
Vaccine Interaction and Protection against Virulent Avian Metapneumovirus (aMPV) Challenge after Combined Administration of Newcastle Disease and aMPV Live Vaccines to Day-Old Turkeys

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Birds
2.2. Vaccines
2.3. Challenge Virus
2.4. Experimental Trial
- TRT group (16 birds)—vaccinated with aMPV strain VCO3;
- B1 group (8 birds)—vaccinated with NDV strain B1;
- VG/GA group (8 birds)—vaccinated with NDV strain VG/GA;
- TRT-B1 group (16 birds)—vaccinated with aMPV strain VCO3 and NDV strain B1, coadministered;
- TRT-VG/GA group (16 birds)—vaccinated with aMPV strain VCO3 and NDV strain VG/GA, coadministered;
- CONTROL group (16 birds)—mock-vaccinated with sterile water.
2.5. aMPV and NDV Detection in Oro-Pharyngeal Swabs
2.6. Serology for aMPV and NDV
2.7. Clinical Signs
- 0: no signs;
- 1: clear nasal exudate;
- 2: turbid nasal exudate;
- 3: swollen infra-orbital sinuses and/or frothy eyes.
2.8. Statistical Analysis
3. Results
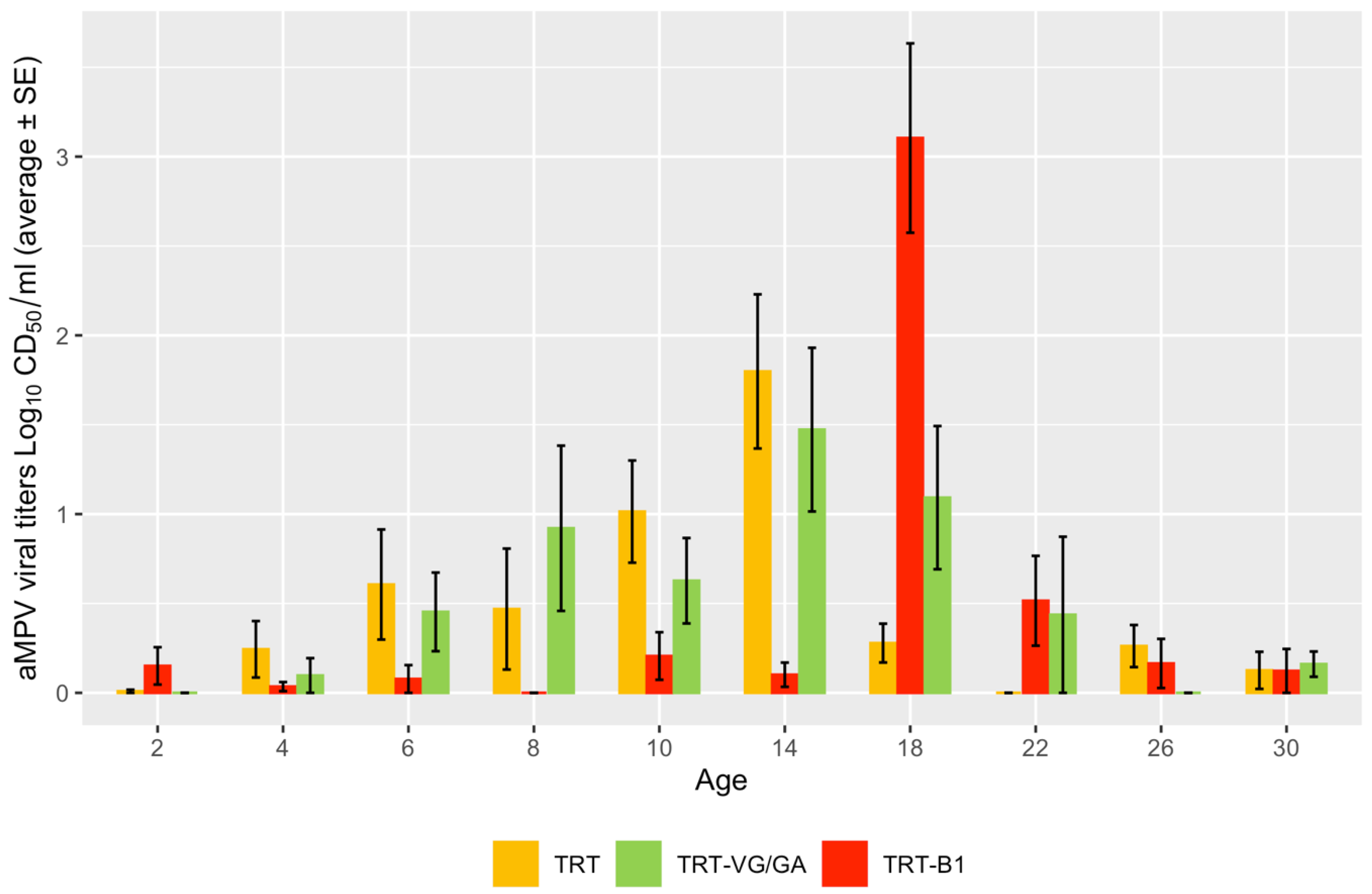
3.1. Shedding of Vaccine Viruses
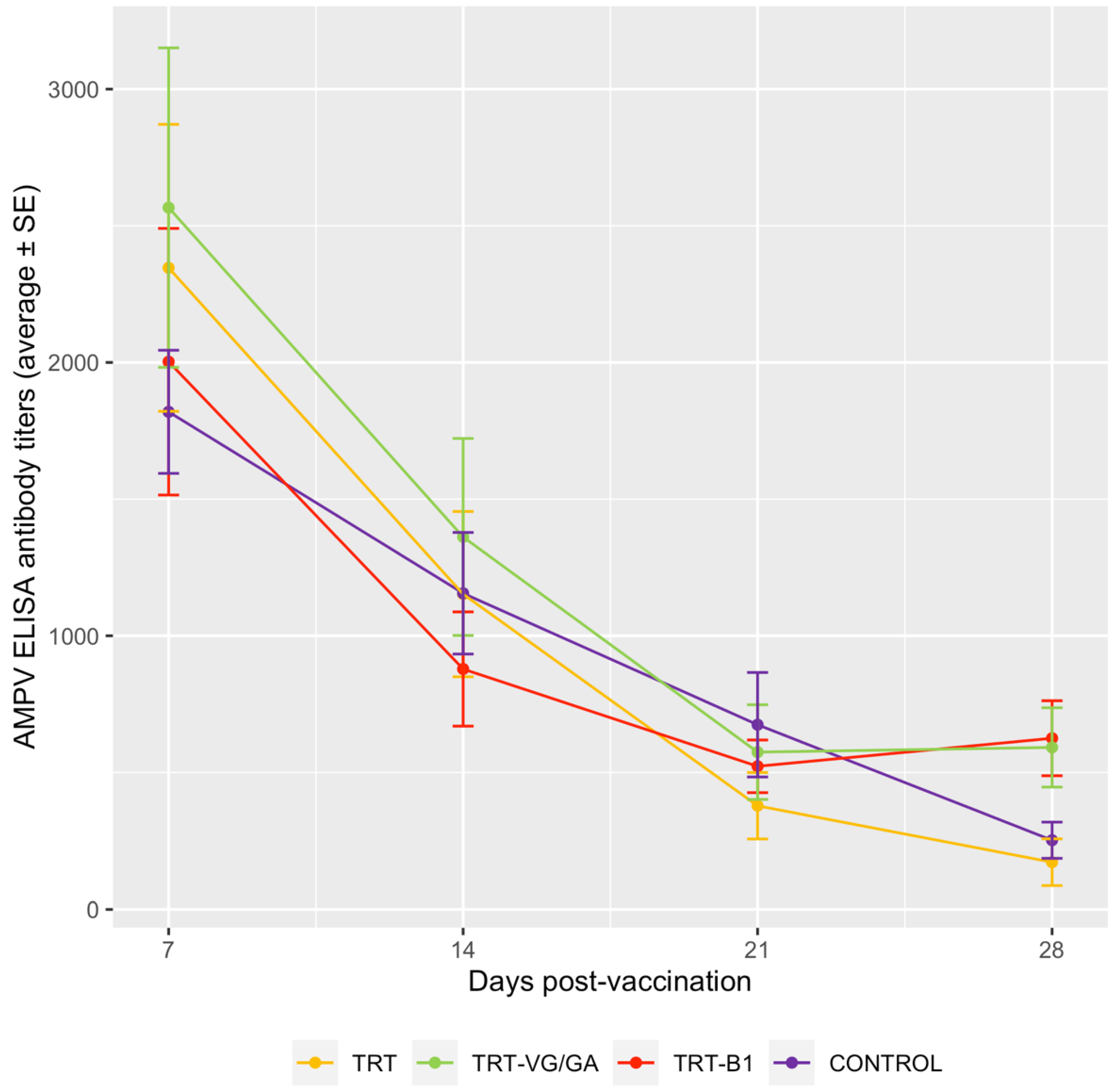
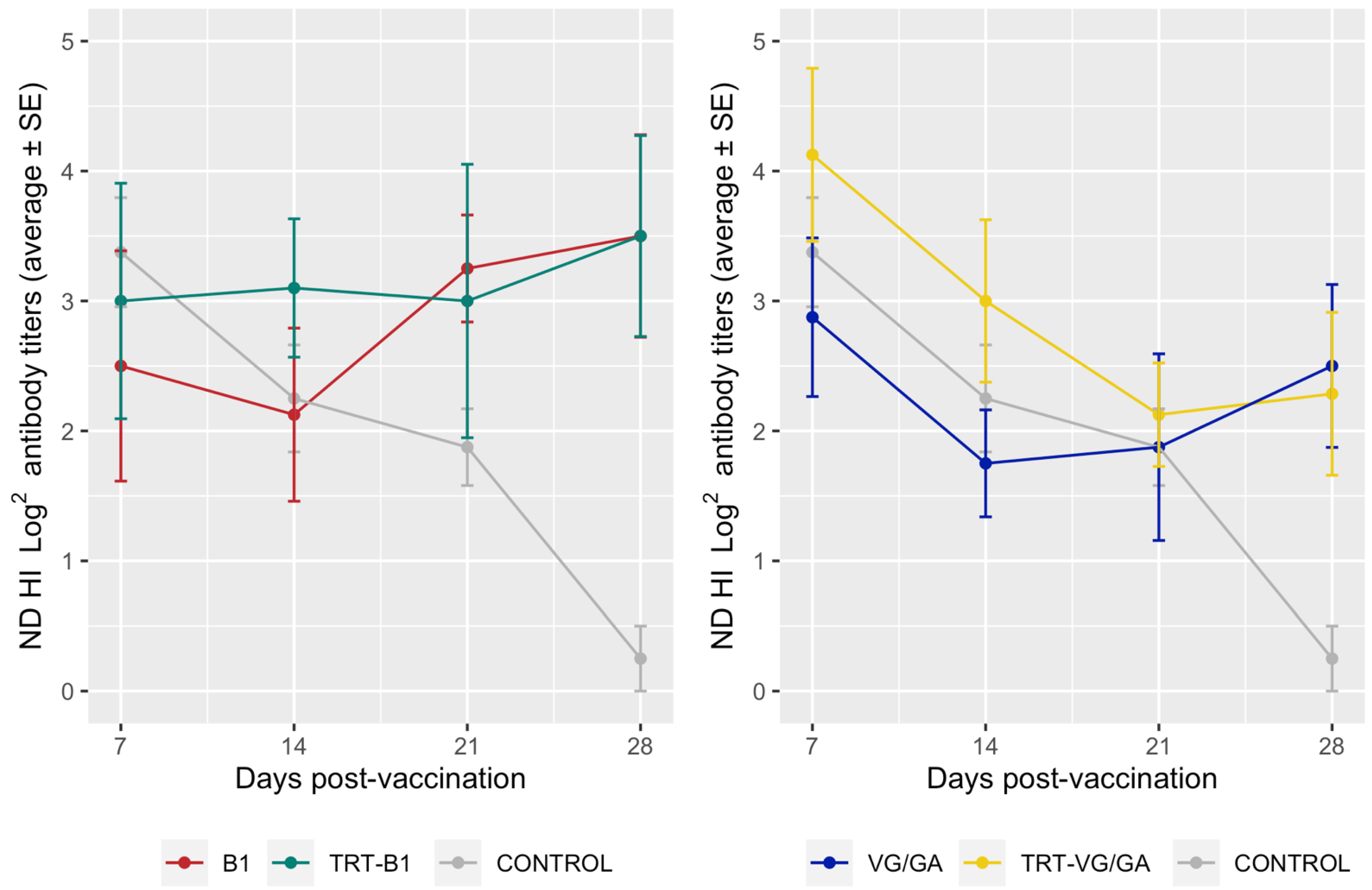
3.2. Serology
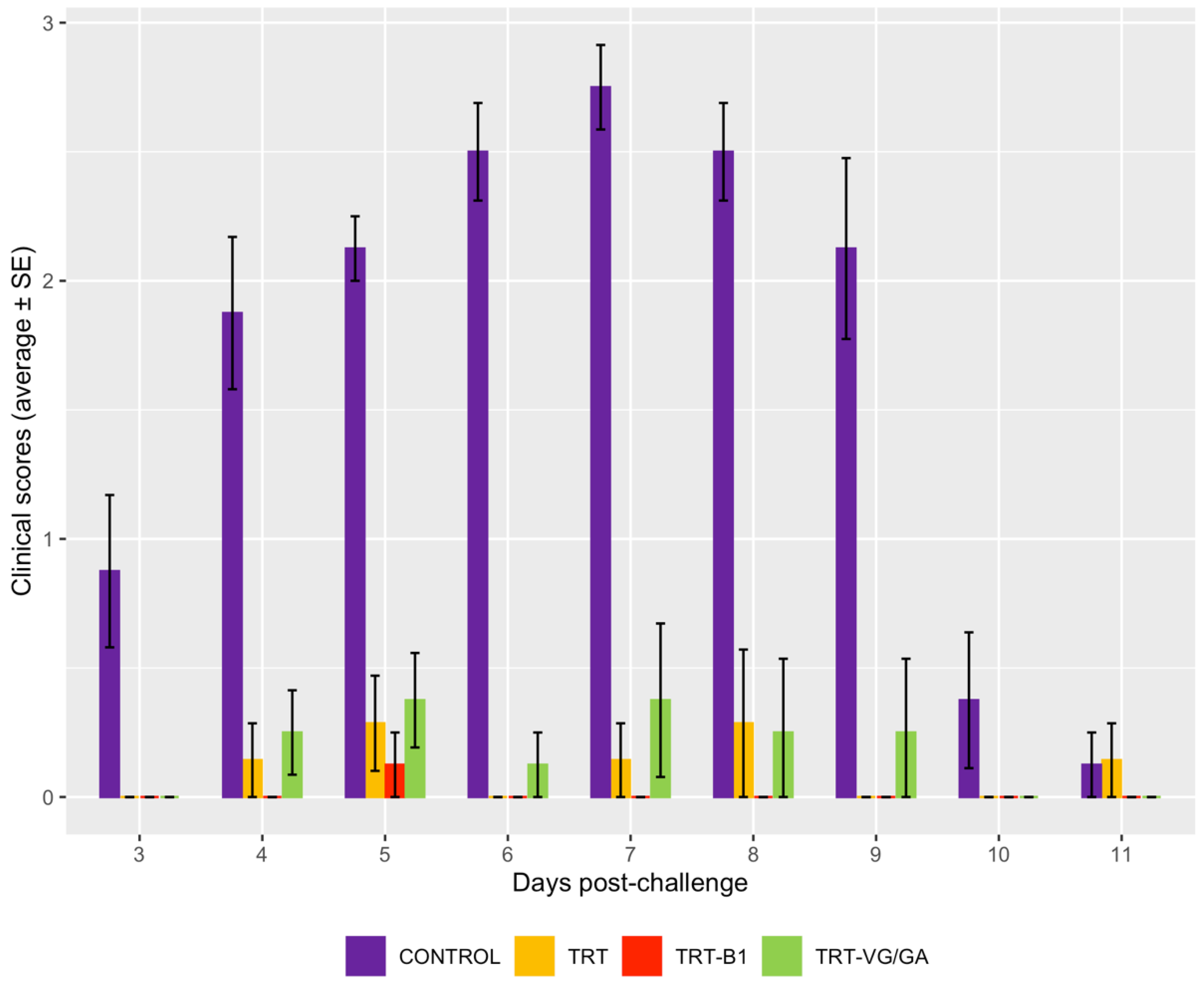
3.3. Clinical Signs after aMPV Challenge
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Available online: https://www.fao.org/faostat/en/ (accessed on 10 November 2022).
- Piacenti, A.M.; King, D.J.; Seal, B.S.; Zhang, J.; Brown, C.C. Pathogenesis of Newcastle disease in commercial and specific pathogen-free turkeys experimentally infected with isolates of different virulence. Vet. Pathol. 2006, 43, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Suarez, D.L.; Miller, P.J.; Koch, G.; Mundt, E.; Rautenschlein, S. Newcastle disease, other avian paramyxoviruses, and avian metapneumovirus infections. In Diseases of Poultry, 14th ed.; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Wit, S., Grimes, T., Johnson, D., Kromm, M., et al., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 109–166. [Google Scholar]
- Kaboudi, K.; Lachheb, J. Avian metapneumovirus infection in turkeys: A review on turkey rhinotracheitis. J. Appl. Poult. Res. 2021, 30, 100211. [Google Scholar] [CrossRef]
- Brown, P.A.; Allée, C.; Courtillon, C.; Szerman, N.; Lemaitre, E.; Toquin, D.; Mangart, J.-M.; Amelot, M.; Eterradossi, N. Host specificity of avian metapneumoviruses. Avian Pathol. 2019, 48, 311–318. [Google Scholar] [CrossRef]
- Franzo, G.; Legnardi, M.; Mescolini, G.; Tucciarone, C.M.; Lupini, C.; Quaglia, G.; Catelli, E.; Cecchinato, M. Avian Metapneumovirus subtype B around Europe: A phylodynamic reconstruction. Vet. Res. 2020, 51, 88. [Google Scholar] [CrossRef] [PubMed]
- Canuti, M.; Kroyer, A.N.; Ojkic, D.; Whitney, H.G.; Robertson, G.J.; Lang, A.S. Discovery and characterization of novel RNA viruses in aquatic North American wild birds. Viruses 2019, 11, 768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Retallack, H.; Clubb, S.; DeRisi, J.L. Genome sequence of a divergent avian metapneumovirus from a monk parakeet (Myiopsitta monachus). Microbiol. Resour. Announc. 2019, 8, e00284-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganapathy, K.; Cargill, P.; Montiel, E.; Jones, R.C. Interaction between live avian pneumovirus and Newcastle disease virus vaccines in specific pathogen free chickens. Avian Pathol. 2005, 34, 297–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganapathy, K.; Todd, V.; Cargill, P.; Montiel, E.; Jones, R.C. Interaction between a live avian pneumovirus vaccine and two different Newcastle disease virus vaccines in broiler chickens with maternal antibodies to Newcastle disease virus. Avian Pathol. 2006, 35, 429–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganapathy, K.; Cox, W.J.; Gough, R.E.; Cargill, P.; Montiel, E.; Jones, R.C. Protection in specific pathogen free chickens with live avian metapneumovirus and Newcastle disease virus vaccines applied singly or in combination. Avian Pathol. 2007, 36, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Tarpey, I.; Huggins, M.B.; Orbell, S.J. The efficacy of an avian metapneumovirus vaccine applied simultaneously with infectious bronchitis and Newcastle disease virus vaccines to specific-pathogen-free chickens. Avian Dis. 2007, 51, 594–596. [Google Scholar] [CrossRef]
- Awad, F.; Forrester, A.; Baylis, M.; Lemiere, S.; Jones, R.C.; Ganapathy, K. Immune responses and interactions following simultaneous application of live Newcastle disease, infectious bronchitis and avian metapneumovirus vaccines in specific-pathogen-free chicks. Res. Vet. Sci. 2015, 98, 127–133. [Google Scholar] [CrossRef] [PubMed]
- World Organization for Animal Health. Turkey rinotracheitis (avian metapneumovirus infection). In OIE Terrestrial Manual; WHO: Rome, Italy, 2022; Available online: https://www.woah.org/fileadmin/Home/fr/Health_standards/tahm/3.03.15_TURKEY_RHINO.pdf (accessed on 10 November 2022).
- Coswig, L.T.; dos Santos, M.B.; Hafez, H.M.; Ferreira, H.L.; Arns, C.W. Propagation of avian metapneumovirus subtypes A and B using chicken embryo related and other cell systems. J. Virol. Methods 2010, 167, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Li, J.; Cook, J.K.A.; Brown, T.D.K.; Shaw, K.; Cavanagh, D. Detection of turkey rhinotracheitis virus in turkeys using the polymerase chain reaction. Avian Pathol. 1993, 22, 771–783. [Google Scholar]
- Cecchinato, M.; Lupini, C.; Munoz Pogoreltseva, O.S.; Listorti, V.; Mondin, A.; Drigo, M.; Catelli, E. Development of a real-time RT-PCR assay for the simultaneous identification, quantitation and differentiation of avian metapneumovirus subtypes A and B. Avian Pathol. 2013, 42, 283–289. [Google Scholar] [CrossRef]
- Wise, M.G.; Suarez, D.L.; Seal, B.S.; Pedersen, J.C.; Senne, D.A.; King, D.J.; Kapczynski, D.R.; Spackman, E. Development of a real-time reverse-transcription PCR for detection of Newcastle disease virus RNA in clinical samples. J. Clin. Microbiol. 2004, 42, 329–338. [Google Scholar] [CrossRef] [Green Version]
- World Organization for Animal Health. Newcastle disease (infection with Newcastle disease virus). In OIE Terrestrial Manual; WHO: Rome, Italy, 2021; Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/3.03.14_NEWCASTLE_DIS.pdf (accessed on 10 March 2023).
- Naylor, C.J.; Jones, R.C. Demonstration of a virulent subpopulation in a prototype live attenuated turkey rhinotracheitis vaccine. Vaccine 1994, 12, 1225–1230. [Google Scholar] [CrossRef]
- Ball, C.; Manswr, B.; Herrmann, A.; Lemiere, S.; Ganapathy, K. Avian metapneumovirus subtype B vaccination in commercial broiler chicks: Heterologous protection and selected host transcription responses to subtype A or B challenge. Avian Pathol. 2022, 51, 181–196. [Google Scholar] [CrossRef]
- Cook, J.K.A. Protection provided by a commercially available vaccine against different strains of turkey rhinotracheitis virus. Vet. Rec. 1995, 136, 392–393. [Google Scholar] [CrossRef]
- Cook, J.K.A.; Huggins, M.B.; Orbell, S.J.; Mawditt, K.; Cavanagh, D. Infectious bronchitis virus vaccine interferes with the replication of avian pneumovirus vaccine in domestic fowl. Avian Pathol. 2001, 30, 233–242. [Google Scholar] [CrossRef]
- Śmiałek, M.; Kowalczyk, J.; Gesek, M.; Kaczorek-Łukowska, E.; Dziewulska, D.; Tykałowski, B.; Koncicki, A. The influence of maternally derived antibodies on protection against aMPV/A infection in TRT vaccinated turkeys. Poult. Sci. 2021, 100, 101086. [Google Scholar] [CrossRef]
- Jones, R.C.; Naylor, C.J.; Al-Afaleq, A.; Worthington, K.J.; Jones, R. Effect of cyclophosphamide immunosuppression on the immunity of turkeys to viral rhinotracheitis. Res. Vet. Sci. 1992, 53, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Perozo, F.; Villegas, P.; Dolz, R.; Afonso, C.L.; Purvis, L.B. The VG/GA strain of Newcastle disease virus: Mucosal immunity, protection against lethal challenge and molecular analysis. Avian Pathol. 2008, 37, 237–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, D.L.; Maraqa, A.D. Protective immunity against Newcastle disease: The role of antibodies specific to Newcastle disease virus polypeptides. Avian Dis. 2000, 44, 138–144. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Oligonucleotide | Sequence | Reference |
|---|---|---|---|
| aMPV | SH forward primer | 5′-TAGTTTTGATCTTCCTTGTTGC-3′ | Cecchinato et al. (2013) [18] |
| SH reverse primer | 5′-GTAGTTGTGCTCAGCTCTGATA-3′ | ||
| MB SH-B probe | 5′ FAM-CGCGATCATTGTGACAGCCAGCTTCACGATCGCG-3′ Iowa Black® FQ | ||
| NDV | M+4100 forward primer | 5′-AGTGATGTGCTCGGACCTTC-3′ | Wise et al. (2004) [19] |
| M−4220 reverse primer | 5′-CCTGAGGAGAGGCATTTGCTA-3′ | ||
| M+4169 probe | 5′ FAM-TTCTCTAGCAGTGGGACAGCCTGC-3′ TAMRA |
| Days Post-Vaccination | NDV-Positive Birds | aMPV-Positive Birds | |||||||
|---|---|---|---|---|---|---|---|---|---|
| B1 | VG/GA | TRT-B1 | TRT- VG/GA | CONTROL | TRT | TRT-B1 | TRT- VG/GA | CONTROL | |
| 2 | 8 | 7 | 8 | 8 | 0 | 0 | 1 | 0 | 0 |
| 4 | 8 | 8 | 7 | 8 | 0 | 1 | 0 | 0 | 0 |
| 6 | 8 | 7 | 8 | 7 | 0 | 4 | 1 | 3 | 0 |
| 8 | 7 | 8 | 7 | 4 | 0 | 2 | 0 | 4 | 0 |
| 10 | 7 | 2 | 4 | 4 | 0 | 5 | 2 | 5 | 0 |
| 14 | 0 | 0 | 0 | 0 | 0 | 7 | 7 | 6 | 0 |
| 18 | 0 | 0 | 0 | 1 | 0 | 4 | 8 | 4 | 0 |
| 22 | 0 | 1 | 0 | 0 | 0 | 0 | 4 | 1 | 0 |
| 26 | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 0 | 0 |
| 30 | 0 | 0 | 0 | 0 | 0 | 2 | 2 | 0 | 0 |
| TOTAL | 38 | 33 | 34 | 32 | 0 | 27 | 26 | 23 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lupini, C.; Legnardi, M.; Graziosi, G.; Cecchinato, M.; Listorti, V.; Terregino, C.; Catelli, E. Vaccine Interaction and Protection against Virulent Avian Metapneumovirus (aMPV) Challenge after Combined Administration of Newcastle Disease and aMPV Live Vaccines to Day-Old Turkeys. Vaccines 2023, 11, 708. https://doi.org/10.3390/vaccines11030708
Lupini C, Legnardi M, Graziosi G, Cecchinato M, Listorti V, Terregino C, Catelli E. Vaccine Interaction and Protection against Virulent Avian Metapneumovirus (aMPV) Challenge after Combined Administration of Newcastle Disease and aMPV Live Vaccines to Day-Old Turkeys. Vaccines. 2023; 11(3):708. https://doi.org/10.3390/vaccines11030708
Chicago/Turabian StyleLupini, Caterina, Matteo Legnardi, Giulia Graziosi, Mattia Cecchinato, Valeria Listorti, Calogero Terregino, and Elena Catelli. 2023. "Vaccine Interaction and Protection against Virulent Avian Metapneumovirus (aMPV) Challenge after Combined Administration of Newcastle Disease and aMPV Live Vaccines to Day-Old Turkeys" Vaccines 11, no. 3: 708. https://doi.org/10.3390/vaccines11030708