complex adaptive immunity to enteric fevers in humans: Lessons learned and the path forward. Front. Immunol. 2014, 5, 516. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaram, P.; Sahay, B.K.; Rao, N.S. Specific immune responses in typhoid fever & after TAB vaccination. Indian J. Med. Res. 1990, 91, 115–119. [Google Scholar] [PubMed]
- Parkhill, J.; Dougan, G.; James, K.D.; Thomson, N.R.; Pickard, D.; Wain, J.; Churcher, C.; Mungall, K.L.; Bentley, S.D.; Holden, M.T.; et al. Complete genome sequence of a multiple drug resistant Salmonella enterica serovar Typhi CT18. Nature 2001, 413, 848–852. [Google Scholar] [CrossRef] [PubMed]
- McClelland, M.; Sanderson, K.E.; Spieth, J.; Clifton, S.W.; Latreille, P.; Courtney, L.; Porwollik, S.; Ali, J.; Dante, M.; Du, F.; et al. Complete genome sequence of Salmonella enterica serovar Typhimurium LT2. Nature 2001, 413, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Raffatellu, M.; Chessa, D.; Wilson, R.P.; Dusold, R.; Rubino, S.; Baumler, A.J. The Vi capsular antigen of Salmonella enterica serotype Typhi reduces Toll-like receptor-dependent interleukin-8 expression in the intestinal mucosa. Infect. Immun. 2005, 73, 3367–3374. [Google Scholar] [CrossRef] [PubMed]
- Wangdi, T.; Lee, C.Y.; Spees, A.M.; Yu, C.; Kingsbury, D.D.; Winter, S.E.; Hastey, C.J.; Wilson, R.P.; Heinrich, V.; Baumler, A.J. The Vi capsular polysaccharide enables Salmonella enterica serovar typhi to evade microbe-guided neutrophil chemotaxis. PLoS Pathog. 2014, 10, e1004306. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.A.; Wiedmann, M. The Cytolethal Distending Toxin Produced by Nontyphoidal Salmonella Serotypes Javiana, Montevideo, Oranienburg, and Mississippi Induces DNA Damage in a Manner Similar to That of Serotype Typhi. MBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Galan, J.E.; Lara-Tejero, M.; Marlovits, T.C.; Wagner, S. Bacterial type III secretion systems: Specialized nanomachines for protein delivery into target cells. Annu. Rev. Microbiol. 2014, 68, 415–438. [Google Scholar] [CrossRef] [PubMed]
- Diard, M.; Hardt, W.D. Basic Processes in Salmonella-Host Interactions: Within-Host Evolution and the Transmission of the Virulent Genotype. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Jennings, E.; Thurston, T.L.M.; Holden, D.W. Salmonella SPI-2 Type III Secretion System Effectors: Molecular Mechanisms and Physiological Consequences. Cell Host Microbe 2017, 22, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Liss, V.; Hensel, M. Take the tube: Remodelling of the endosomal system by intracellular Salmonella enterica. Cell Microbiol. 2015, 17, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Van der Heijden, J.; Finlay, B.B. Type III effector-mediated processes in Salmonella infection. Future Microbiol. 2012, 7, 685–703. [Google Scholar] [CrossRef] [PubMed]
- Agbor, T.A.; McCormick, B.A. Salmonella effectors: Important players modulating host cell function during infection. Cell Microbiol. 2011, 13, 1858–1869. [Google Scholar] [CrossRef] [PubMed]
- Fass, E.; Groisman, E.A. Control of Salmonella pathogenicity island-2 gene expression. Curr. Opin. Microbiol. 2009, 12, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Ellermeier, J.R.; Slauch, J.M. Adaptation to the host environment: Regulation of the SPI1 type III secretion system in Salmonella enterica serovar Typhimurium. Curr. Opin. Microbiol. 2007, 10, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Knodler, L.A.; Steele-Mortimer, O. Taking possession: Biogenesis of the Salmonella-containing vacuole. Traffic 2003, 4, 587–599. [Google Scholar] [CrossRef] [PubMed]
- LaRock, D.L.; Chaudhary, A.; Miller, S.I. Salmonellae interactions with host processes. Nat. Rev. Microbiol. 2015, 13, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Forest, C.G.; Ferraro, E.; Sabbagh, S.C.; Daigle, F. Intracellular survival of Salmonella enterica serovar Typhi in human macrophages is independent of Salmonella pathogenicity island (SPI)-2. Microbiology 2010, 156, 3689–3698. [Google Scholar] [CrossRef] [PubMed]
- Sabbagh, S.C.; Lepage, C.; McClelland, M.; Daigle, F. Selection of Salmonella enterica serovar Typhi genes involved during interaction with human macrophages by screening of a transposon mutant library. PLoS ONE 2012, 7, e36643. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Morales, F. Impact of Salmonella enterica Type III Secretion System Effectors on the Eukaryotic Host Cell. ISRN Cell Biol. 2012, 2012. [Google Scholar] [CrossRef]
- Spano, S.; Galan, J.E. A Rab32-dependent pathway contributes to Salmonella typhi host restriction. Science 2012, 338, 960–963. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Galan, J.E. A Salmonella protein antagonizes Rac-1 and Cdc42 to mediate host-cell recovery after bacterial invasion. Nature 1999, 401, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Byrne, A.; Berger, C.N.; Klemm, E.; Crepin, V.F.; Dougan, G.; Frankel, G. The Type III Secretion System Effector SptP of Salmonella enterica Serovar Typhi. J. Bacteriol. 2017, 199. [Google Scholar] [CrossRef] [PubMed]
- Darton, T.C.; Zhou, L.; Blohmke, C.J.; Jones, C.; Waddington, C.S.; Baker, S.; Pollard, A.J. Blood culture-PCR to optimise typhoid fever diagnosis after controlled human infection identifies frequent asymptomatic cases and evidence of primary bacteraemia. J. Infect. 2017, 74, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, S.; Ahirwar, S.K.; Nath, G. Proteomics-based identification of plasma proteins and their association with the host-pathogen interaction in chronic typhoid carriers. Int. J. Infect. Dis. 2014, 19, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Charles, R.C.; Sultana, T.; Alam, M.M.; Yu, Y.; Wu-Freeman, Y.; Bufano, M.K.; Rollins, S.M.; Tsai, L.; Harris, J.B.; LaRocque, R.C.; et al. Identification of immunogenic Salmonella enterica serotype Typhi antigens expressed in chronic biliary carriers of S. Typhi in Kathmandu, Nepal. PLoS Negl. Trop. Dis. 2013, 7, e2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.H.; Cho, H.; Park, M.Y.; Kim, Y.S.; Moon, H.B.; Kim, Y.K.; Pai, C.H. Detection of the H1-j strain of Salmonella typhi among Korean isolates by the polymerase chain reaction. Am. J. Trop. Med. Hyg. 1994, 50, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhou, Y.; Bao, H.; Liu, H.W. Vi antigen biosynthesis in Salmonella typhi: Characterization of UDP-N-acetylglucosamine C-6 dehydrogenase (TviB) and UDP-N-acetylglucosaminuronic acid C-4 epimerase (TviC). Biochemistry 2006, 45, 8163–8173. [Google Scholar] [CrossRef] [PubMed]
- Nolan, C.M.; White, P.C., Jr.; Feeley, J.C.; Hambie, E.A.; Brown, S.L.; Wong, K.H. Vi serology in the detection of typhoid carriers. Lancet 1981, 1, 583–585. [Google Scholar] [CrossRef]
- Nolan, C.M.; Feeley, J.C.; White, P.C.; Hambie, E.A., Jr.; Brown, S.L.; Wong, K.H. Evaluation of a new assay for Vi antibody in chronic carriers of Salmonella typhi. J. Clin. Microbiol. 1980, 12, 22–26. [Google Scholar] [PubMed]
- Winter, S.E.; Winter, M.G.; Thiennimitr, P.; Gerriets, V.A.; Nuccio, S.P.; Russmann, H.; Baumler, A.J. The TviA auxiliary protein renders the Salmonella enterica serotype Typhi RcsB regulon responsive to changes in osmolarity. Mol. Microbiol. 2009, 74, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Santander, J.; Roland, K.L.; Curtiss, R., 3rd. Regulation of Vi capsular polysaccharide synthesis in Salmonella enterica serotype Typhi. J. Infect. Dev. Ctries. 2008, 2, 412–420. [Google Scholar] [PubMed]
- Tacket, C.O.; Pasetti, M.F.; Sztein, M.B.; Livio, S.; Levine, M.M. Immune responses to an oral typhoid vaccine strain that is modified to constitutively express Vi capsular polysaccharide. J. Infect. Dis. 2004, 190, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Looney, R.J.; Steigbigel, R.T. Role of the Vi antigen of Salmonella typhi in resistance to host defense in vitro. J. Lab. Clin. Med. 1986, 108, 506–516. [Google Scholar] [PubMed]
- Fowler, C.C.; Chang, S.J.; Gao, X.; Geiger, T.; Stack, G.; Galan, J.E. Emerging insights into the biology of typhoid toxin. Curr. Opin. Microbiol. 2017, 35, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Haghjoo, E.; Galan, J.E. Salmonella typhi encodes a functional cytolethal distending toxin that is delivered into host cells by a bacterial-internalization pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 4614–4619. [Google Scholar] [CrossRef] [PubMed]
- Spano, S.; Ugalde, J.E.; Galan, J.E. Delivery of a Salmonella typhi exotoxin from a host intracellular compartment. Cell Host Microbe 2008, 3, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.J.; Song, J.; Galan, J.E. Receptor-Mediated Sorting of Typhoid Toxin during Its Export from Salmonella typhi-Infected Cells. Cell Host Microbe 2016, 20, 682–689. [Google Scholar] [CrossRef] [PubMed]
- Raffatellu, M.; Wilson, R.P.; Winter, S.E.; Baumler, A.J. Clinical pathogenesis of typhoid fever. J. Infect. Dev. Ctries. 2008, 2, 260–266. [Google Scholar] [PubMed]
- Sztein, M.B. Cell-mediated immunity and antibody responses elicited by attenuated Salmonella enterica Serovar Typhi strains used as live oral vaccines in humans. Clin. Infect. Dis. 2007, 45, S15–S19. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Ravenhall, M.; Pickard, D.; Dougan, G.; Byrne, A.; Frankel, G. Comparison of Salmonella enterica Serovars Typhi and Typhimurium Reveals Typhoidal Serovar-Specific Responses to Bile. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [PubMed]
- Milligan, R.; Paul, M.; Richardson, M.; Neuberger, A. Vaccines for preventing typhoid fever. Cochrane Database Syst. Rev. 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Amicizia, D.; Arata, L.; Zangrillo, F.; Panatto, D.; Gasparini, R. Overview of the impact of Typhoid and Paratyphoid fever. Utility of Ty21a vaccine (Vivotif(R)). J. Prev. Med. Hyg. 2017, 58, E1–E8. [Google Scholar] [PubMed]
- Mastroeni, P.; Chabalgoity, J.A.; Dunstan, S.J.; Maskell, D.J.; Dougan, G. Salmonella: Immune responses and vaccines. Vet. J. 2001, 161, 132–164. [Google Scholar] [CrossRef] [PubMed]
- Germanier, R.; Fuer, E. Isolation and characterization of Gal E mutant Ty 21a of Salmonella typhi: A candidate strain for a live, oral typhoid vaccine. J. Infect. Dis. 1975, 131, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Cisar, J.O.; Poly, F.; Yang, J.; Albanese, J.; Dharmasena, M.; Wai, T.; Guerry, P.; Kopecko, D.J. Genome Sequence of Salmonella enterica Serovar Typhi Oral Vaccine Strain Ty21a. Genome Announc. 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Zuckerman, J.N.; Hatz, C.; Kantele, A. Review of current typhoid fever vaccines, cross-protection against paratyphoid fever, and the European guidelines. Expert Rev. Vaccines 2017, 16, 1029–1043. [Google Scholar] [CrossRef] [PubMed]
- Fraser, A.; Paul, M.; Goldberg, E.; Acosta, C.J.; Leibovici, L. Typhoid fever vaccines: Systematic review and meta-analysis of randomised controlled trials. Vaccine 2007, 25, 7848–7857. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Pollard, A.J. Seroefficacy of Vi Polysaccharide-Tetanus Toxoid Typhoid Conjugate Vaccine (Typbar TCV). Clin. Infect. Dis. 2018, 67, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.; Lee, C.K.; Ahn, C.; Shin, J.; Knezevic, I. Working Group on quality, safety and efficacy of typhoid Vi capsular polysaccharide conjugate, vaccines, Jeju, Republic of Korea, 5–7 September 2012. Vaccine 2013, 31, 4466–4469. [Google Scholar] [CrossRef] [PubMed]
- Szu, S.C. Development of Vi conjugate—A new generation of typhoid vaccine. Expert Rev. Vaccines 2013, 12, 1273–1286. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.Y.; Ho, V.A.; Khiem, H.B.; Trach, D.D.; Bay, P.V.; Thanh, T.C.; Kossaczka, Z.; Bryla, D.A.; Shiloach, J.; Robbins, J.B.; et al. The efficacy of a Salmonella typhi Vi conjugate vaccine in two-to-five-year-old children. N. Engl. J. Med. 2001, 344, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Bhutta, Z.A.; Capeding, M.R.; Bavdekar, A.; Marchetti, E.; Ariff, S.; Soofi, S.B.; Anemona, A.; Habib, M.A.; Alberto, E.; Juvekar, S.; et al. Immunogenicity and safety of the Vi-CRM197 conjugate vaccine against typhoid fever in adults, children, and infants in south and southeast Asia: Results from two randomised, observer-blind, age de-escalation, phase 2 trials. Lancet Infect. Dis. 2014, 14, 119–129. [Google Scholar] [CrossRef]
- Park, S.E.; Marks, F. A conjugate vaccine against typhoid fever. Lancet Infect. Dis. 2014, 14, 90–91. [Google Scholar] [CrossRef]
- Gat, O.; Galen, J.E.; Tennant, S.; Simon, R.; Blackwelder, W.C.; Silverman, D.J.; Pasetti, M.F.; Levine, M.M. Cell-associated flagella enhance the protection conferred by mucosally-administered attenuated Salmonella Paratyphi A vaccines. PLoS Negl. Trop. Dis. 2011, 5, e1373. [Google Scholar] [CrossRef] [PubMed]
- Konadu, E.; Shiloach, J.; Bryla, D.A.; Robbins, J.B.; Szu, S.C. Synthesis, characterization, and immunological properties in mice of conjugates composed of detoxified lipopolysaccharide of Salmonella paratyphi A bound to tetanus toxoid with emphasis on the role of O acetyls. Infect. Immun. 1996, 64, 2709–2715. [Google Scholar] [PubMed]
- Micoli, F.; Rondini, S.; Gavini, M.; Lanzilao, L.; Medaglini, D.; Saul, A.; Martin, L.B. O:2-CRM(197) conjugates against Salmonella Paratyphi A. PLoS ONE 2012, 7, e47039. [Google Scholar] [CrossRef] [PubMed]
- Kothari, S.; Kim, J.A.; Kothari, N.; Jones, C.; Choe, W.S.; Carbis, R. Purification of O-specific polysaccharide from lipopolysaccharide produced by Salmonella enterica serovar Paratyphi A. Vaccine 2014, 32, 2457–2462. [Google Scholar] [CrossRef] [PubMed]
- Saylor, C.; Dadachova, E.; Casadevall, A. Monoclonal antibody-based therapies for microbial diseases. Vaccine 2009, 27, G38–G46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casadevall, A.; Dadachova, E.; Pirofski, L.A. Passive antibody therapy for infectious diseases. Nat. Rev. Microbiol. 2004, 2, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Pfarr, D.S.; Losonsky, G.A.; Kiener, P.A. Immunoprophylaxis of RSV infection: Advancing from RSV-IGIV to palivizumab and motavizumab. Curr. Top. Microbiol. Immunol. 2008, 317, 103–123. [Google Scholar] [PubMed]
- Singh, S.P.; Williams, Y.U.; Benjamin, W.H.; Klebba, P.E.; Boyd, D. Immunoprotection by monoclonal antibodies to the porins and lipopolysaccharide of Salmonella typhimurium. Microb. Pathog. 1996, 21, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Carlin, N.I.; Svenson, S.B.; Lindberg, A.A. Role of monoclonal O-antigen antibody epitope specificity and isotype in protection against experimental mouse typhoid. Microb. Pathog. 1987, 2, 171–183. [Google Scholar] [CrossRef]
- Goh, Y.S.; Clare, S.; Micoli, F.; Saul, A.; Mastroeni, P.; MacLennan, C.A. Monoclonal Antibodies of a Diverse Isotype Induced by an O-Antigen Glycoconjugate Vaccine Mediate In Vitro and In Vivo Killing of African Invasive Nontyphoidal Salmonella. Infect. Immun. 2015, 83, 3722–3731. [Google Scholar] [CrossRef] [PubMed]
- Desranleau, J.M. Progress in the treatment of typhoid fever with Vi bacteriophages. Can. J. Public Health 1949, 40, 473–478. [Google Scholar] [PubMed]
- Kakabadze, E.; Makalatia, K.; Grdzelishvili, N.; Bakuradze, N.; Goderdzishvili, M.; Kusradze, I.; Phoba, M.F.; Lunguya, O.; Lood, C.; Lavigne, R.; et al. Selection of Potential Therapeutic Bacteriophages that Lyse a CTX-M-15 Extended Spectrum beta-Lactamase Producing Salmonella enterica Serovar Typhi Strain from the Democratic Republic of the Congo. Viruses 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Serna, A.; Boedeker, E.C. Pathogenesis and treatment of Shiga toxin-producing Escherichia coli infections. Curr. Opin. Gastroenterol. 2008, 24, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Bitzan, M.; Poole, R.; Mehran, M.; Sicard, E.; Brockus, C.; Thuning-Roberson, C.; Riviere, M. Safety and pharmacokinetics of chimeric anti-Shiga toxin 1 and anti-Shiga toxin 2 monoclonal antibodies in healthy volunteers. Antimicrob. Agents Chemother. 2009, 53, 3081–3087. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, J.M.; Mukherjee, J.; Leysath, C.E.; Debatis, M.; Ofori, K.; Baldwin, K.; Boucher, C.; Peters, R.; Beamer, G.; Sheoran, A.; et al. A single VHH-based toxin-neutralizing agent and an effector antibody protect mice against challenge with Shiga toxins 1 and 2. Infect. Immun. 2013, 81, 4592–4603. [Google Scholar] [CrossRef] [PubMed]
- Sepulveda, J.; Mukherjee, J.; Tzipori, S.; Simpson, L.L.; Shoemaker, C.B. Efficient serum clearance of botulinum neurotoxin achieved using a pool of small antitoxin binding agents. Infect. Immun. 2010, 78, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Moayeri, M.; Tremblay, J.M.; Debatis, M.; Dmitriev, I.P.; Kashentseva, E.A.; Yeh, A.J.; Cheung, G.Y.; Curiel, D.T.; Leppla, S.; Shoemaker, C.B. Adenoviral Expression of a Bispecific VHH-Based Neutralizing Agent That Targets Protective Antigen Provides Prophylactic Protection from Anthrax in Mice. Clin. Vaccine Immunol. 2016, 23, 213–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitov, P.I.; Sadowska, J.M.; Mulvey, G.; Armstrong, G.D.; Ling, H.; Pannu, N.S.; Read, R.J.; Bundle, D.R. Shiga-like toxins are neutralized by tailored multivalent carbohydrate ligands. Nature 2000, 403, 669–672. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Rademacher, C.; Nycholat, C.M.; Futakawa, S.; Lemme, K.; Ernst, B.; Paulson, J.C. High affinity sialoside ligands of myelin associated glycoprotein. Bioorg. Med. Chem. Lett. 2011, 21, 5045–5049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, S.; Linstedt, A.D. Retrograde trafficking of AB(5) toxins: Mechanisms to therapeutics. J. Mol. Med. 2013, 91, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Saenz, J.B.; Doggett, T.A.; Haslam, D.B. Identification and characterization of small molecules that inhibit intracellular toxin transport. Infect. Immun. 2007, 75, 4552–4561. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.-A.; Chong, A.; Song, J. Why Is Eradicating Typhoid Fever So Challenging: Implications for Vaccine and Therapeutic Design. Vaccines 2018, 6, 45. https://doi.org/10.3390/vaccines6030045
Yang Y-A, Chong A, Song J. Why Is Eradicating Typhoid Fever So Challenging: Implications for Vaccine and Therapeutic Design. Vaccines. 2018; 6(3):45. https://doi.org/10.3390/vaccines6030045
Chicago/Turabian StyleYang, Yi-An, Alexander Chong, and Jeongmin Song. 2018. "Why Is Eradicating Typhoid Fever So Challenging: Implications for Vaccine and Therapeutic Design" Vaccines 6, no. 3: 45. https://doi.org/10.3390/vaccines6030045