
Innate Immune Response Assessment in Cyprinus carpio L. upon Experimental Administration with Artemia salina Bio-Encapsulated Aeromonas hydrophila Bacterin

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Experimental Design
2.3. Preparation of Inactivated A. hydrophila
2.4. Preparation of Bio-Encapsulated Antigen
2.4.1. Hatching of Cysts
2.4.2. Optimization of Bio-Encapsulation
2.4.3. Bio-Encapsulation
2.5. Preparation of Antigen for Immersion
2.6. Fish Sampling
2.7. Differential Leucocyte Count of Blood Sample (DLC)
2.8. Non-Specific Immune Parameters
2.8.1. Lysozyme
2.8.2. Myeloperoxidase
2.8.3. Respiratory Burst Activity
2.8.4. Serum Antiprotease Activity
2.8.5. Alkaline Phosphatase
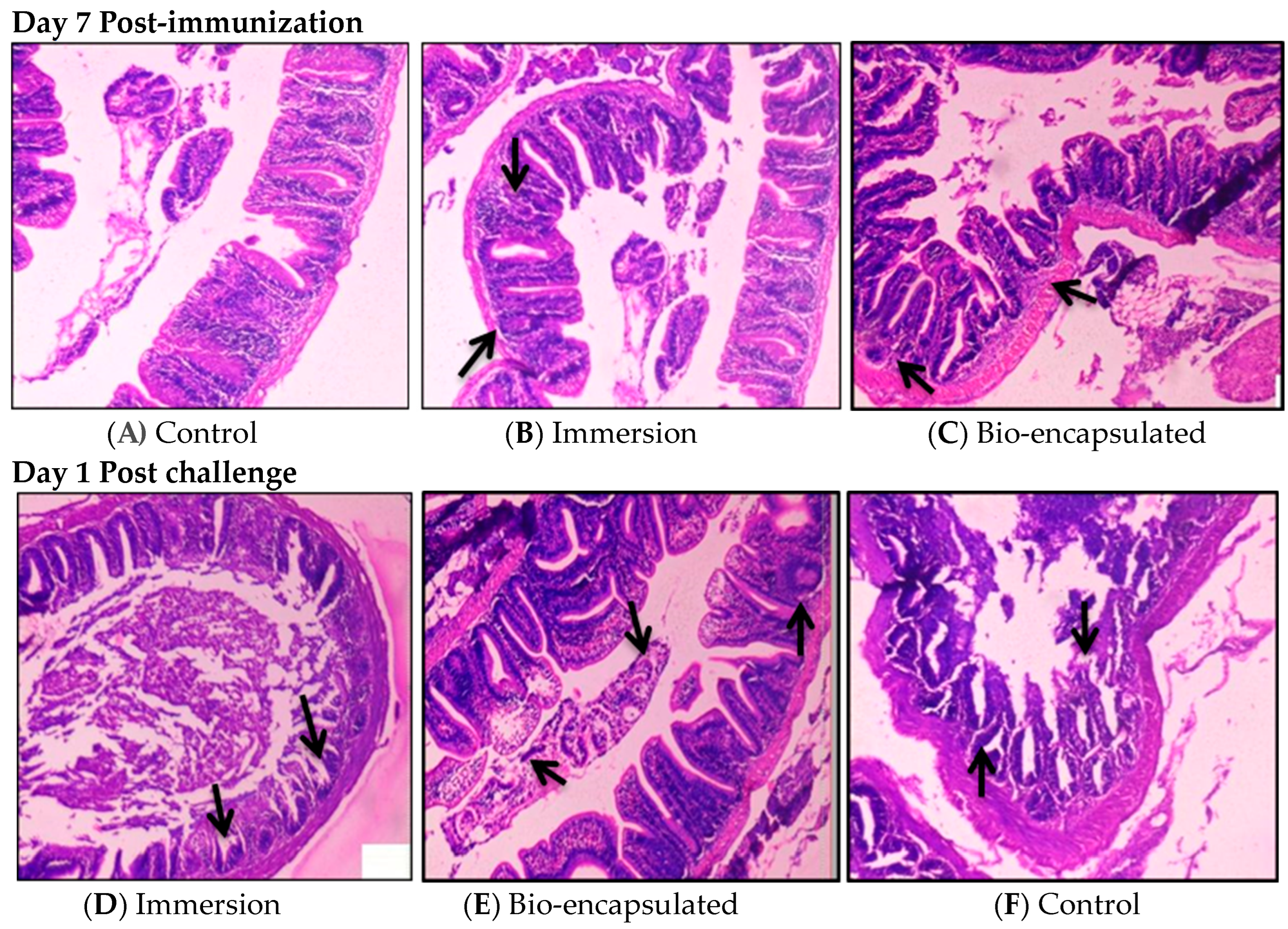
2.8.6. Histology
2.9. Challenge with Live Aeromonas hydrophila
2.10. Statistical Analyses
3. Results
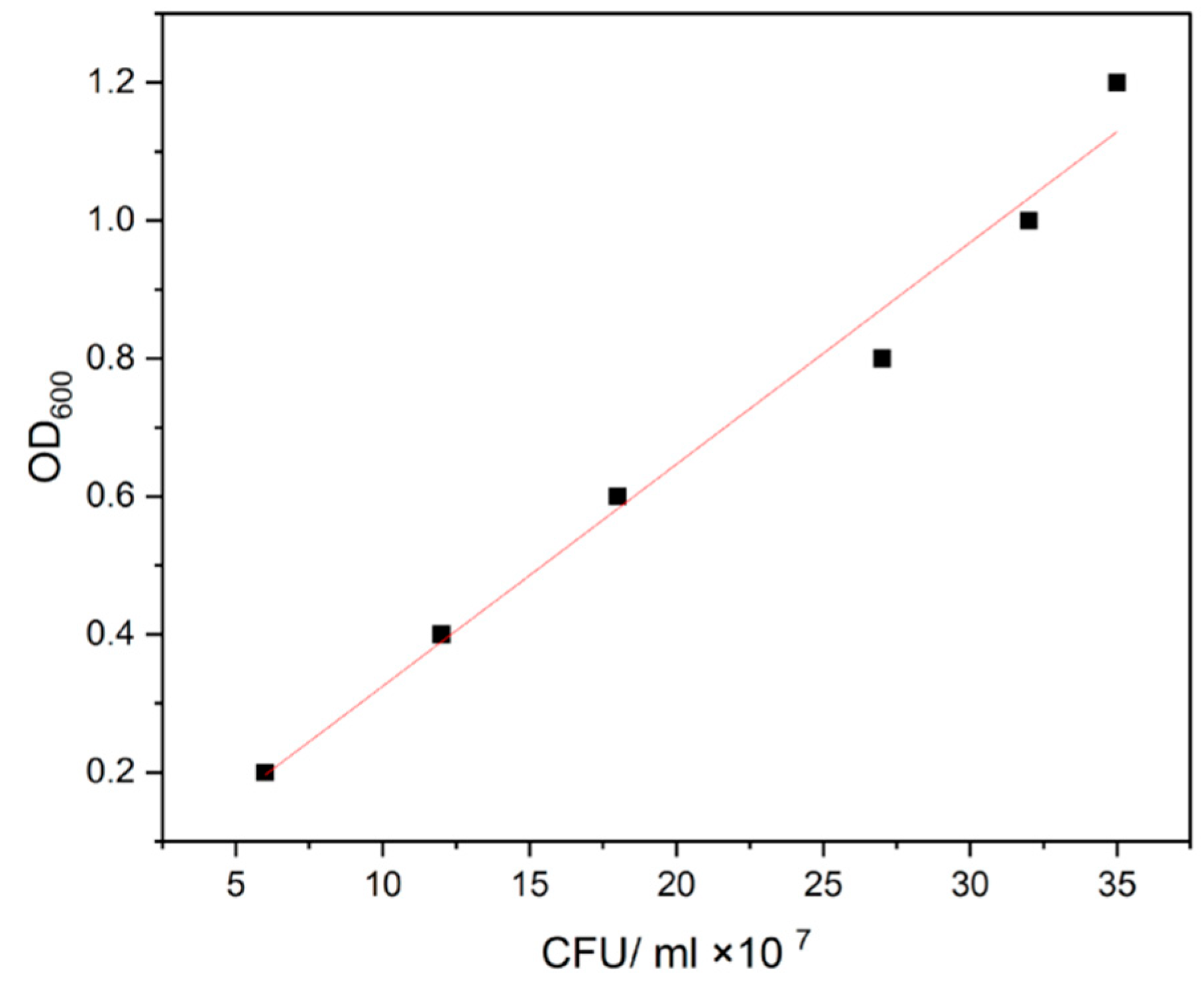
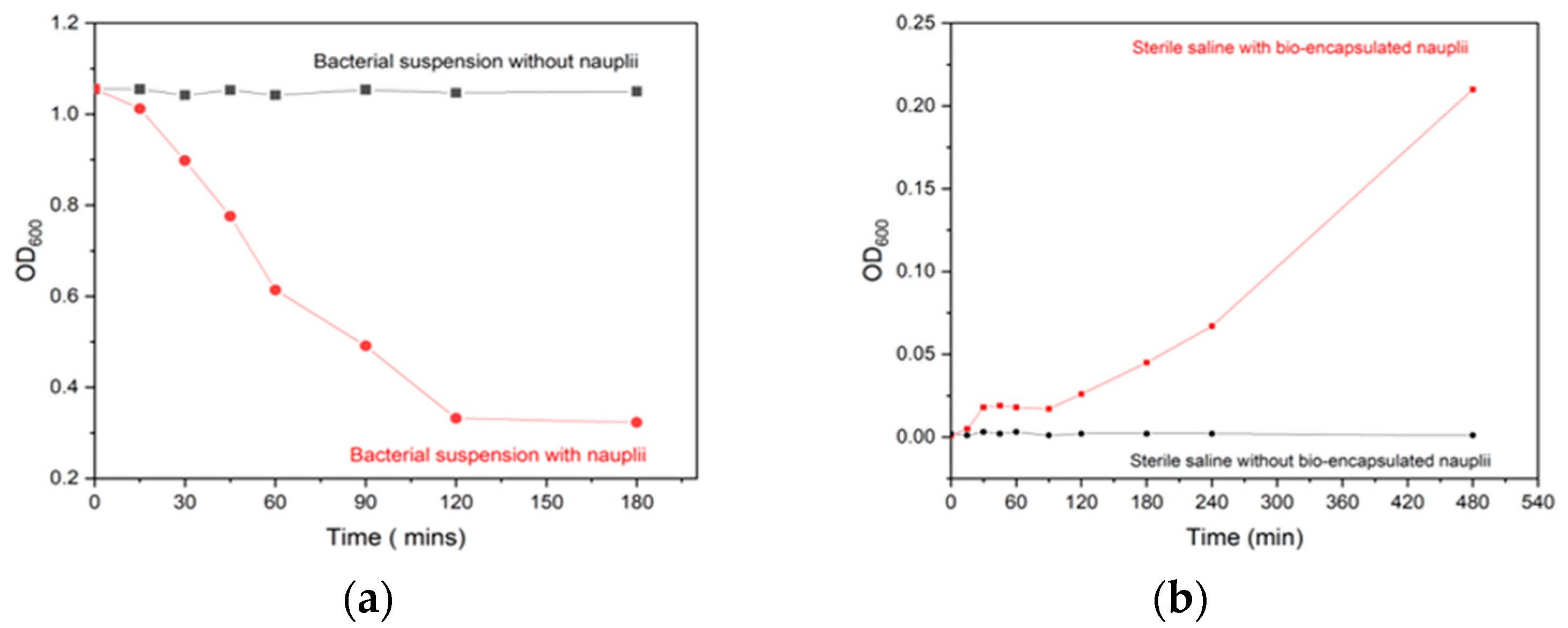
3.1. Bio-Encapsulation
3.2. Differential Leucocyte Count (DLC)
3.3. Non-Specific Immune Parameters
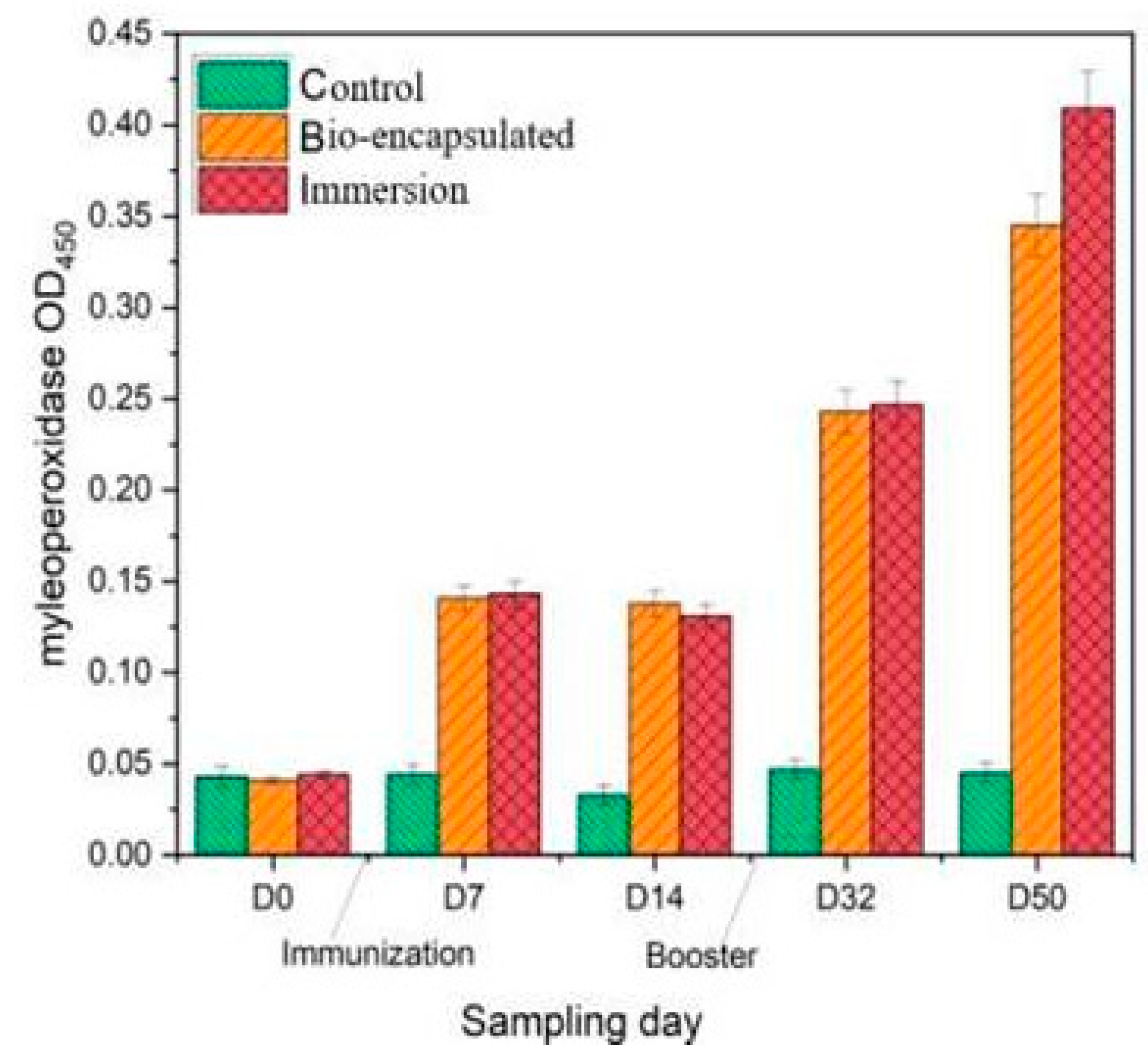
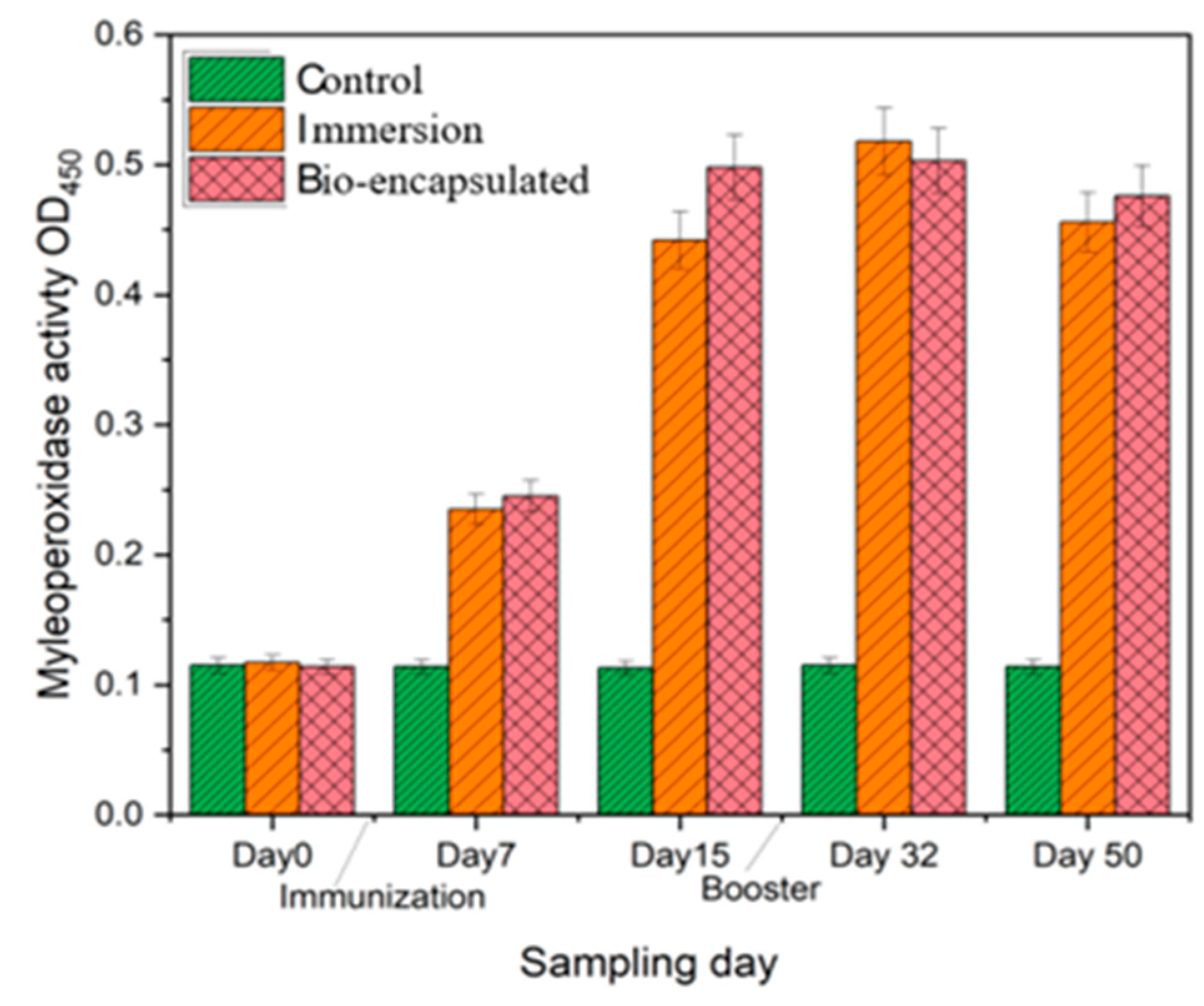
3.3.1. Myeloperoxidase
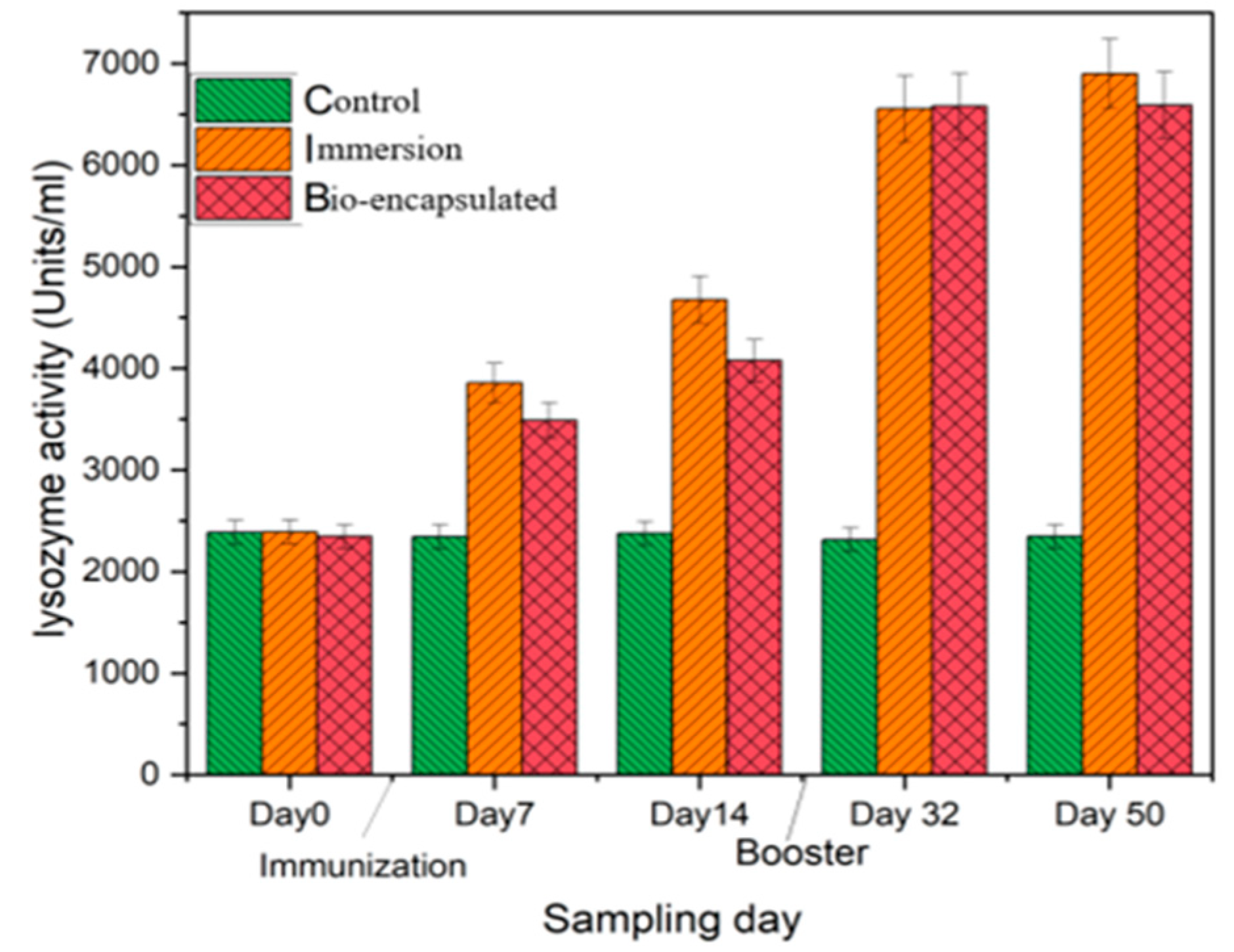
3.3.2. Lysozyme
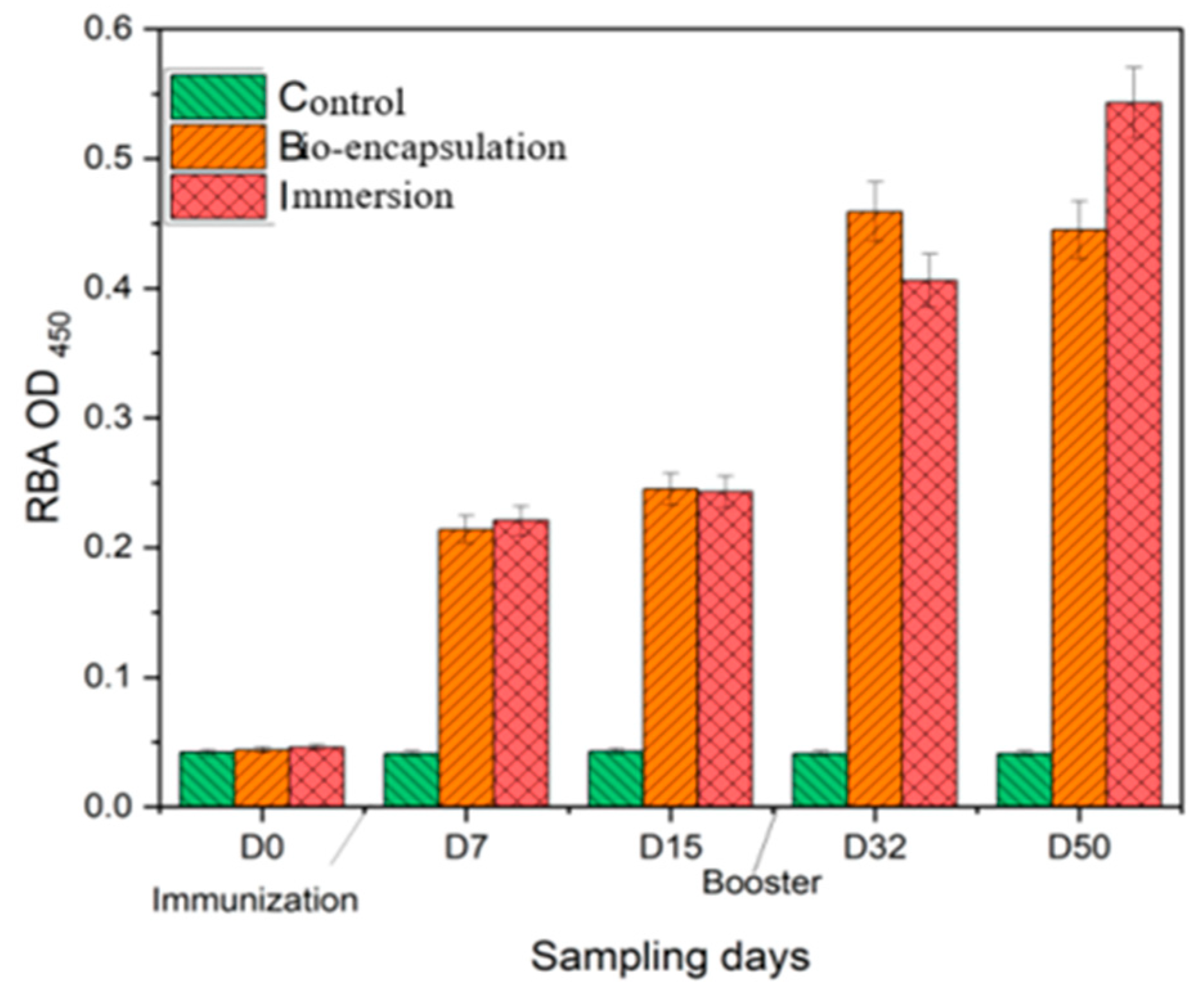
3.3.3. Respiratory Burst Activity (RBA)
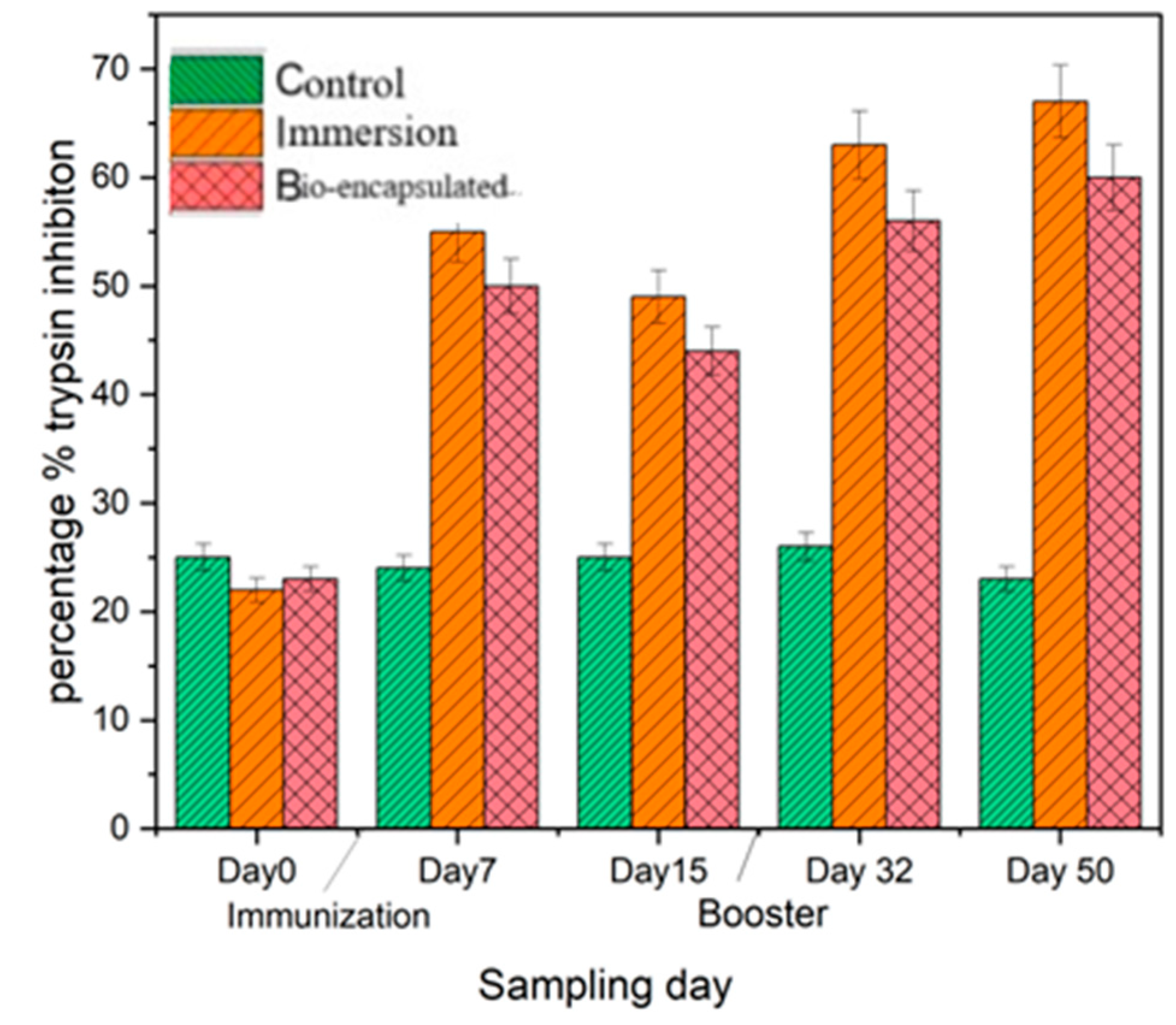
3.3.4. Serum Antiprotease Activity
3.3.5. Intestinal Tissue Myeloperoxidase (MPO) Activity
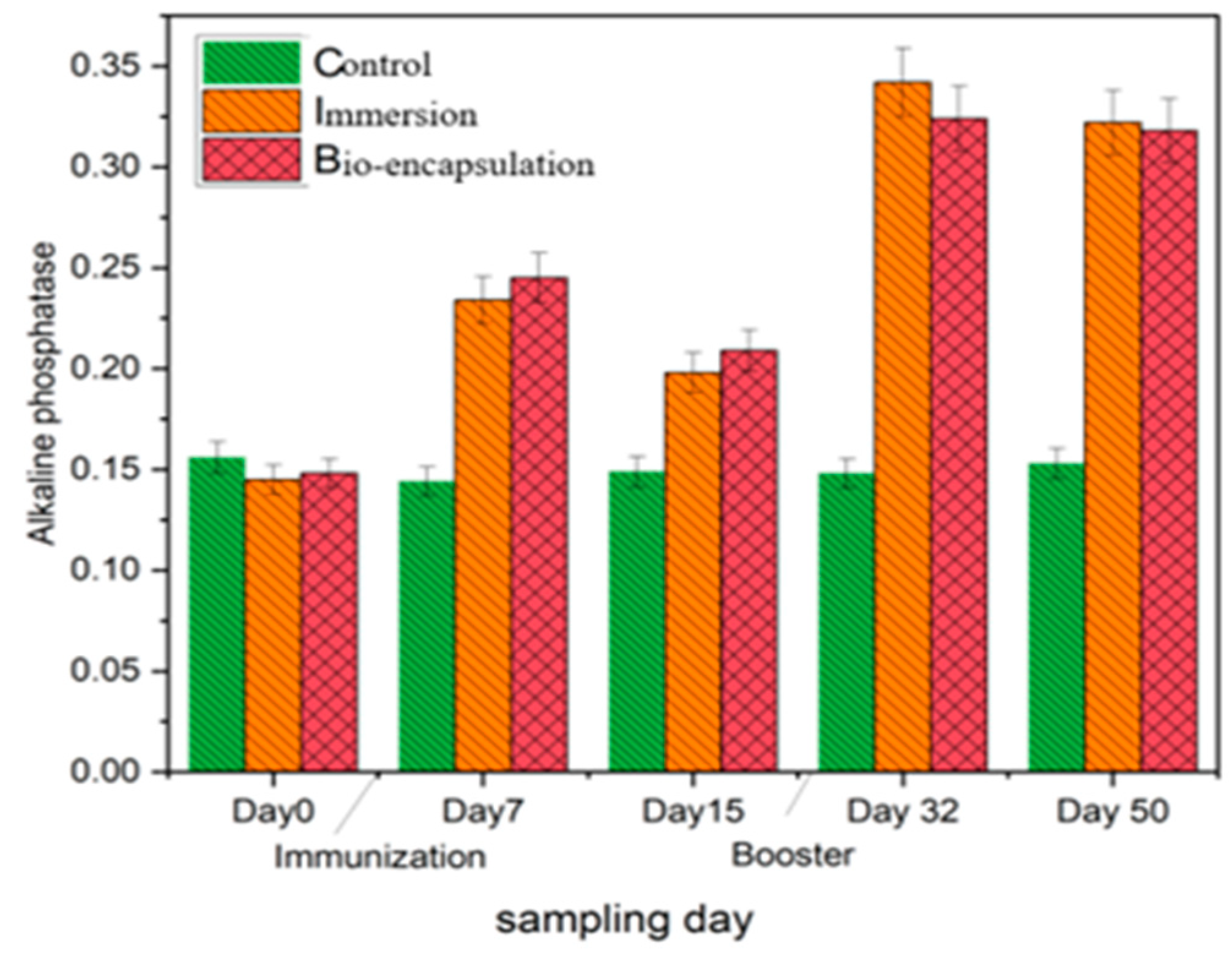
3.3.6. Alkaline Phosphatase (AKP)
3.3.7. Intestinal Tissue Lysozyme Activity
3.3.8. Histology
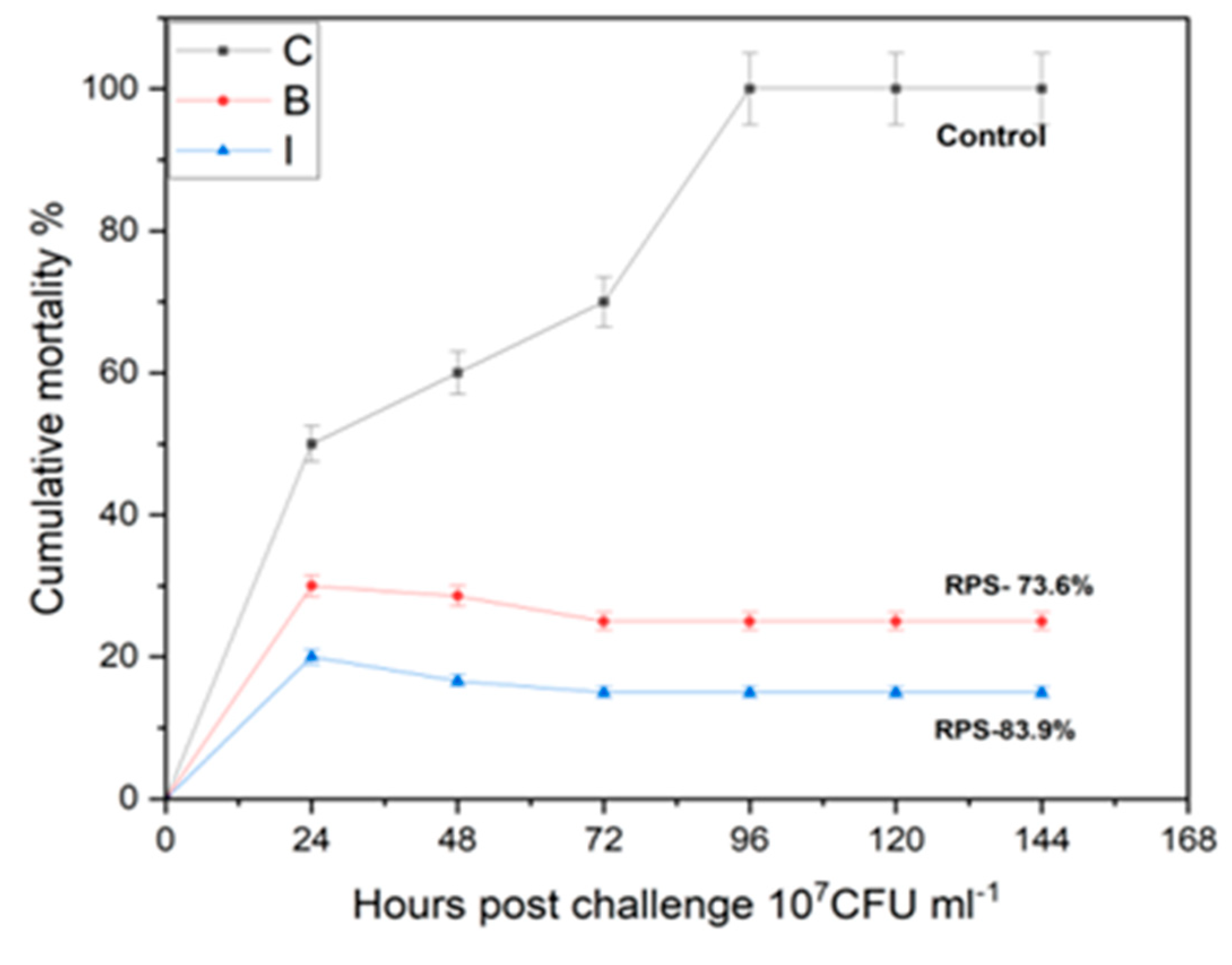
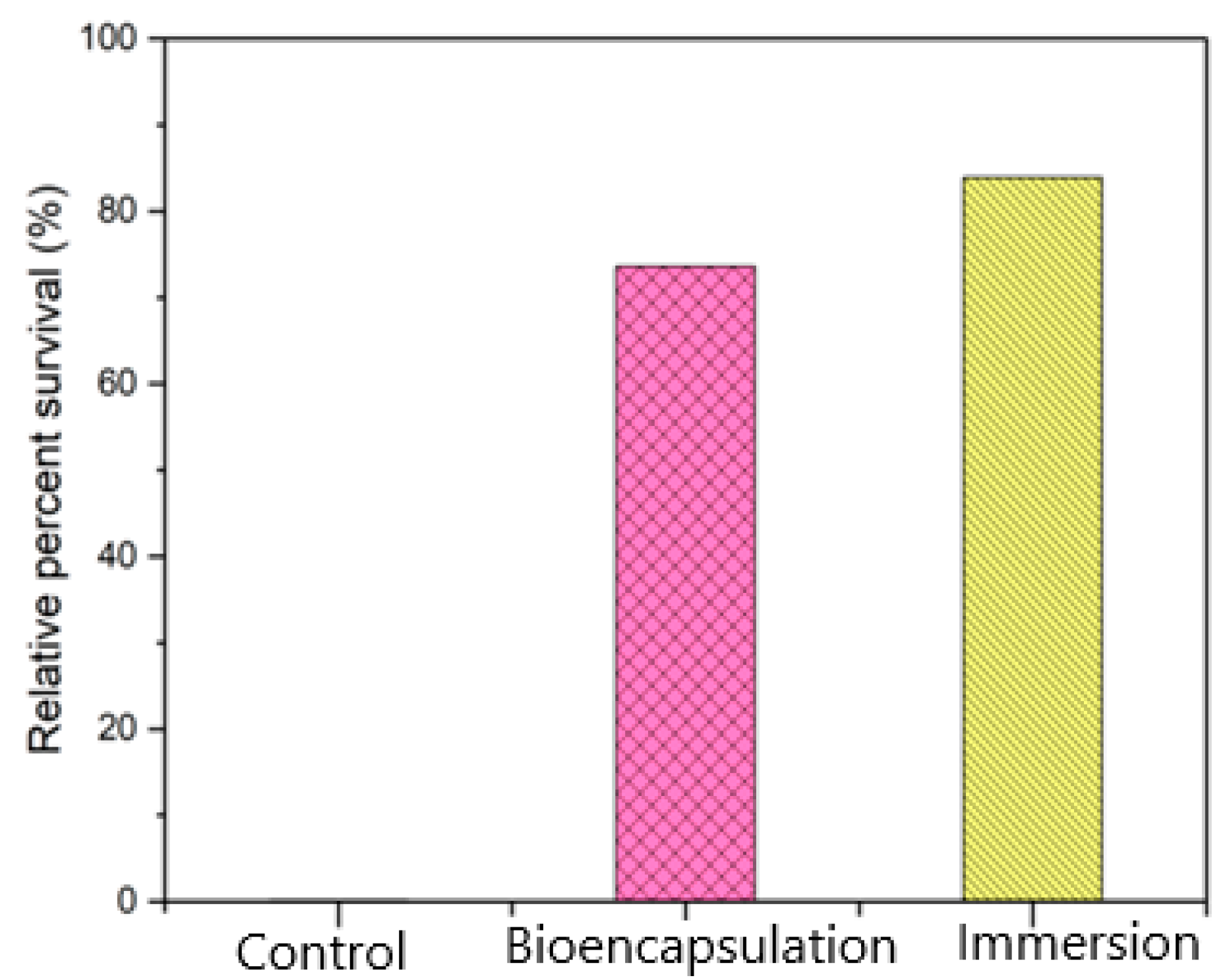
3.4. Challenge with Live Aeromonas hydrophila
Cumulative Mortality Index (CMI) and Post Challenge Symptoms
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boyd, C.E.; McNevin, A.A. Aquaculture, Resource Use, and the Environment; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Okamura, B.; Feist, S.W. Emerging diseases in freshwater systems. Freshw. Biol. 2011, 56, 627–637. [Google Scholar] [CrossRef]
- Akbar Ali, I.; Radhakrishnan, D.K.; Kumar, S. Immunostimulants and their uses in aquaculture. In Aquaculture Science and Engineering; Balasubramanian, B., Liu, W.-C., Sattanathan, G., Eds.; Springer Nature: Singapore, 2022; pp. 291–322. [Google Scholar] [CrossRef]
- Rombout, J.H.W.M.; Yang, G.; Kiron, V. Adaptive immune responses at mucosal surfaces of teleost fish. Fish Shellfish Immunol. 2014, 40, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Uribe, C.; Folch, H.; Enriquez, R.; Moran, G. Innate and adaptive immunity in teleost fish: A review. Vet. Med. 2011, 56, 486–503. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, W.; Wu, H.; Gong, X.; Li, A. Multilocus sequence typing revealed a clonal lineage of Aeromonas hydrophila caused motile Aeromonas septicemia outbreaks in pond-cultured cyprinid fish in an epidemic area in central China. Aquaculture 2014, 432, 1–6. [Google Scholar] [CrossRef]
- Abowei, J.F. Salinity, dissolved oxygen, pH and surface water temperature conditions in Nkoro River, Niger Delta, Nigeria. Adv. J. Food Sci. Technol. 2010, 2, 36–40. [Google Scholar]
- Joosten, P.H.M.; Avilés-Trigueros, M.; Sorgeloos, P.; Rombout, J.H.W.M. Oral vaccination of juvenile carp (Cyprinus carpio) and gilthead seabream (Sparus aurata) with bioencapsulated Vibrio anguillarum bacterin. Fish Shellfish Immunol. 1995, 5, 289–299. [Google Scholar] [CrossRef]
- Yasumoto, S.; Miyanari, S.; Yoshimura, S.; Kondo, M.; Takahashi, Y. Effect of oral administration of 5-aminolevulinic acid in Japanese eel Anguilla japonica. Aquac. Sci. 2012, 60, 411–412. [Google Scholar]
- Farias, T.H.V.; Arijo, S.; Medina, A.; Pala, G.; da Rosa Prado, E.J.; Montassier, H.J.; Pilarski, F.; de Andrade Belo, M.A. Immune responses induced by inactivated vaccine against Aeromonas hydrophila in pacu, Piaractus mesopotamicus. Fish Shellfish Immunol. 2020, 101, 186–191. [Google Scholar] [CrossRef]
- Lavens, P.; Sorgeloos, P. The cryptobiotic state of Artemia cysts, its diapause deactivation and hatching: A review. Artemia Res. Its Appl. 1987, 3, 27–63. [Google Scholar]
- Gomez-Gil, B.; Herrera-Vega, M.A.; Abreu-Grobois, F.A.; Roque, A. Bioencapsulation of two different vibrio species in nauplii of the brine shrimp (Artemia franciscana). Appl. Environ. Microbiol. 1998, 64, 2318–2322. [Google Scholar] [CrossRef]
- Akbar, I.; Radhakrishnan, D.K.; Venkatachalam, R.; Sathrajith, A.T.; Sureshkumar, S. Standardization of the bioencapsulation of probiotics and oil emulsion in Artemia parthenogenetica. Int. J. Res. Fish. Aquac. 2014, 4, 122–125. [Google Scholar]
- Hamsah, H.; Widanarni, W.; Alimuddin, A.; Yuhana, M.; Junior, M.Z.; Hidayatullah, D. Immune response and resistance of Pacific white shrimp larvae administered probiotic, prebiotic, and synbiotic through the bio-encapsulation of Artemia sp. Aquacult. Int. 2019, 27, 567–580. [Google Scholar] [CrossRef]
- Lamers, C.H.J.; Haas, M.J.H.; Muiswinkel, W.B. The reaction of the immune system of fish to vaccination: Development of immunological memory in carp, Cyprinus carpio L.; following direct immersion in Aeromonas hydrophila bacterin. J. Fish Dis. 1985, 8, 253–262. [Google Scholar] [CrossRef]
- Dang, Z.; Arena, M.; Kienzler, A. Fish toxicity testing for identification of thyroid disrupting chemicals. Environ. Pollut. 2021, 284, 117374. [Google Scholar] [CrossRef]
- Mugimba, K.K.; Lamkhannat, M.; Dubey, S.; Mutoloki, S.; Munang’andu, H.M.; Evensen, Ø. Tilapia lake virus downplays innate immune responses during early stage of infection in Nile tilapia (Oreochromis niloticus). Sci. Rep. 2020, 10, 20364. [Google Scholar] [CrossRef]
- Tripathi, N.K.; Latimer, K.S.; Burnley, V.V. Hematologic reference intervals for koi (Cyprinus carpio), including blood cell morphology, cytochemistry, and ultrastructure. Vet. Clin. Pathol. 2004, 33, 74–83. [Google Scholar] [CrossRef]
- Hutchinson, T.H.; Manning, M.J. Seasonal trends in serum lysozyme activity and total protein concentration in dab (Limanda limanda L.) sampled from Lyme Bay, U.K. Fish Shellfish Immunol. 1996, 6, 473–482. [Google Scholar] [CrossRef]
- Quade, M.J.; Roth, J.A. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunopathol. 1997, 58, 239–248. [Google Scholar] [CrossRef]
- Yeh, S.-P.; Chang, C.-A.; Chang, C.-Y.; Liu, C.-H.; Cheng, W. Dietary sodium alginate administration affects fingerling growth and resistance to Streptococcus sp. and iridovirus, and juvenile non-specific immune responses of the orange-spotted grouper, Epinephelus coioides. Fish Shellfish Immunol. 2008, 25, 19–27. [Google Scholar] [CrossRef]
- Zuo, X.; Woo, P.T. Natural anti-proteases in rainbow trout, Oncorhynchus mykiss and brook charr, Salvelinus fontinalis and the in vitro neutralization of fish alpha 2-macroglobulin by the metalloprotease from the pathogenic haemoflagellate, Cryptobiasalmositica. Parasitology 1997, 114, 375–382. [Google Scholar] [CrossRef]
- Walter, K.; Schütt, C. Acid and alkaline phosphatase in serum. In Methods of Enzymatic Analysis; Elsevier: Amsterdam, The Netherlands, 1974; pp. 856–860. [Google Scholar] [CrossRef]
- Sivakamavalli, J.; Vaseeharan, B. Variations in biochemical and histological characteristics of WSSV infected green tiger shrimp Penaeus semisulcatus. J. Recept. Signal Transduct Res. 2014, 34, 386–395. [Google Scholar] [CrossRef]
- Ardó, L.; Jeney, Z.; Adams, A.; Jeney, G. Immune responses of resistant and sensitive common carp families following experimental challenge with Aeromonas hydrophila. Fish Shellfish Immunol. 2010, 29, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.A.; Amend, D.F. Potential for immersion vaccination against Aeromonas salmonicida. J. Fish Dis. 1984, 7, 101–105. [Google Scholar] [CrossRef]
- Tobar, J.A.; Jerez, S.; Caruffo, M.; Bravo, C.; Contreras, F.; Bucarey, S.A.; Harel, M. Oral vaccination of Atlantic salmon (Salmo salar) against salmonid rickettsial septicaemia. Vaccine 2011, 29, 2336–2340. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.K. Current prospects and challenges in fish vaccine development in India with special reference to Aeromonas hydrophila vaccine. Fish Shellfish Immunol. 2020, 100, 283–299. [Google Scholar] [CrossRef]
- Azad, I.S.; Shankar, K.M.; Mohan And, C.V.; Kalita, B. Biofilm vaccine of Aeromonas hydrophila—Standardization of dose and duration for oral vaccination of carps. Fish Shellfish Immunol. 1999, 9, 519–528. [Google Scholar] [CrossRef]
- Campbell, R.; Adams, A.; Tatner, M.F.; Chair, M.; Sorgeloos, P. Uptake of Vibrio anguillarum vaccine by Artemia salina as a potential oral delivery system to fish fry. Fish Shellfish Immunol. 1993, 3, 451–459. [Google Scholar] [CrossRef]
- Dang, M.; Cao, T.; Vasquez, I.; Hossain, A.; Gnanagobal, H.; Kumar, S.; Hall, J.R.; Monk, J.; Boyce, D.; Westcott, J.; et al. Oral Immunization of Larvae and Juvenile of Lumpfish (Cyclopterus lumpus) against Vibrio anguillarum Does Not Influence Systemic Immunity. Vaccines 2021, 9, 819. [Google Scholar] [CrossRef]
- Das, A.; Sahoo, P.K.; Mohanty, B.R.; Jena, J.K. Pathophysiology of experimental Aeromonas hydrophila infection in Puntius sarana: Early changes in blood and aspects of the innate immune-related gene expression in survivors. Vet. Immunol. Immunopathol. 2011, 142, 207–218. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Y.; Sun, G.; Li, X.; Liu, Z. Growth, immune response, antioxidant capability, and disease resistance of juvenile Atlantic salmon (Salmo salar L.) fed Bacillus velezensis V4 and Rhodotorula mucilaginosa compound. Aquaculture 2019, 500, 65–74. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Devi, G.; Balasundaram, C.; Van Doan, H.; Jaturasitha, S.; Saravanan, K.; RingØ, E. Impact of cinnamaldehyde on innate immunity and immune gene expression in Channa striatus against Aphanomyces invadans. Fish Shellfish Immunol. 2021, 117, 1–16. [Google Scholar] [CrossRef]
- Priyadarshini, S.K.; Subramani, P.A.; Michael, R.D. Modulation of the innate immune responses in the striped snakehead murrel, Channa striata upon experimental infection with live and heat killed Aeromonas hydrophila. Open Vet. J. 2017, 7, 157–164. [Google Scholar] [CrossRef]
- Kumaresan, V.; Harikrishnan, R.; Arockiaraj, J. A potential Kazal-type serine protease inhibitor involves in kinetics of protease inhibition and bacteriostatic activity. Fish Shellfish Immunol. 2015, 42, 430–438. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, X.; Zhao, F.; Zhou, Z.; Zhao, F.; Wang, J.; Liu, T.; Yang, X.; Zhang, X.; Li, Z. Stress responses of the intestinal digestion, antioxidant status, microbiota and non-specific immunity in Songpu mirror carp (Cyprinus carpio L.) under starvation. Fish Shellfish Immunol. 2022, 120, 411–420. [Google Scholar] [CrossRef]
- AlYahya, S.A.; Ameen, F.; Al-Niaeem, K.S.; Al-Sa’adi, B.A.; Hadi, S.; Mostafa, A.A. Histopathological studies of experimental Aeromonas hydrophila infection in blue tilapia, Oreochromis aureus. Saudi J. Biol. Sci. 2018, 25, 182–185. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types of Cells | Control | Immersion | Bioencapsulation |
|---|---|---|---|
| Lymphocytes | 85 ± 1.0 | 82.6 ± 1.15 | 82.6 ± 2.08 |
| Monocytes | 4.6 ± 0.5 | 4.6 ± 1.15 | 3.6 ± 1.5 |
| Granulocytes | 11 ± 1.0 | 12.6 ± 2.0 | 12 ± 2.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radhakrishnan, A.; Prabakaran, D.S.; Ramesh, T.; Sakthivel, R.; Ramasamy, K.; Han, H.-S.; Jeyachandran, S. Innate Immune Response Assessment in Cyprinus carpio L. upon Experimental Administration with Artemia salina Bio-Encapsulated Aeromonas hydrophila Bacterin. Vaccines 2023, 11, 877. https://doi.org/10.3390/vaccines11040877
Radhakrishnan A, Prabakaran DS, Ramesh T, Sakthivel R, Ramasamy K, Han H-S, Jeyachandran S. Innate Immune Response Assessment in Cyprinus carpio L. upon Experimental Administration with Artemia salina Bio-Encapsulated Aeromonas hydrophila Bacterin. Vaccines. 2023; 11(4):877. https://doi.org/10.3390/vaccines11040877
Chicago/Turabian StyleRadhakrishnan, Akshaya, D. S. Prabakaran, Thiyagarajan Ramesh, Ramalingam Sakthivel, Kavikumar Ramasamy, Hyo-Shim Han, and Sivakamavalli Jeyachandran. 2023. "Innate Immune Response Assessment in Cyprinus carpio L. upon Experimental Administration with Artemia salina Bio-Encapsulated Aeromonas hydrophila Bacterin" Vaccines 11, no. 4: 877. https://doi.org/10.3390/vaccines11040877