
Tracking B Cell Memory to SARS-CoV-2 Using Rare Cell Analysis System


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Vaccinated Subjects
2.2. Peripheral Blood Sample Processing
2.3. Detecting SARS-CoV-2 Binding and Neutralizing Antibody
2.4. Detecting SARS-CoV-2 Spike-RBD Specific Memory B Cells
2.5. In Vitro Differentiation of SRAS-CoV-2 Spike-RBD-Specific Memory B Cells to Antibody Secreting Cells
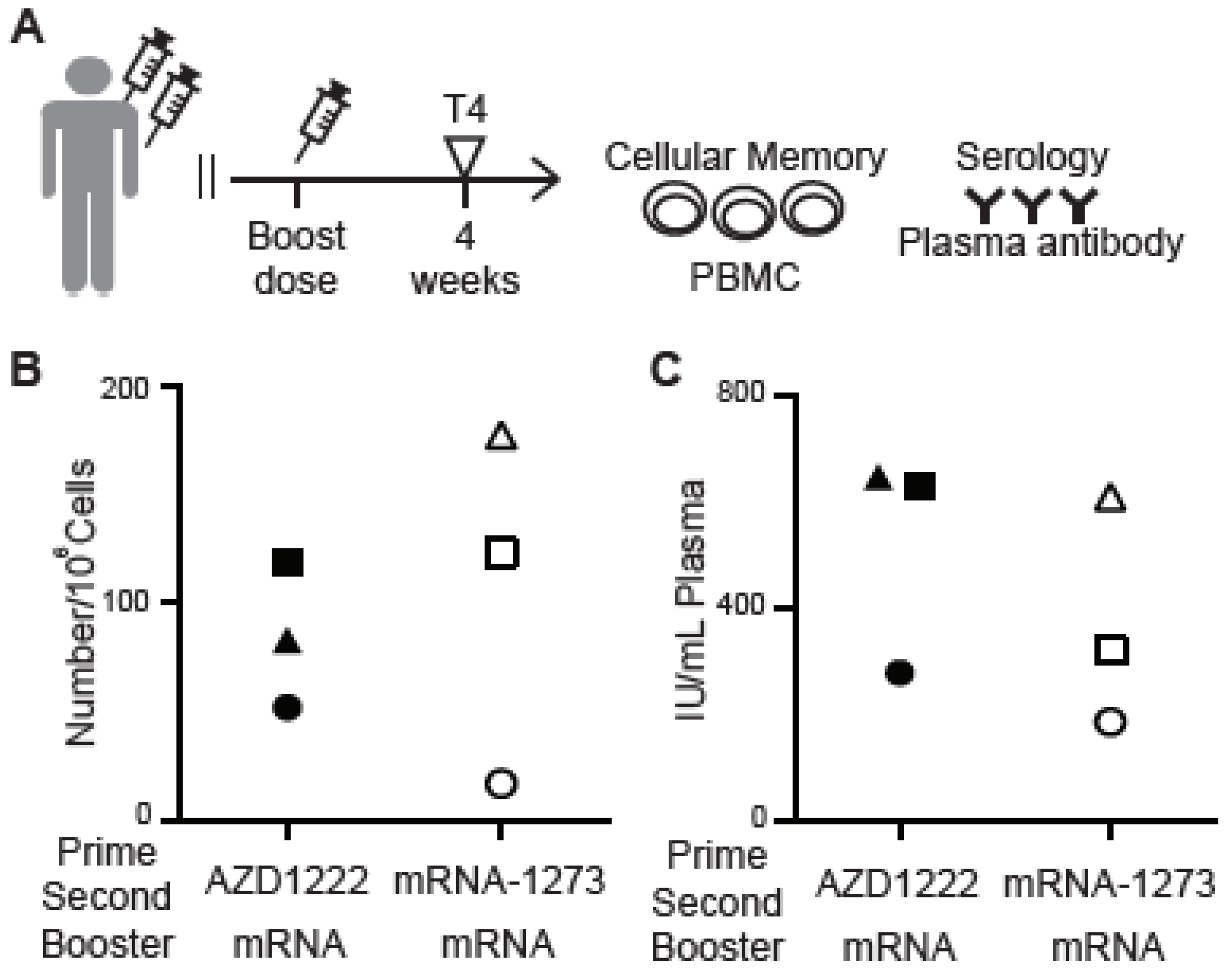
3. Results
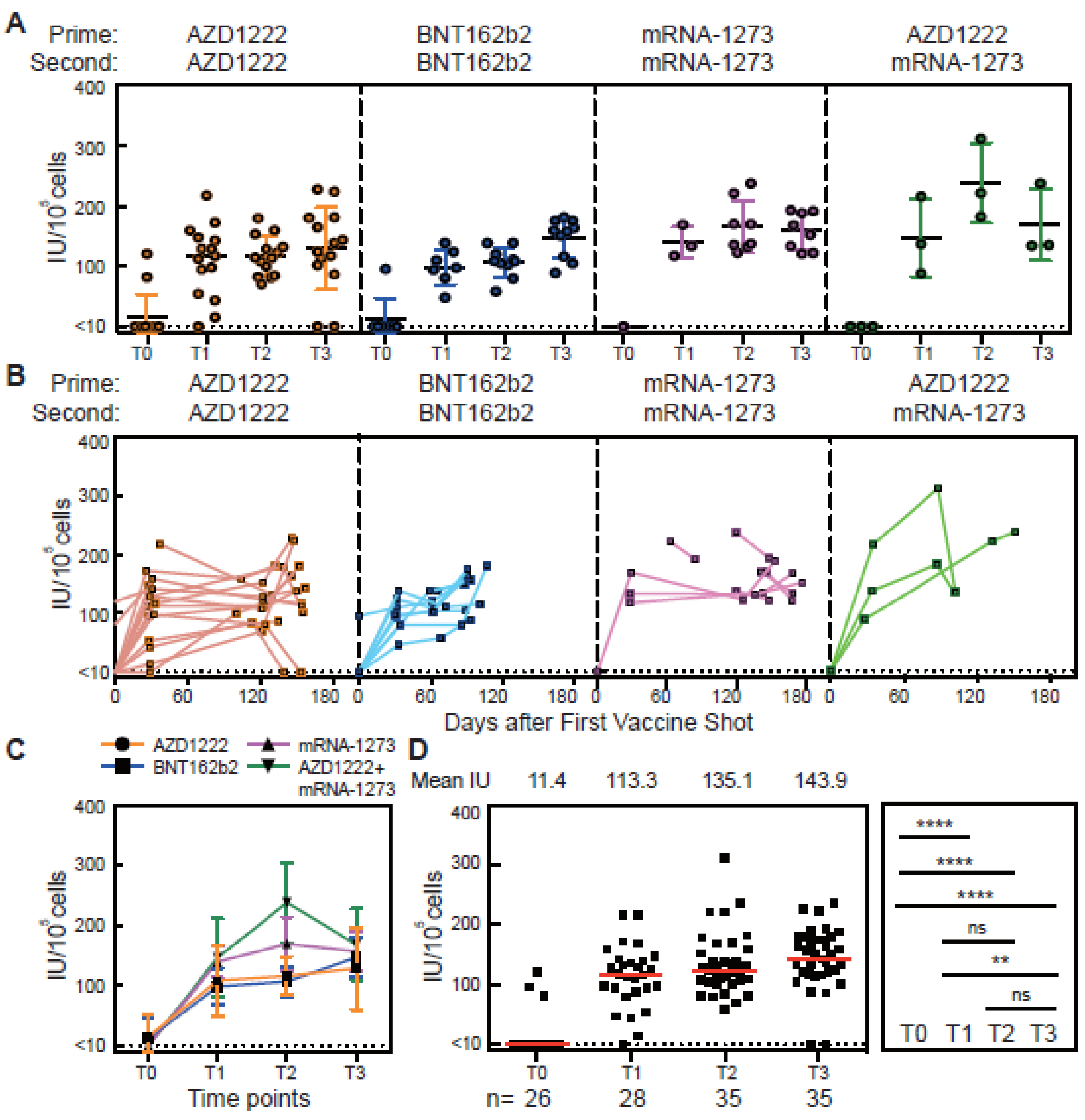
3.1. Antibody Responses of Vaccinated Subjects
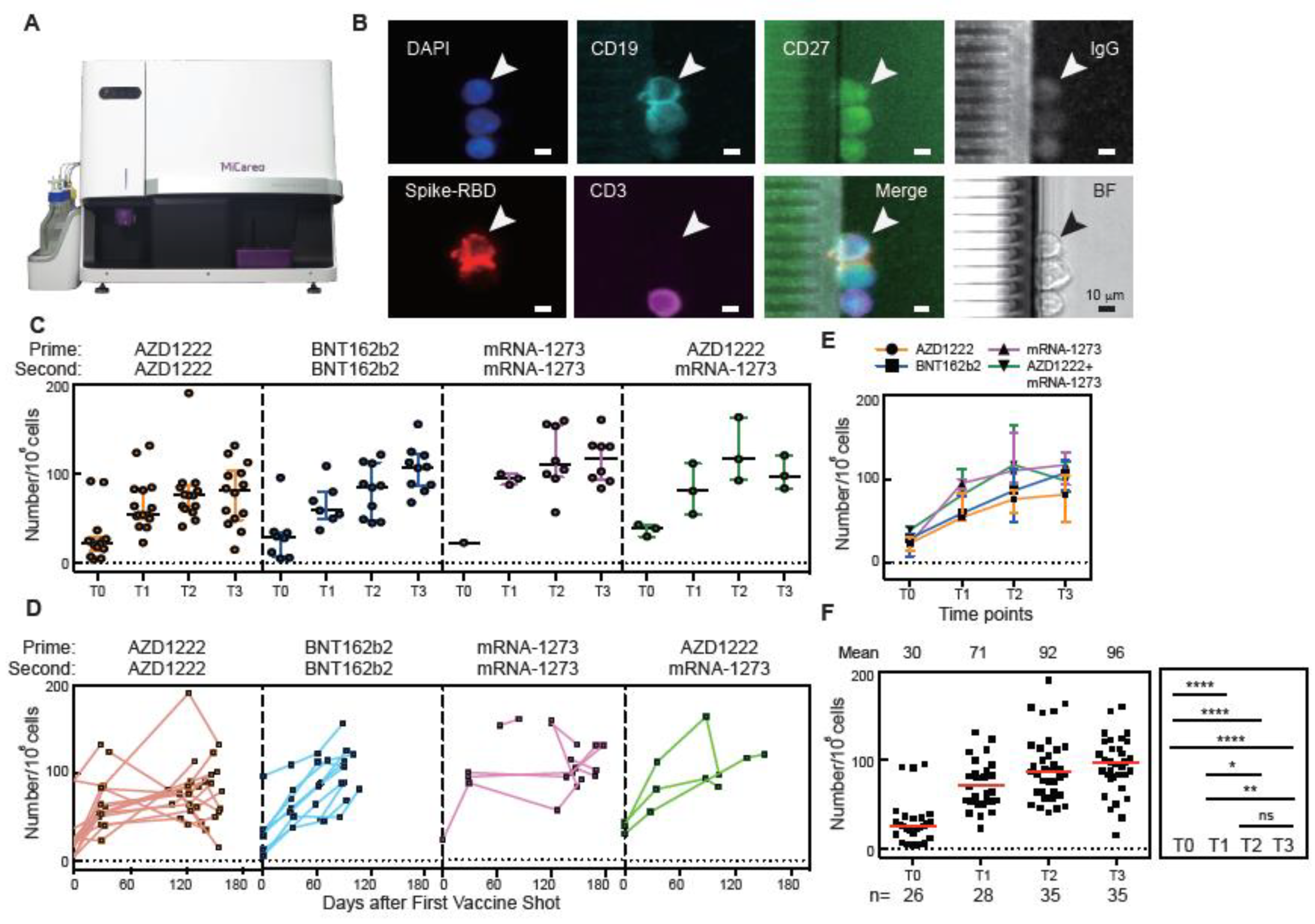
3.2. Spike-RBD-Specific Memory B Cells Detected by the MiSelect R II System
3.3. Cellular Memory Responses from PBMCs of Vaccinated Subjects
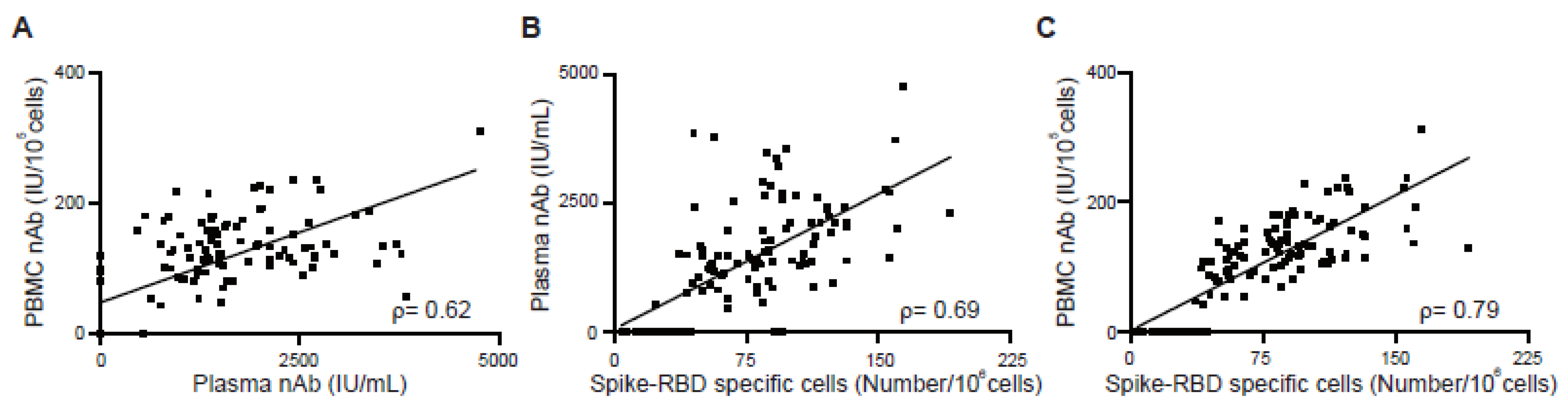
3.4. Correlation between Antibody Response and Cellular Memory from Vaccinated Subjects
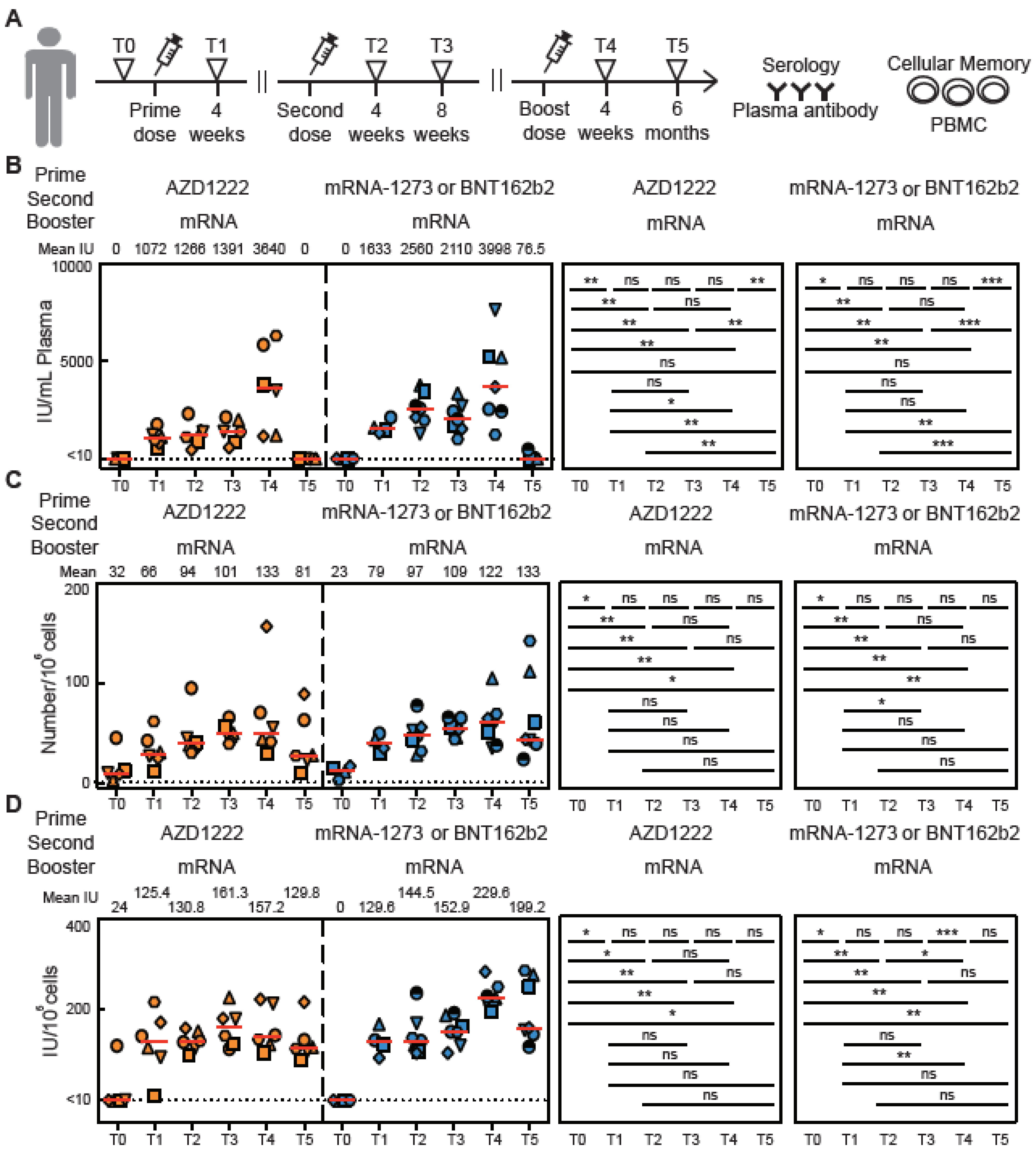
3.5. Cellular Memory to Major Variants of Concern (VOC)
4. Discussion
Limitations of the Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Callaway, E. The next generation of coronavirus vaccines: A graphical guide. Nature 2023, 614, 22–25. [Google Scholar] [CrossRef]
- Joseph, M.; Wu, Y.; Dannebaum, R.; Rubelt, F.; Zlatareva, I.; Lorenc, A.; Du, Z.G.; Davies, D.; Kyle-Cezar, F.; Das, A.; et al. Global patterns of antigen receptor repertoire disruption across adaptive immune compartments in COVID-19. Proc. Natl. Acad. Sci. USA 2022, 119, e2201541119. [Google Scholar] [CrossRef] [PubMed]
- Sette, A.; Crotty, S. Immunological memory to SARS-CoV-2 infection and COVID-19 vaccines. Immunol. Rev. 2022, 310, 27–46. [Google Scholar] [CrossRef] [PubMed]
- Vardhana, S.; Baldo, L.; Morice, W.G., 2nd; Wherry, E.J. Understanding T cell responses to COVID-19 is essential for informing public health strategies. Sci. Immunol. 2022, 7, eabo1303. [Google Scholar] [CrossRef] [PubMed]
- Saggau, C.; Martini, G.R.; Rosati, E.; Meise, S.; Messner, B.; Kamps, A.-K.; Bekel, N.; Gigla, J.; Rose, R.; Voß, M.; et al. The pre-exposure SARS-CoV-2-specific T cell repertoire determines the quality of the immune response to vaccination. Immunity 2022, 55, 1924–1939.e5. [Google Scholar] [CrossRef]
- Wang, Z.; Schmidt, F.; Weisblum, Y.; Muecksch, F.; Barnes, C.O.; Finkin, S.; Schaefer-Babajew, D.; Cipolla, M.; Gaebler, C.; Lieberman, J.A.; et al. mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants. Nature 2021, 592, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Tay, M.Z.; Rouers, A.; Fong, S.-W.; Goh, Y.S.; Chan, Y.-H.; Chang, Z.W.; Xu, W.; Tan, C.W.; Chia, W.N.; Torres-Ruesta, A.; et al. Decreased memory B cell frequencies in COVID-19 delta variant vaccine breakthrough infection. EMBO Mol. Med. 2022, 14, e15227. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.; Conklin, J. Standardization of two SARS-CoV-2 serology assays to the WHO 20/136 human standard reference material. J. Virol. Methods 2022, 300, 114430. [Google Scholar] [CrossRef]
- Yuan, M.; Wu, N.C.; Zhu, X.; Lee, C.-C.D.; So, R.T.Y.; Lv, H.; Mok, C.K.P.; Wilson, I.A. A highly conserved cryptic epitope in the receptor binding domains of SARS-CoV-2 and SARS-CoV. Science 2020, 368, 630–633. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.S.; Xu, S.; Yu, H.-M.; Fang, W.-F.; Qin, Y.; Wu, L.; Wang, J.; Zhao, M.; Schiro, P.G.; Fujimoto, B.; et al. Isolating Rare Cells and Circulating Tumor Cells with High Purity by Sequential eDAR. Anal. Chem. 2019, 91, 14605–14610. [Google Scholar] [CrossRef]
- Buckner, C.M.; Kardava, L.; El Merhebi, O.; Narpala, S.R.; Serebryannyy, L.; Lin, B.C.; Wang, W.; Zhang, X.; Lopes de Assis, F.; Kelly, S.E.M.; et al. Interval between prior SARS-CoV-2 infection and booster vaccination impacts magnitude and quality of antibody and B cell responses. Cell 2022, 185, 4333–4346.e14. [Google Scholar] [CrossRef]
- Jahnmatz, M.; Kesa, G.; Netterlid, E.; Buisman, A.-M.; Thorstensson, R.; Ahlborg, N. Optimization of a human IgG B-cell ELISpot assay for the analysis of vaccine-induced B-cell responses. J. Immunol. Methods 2013, 391, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, D.-Y.; Hung, K.-H.; Lin, I.-Y.; Su, S.-T.; Wu, S.-Y.; Chung, C.-H.; Wang, T.-C.; Li, W.-H.; Shih, A.C.-C.; Lin, K.-I. Uncovering MicroRNA Regulatory Hubs that Modulate Plasma Cell Differentiation. Sci. Rep. 2015, 5, 17957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Yin, S.; Tong, X.; Tao, Y.; Ni, J.; Pan, J.; Li, M.; Wan, Y.; Mao, M.; Xiong, Y.; et al. Dynamic SARS-CoV-2-specific B-cell and T-cell responses following immunization with an inactivated COVID-19 vaccine. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2022, 28, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Dou, X.; Wang, E.; Jiang, R.; Li, M.; Xiong, D.; Sun, B.; Zhang, X. Longitudinal profile of neutralizing and binding antibodies in vaccinated and convalescent COVID-19 cohorts by chemiluminescent immunoassays. Immun. Inflamm. Dis. 2022, 10, e612. [Google Scholar] [CrossRef]
- Zhang, J.; Xing, S.; Liang, D.; Hu, W.; Ke, C.; He, J.; Yuan, R.; Huang, Y.; Li, Y.; Liu, D.; et al. Differential Antibody Response to Inactivated COVID-19 Vaccines in Healthy Subjects. Front. Cell. Infect. Microbiol. 2021, 11, 791660. [Google Scholar] [CrossRef]
- Song, G.; He, W.-T.; Callaghan, S.; Anzanello, F.; Huang, D.; Ricketts, J.; Torres, J.L.; Beutler, N.; Peng, L.; Vargas, S.; et al. Cross-reactive serum and memory B-cell responses to spike protein in SARS-CoV-2 and endemic coronavirus infection. Nat. Commun. 2021, 12, 2938. [Google Scholar] [CrossRef]
- Bartolo, L.; Afroz, S.; Pan, Y.-G.; Xu, R.; Williams, L.; Lin, C.-F.; Tanes, C.; Bittinger, K.; Friedman, E.S.; Gimotty, P.A.; et al. SARS-CoV-2-specific T cells in unexposed adults display broad trafficking potential and cross-react with commensal antigens. Sci. Immunol. 2022, 7, eabn3127. [Google Scholar] [CrossRef]
- Mesin, L.; Ersching, J.; Victora, G.D. Germinal Center B Cell Dynamics. Immunity 2016, 45, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Nutt, S.L.; Hodgkin, P.D.; Tarlinton, D.M.; Corcoran, L.M. The generation of antibody-secreting plasma cells. Nat. Rev. Immunol. 2015, 15, 160–171. [Google Scholar] [CrossRef]
- Mantus, G.; Nyhoff, L.E.; Edara, V.-V.; Zarnitsyna, V.I.; Ciric, C.R.; Flowers, M.W.; Norwood, C.; Ellis, M.; Hussaini, L.; Manning, K.E.; et al. Pre-existing SARS-CoV-2 immunity influences potency, breadth, and durability of the humoral response to SARS-CoV-2 vaccination. Cell Rep. Med. 2022, 3, 100603. [Google Scholar] [CrossRef]
- Pape, K.A.; Dileepan, T.; Kabage, A.J.; Kozysa, D.; Batres, R.; Evert, C.; Matson, M.; Lopez, S.; Krueger, P.D.; Graiziger, C.; et al. High-affinity memory B cells induced by SARS-CoV-2 infection produce more plasmablasts and atypical memory B cells than those primed by mRNA vaccines. Cell Rep. 2021, 37, 109823. [Google Scholar] [CrossRef]
- Goel, R.R.; Apostolidis, S.A.; Painter, M.M.; Mathew, D.; Pattekar, A.; Kuthuru, O.; Gouma, S.; Hicks, P.; Meng, W.; Rosenfeld, A.M.; et al. Distinct antibody and memory B cell responses in SARS-CoV-2 naïve and recovered individuals following mRNA vaccination. Sci. Immunol. 2021, 6, abi6950. [Google Scholar] [CrossRef] [PubMed]
- Atmar, R.L.; Lyke, K.E.; Deming, M.E.; Jackson, L.A.; Branche, A.R.; El Sahly, H.M.; Rostad, C.A.; Martin, J.M.; Johnston, C.; Rupp, R.E.; et al. Homologous and Heterologous Covid-19 Booster Vaccinations. N. Engl. J. Med. 2022, 386, 1046–1057. [Google Scholar] [CrossRef]
- Jalkanen, P.; Kolehmainen, P.; Häkkinen, H.K.; Huttunen, M.; Tähtinen, P.A.; Lundberg, R.; Maljanen, S.; Reinholm, A.; Tauriainen, S.; Pakkanen, S.H.; et al. COVID-19 mRNA vaccine induced antibody responses against three SARS-CoV-2 variants. Nat. Commun. 2021, 12, 3991. [Google Scholar] [CrossRef] [PubMed]
- Muik, A.; Lui, B.G.; Wallisch, A.-K.; Bacher, M.; Mühl, J.; Reinholz, J.; Ozhelvaci, O.; Beckmann, N.; Güimil Garcia, R.D.L.C.; Poran, A.; et al. Neutralization of SARS-CoV-2 Omicron by BNT162b2 mRNA vaccine-elicited human sera. Science 2022, 375, 678–680. [Google Scholar] [CrossRef]
- Goel, R.R.; Painter, M.M.; Apostolidis, S.A.; Mathew, D.; Meng, W.; Rosenfeld, A.M.; Lundgreen, K.A.; Reynaldi, A.; Khoury, D.S.; Pattekar, A.; et al. mRNA vaccines induce durable immune memory to SARS-CoV-2 and variants of concern. Science 2021, 374, abm0829. [Google Scholar] [CrossRef]
- van den Hoogen, L.L.; Verheul, M.K.; Vos, E.R.A.; van Hagen, C.C.E.; van Boven, M.; Wong, D.; Wijmenga-Monsuur, A.J.; Smits, G.; Kuijer, M.; van Rooijen, D.; et al. SARS-CoV-2 Spike S1-specific IgG kinetic profiles following mRNA or vector-based vaccination in the general Dutch population show distinct kinetics. Sci. Rep. 2022, 12, 5935. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Zhao, M.; Slotkin, R.; Sheth, A.H.; Pischel, L.; Kyriakides, T.C.; Emu, B.; McNamara, C.; Shi, Q.; Delgobbo, J.; Xu, J.; et al. Serum Neutralizing Antibody Titers 12 months after COVID-19 mRNA Vaccination: Correlation to Clinical Variables in an Adult, US-Population. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2022, 76, e391–e399. [Google Scholar] [CrossRef]
- Piano Mortari, E.; Russo, C.; Vinci, M.R.; Terreri, S.; Fernandez Salinas, A.; Piccioni, L.; Alteri, C.; Colagrossi, L.; Coltella, L.; Ranno, S.; et al. Highly Specific Memory B Cells Generation after the 2nd Dose of BNT162b2 Vaccine Compensate for the Decline of Serum Antibodies and Absence of Mucosal IgA. Cells 2021, 10, 2541. [Google Scholar] [CrossRef]
- DeVilbiss, A.W.; Zhao, Z.; Martin-Sandoval, M.S.; Ubellacker, J.M.; Tasdogan, A.; Agathocleous, M.; Mathews, T.P.; Morrison, S.J. Metabolomic profiling of rare cell populations isolated by flow cytometry from tissues. eLife 2021, 10, e61980. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Yanikkaya Demirel, G.; Keller, T.; Preijers, F.; Psarra, K.; Schiemann, M.; Özçürümez, M.; Sack, U. Flow Cytometric Analyses of Lymphocyte Markers in Immune Oncology: A Comprehensive Guidance for Validation Practice According to Laws and Standards. Front. Immunol. 2020, 11, 2169. [Google Scholar] [CrossRef] [PubMed]
- van Dongen, J.J.M.; van der Burg, M.; Kalina, T.; Perez-Andres, M.; Mejstrikova, E.; Vlkova, M.; Lopez-Granados, E.; Wentink, M.; Kienzler, A.-K.; Philippé, J.; et al. EuroFlow-Based Flowcytometric Diagnostic Screening and Classification of Primary Immunodeficiencies of the Lymphoid System. Front. Immunol. 2019, 10, 1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelanew, T.; Mulu, A.; Abebe, M.; Bates, T.A.; Wassie, L.; Teferi, M.; Fentahun, D.; Alemu, A.; Tamiru, F.; Assefa, G.; et al. A Single Dose of ChAdOx1 nCoV-19 Vaccine Elicits High Antibody Responses in Individuals with Prior SARS-CoV-2 Infection Comparable to That of Two-Dose-Vaccinated, SARS-CoV-2-Infection-Naïve Individuals: A Longitudinal Study in Ethiopian Health Workers. Vaccines 2022, 10, 859. [Google Scholar] [CrossRef] [PubMed]
- Guiomar, R.; Santos, A.J.; Melo, A.M.; Costa, I.; Matos, R.; Rodrigues, A.P.; Kislaya, I.; Silva, A.S.; Roque, C.; Nunes, C.; et al. Monitoring of SARS-CoV-2 Specific Antibodies after Vaccination. Vaccines 2022, 10, 154. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, D.-Y.; Wang, C.-H.; Schiro, P.G.; Chen, N.; Tseng, J.-Y. Tracking B Cell Memory to SARS-CoV-2 Using Rare Cell Analysis System. Vaccines 2023, 11, 735. https://doi.org/10.3390/vaccines11040735
Tsai D-Y, Wang C-H, Schiro PG, Chen N, Tseng J-Y. Tracking B Cell Memory to SARS-CoV-2 Using Rare Cell Analysis System. Vaccines. 2023; 11(4):735. https://doi.org/10.3390/vaccines11040735
Chicago/Turabian StyleTsai, Dong-Yan, Chun-Hung Wang, Perry G. Schiro, Nathan Chen, and Ju-Yu Tseng. 2023. "Tracking B Cell Memory to SARS-CoV-2 Using Rare Cell Analysis System" Vaccines 11, no. 4: 735. https://doi.org/10.3390/vaccines11040735