
Involvement of the MGF 110-11L Gene in the African Swine Fever Replication and Virulence

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Design and Assembly
2.2. Production and Isolation of Mutants
2.3. Titer Determination
2.4. Preparation and Culture of Porcine Alveolar Macrophages (PAMs)
2.5. Copy Number Determination
2.6. Sequencing
2.7. Statistical Analysis
2.8. Animal Experiments
2.9. Collection of Blood Samples
2.10. ELISA Test
2.11. Real-Time PCR
3. Results
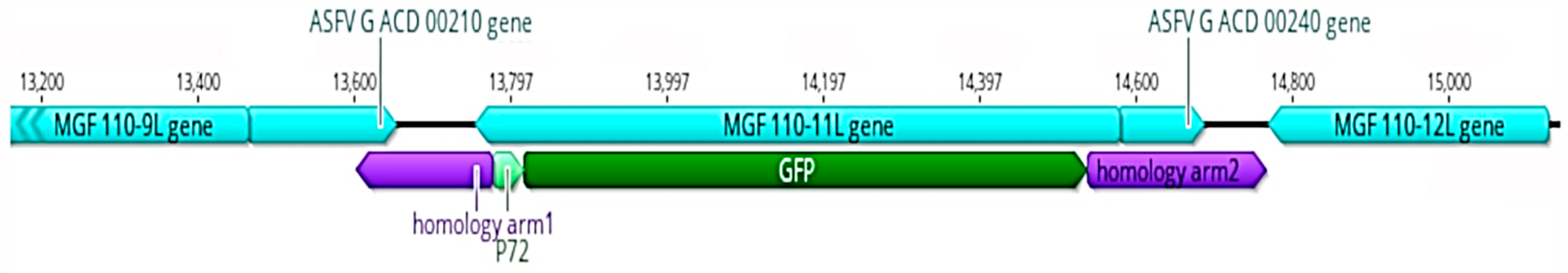
3.1. Properties of MGF 110-11L Gene
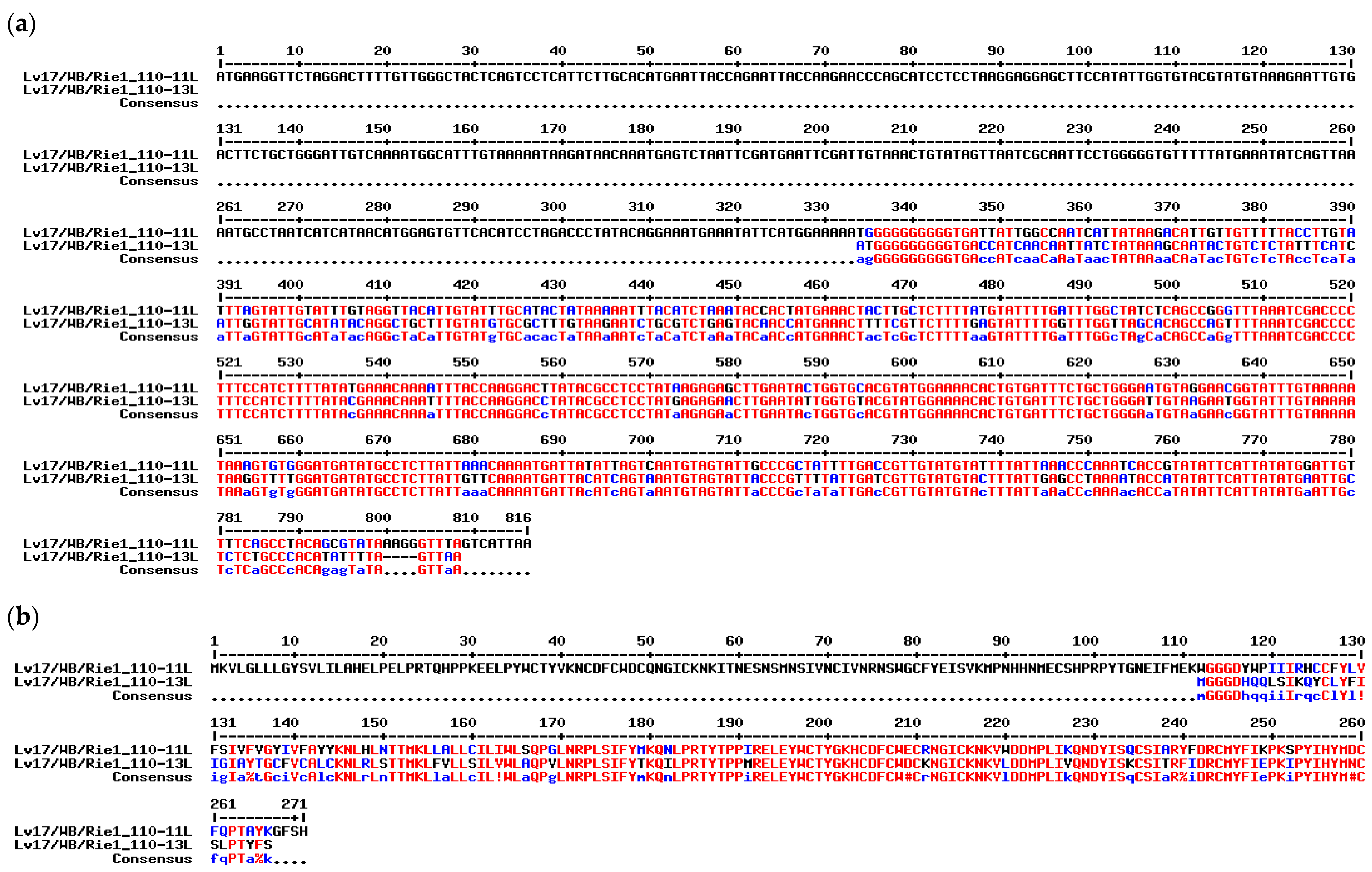
3.2. Generation and Sequencing the Lv17/WB/Rie1/d110-11L Virus
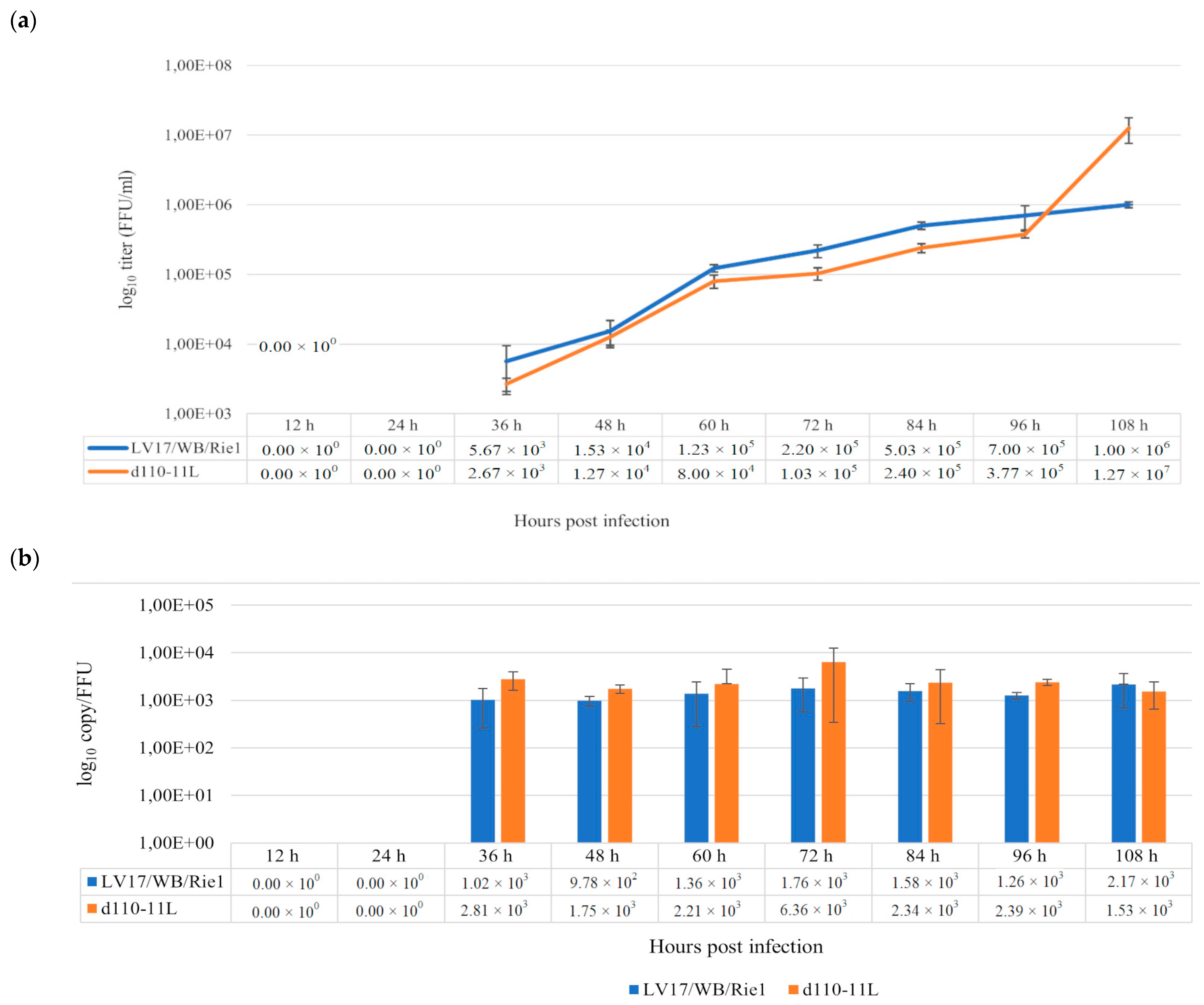
3.3. Replication of Lv17/WB/Rie1/d110-11L in Cultured PAMs
3.4. In Vivo Characteristics of Lv17/WB/Rie1/d110-11L as a Vaccine Candidate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eustace Montgomery, R. On A Form of Swine Fever Occurring in British East Africa (Kenya Colony). J. Comp. Pathol. Ther. 1921, 34, 159–191. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.; Plowright, W.; Pierce, M.A. The Epizootiology of African Swine Fever in Africa. Vet. Rec. 1969, 85, 668–674. [Google Scholar] [PubMed]
- Galindo, I.; Alonso, C. African Swine Fever Virus: A Review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costard, S.; Mur, L.; Lubroth, J.; Sanchez-Vizcaino, J.M.; Pfeiffer, D.U. Epidemiology of African Swine Fever Virus. Virus Res. 2013, 173, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Correa-Fiz, F.; Blanco-Fuertes, M.; Navas, M.J.; Lacasta, A.; Bishop, R.P.; Githaka, N.; Onzere, C.; Le Potier, M.-F.; Almagro-Delgado, V.; Martinez, J.; et al. Comparative Analysis of the Fecal Microbiota from Different Species of Domesticated and Wild Suids. Sci. Rep. 2019, 9, 13616. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, C.; Sánchez, E.G.; Pérez-Núñez, D.; Nogal, M.; de León, P.; Carrascosa, Á.L.; Nieto, R.; Soler, A.; Arias, M.L.; Revilla, Y. African Swine Fever Virus (ASFV) Protection Mediated by NH/P68 and NH/P68 Recombinant Live-Attenuated Viruses. Vaccine 2018, 36, 2694–2704. [Google Scholar] [CrossRef]
- Zhu, J.J. African Swine Fever Vaccinology: The Biological Challenges from Immunological Perspectives. Viruses 2022, 14, 2021. [Google Scholar] [CrossRef]
- Alonso, C.; Borca, M.; Dixon, L.; Revilla, Y.; Rodriguez, F.; Escribano, J.M. ICTV Report Consortium ICTV Virus Taxonomy Profile: Asfarviridae. J. Gen. Virol. 2018, 99, 613–614. [Google Scholar] [CrossRef]
- Alejo, A.; Matamoros, T.; Guerra, M.; Andrés, G. A Proteomic Atlas of the African Swine Fever Virus Particle. J. Virol. 2018, 92, e01293-18. [Google Scholar] [CrossRef] [Green Version]
- Dixon, L.K.; Chapman, D.A.G.; Netherton, C.L.; Upton, C. African Swine Fever Virus Replication and Genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef]
- Cackett, G.; Matelska, D.; Sýkora, M.; Portugal, R.; Malecki, M.; Bähler, J.; Dixon, L.; Werner, F. The African Swine Fever Virus Transcriptome. J. Virol. 2020, 94, e00119-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Vega, I.; Viñuela, E.; Blasco, R. Genetic Variation and Multigene Families in African Swine Fever Virus. Virology 1990, 179, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Chapman, D.A.G.; Tcherepanov, V.; Upton, C.; Dixon, L.K. Comparison of the Genome Sequences of Non-Pathogenic and Pathogenic African Swine Fever Virus Isolates. J. Gen. Virol. 2008, 89, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Blasco, R.; Agüero, M.; Almendral, J.; Viñuela, E. Variable and Constant Regions in African Swine Fever Virus DNA. Virology 1989, 168, 330–338. [Google Scholar] [CrossRef]
- Yozawa, T.; Kutish, G.F.; Afonso, C.L.; Lu, Z.; Rock, D.L. Two Novel Multigene Families, 530 and 300, in the Terminal Variable Regions of African Swine Fever Virus Genome. Virology 1994, 202, 997–1002. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, H.; Liu, L.; Cao, Y.; Jiang, T.; Zou, Y.; Peng, Y. Classification and Characterization of Multigene Family Proteins of African Swine Fever Viruses. Brief Bioinform. 2021, 22, bbaa380. [Google Scholar] [CrossRef] [PubMed]
- Afonso, C.L.; Piccone, M.E.; Zaffuto, K.M.; Neilan, J.; Kutish, G.F.; Lu, Z.; Balinsky, C.A.; Gibb, T.R.; Bean, T.J.; Zsak, L.; et al. African Swine Fever Virus Multigene Family 360 and 530 Genes Affect Host Interferon Response. J. Virol. 2004, 78, 1858–1864. [Google Scholar] [CrossRef] [Green Version]
- Neilan, J.G.; Zsak, L.; Lu, Z.; Kutish, G.F.; Afonso, C.L.; Rock, D.L. Novel Swine Virulence Determinant in the Left Variable Region of the African Swine Fever Virus Genome. J. Virol. 2002, 76, 3095–3104. [Google Scholar] [CrossRef] [Green Version]
- Zsak, L.; Lu, Z.; Burrage, T.G.; Neilan, J.G.; Kutish, G.F.; Moore, D.M.; Rock, D.L. African Swine Fever Virus Multigene Family 360 and 530 Genes Are Novel Macrophage Host Range Determinants. J. Virol. 2001, 75, 3066–3076. [Google Scholar] [CrossRef] [Green Version]
- Agüero, M.; Blasco, R.; Wilkinson, P.; Viñuela, E. Analysis of Naturally Occurring Deletion Variants of African Swine Fever Virus: Multigene Family 110 Is Not Essential for Infectivity or Virulence in Pigs. Virology 1990, 176, 195–204. [Google Scholar] [CrossRef]
- Upton, C. Multigene Families in African Swine Fever Virus. Available online: https://4virology.net/organisms/dsdna-viruses/asfarviridae/mgf-110/ (accessed on 20 July 2020).
- Almendral, J.M.; Almazán, F.; Blasco, R.; Viñuela, E. Multigene Families in African Swine Fever Virus: Family 110. J. Virol. 1990, 64, 2064–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, S.; Ribeiro, G.; Costa, J.V. Sequence and Organization of the Left Multigene Family 110 Region of the Vero-Adapted L60V Strain of African Swine Fever Virus. Virus Genes 1997, 15, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Zani, L.; Forth, J.H.; Forth, L.; Nurmoja, I.; Leidenberger, S.; Henke, J.; Carlson, J.; Breidenstein, C.; Viltrop, A.; Höper, D.; et al. Deletion at the 5’-End of Estonian ASFV Strains Associated with an Attenuated Phenotype. Sci. Rep. 2018, 8, 6510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Cordón, P.J.; Chapman, D.; Jabbar, T.; Reis, A.L.; Goatley, L.; Netherton, C.L.; Taylor, G.; Montoya, M.; Dixon, L. Different Routes and Doses Influence Protection in Pigs Immunised with the Naturally Attenuated African Swine Fever Virus Isolate OURT88/3. Antivir. Res. 2017, 138, 1–8. [Google Scholar] [CrossRef]
- Ramirez-Medina, E.; Vuono, E.; Pruitt, S.; Rai, A.; Silva, E.; Espinoza, N.; Zhu, J.; Velazquez-Salinas, L.; Borca, M.V.; Gladue, D.P. Development and In Vivo Evaluation of a MGF110-1L Deletion Mutant in African Swine Fever Strain Georgia. Viruses 2021, 13, 286. [Google Scholar] [CrossRef]
- Netherton, C.; Rouiller, I.; Wileman, T. The Subcellular Distribution of Multigene Family 110 Proteins of African Swine Fever Virus Is Determined by Differences in C-Terminal KDEL Endoplasmic Reticulum Retention Motifs. J. Virol. 2004, 78, 3710–3721. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Medina, E.; Vuono, E.; Silva, E.; Rai, A.; Valladares, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Borca, M.V.; Gladue, D.P. Evaluation of the Deletion of MGF110-5L-6L on Swine Virulence from the Pandemic Strain of African Swine Fever Virus and Use as a DIVA Marker in Vaccine Candidate ASFV-G-ΔI177L. J. Virol. 2022, 96, e00597-22. [Google Scholar] [CrossRef]
- Zhong, H.; Fan, S.; Du, Y.; Zhang, Y.; Zhang, A.; Jiang, D.; Han, S.; Wan, B.; Zhang, G. African Swine Fever Virus MGF110-7L Induces Host Cell Translation Suppression and Stress Granule Formation by Activating the PERK/PKR-EIF2α Pathway. Microbiol. Spectr. 2022, 10, e03282-22. [Google Scholar] [CrossRef]
- Li, D.; Liu, Y.; Qi, X.; Wen, Y.; Li, P.; Ma, Z.; Liu, Y.; Zheng, H.; Liu, Z. African Swine Fever Virus MGF-110-9L-Deficient Mutant Has Attenuated Virulence in Pigs. Virol. Sin. 2021, 36, 187–195. [Google Scholar] [CrossRef]
- Barasona, J.A.; Cadenas-Fernández, E.; Kosowska, A.; Barroso-Arévalo, S.; Rivera, B.; Sánchez, R.; Porras, N.; Gallardo, C.; Sánchez-Vizcaíno, J.M. Safety of African Swine Fever Vaccine Candidate Lv17/WB/Rie1 in Wild Boar: Overdose and Repeated Doses. Front. Immunol. 2021, 12, 761753. [Google Scholar] [CrossRef]
- Gallardo, C.; Soler, A.; Rodze, I.; Nieto, R.; Cano-Gómez, C.; Fernandez-Pinero, J.; Arias, M. Attenuated and Non-haemadsorbing (Non- HAD ) Genotype II African Swine Fever Virus ( ASFV ) Isolated in Europe, Latvia 2017. Transbound. Emerg. Dis. 2019, 66, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Barasona, J.A.; Gallardo, C.; Cadenas-Fernández, E.; Jurado, C.; Rivera, B.; Rodríguez-Bertos, A.; Arias, M.; Sánchez-Vizcaíno, J.M. First Oral Vaccination of Eurasian Wild Boar Against African Swine Fever Virus Genotype II. Front. Vet. Sci. 2019, 6, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hübner, A.; Petersen, B.; Keil, G.M.; Niemann, H.; Mettenleiter, T.C.; Fuchs, W. Efficient Inhibition of African Swine Fever Virus Replication by CRISPR/Cas9 Targeting of the Viral P30 Gene (CP204L). Sci. Rep. 2018, 8, 1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borca, M.V.; Holinka, L.G.; Berggren, K.A.; Gladue, D.P. CRISPR-Cas9, a Tool to Efficiently Increase the Development of Recombinant African Swine Fever Viruses. Sci. Rep. 2018, 8, 3154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE—World Organisation for Animal Health (Ed.) Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 8th ed.; OIE: Paris, France, 2018; ISBN 978-92-95108-18-9. [Google Scholar]
- Olasz, F.; Mészáros, I.; Marton, S.; Kaján, G.L.; Tamás, V.; Locsmándi, G.; Magyar, T.; Bálint, Á.; Bányai, K.; Zádori, Z. A Simple Method for Sample Preparation to Facilitate Efficient Whole-Genome Sequencing of African Swine Fever Virus. Viruses 2019, 11, 1129. [Google Scholar] [CrossRef] [Green Version]
- Corpet, F. Multiple Sequence Alignment with Hierarchical Clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]
- BLAST. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 18 June 2020).
- Zhao, Y.; O’Neil, N.J.; Rose, A.M. Poly-G/Poly-C Tracts in the Genomes of Caenorhabditis. BMC Genom. 2007, 8, 403. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.G.; Wellesley, F.C.; Savery, N.J.; Szczelkun, M.D. Length Heterogeneity at Conserved Sequence Block 2 in Human Mitochondrial DNA Acts as a Rheostat for RNA Polymerase POLRMT Activity. Nucleic Acids Res. 2016, 44, 7817–7829. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.S.; Hwang, C.K.; Song, K.Y.; Law, P.-Y.; Wei, L.-N.; Loh, H.H. Poly(C)-Binding Proteins as Transcriptional Regulators of Gene Expression. Biochem. Biophys. Res. Commun. 2009, 380, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Penza, V.; Russell, S.J.; Schulze, A.J. The Long-Lasting Enigma of Polycytidine (PolyC) Tract. PLoS Pathog. 2021, 17, e1009739. [Google Scholar] [CrossRef]
- Martin, L.R.; Neal, Z.C.; McBride, M.S.; Palmenberg, A.C. Mengovirus and Encephalomyocarditis Virus Poly(C) Tract Lengths Can Affect Virus Growth in Murine Cell Culture. J. Virol. 2000, 74, 3074–3081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cackett, G.; Portugal, R.; Matelska, D.; Dixon, L.; Werner, F. African Swine Fever Virus and Host Response: Transcriptome Profiling of the Georgia 2007/1 Strain and Porcine Macrophages. J. Virol. 2022, 96, e01939-21. [Google Scholar] [CrossRef] [PubMed]
- Vuono, E.A.; Ramirez-Medina, E.; Pruitt, S.; Rai, A.; Espinoza, N.; Silva, E.; Velazquez-Salinas, L.; Gladue, D.P.; Borca, M.V. Deletion of the ASFV DUTPase Gene E165R from the Genome of Highly Virulent African Swine Fever Virus Georgia 2010 Does Not Affect Virus Replication or Virulence in Domestic Pigs. Viruses 2022, 14, 1409. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Holinka, L.G.; Velazquez-Salinas, L.; Zhu, J.; Gladue, D.P. Development of a Highly Effective African Swine Fever Virus Vaccine by Deletion of the I177L Gene Results in Sterile Immunity against the Current Epidemic Eurasia Strain. J. Virol. 2020, 94, e02017-19. [Google Scholar] [CrossRef]
- Zhou, P.; Li, L.-F.; Zhang, K.; Wang, B.; Tang, L.; Li, M.; Wang, T.; Sun, Y.; Li, S.; Qiu, H.-J. Deletion of the H240R Gene of African Swine Fever Virus Decreases Infectious Progeny Virus Production Due to Aberrant Virion Morphogenesis and Enhances Inflammatory Cytokine Expression in Porcine Macrophages. J. Virol. 2022, 96, e01667-21. [Google Scholar] [CrossRef]
- Zhang, Y.; Ke, J.; Zhang, J.; Yang, J.; Yue, H.; Zhou, X.; Qi, Y.; Zhu, R.; Miao, F.; Li, Q.; et al. African Swine Fever Virus Bearing an I226R Gene Deletion Elicits Robust Immunity in Pigs to African Swine Fever. J. Virol. 2021, 95, e01199-21. [Google Scholar] [CrossRef]
- Ding, M.; Dang, W.; Liu, H.; Zhang, K.; Xu, F.; Tian, H.; Huang, H.; Shi, Z.; Sunkang, Y.; Qin, X.; et al. Sequential Deletions of Interferon Inhibitors MGF110-9L and MGF505-7R Result in Sterile Immunity against the Eurasia Strain of Africa Swine Fever. J. Virol. 2022, 96, e01192-22. [Google Scholar] [CrossRef]
- Rathakrishnan, A.; Connell, S.; Petrovan, V.; Moffat, K.; Goatley, L.C.; Jabbar, T.; Sánchez-Cordón, P.J.; Reis, A.L.; Dixon, L.K. Differential Effect of Deleting Members of African Swine Fever Virus Multigene Families 360 and 505 from the Genotype II Georgia 2007/1 Isolate on Virus Replication, Virulence, and Induction of Protection. J. Virol. 2022, 96, e01899-21. [Google Scholar] [CrossRef]
- Li, D.; Yang, W.; Li, L.; Li, P.; Ma, Z.; Zhang, J.; Qi, X.; Ren, J.; Ru, Y.; Niu, Q.; et al. African Swine Fever Virus MGF-505-7R Negatively Regulates CGAS–STING-Mediated Signaling Pathway. J. Immunol. 2021, 206, 1844–1857. [Google Scholar] [CrossRef]
- Mazur-Panasiuk, N.; Woźniakowski, G.; Niemczuk, K. The First Complete Genomic Sequences of African Swine Fever Virus Isolated in Poland. Sci. Rep. 2019, 9, 4556. [Google Scholar] [CrossRef] [Green Version]
- Dodantenna, N.; Ranathunga, L.; Chathuranga, W.A.G.; Weerawardhana, A.; Cha, J.-W.; Subasinghe, A.; Gamage, N.; Haluwana, D.K.; Kim, Y.; Jheong, W.; et al. African Swine Fever Virus EP364R and C129R Target Cyclic GMP-AMP To Inhibit the CGAS-STING Signaling Pathway. J. Virol. 2022, 96, e01022-22. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Nesvizhskii, A.I.; Rani, P.G.; Hahn, S.; Aebersold, R.; Ranish, J.A. The Transcription Elongation Factor TFIIS Is a Component of RNA Polymerase II Preinitiation Complexes. Proc. Natl. Acad. Sci. USA 2007, 104, 16068–16073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrés, G.; Simón-Mateo, C.; Viñuela, E. Assembly of African Swine Fever Virus: Role of Polyprotein Pp220. J. Virol. 1997, 71, 2331–2341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrés, G.; García-Escudero, R.; Salas, M.L.; Rodríguez, J.M. Repression of African Swine Fever Virus Polyprotein Pp220-Encoding Gene Leads to the Assembly of Icosahedral Core-Less Particles. J. Virol. 2002, 76, 2654–2666. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Covered and Amplified Region | Name of the Primer | Sequence |
|---|---|---|
| pUC19 + homology arm 1 | reg1_d11_F | 5′-ACGGCCAGTGAATTCGAGCTCGGTA CGTTATGTTGATAGTGTA-3′ |
| homology arm 1 + p72 promoter | reg1_ d11_R | 5′-ATATAATGTTATAAAAATAATTTATTGTT TTTATTAAATACGTATAAAGGGTTTAGTCATTAATAG-3′ |
| p72 promoter + eGFP | reg2_d11_F | 5′-TATTTAATAAAAACAATAAATTATT TTTATAACATTATATATGGTGAGCAAGGGCGAGGAGC-3′ |
| homology arm 2 + eGFP | reg2_d11_R | 5′-GCTACTCAGTCCTC ATTTTACTTGTACAGCTCGTCCATG-3′ |
| eGFP + homology arm 2 | reg3_d11_F | 5′-GCATGGACGAGCTG TACAAGTAAAATGAGGACTGAGTAGCC-3′ |
| pUC19 + homology arm 2 | reg3_d11_R | 5′-GGAAACAGCTATGACCATGAT TACGCCAAGCTTGCATGCCTCTATAAAGCAATACTGTC-3′ |
| Name of the Oligonucleotide | Sequence |
|---|---|
| CRISPR11_p72F | 5′-CACCG CTAAACCCTTTATACGCTGT-3′ |
| CRISPR11L_p72R | 5′-AAAC ACAGCGTATAAAGGGTTTAG C-3′ |
| CRISPR11L_endF | 5′-CACCG GTAATTCATGTGCAAGAATG-3′ |
| CRISPR11L_endR | 5′-AAAC CATTCTTGCACATGAATTAC C-3′ |
| Group | No. of Pigs | Vaccine Candidate/ Strain | Virus Concentration in FFU * in One Dose (2 mL) | Inoculation Route |
|---|---|---|---|---|
| Lv17d110-11L | 5 | Lv17/WB/Rie1/d110-11L | 102 | Intramuscular |
| Lv17 | 5 | Lv17/WB/Rie1 | 102 | Intramuscular |
| control | 3 | Unvaccinated controls | ||
| MGF 110-11L Gene | |
|---|---|
| Length of the Homopolymer G Region | Number of Isolates |
| 4 bp | 1 |
| 7 bp | 1 |
| 8 bp | 50 |
| 9 bp | 23 |
| 10 bp | 23 |
| 11 bp | 17 |
| 12 bp | 10 |
| 13 bp | 21 |
| 14 bp | 15 |
| 17 bp | 1 |
| 19 bp | 1 |
| Total: 163 | |
| Position in LV17/WB/Rie1 Genome | Change in d110-11L (Nucleotide) | Change in d110-11L (Amino Acid) | Gene | Other Information | Major Variants |
|---|---|---|---|---|---|
| 11,363 | C > T | - | NCR | promoter/regulator region of 285L gene | 95.9% |
| 38,598 | G > A | Val > Ile | MGF 505-5R | both the original and the substituted aa has an aliphatic side-chain | 95.6% |
| 76,238 | insertion of A | frameshift | EP364R | early termination, the length has reduced from 364 to 232 aa | 85.4% |
| 112,655 | C > A | Lys > Asn | G1340L | basic to polar aa | 70.3% |
| 115,161 | G > T | Gly > Cys | G1211R | apolar aa to polar aa | 92.4% |
| 119,295 | C > Y | Gly > Gly/Asp | CP2475L | ambiguous aa | C/T = 56.8/41.2% |
| 130,645 | C > Y | Ala > Ala/Thr | NP1450L | ambiguous aa | C/T = 55.4/42.4% |
| 134,614 | G > R | Ala >Ala/Val | NP419L | ambiguous aa | A/G = 55.6/42.9% |
| 166,363 | G > K | Pro > Pro/Gln | E146L | ambiguous aa | G/T = 41.1/53.1% |
| 171,591 | insertion of A | NCR | promoter/regulator region of the I267L gene | 73.7% | |
| 172,645 | deletion of T | frameshift | I243L | early termination, length has reduced from 243 to 158 aa | 70.0% |
| 176,321 | C > T | Asp > Asn | I196L | acidic to polar aa | 98.4% |
| 180,817 | insertion of T | frameshift | MGF 100-2L | early termination, the length has reduced from 141 to 80 aa | 69.7% |
| 185,263 | C > T | Glu > Glu | DP71L | synonymous mutation | 85.6% |
| Group | Pig # | Lymph Nodes † | Tonsils | Lungs | Spleen | Kidney | Heart | Skin †† | Intestine |
|---|---|---|---|---|---|---|---|---|---|
| Lv17d110-11L | 1 | - | - | - | - | - | - | - | - |
| 2 | - | - | +/d | - | - | +/l | - | - | |
| 3 | +/m | - | - | - | - | +/l, n | - | - | |
| 4 | +/m | - | - | - | - | - | - | - | |
| 5 | - | - | - | - | - | +/o | - | - | |
| Lv17 | 6 | +/a | - | - | +/b | - | - | +/h | - |
| 7 | +/a | - | - | +/b | - | - | - | - | |
| 8 | +/a | - | - | +/b | - | - | - | +/i | |
| 9 | +/a | - | - | - | +/e | - | - | - | |
| 10 | +/a | - | +/d | +/b | - | - | +/g | - | |
| control | 11 | +/a | - | - | +/b | +/e | +/e | - | - |
| 12 | +/a | +/f | - | +/b | +/e | +/e | - | - | |
| 13 | +/a | - | +/d | +/b | +/e | +/c | - | - |
| Strain/Controls | Days Post-Vaccination (DPV) | Days Post-Challenge (DPC) | ||||
|---|---|---|---|---|---|---|
| 0 | 7 | 14 | 0 * | 7 ** | 14 *** | |
| Lv17/WB/Rie1/d110-11L | - | - | 5 (70.7%) a,b | 5 (72.2%) | 5 (90.8%) | 5 (95.6%) |
| Lv17/WB/Rie1 | - | - | 5 (78.2%) | 3 (78.4%) c | 3 (89.8%) c | 3 (93.3%) c |
| Controls | - | - | - | - | - | 1 (80.37%) d |
| Strain | Days Post-Vaccination (DPV) | Days Post-Challenge (DPC) | ||||
|---|---|---|---|---|---|---|
| 0 | 7 | 14 | 0 * | 7 ** | 14 *** | |
| Lv17/WB/Rie1/d110-11L | - | 5 (35.2) a,b | 4 (37.4) | 1 (37.41) | - | - |
| Lv17/WB/Rie1 | - | 5 (32.7) | 3 (33.8) | 1 (36.5) c | - | - |
| Unvaccinated controls | - | - | - | - | 2 (24.2) | 2 (29.38) d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamás, V.; Righi, C.; Mészáros, I.; D’Errico, F.; Olasz, F.; Casciari, C.; Zádori, Z.; Magyar, T.; Petrini, S.; Feliziani, F. Involvement of the MGF 110-11L Gene in the African Swine Fever Replication and Virulence. Vaccines 2023, 11, 846. https://doi.org/10.3390/vaccines11040846
Tamás V, Righi C, Mészáros I, D’Errico F, Olasz F, Casciari C, Zádori Z, Magyar T, Petrini S, Feliziani F. Involvement of the MGF 110-11L Gene in the African Swine Fever Replication and Virulence. Vaccines. 2023; 11(4):846. https://doi.org/10.3390/vaccines11040846
Chicago/Turabian StyleTamás, Vivien, Cecilia Righi, István Mészáros, Federica D’Errico, Ferenc Olasz, Cristina Casciari, Zoltán Zádori, Tibor Magyar, Stefano Petrini, and Francesco Feliziani. 2023. "Involvement of the MGF 110-11L Gene in the African Swine Fever Replication and Virulence" Vaccines 11, no. 4: 846. https://doi.org/10.3390/vaccines11040846