
Immunotherapy of Equine Sarcoids—From Early Approaches to Innovative Vaccines


Abstract
:1. Introduction
2. Association of Bovine Papillomaviruses with Sarcoid Disease
3. BPV Immune Escape in the Equid Host
4. Immunotherapy of Sarcoids
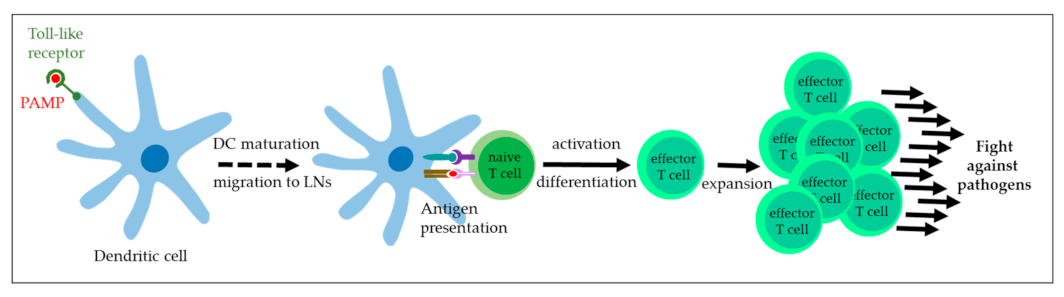
4.1. Toll-like Receptor Agonists
4.2. Immunostimulatory Cytokines
4.3. Recombinant Virus-like Particles
4.4. Autologous Vaccination
4.5. Influenza Virus Vector-Mediated Immunotherapy
5. Future and New Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knottenbelt, D.C. A suggested clinical classification for the equine sarcoid. Clin. Tech. Equine Pract. 2005, 4, 278–295. [Google Scholar] [CrossRef]
- Nasir, L.; Reid, S.W.J. Bovine papillomaviruses and equine sarcoids. In Papillomavirus Research: From Natural History to Vaccines and Beyond; Caister Academic Press: Norfolk, UK, 2006; Volume 1, pp. 389–397. [Google Scholar]
- Scott, D.W.; Miller, W.H., Jr. Sarcoid. Equine Dermatol. 2003, 1, 719–731. [Google Scholar]
- de Villiers, E.M.; Fauquet, C.; Broker, T.R.; Bernard, H.U.; zur Hausen, H. Classification of papillomaviruses. Virology 2004, 324, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Campo, M.S. Introduction. In Papillomavirus Research: From Natural History to Vaccines and Beyond; Caister Academic Press: Norfolk, UK, 2006; pp. 1–2. [Google Scholar]
- Campo, M.S. Bovine papillomavirus: Old system, new lessons? In Papillomavirus Research: From Natural History to Vaccines and Beyond; Caister Academic Press: Norfolk, UK, 2006; pp. 373–387. [Google Scholar]
- Olthof, N.C.; Huebbers, C.U.; Kolligs, J.; Henfling, M.; Ramaekers, F.C.; Cornet, I.; van Lent-Albrechts, J.A.; Stegmann, A.P.; Silling, S.; Wieland, U.; et al. Viral load, gene expression and mapping of viral integration sites in HPV16-associated HNSCC cell lines. Int. J. Cancer 2015, 136, E207–E218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pett, M.; Coleman, N. Integration of high-risk human papillomavirus: A key event in cervical carcinogenesis? J. Pathol. 2007, 212, 356–367. [Google Scholar] [CrossRef]
- Walline, H.M.; Komarck, C.M.; McHugh, J.B.; Bellile, E.L.; Brenner, J.C.; Prince, M.E.; McKean, E.L.; Chepeha, D.B.; Wolf, G.T.; Worden, F.P.; et al. Genomic Integration of High-Risk HPV Alters Gene Expression in Oropharyngeal Squamous Cell Carcinoma. Mol. Cancer Res. 2016, 14, 941–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, C., Jr.; Cook, R.H. Cutaneous sarcoma-like lesions of the horse caused by the agent of bovine papilloma. Exp. Biol. Med. 1951, 77, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Chambers, G.; Ellsmore, V.A.; O’Brien, P.M.; Reid, S.W.; Love, S.; Campo, M.S.; Nasir, L. Association of bovine papillomavirus with the equine sarcoid. J. Gen. Virol. 2003, 84, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Lunardi, M.; de Alcantara, B.K.; Otonel, R.A.; Rodrigues, W.B.; Alfieri, A.F.; Alfieri, A.A. Bovine papillomavirus type 13 DNA in equine sarcoids. J. Clin. Microbiol. 2013, 51, 2167–2171. [Google Scholar] [CrossRef] [Green Version]
- Jindra, C.; Kamjunke, A.K.; Jones, S.; Brandt, S. Screening for bovine papillomavirus type 13 (BPV13) in a European population of sarcoid-bearing equids. Equine Vet. J. 2021, 54, 662–669. [Google Scholar] [CrossRef]
- Amtmann, E.; Muller, H.; Sauer, G. Equine connective tissue tumors contain unintegrated bovine papilloma virus DNA. J. Virol. 1980, 35, 962–964. [Google Scholar] [CrossRef] [Green Version]
- Bogaert, L.; Martens, A.; Kast, W.M.; Van Marck, E.; De Cock, H. Bovine papillomavirus DNA can be detected in keratinocytes of equine sarcoid tumors. Vet. Microbiol. 2010, 146, 269–275. [Google Scholar] [CrossRef]
- Brandt, S.; Haralambus, R.; Shafti-Keramat, S.; Steinborn, R.; Stanek, C.; Kirnbauer, R. A subset of equine sarcoids harbours BPV-1 DNA in a complex with L1 major capsid protein. Virology 2008, 375, 433–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, A.D.; Armstrong, E.L.; Gofton, R.G.; Mason, J.; De Toit, N.; Day, M.J. Characterisation of early and late bovine papillomavirus protein expression in equine sarcoids. Vet. Microbiol. 2013, 162, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Bogaert, L.; Martens, A.; Van Poucke, M.; Ducatelle, R.; De Cock, H.; Dewulf, J.; De Baere, C.; Peelman, L.; Gasthuys, F. High prevalence of bovine papillomaviral DNA in the normal skin of equine sarcoid-affected and healthy horses. Vet. Microbiol. 2008, 129, 58–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, E.A.; Theon, A.P.; Madewell, B.R.; Griffey, S.M.; Hitchcock, M.E. Bovine papillomavirus DNA in neoplastic and nonneoplastic tissues obtained from horses with and without sarcoids in the western United States. Am. J. Vet. Res. 2001, 62, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Martens, A.; De Moor, A.; Ducatelle, R. PCR detection of bovine papilloma virus DNA in superficial swabs and scrapings from equine sarcoids. Vet. J. 2001, 161, 280–286. [Google Scholar] [CrossRef]
- Trenfield, K.; Spradbrow, P.B.; Vanselow, B. Sequences of papillomavirus DNA in equine sarcoids. Equine Vet. J. 1985, 17, 449–452. [Google Scholar] [CrossRef]
- Knottenbelt, D.C. Sarcoid. In Pascoe’s Principles and Practice of Equine Dermatology; Knottenbelt, D.C., Ed.; Saunders Elsevier: London, UK, 2009; pp. 387–407. [Google Scholar]
- Knottenbelt, D.C. The Equine Sarcoid: Why Are There so Many Treatment Options? Vet. Clin. N. Am. Equine Pract. 2019, 35, 243–262. [Google Scholar] [CrossRef]
- Tarwid, J.N.; Fretz, P.B.; Clark, E.G. Equine sarcoids: A study with emphasis on pathological diagnosis. Compend. Contin. Educ. Pract. Vet. 1985, 7, 293–300. [Google Scholar]
- Chow, L.T.; Broker, T.R. Mechanisms and regulation of papillomavirus DNA replication. In Papillomavirus Research: From Natural History to Vaccines and Beyond; Caister Academic Press: Norfolk, UK, 2006; pp. 53–71. [Google Scholar]
- Hainisch, E.K.; Jindra, C.; Reicher, P.; Miglinci, L.; Brodesser, D.M.; Brandt, S. Bovine Papillomavirus Type 1 or 2 Virion-Infected Primary Fibroblasts Constitute a Near-Natural Equine Sarcoid Model. Viruses 2022, 14, 2658. [Google Scholar] [CrossRef]
- Yuan, Z.; Gault, E.A.; Campo, M.S.; Nasir, L. Different contribution of bovine papillomavirus type 1 oncoproteins to the transformation of equine fibroblasts. J. Gen. Virol. 2011, 92, 773–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.Q.; Gault, E.A.; Gobeil, P.; Nixon, C.; Campo, M.S.; Nasir, L. Establishment and characterization of equine fibroblast cell lines transformed in vivo and in vitro by BPV-1: Model systems for equine sarcoids. Virology 2008, 373, 352–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borzacchiello, G.; Iovane, G.; Marcante, M.L.; Poggiali, F.; Roperto, F.; Roperto, S.; Venuti, A. Presence of bovine papillomavirus type 2 DNA and expression of the viral oncoprotein E5 in naturally occurring urinary bladder tumours in cows. J. Gen. Virol. 2003, 84, 2921–2926. [Google Scholar] [CrossRef]
- Klingelhutz, A.J.; Foster, S.A.; McDougall, J.K. Telomerase activation by the E6 gene product of human papillomavirus type 16. Nature 1996, 380, 79–82. [Google Scholar] [CrossRef]
- Petti, L.; DiMaio, D. Specific interaction between the bovine papillomavirus E5 transforming protein and the beta receptor for platelet-derived growth factor in stably transformed and acutely transfected cells. J. Virol. 1994, 68, 3582–3592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suprynowicz, F.A.; Campo, M.S.; Schlegel, R. Biologic activities of papillomavirus E5 proteins. In Papillomavirus Research: From Natural History to Vaccines and Beyond; Caister Academic Press: Norfolk, UK, 2006; Volume 1, pp. 97–114. [Google Scholar]
- Tong, X.; Howley, P.M. The bovine papillomavirus E6 oncoprotein interacts with paxillin and disrupts the actin cytoskeleton. Proc. Natl. Acad. Sci. USA 1997, 94, 4412–4417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeMasi, J.; Huh, K.W.; Nakatani, Y.; Munger, K.; Howley, P.M. Bovine papillomavirus E7 transformation function correlates with cellular p600 protein binding. Proc. Natl. Acad. Sci. USA 2005, 102, 11486–11491. [Google Scholar] [CrossRef] [Green Version]
- Brandt, S. Immune response to bovine papillomavirus type 1 in equine sarcoid. Vet. J. 2016, 216, 107–108. [Google Scholar] [CrossRef]
- Hainisch, E.K.; Abel-Reichwald, H.; Shafti-Keramat, S.; Pratscher, B.; Corteggio, A.; Borzacchiello, G.; Wetzig, M.; Jindra, C.; Tichy, A.; Kirnbauer, R.; et al. Potential of a BPV1 L1 VLP vaccine to prevent BPV1- or BPV2-induced pseudo-sarcoid formation and safety and immunogenicity of EcPV2 L1 VLPs in the horse. J. Gen. Virol. 2016, 98, 230–241. [Google Scholar] [CrossRef]
- Hainisch, E.K.; Brandt, S.; Shafti-Keramat, S.; Van den Hoven, R.; Kirnbauer, R. Safety and immunogenicity of BPV-1 L1 virus-like particles in a dose-escalation vaccination trial in horses. Equine Vet. J. 2012, 44, 107–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartl, B.; Hainisch, E.K.; Shafti-Keramat, S.; Kirnbauer, R.; Corteggio, A.; Borzacchiello, G.; Tober, R.; Kainzbauer, C.; Pratscher, B.; Brandt, S. Inoculation of young horses with bovine papillomavirus type 1 virions leads to early infection of PBMCs prior to pseudo-sarcoid formation. J. Gen. Virol. 2011, 92, 2437–2445. [Google Scholar] [CrossRef] [PubMed]
- Ragland, W.L.; Spencer, G.R. Attempts to relate bovine papilloma virus to the cause of equine sarcoid: Immunity to bovine papilloma virus. Am. J. Vet. Res. 1968, 29, 1363–1366. [Google Scholar] [PubMed]
- Ragland, W.L.; Spencer, G.R. Attempts to relate bovine papilloma virus to the cause of equine sarcoid: Equidae inoculated intradermally with bovine papilloma virus. Am. J. Vet. Res. 1969, 30, 743–752. [Google Scholar]
- Voss, J.L. Transmission of equine sarcoid. Am. J. Vet. Res. 1969, 30, 183–191. [Google Scholar] [PubMed]
- Stanley, M. Host defence and persistent human papillomavirus infection. Curr. Opin. Virol. 2021, 51, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.Q.; Bennett, L.; Campo, M.S.; Nasir, L. Bovine papillomavirus type 1 E2 and E7 proteins down-regulate Toll Like Receptor 4 (TLR4) expression in equine fibroblasts. Virus Res. 2010, 149, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.Q.; Nicolson, L.; Marchetti, B.; Gault, E.A.; Campo, M.S.; Nasir, L. Transcriptional changes induced by bovine papillomavirus type 1 in equine fibroblasts. J. Virol. 2008, 82, 6481–6491. [Google Scholar] [CrossRef] [Green Version]
- Nasir, L.; Brandt, S. Papillomavirus associated diseases of the horse. Vet. Microbiol. 2013, 167, 159–167. [Google Scholar] [CrossRef]
- Marchetti, B.; Gault, E.A.; Cortese, M.S.; Yuan, Z.; Ellis, S.A.; Nasir, L.; Campo, M.S. Bovine papillomavirus type 1 oncoprotein E5 inhibits equine MHC class I and interacts with equine MHC I heavy chain. J. Gen. Virol. 2009, 90, 2865–2870. [Google Scholar] [CrossRef]
- Steinbach, A.; Riemer, A.B. Immune evasion mechanisms of human papillomavirus: An update. Int. J. Cancer 2018, 142, 224–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Falco, F.; Cutarelli, A.; Catoi, A.F.; Uberti, B.D.; Cuccaro, B.; Roperto, S. Bovine delta papillomavirus E5 oncoprotein negatively regulates the cGAS-STING signaling pathway in cattle in a spontaneous model of viral disease. Front. Immunol. 2022, 13, 937736. [Google Scholar] [CrossRef]
- De Falco, F.; Cutarelli, A.; Gentile, I.; Cerino, P.; Uleri, V.; Catoi, A.F.; Roperto, S. Bovine Delta Papillomavirus E5 Oncoprotein Interacts with TRIM25 and Hampers Antiviral Innate Immune Response Mediated by RIG-I-Like Receptors. Front. Immunol. 2021, 12, 658762. [Google Scholar] [CrossRef]
- Brostrom, H. Equine sarcoids. A clinical and epidemiological study in relation to equine leucocyte antigens (ELA). Acta Vet. Scand. 1995, 36, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Broström, H.; Fahlbrink, E.; Dubath, M.L.; Lazary, S. Association between equine leucocyte antigens (ELA) and equine sarcoid tumors in the population of Swedish halfbreds and some of their families. Vet. Immunol. Immunopathol. 1988, 19, 215–223. [Google Scholar] [CrossRef]
- Meredith, D.; Elser, A.H.; Wolf, B.; Soma, L.R.; Donawick, W.J.; Lazary, S. Equine leukocyte antigens: Relationships with sarcoid tumors and laminitis in two pure breeds. Immunogenetics 1986, 23, 221–225. [Google Scholar] [CrossRef]
- Triulzi, T.; Tagliabue, E.; Balsari, A.; Casalini, P. FOXP3 expression in tumor cells and implications for cancer progression. J. Cell Physiol. 2013, 228, 30–35. [Google Scholar] [CrossRef]
- Gajewski, T.F. The expanding universe of regulatory T cell subsets in cancer. Immunity 2007, 27, 185–187. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ma, Y.; Fang, Y.; Wu, S.; Liu, L.; Fu, D.; Shen, X. Regulatory T cell: A protection for tumour cells. J. Cell. Mol. Med. 2012, 16, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.D.; Hicks, C. Both tumour cells and infiltrating T-cells in equine sarcoids express FOXP3 associated with an immune-supressed cytokine microenvironment. Vet. Res. 2016, 47, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisshusler, H.; Marti, E.; Stoffel, M.H.; Kuhni, K.; Stojiljkovic, A.; von Tscharner, C.; Vidondo, B.; Gerber, V.; Koch, C. Quantitative analysis of infiltrating immune cells and bovine papillomavirus type 1 E2-positive cells in equine sarcoids. Vet. J. 2016, 216, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Ebert, L.M.; Tan, B.S.; Browning, J.; Svobodova, S.; Russell, S.E.; Kirkpatrick, N.; Gedye, C.; Moss, D.; Ng, S.P.; MacGregor, D.; et al. The regulatory T cell-associated transcription factor FoxP3 is expressed by tumor cells. Cancer Res. 2008, 68, 3001–3009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Q.; Zhang, S.; Wei, H.; Pang, X.; Zhang, H. Roles of Foxp3 in the occurrence and development of cervical cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 8717–8730. [Google Scholar] [PubMed]
- Wang, X.; Lang, M.; Zhao, T.; Feng, X.; Zheng, C.; Huang, C.; Hao, J.; Dong, J.; Luo, L.; Li, X.; et al. Cancer-FOXP3 directly activated CCL5 to recruit FOXP3(+)Treg cells in pancreatic ductal adenocarcinoma. Oncogene 2017, 36, 3048–3058. [Google Scholar] [CrossRef]
- Ashrafi, G.H.; Piuko, K.; Burden, F.; Yuan, Z.; Gault, E.A.; Muller, M.; Trawford, A.; Reid, S.W.; Nasir, L.; Campo, M.S. Vaccination of sarcoid-bearing donkeys with chimeric virus-like particles of bovine papillomavirus type 1. J. Gen. Virol. 2008, 89, 148–157. [Google Scholar] [CrossRef]
- Vacchelli, E.; Aranda, F.; Obrist, F.; Eggermont, A.; Galon, J.; Cremer, I.; Zitvogel, L.; Kroemer, G.; Galluzzi, L. Trial watch: Immunostimulatory cytokines in cancer therapy. Oncoimmunology 2014, 3, e29030. [Google Scholar] [CrossRef] [Green Version]
- Mattil-Fritz, S.; Scharner, D.; Piuko, K.; Thones, N.; Gissmann, L.; Muller, H.; Muller, M. Immunotherapy of equine sarcoid: Dose-escalation trial for the use of chimeric papillomavirus-like particles. J. Gen. Virol. 2008, 89, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Jindra, C.; Hainisch, E.K.; Rummele, A.; Wolschek, M.; Muster, T.; Brandt, S. Influenza virus vector iNS1 expressing bovine papillomavirus 1 (BPV1) antigens efficiently induces tumour regression in equine sarcoid patients. PLoS ONE 2021, 16, e0260155. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef] [Green Version]
- Dajon, M.; Iribarren, K.; Cremer, I. Toll-like receptor stimulation in cancer: A pro- and anti-tumor double-edged sword. Immunobiology 2017, 222, 89–100. [Google Scholar] [CrossRef]
- Calmette, A. Preventive Vaccination Against Tuberculosis with BCG. Proc. R. Soc. Med. 1931, 24, 1481–1490. [Google Scholar] [CrossRef] [Green Version]
- Cardillo, F.; Bonfim, M.; da Silva Vasconcelos Sousa, P.; Mengel, J.; Ribeiro Castello-Branco, L.R.; Pinho, R.T. Bacillus Calmette-Guerin Immunotherapy for Cancer. Vaccines 2021, 9, 439. [Google Scholar] [CrossRef]
- Murphy, J.M.; Severin, G.A.; Lavach, J.D.; Hepler, D.I.; Lueker, D.C. Immunotherapy in ocular equine sarcoid. J. Am. Vet. Med. Assoc. 1979, 174, 269–272. [Google Scholar] [PubMed]
- Wyman, M.; Rings, M.D.; Tarr, M.J.; Alden, C.L. Immunotherapy in equine sarcoid: A report of two cases. J. Am. Vet. Med. Assoc. 1977, 171, 751–779. [Google Scholar]
- Klein, W.R.; Bras, G.E.; Misdorp, W.; Steerenberg, P.A.; De Jong, W.H.; Tiesjema, R.H.; Kersjes, A.W.; Ruitenberg, E.J. Equine sarcoid: BCG immunotherapy compared to cryosurgery in a prospective randomised clinical trial. Cancer Immunol. Immunother. 1986, 21, 133–140. [Google Scholar] [CrossRef]
- Knottenbelt, D.C.; Kelly, D.F. The diagnosis and treatment of periorbital sarcoid in the horse: 445 cases from 1974 to 1999. Vet. Ophthalmol. 2000, 3, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Martens, A.; De Moor, A.; Vlaminck, L.; Pille, F.; Steenhaut, M. Evaluation of excision, cryosurgery and local BCG vaccination for the treatment of equine sarcoids. Vet. Rec. 2001, 149, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Komaromy, A.M.; Andrew, S.E.; Brooks, D.E.; Detrisac, C.J.; Gelatt, K.N. Periocular sarcoid in a horse. Vet. Ophthalmol. 2004, 7, 141–146. [Google Scholar] [CrossRef]
- Lavach, J.D.; Severin, G.A.; Lueker, D. Immunotherapy of periocular sarcoids in horses. Vet. Clin. N. Am. Large Anim. Pract. 1984, 6, 513–518. [Google Scholar] [CrossRef]
- Owen, R.A.; Jagger, D.W. Clinical observations on the use of BCG cell wall fraction for treatment of periocular and other equine sarcoids. Vet. Rec. 1987, 120, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Klein, W.R. Immunotherapy of squamous cell carcinoma of the bovine eye and of equine sarcoid. Tijdschr Diergeneeskd 1990, 115, 1149–1155. [Google Scholar] [PubMed]
- EMA. List of Nationally Authorised Medicinal Products—Active Substance: BCG Vaccine, PSUS_A/00000304/201803; European Medicines Agency: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Kollipara, R.; Ekhlassi, E.; Downing, C.; Guidry, J.; Lee, M.; Tyring, S.K. Advancements in Pharmacotherapy for Noncancerous Manifestations of HPV. J. Clin. Med. 2015, 4, 832–846. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, S.A.; Torres, S.M.; Malone, E.D.; Diaz, S.F.; Jessen, C.; Gilbert, S. Efficacy of imiquimod 5% cream in the treatment of equine sarcoids: A pilot study. Vet. Dermatol. 2006, 17, 259–265. [Google Scholar] [CrossRef]
- Pettersson, C.M.; Brostrom, H.; Humblot, P.; Bergvall, K.E. Topical treatment of equine sarcoids with imiquimod 5% cream or Sanguinaria canadensis and zinc chloride—An open prospective study. Vet. Dermatol. 2020, 31, 471-e126. [Google Scholar] [CrossRef] [PubMed]
- Haspeslagh, M.; Vlaminck, L.E.; Martens, A.M. Treatment of sarcoids in equids: 230 cases (2008–2013). J. Am. Vet. Med. Assoc. 2016, 249, 311–318. [Google Scholar] [CrossRef]
- Barros, M.R., Jr.; de Oliveira, T.H.A.; de Melo, C.M.L.; Venuti, A.; de Freitas, A.C. Viral Modulation of TLRs and Cytokines and the Related Immunotherapies for HPV-Associated Cancers. J. Immunol. Res. 2018, 2018, 2912671. [Google Scholar] [CrossRef]
- Jebreel, A.; Mistry, D.; Loke, D.; Dunn, G.; Hough, V.; Oliver, K.; Stafford, N.; Greenman, J. Investigation of interleukin 10, 12 and 18 levels in patients with head and neck cancer. J. Laryngol. Otol. 2007, 121, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Peghini, B.C.; Abdalla, D.R.; Barcelos, A.C.; Teodoro, L.; Murta, E.F.; Michelin, M.A. Local cytokine profiles of patients with cervical intraepithelial and invasive neoplasia. Hum. Immunol. 2012, 73, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Torres-Poveda, K.; Bahena-Roman, M.; Madrid-Gonzalez, C.; Burguete-Garcia, A.I.; Bermudez-Morales, V.H.; Peralta-Zaragoza, O.; Madrid-Marina, V. Role of IL-10 and TGF-beta1 in local immunosuppression in HPV-associated cervical neoplasia. World J. Clin. Oncol. 2014, 5, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Smola-Hess, S.; Pfister, H. Immune evasion in genital papillomavirus infection and cervical cancer: Role of cytokines and chemokines. In Papillomavirus Research—From Natural History to Vaccines and Beyond; Caister Academic Press: Norfolk, UK, 2006; pp. 321–339. [Google Scholar]
- Klein, C.; Waldhauer, I.; Nicolini, V.G.; Freimoser-Grundschober, A.; Nayak, T.; Vugts, D.J.; Dunn, C.; Bolijn, M.; Benz, J.; Stihle, M.; et al. Cergutuzumab amunaleukin (CEA-IL2v), a CEA-targeted IL-2 variant-based immunocytokine for combination cancer immunotherapy: Overcoming limitations of aldesleukin and conventional IL-2-based immunocytokines. Oncoimmunology 2017, 6, e1277306. [Google Scholar] [CrossRef] [PubMed]
- Paradkar, P.H.; Joshi, J.V.; Mertia, P.N.; Agashe, S.V.; Vaidya, R.A. Role of cytokines in genesis, progression and prognosis of cervical cancer. Asian Pac. J. Cancer Prev. 2014, 15, 3851–3864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrangle, J.M.; Patterson, A.; Johnson, C.B.; Neitzke, D.J.; Mehrotra, S.; Denlinger, C.E.; Paulos, C.M.; Li, Z.; Cole, D.J.; Rubinstein, M.P. IL-2 and Beyond in Cancer Immunotherapy. J. Interferon Cytokine Res. 2018, 38, 45–68. [Google Scholar] [CrossRef] [PubMed]
- Den Otter, W.; Hill, F.W.; Klein, W.R.; Koten, J.W.; Steerenberg, P.A.; De Mulder, P.H.; Rhode, C.; Stewart, R.; Faber, J.A.; Ruitenberg, E.J.; et al. Therapy of bovine ocular squamous-cell carcinoma with local doses of interleukin-2: 67% complete regressions after 20 months of follow-up. Cancer Immunol. Immunother. 1995, 41, 10–14. [Google Scholar] [CrossRef]
- Spoormakers, T.J.; Klein, W.R.; Jacobs, J.J.; Van Den Ingh, T.S.; Koten, J.W.; Den Otter, W. Comparison of the efficacy of local treatment of equine sarcoids with IL-2 or cisplatin/IL-2. Cancer Immunol. Immunother. 2003, 52, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Loschelder-Ostrowski, J.; Winter, J.C.; Merle, R.; Klopfleisch, R.; Gehlen, H. Treatment of equine sarcoids using recombinant poxviruses expressing feline interleukin-2. Vet. Dermatol. 2021, 32, 283-e277. [Google Scholar] [CrossRef]
- Saba, C.; Eggleston, R.; Parks, A.; Peroni, J.; Sjoberg, E.; Rice, S.; Tyma, J.; Williams, J.; Grosenbaugh, D.; Leard, A.T. ALVAC-fIL2, a feline interleukin-2 immunomodulator, as a treatment for sarcoids in horses: A pilot study. J. Vet. Intern. Med. 2022, 36, 1179–1184. [Google Scholar] [CrossRef]
- Kirnbauer, R. Papillomavirus-like particles for serology and vaccine development. Intervirology 1996, 39, 54–61. [Google Scholar] [CrossRef]
- Schiller, J.T. Papillomavirus Vaccines. Papillomaviruses 2007, 1, 337–369. [Google Scholar]
- Kirnbauer, R.; Chandrachud, L.M.; O’Neil, B.W.; Wagner, E.R.; Grindlay, G.J.; Armstrong, A.; McGarvie, G.M.; Schiller, J.T.; Lowy, D.R.; Campo, M.S. Virus-like particles of bovine papillomavirus type 4 in prophylactic and therapeutic immunization. Virology 1996, 219, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Espy, B.M.K. How to Treat Equine Sarcoids by Autologous Implantation. AAEP Proc. 2008, 54, 68–73. [Google Scholar]
- Rothacker, C.C.; Boyle, A.G.; Levine, D.G. Autologous vaccination for the treatment of equine sarcoids: 18 cases (2009–2014). Can. Vet. J. 2015, 56, 709–714. [Google Scholar] [PubMed]
- Figge, G. Multimodale, Kombinierte Tumortherapie und deren Behandlungserfolg bei hochgradigen Equinen Sarkoiden an der Pferdeklinik der Veterinärmedizinischen Universität Wien von 2001–2020. Master Thesis, Veterinary University, Vienna, Austria, 2022. [Google Scholar]
- Garcia-Sastre, A.; Egorov, A.; Matassov, D.; Brandt, S.; Levy, D.E.; Durbin, J.E.; Palese, P.; Muster, T. Influenza A virus lacking the NS1 gene replicates in interferon-deficient systems. Virology 1998, 252, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egorov, A.; Brandt, S.; Sereinig, S.; Romanova, J.; Ferko, B.; Katinger, D.; Grassauer, A.; Alexandrova, G.; Katinger, H.; Muster, T. Transfectant influenza A viruses with long deletions in the NS1 protein grow efficiently in Vero cells. J. Virol. 1998, 72, 6437–6441. [Google Scholar] [CrossRef] [Green Version]
- Romanova, J.; Krenn, B.M.; Wolschek, M.; Ferko, B.; Romanovskaja-Romanko, E.; Morokutti, A.; Shurygina, A.P.; Nakowitsch, S.; Ruthsatz, T.; Kiefmann, B.; et al. Preclinical evaluation of a replication-deficient intranasal DeltaNS1 H5N1 influenza vaccine. PLoS ONE 2009, 4, e5984. [Google Scholar] [CrossRef] [PubMed]
- Wacheck, V.; Egorov, A.; Groiss, F.; Pfeiffer, A.; Fuereder, T.; Hoeflmayer, D.; Kundi, M.; Popow-Kraupp, T.; Redlberger-Fritz, M.; Mueller, C.A.; et al. A novel type of influenza vaccine: Safety and immunogenicity of replication-deficient influenza virus created by deletion of the interferon antagonist NS1. J. Infect. Dis. 2010, 201, 354–362. [Google Scholar] [CrossRef]
- Efferson, C.L.; Tsuda, N.; Kawano, K.; Nistal-Villan, E.; Sellappan, S.; Yu, D.; Murray, J.L.; Garcia-Sastre, A.; Ioannides, C.G. Prostate tumor cells infected with a recombinant influenza virus expressing a truncated NS1 protein activate cytolytic CD8+ cells to recognize noninfected tumor cells. J. Virol. 2006, 80, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Ogbomo, H.; Michaelis, M.; Geiler, J.; van Rikxoort, M.; Muster, T.; Egorov, A.; Doerr, H.W.; Cinatl, J., Jr. Tumor cells infected with oncolytic influenza A virus prime natural killer cells for lysis of resistant tumor cells. Med. Microbiol. Immunol. 2010, 199, 93–101. [Google Scholar] [CrossRef]
- Ferko, B.; Kittel, C.; Romanova, J.; Sereinig, S.; Katinger, H.; Egorov, A. Live attenuated influenza virus expressing human interleukin-2 reveals increased immunogenic potential in young and aged hosts. J. Virol. 2006, 80, 11621–11627. [Google Scholar] [CrossRef] [Green Version]
- Kittel, C.; Ferko, B.; Kurz, M.; Voglauer, R.; Sereinig, S.; Romanova, J.; Stiegler, G.; Katinger, H.; Egorov, A. Generation of an influenza A virus vector expressing biologically active human interleukin-2 from the NS gene segment. J. Virol. 2005, 79, 10672–10677. [Google Scholar] [CrossRef] [Green Version]
- Sereinig, S.; Stukova, M.; Zabolotnyh, N.; Ferko, B.; Kittel, C.; Romanova, J.; Vinogradova, T.; Katinger, H.; Kiselev, O.; Egorov, A. Influenza virus NS vectors expressing the mycobacterium tuberculosis ESAT-6 protein induce CD4+ Th1 immune response and protect animals against tuberculosis challenge. Clin. Vaccine Immunol. 2006, 13, 898–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Rikxoort, M.; Michaelis, M.; Wolschek, M.; Muster, T.; Egorov, A.; Seipelt, J.; Doerr, H.W.; Cinatl, J., Jr. Oncolytic effects of a novel influenza A virus expressing interleukin-15 from the NS reading frame. PLoS ONE 2012, 7, e36506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolschek, M.; Samm, E.; Seper, H.; Sturlan, S.; Kuznetsova, I.; Schwager, C.; Khassidov, A.; Kittel, C.; Muster, T.; Egorov, A.; et al. Establishment of a chimeric, replication-deficient influenza A virus vector by modulation of splicing efficiency. J. Virol. 2011, 85, 2469–2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jindra, C.; Huber, B.; Shafti-Keramat, S.; Wolschek, M.; Ferko, B.; Muster, T.; Brandt, S.; Kirnbauer, R. Attenuated Recombinant Influenza A Virus Expressing HPV16 E6 and E7 as a Novel Therapeutic Vaccine Approach. PLoS ONE 2015, 10, e0138722. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
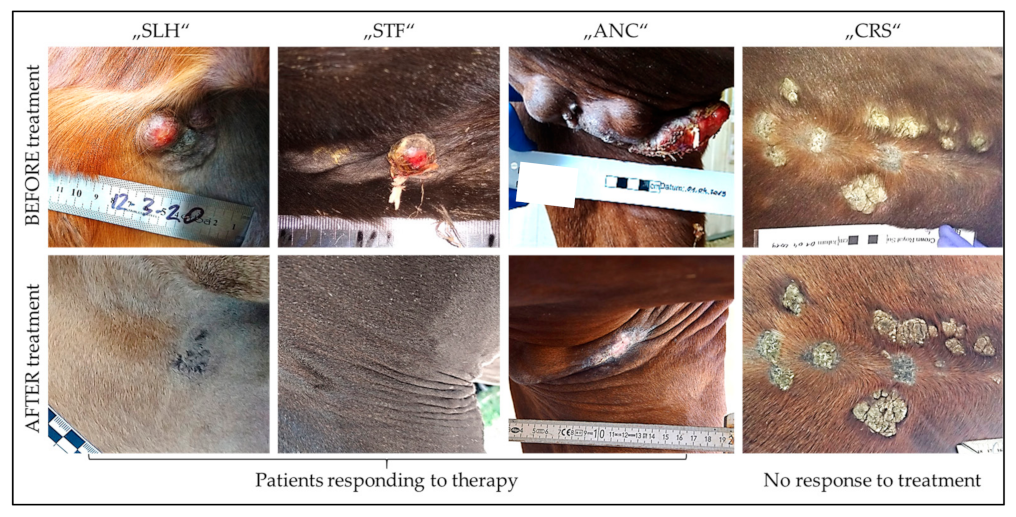
| Treatment Schedule | Used Vaccines | Number and Types of Cases | Therapeutic Outcome | |||
|---|---|---|---|---|---|---|
| COR | PAR | STD | PRD | |||
| Days 1, 3, 5/8, 10, 12 | AAA/BBB | 7 severe | 2/7 | - | 2/7 | 3/7 |
| 1 moderate | 1/1 | - | - | - | ||
| Every second month on average until COR or no further improvement | AAA | 4 severe | 1/4 | 1/4 | - | 2/4 |
| 4 moderate | 2/2 | 2/2 § | - | - | ||
| 2 mild | 2/2 | - | - | - | ||
| AA, then B * | 3 severe | 1/3 | 1/3 | - | 1/3 | |
| 1 mild | 1/1 | - | - | - | ||
| BBB | 3 severe | 1/3 | - | 2/3 | - | |
| BB, then A * | 2 severe | 1/2 | - | 1/2 | - | |
| 2 mild | 2/2 | - | - | - | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jindra, C.; Hainisch, E.K.; Brandt, S. Immunotherapy of Equine Sarcoids—From Early Approaches to Innovative Vaccines. Vaccines 2023, 11, 769. https://doi.org/10.3390/vaccines11040769
Jindra C, Hainisch EK, Brandt S. Immunotherapy of Equine Sarcoids—From Early Approaches to Innovative Vaccines. Vaccines. 2023; 11(4):769. https://doi.org/10.3390/vaccines11040769
Chicago/Turabian StyleJindra, Christoph, Edmund K. Hainisch, and Sabine Brandt. 2023. "Immunotherapy of Equine Sarcoids—From Early Approaches to Innovative Vaccines" Vaccines 11, no. 4: 769. https://doi.org/10.3390/vaccines11040769