
Characterization of the Burkholderia cenocepacia TonB Mutant as a Potential Live Attenuated Vaccine


Abstract
:1. Introduction
2. Material and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Growth Kinetics Assay
2.3. DNA, PCR and Cloning Methods
2.4. pGPTCR Plasmid Construction
2.5. Construction of the GAP001 Strain
2.6. Iron Utilization Assay
2.7. Siderophore Secretion Assay
2.8. In Vitro Survival Assays
2.9. In Vivo Survival Study
2.10. Vaccine Study
2.11. Specific Immunoglobulin Analysis
3. Results
3.1. tonB Mutant Generation
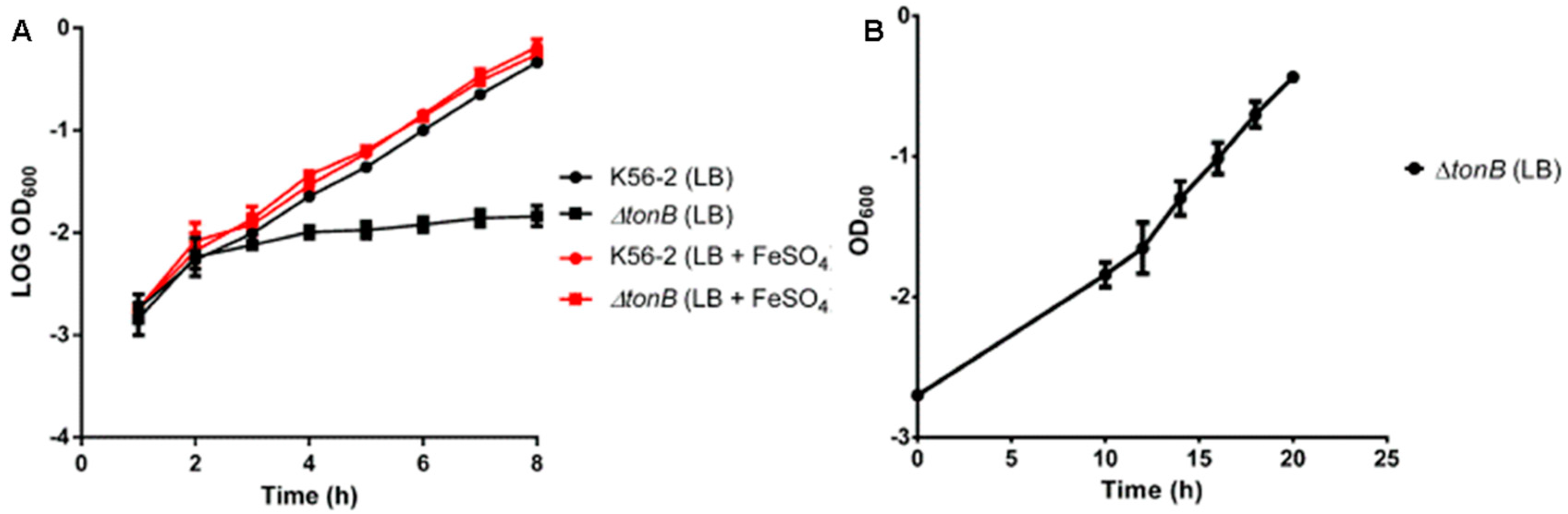
3.2. GAP001 Iron Utilization and Growth Characterization
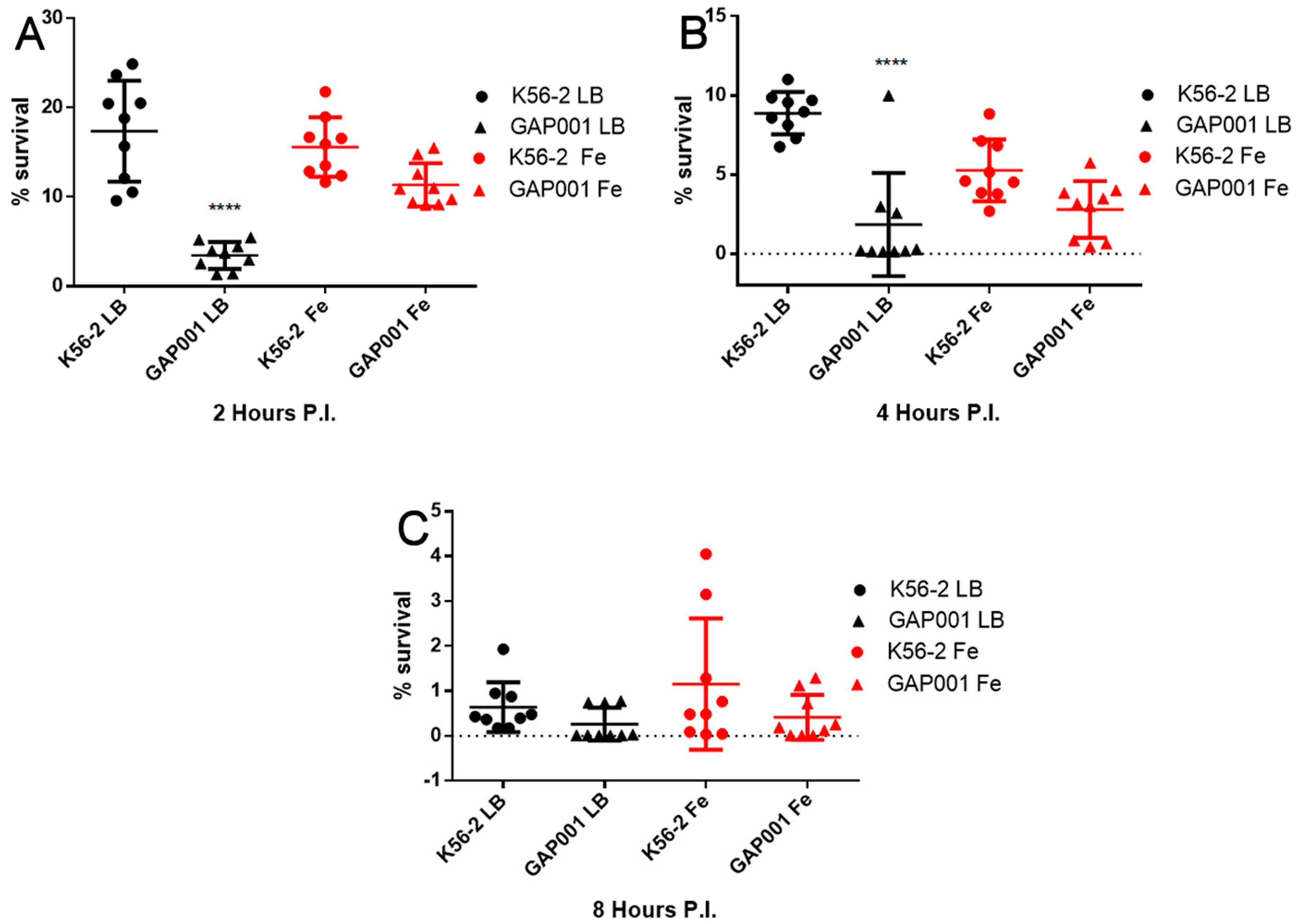
3.3. GAP001 Intracellular Uptake and Survival Characterization
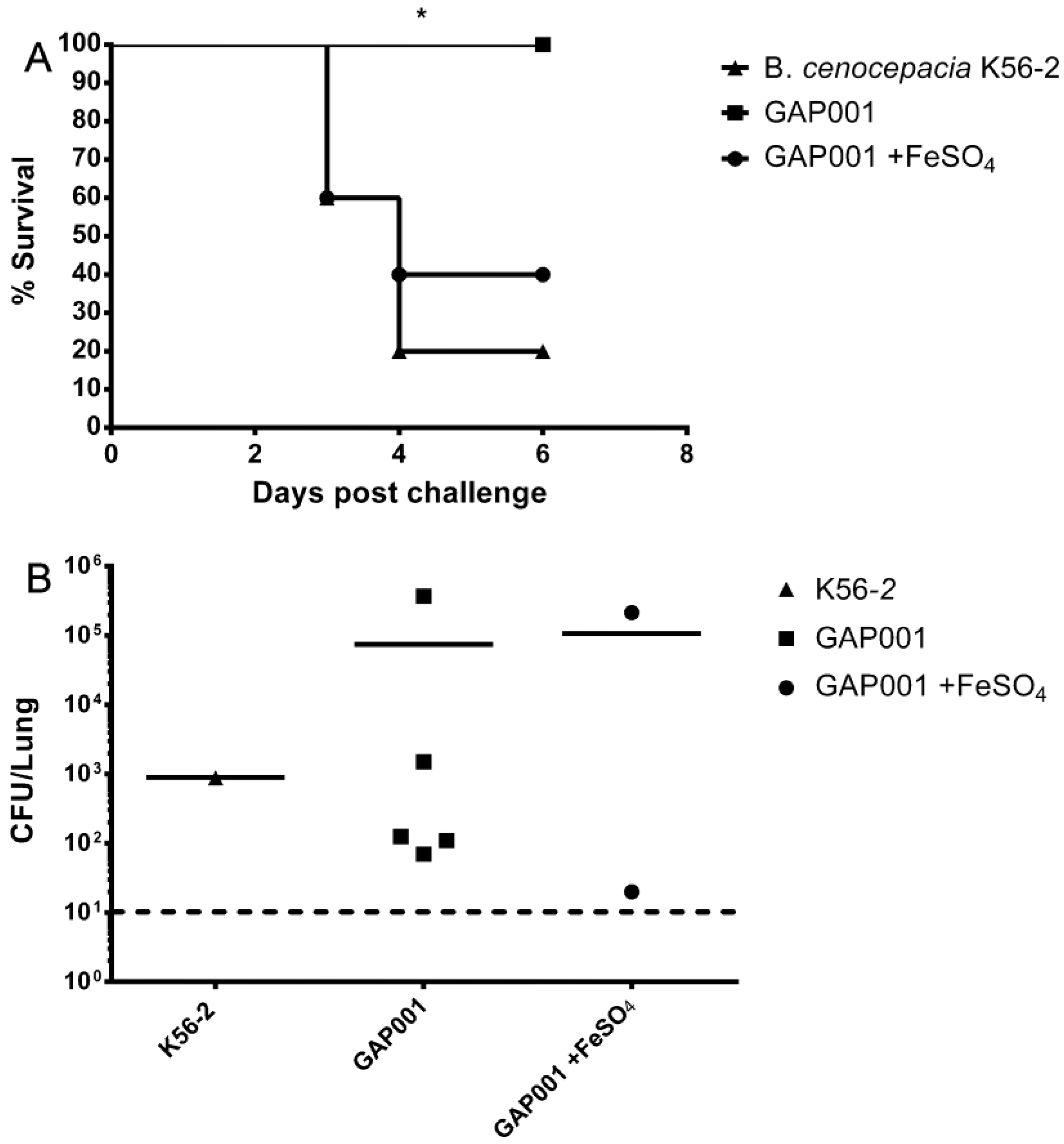
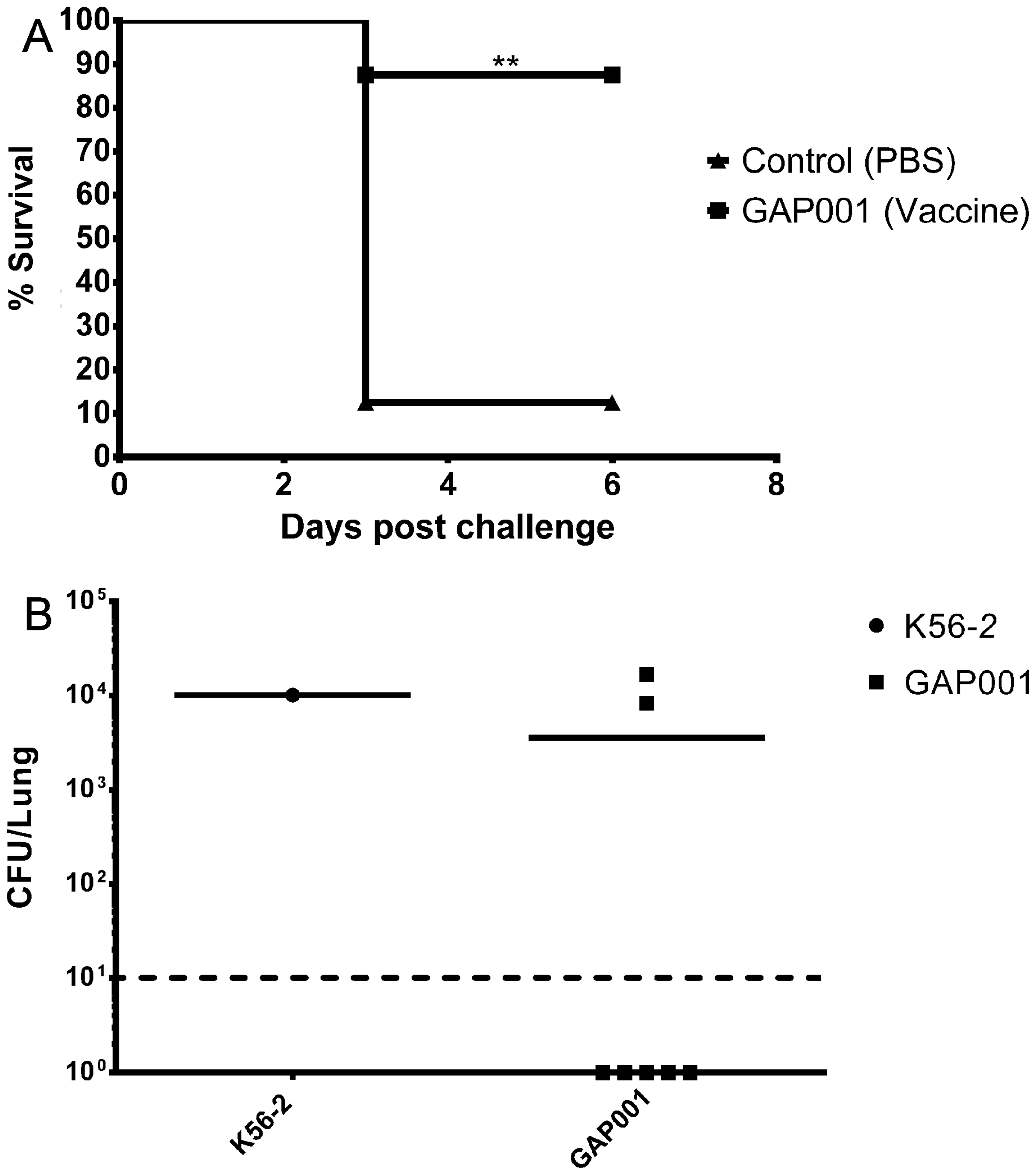
3.4. GAP001 In Vivo Characterization
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Coenye, T.; Vandamme, P. Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ. Microbiol. 2003, 5, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, P.; Dawyndt, P. Classification and identification of the Burkholderia cepacia complex: Past, present and future. Syst. Appl. Microbiol. 2011, 34, 87–95. [Google Scholar]
- LiPuma, J.J. Assessing Airway Microbiota in Cystic Fibrosis: What More Should Be Done? J. Clin. Microbiol. 2015, 53, 2006–2007. [Google Scholar] [CrossRef] [PubMed]
- Pradenas, G.A.; Ross, B.N.; Torres, A.G. Burkholderia cepacia Complex Vaccines: Where Do We Go from here? Vaccines (Basel) 2016, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Courtney, J.M.; Dunbar, K.E.; McDowell, A.; Moore, J.E.; Warke, T.J.; Stevenson, M.; Elborn, J.S. Clinical outcome of Burkholderia cepacia complex infection in cystic fibrosis adults. J. Cyst. Fibros. 2004, 3, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Loutet, S.A.; Valvano, M.A. A decade of Burkholderia cenocepacia virulence determinant research. Infect. Immun. 2010, 78, 4088–4100. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Sajjan, U.S. Host evasion by Burkholderia cenocepacia. Front. Cell Infect. Microbiol. 2011, 1, 25. [Google Scholar] [CrossRef] [PubMed]
- Saldias, M.S.; Valvano, M.A. Interactions of Burkholderia cenocepacia and other Burkholderia cepacia complex bacteria with epithelial and phagocytic cells. Microbiology 2009, 155, 2809–2817. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.R.; Torres, A.G. The art of persistence-the secrets to Burkholderia chronic infections. Pathog. Dis. 2016. [Google Scholar] [CrossRef] [PubMed]
- Bertot, G.M.; Restelli, M.A.; Galanternik, L.; Urey, R.C.A.; Valvano, M.A.; Grinstein, S. Nasal immunization with Burkholderia multivorans outer membrane proteins and the mucosal adjuvant adamantylamide dipeptide confers efficient protection against experimental lung infections with B. multivorans and B. cenocepacia. Infect. Immun. 2007, 75, 2740–2752. [Google Scholar] [CrossRef] [PubMed]
- McClean, S.; Healy, M.E.; Collins, C.; Carberry, S.; O’Shaughnessy, L.; Dennehy, R.; Adams, Á.; Kennelly, H.; Corbett, J.M.; Carty, F.; et al. Linocin and OmpW Are Involved in Attachment of the Cystic Fibrosis-Associated Pathogen Burkholderia cepacia Complex to Lung Epithelial Cells and Protect Mice against Infection. Infect. Immun. 2016, 84, 1424–1437. [Google Scholar] [CrossRef] [PubMed]
- Makidon, P.E.; Knowlton, J.; Groom, J.V., 2nd; Blanco, L.P.; LiPuma, J.J.; Bielinska, A.U.; Baker, J.R., Jr. Induction of immune response to the 17 kDa OMPA Burkholderia cenocepacia polypeptide and protection against pulmonary infection in mice after nasal vaccination with an OMP nanoemulsion-based vaccine. Med. Microbiol. Immunol. 2010, 199, 81–92. [Google Scholar] [PubMed]
- Hatcher, C.L.; Muruato, L.A.; Torres, A.G. Recent Advances in Burkholderia mallei and B. pseudomallei Research. Curr. Trop. Med. Rep. 2015, 2, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Mott, T.M.; Vijayakumar, S.; Sbrana, E.; Endsley, J.J.; Torres, A.G. Characterization of the Burkholderia mallei tonB mutant and Its potential as a backbone strain for vaccine development. PLoS Negl. Trop. Dis. 2015, 9, e0003863. [Google Scholar] [CrossRef] [PubMed]
- Skaar, E.P. The battle for iron between bacterial pathogens and their vertebrate hosts. PLoS Pathog. 2010, 6, e1000949. [Google Scholar] [CrossRef] [PubMed]
- Mira, N.P.; Madeira, A.; Moreira, A.S.; Coutinho, C.P.; Sá-Correia, I. Genomic expression analysis reveals strategies of Burkholderia cenocepacia to adapt to cystic fibrosis patients’ airways and antimicrobial therapy. PLoS ONE 2011, 6, e28831. [Google Scholar] [CrossRef] [PubMed]
- Visser, M.B.; Majumdar, S.; Hani, E.; Sokol, P.A. Importance of the ornibactin and pyochelin siderophore transport systems in Burkholderia cenocepacia lung infections. Infect. Immun. 2004, 72, 2850–2857. [Google Scholar] [CrossRef] [PubMed]
- Tuanyok, A.; Kim, H.S.; Nierman, W.C.; Yu, Y.; Dunbar, J.; Moore, R.A.; Baker, P.; Tom, M.; Ling, J.M.; Woods, D.E. Genome-wide expression analysis of iron regulation in Burkholderia pseudomallei and Burkholderia mallei using DNA microarrays. FEMS Microbiol. Lett. 2005, 252, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, B.; Alvarez, J.; Menéndez, A.; Guijarro, J.A. A mutant in one of two exbD loci of a TonB system in Flavobacterium psychrophilum shows attenuated virulence and confers protection against cold water disease. Microbiology 2008, 154, 1144–1151. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.F.; Lin, T.L.; Lee, C.Z.; Tsai, S.F.; Wang, J.T. Serum-induced iron-acquisition systems and TonB contribute to virulence in Klebsiella pneumoniae causing primary pyogenic liver abscess. J. Infect. Dis. 2008, 197, 1717–1727. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, C.L.; Mott, T.M.; Muruato, L.A.; Sbrana, E.; Torres, A.G. Burkholderia mallei CLH001 Attenuated Vaccine Strain Is Immunogenic and Protects against Acute Respiratory Glanders. Infect. Immun. 2016, 84, 2345–2354. [Google Scholar] [CrossRef] [PubMed]
- Mahenthiralingam, E.; Coenye, T.; Chung, J.W.; Speert, D.P.; Govan, J.R.; Taylor, P.; Vandamme, P. Diagnostically and experimentally useful panel of strains from the Burkholderia cepacia complex. J. Clin. Microbiol. 2000, 38, 910–913. [Google Scholar] [PubMed]
- Torres, A.G.; Payne, S.M. Haem iron-transport system in enterohaemorrhagic Escherichia coli O157:H7. Mol. Microbiol. 1997, 23, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of Blue Agar CAS Assay for Siderophore Detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar]
- Lewis, E.R.G.; Kilgore, P.B.; Mott, T.M.; Pradenas, G.A.; Torres, A.G. Comparing in vitro and in vivo virulence phenotypes of Burkholderia pseudomallei type G strains. PLoS ONE 2017, 12, e0175983. [Google Scholar] [CrossRef] [PubMed]
- Hamad, M.A.; Zajdowicz, S.L.; Holmes, R.K.; Voskuil, M.I. An allelic exchange system for compliant genetic manipulation of the select agents Burkholderia pseudomallei and Burkholderia mallei. Gene 2009, 430, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, G.M.; Sullivan, T.D.; Gallardo, S.S.; Brandhorst, T.T.; Vanden Wymelenberg, A.J.; Cuomo, C.A.; Suen, G.; Currie, C.R.; Klein, B.S. SREB, a GATA transcription factor that directs disparate fates in Blastomyces dermatitidis including morphogenesis and siderophore biosynthesis. PLoS Pathog. 2010, 6, e1000846. [Google Scholar] [CrossRef] [PubMed]
- Speert, D.P.; Steen, B.; Halsey, K.; Kwan, E. A murine model for infection with Burkholderia cepacia with sustained persistence in the spleen. Infect. Immun. 1999, 67, 4027–4032. [Google Scholar] [PubMed]
- Tiringer, K.; Treis, A.; Fucik, P.; Gona, M.; Gruber, S.; Renner, S.; Dehlink, E.; Nachbaur, E.; Horak, F.; Jaksch, P.; et al. A Th17- and Th2-skewed cytokine profile in cystic fibrosis lungs represents a potential risk factor for Pseudomonas aeruginosa infection. Am. J. Respir. Crit. Care Med. 2013, 187, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Moser, C.; Kjaergaard, S.; Pressler, T.; Kharazmi, A.; Koch, C.; Høiby, N. The immune response to chronic Pseudomonas aeruginosa lung infection in cystic fibrosis patients is predominantly of the Th2 type. APMIS 2000, 108, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Moser, C.; Jensen, P.Ø.; Pressler, T.; Frederiksen, B.; Lanng, S.; Kharazmi, A.; Koch, C.; Høiby, N. Serum concentrations of GM-CSF and G-CSF correlate with the Th1/Th2 cytokine response in cystic fibrosis patients with chronic Pseudomonas aeruginosa lung infection. APMIS 2005, 113, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Hartl, D. Immunological mechanisms behind the cystic fibrosis-ABPA link. Med. Mycol. 2009, 47, S183–S191. [Google Scholar] [CrossRef] [PubMed]
- Hartl, D.; Griese, M.; Kappler, M.; Zissel, G.; Reinhardt, D.; Rebhan, C.; Schendel, D.J.; Krauss-Etschmann, S. Pulmonary T(H)2 response in Pseudomonas aeruginosa-infected patients with cystic fibrosis. J. Allergy. Clin. Immunol. 2006, 117, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Wojnarowski, C.; Frischer, T.; Hofbauer, E.; Grabner, C.; Mosgoeller, W.; Eichler, I.; Ziesche, R. Cytokine expression in bronchial biopsies of cystic fibrosis patients with and without acute exacerbation. Eur. Respir. J. 1999, 14, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Moser, C.; Jensen, P.O.; Kobayashi, O.; Hougen, H.P.; Song, Z.; Rygaard, J.; Kharazmi, A.; Høiby, N. Improved outcome of chronic Pseudomonas aeruginosa lung infection is associated with induction of a Th1-dominated cytokine response. Clin. Exp. Immunol. 2002, 127, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.B.; Dow, S.W. Development of Burkholderia mallei and pseudomallei vaccines. Front. Cell Infect. Microbiol. 2013, 3, 10. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Relevant Features a | Reference or Source |
|---|---|---|
| B. cenocepacia K56-2 | Human clinical isolate; Tets Pbr | [22] |
| GAP001 | B. cenocepacia K56-2 ΔtonB | This study |
| B. cenocepacia K56-2 b | Human clinical isolate; Nacr Pbr | This study |
| E. coli S17-1 (pGPTcR) | E. coli strain containing pGPTcR (pMO130 tet) Pbs Tetr | This study |
| E. coli S17-1 (pGPTcR-tonB) | Donor strain containing pGPTcR-tonB plasmid; Pbs Tetr | This study |
| Strain | LB | LB + Fe |
|---|---|---|
| B. cenocepacia K56-2 | 22.00 ± 0.36 | 21.67 ± 0.49 |
| GAP001 b | 41.00 ± 1.26 **** | 36.17 ± 0.60 **** |
| Strain | FeSO4 | Hemoglobin | Hemin | Lactoferrin | Transferrin |
|---|---|---|---|---|---|
| B. cenocepacia K56-2 b | + | + | + | + | + |
| GAP001 c | + | − | − | − | − |
| Treatment a | Serum titer b IgG | IgG2a | IgG1 |
|---|---|---|---|
| PBS | ND | ND | ND |
| GAP001/5.4 × 107 CFU | 51,200 ± 0 | 8533 ± 3200 | ND |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pradenas, G.A.; Myers, J.N.; Torres, A.G. Characterization of the Burkholderia cenocepacia TonB Mutant as a Potential Live Attenuated Vaccine. Vaccines 2017, 5, 33. https://doi.org/10.3390/vaccines5040033
Pradenas GA, Myers JN, Torres AG. Characterization of the Burkholderia cenocepacia TonB Mutant as a Potential Live Attenuated Vaccine. Vaccines. 2017; 5(4):33. https://doi.org/10.3390/vaccines5040033
Chicago/Turabian StylePradenas, Gonzalo A., Julia N. Myers, and Alfredo G. Torres. 2017. "Characterization of the Burkholderia cenocepacia TonB Mutant as a Potential Live Attenuated Vaccine" Vaccines 5, no. 4: 33. https://doi.org/10.3390/vaccines5040033