
Post-Vaccination and Post-Infection Immunity to the Hepatitis B Virus and Circulation of Immune-Escape Variants in the Russian Federation 20 Years after the Start of Mass Vaccination
 , , , ,
, , , ,  , , , , add
Show full author list
, , , , add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
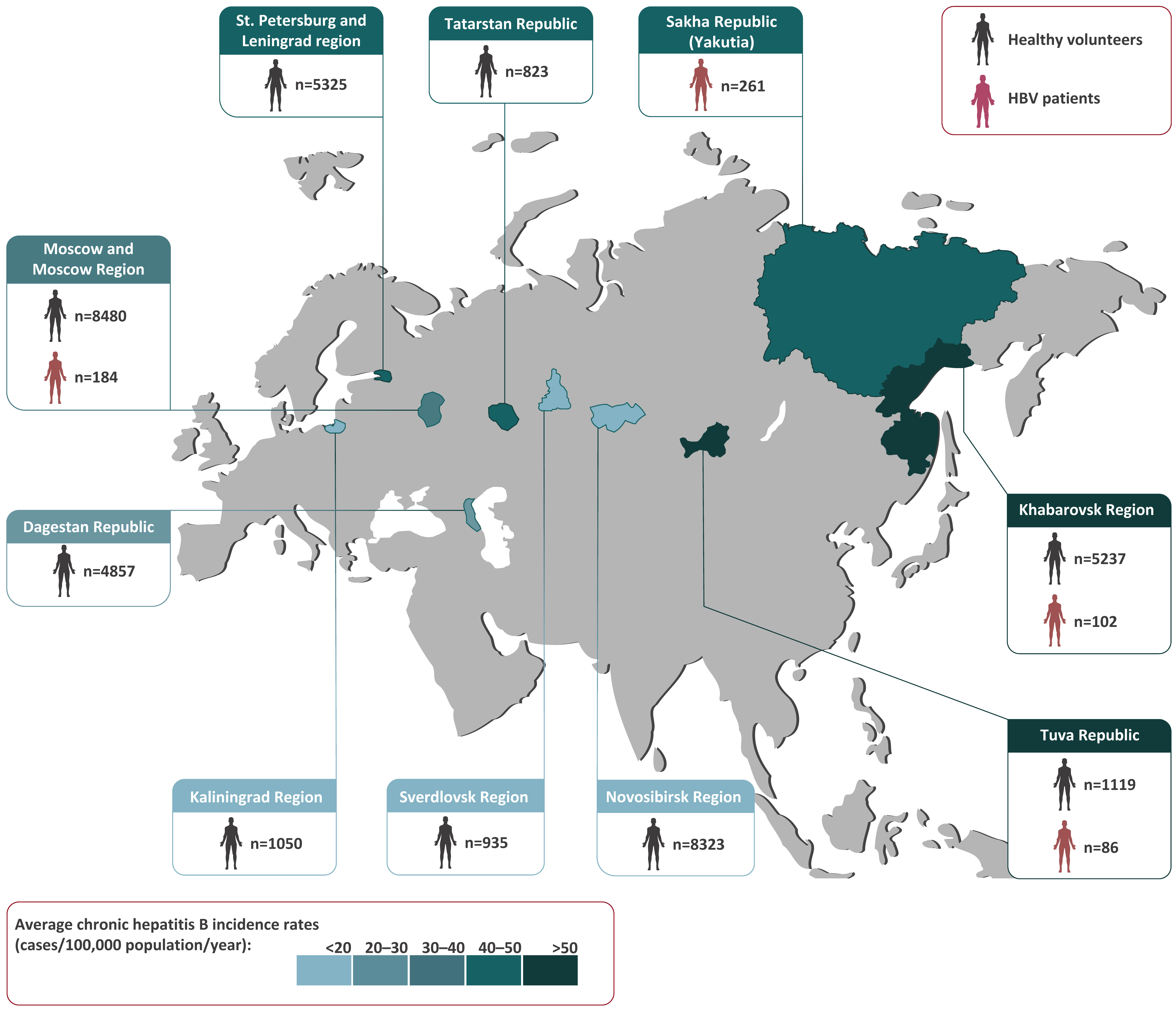
2.1. Serum Samples
2.2. HBV Testing
2.3. HBV Sequence Analysis
2.4. Statistical Analysis
3. Results
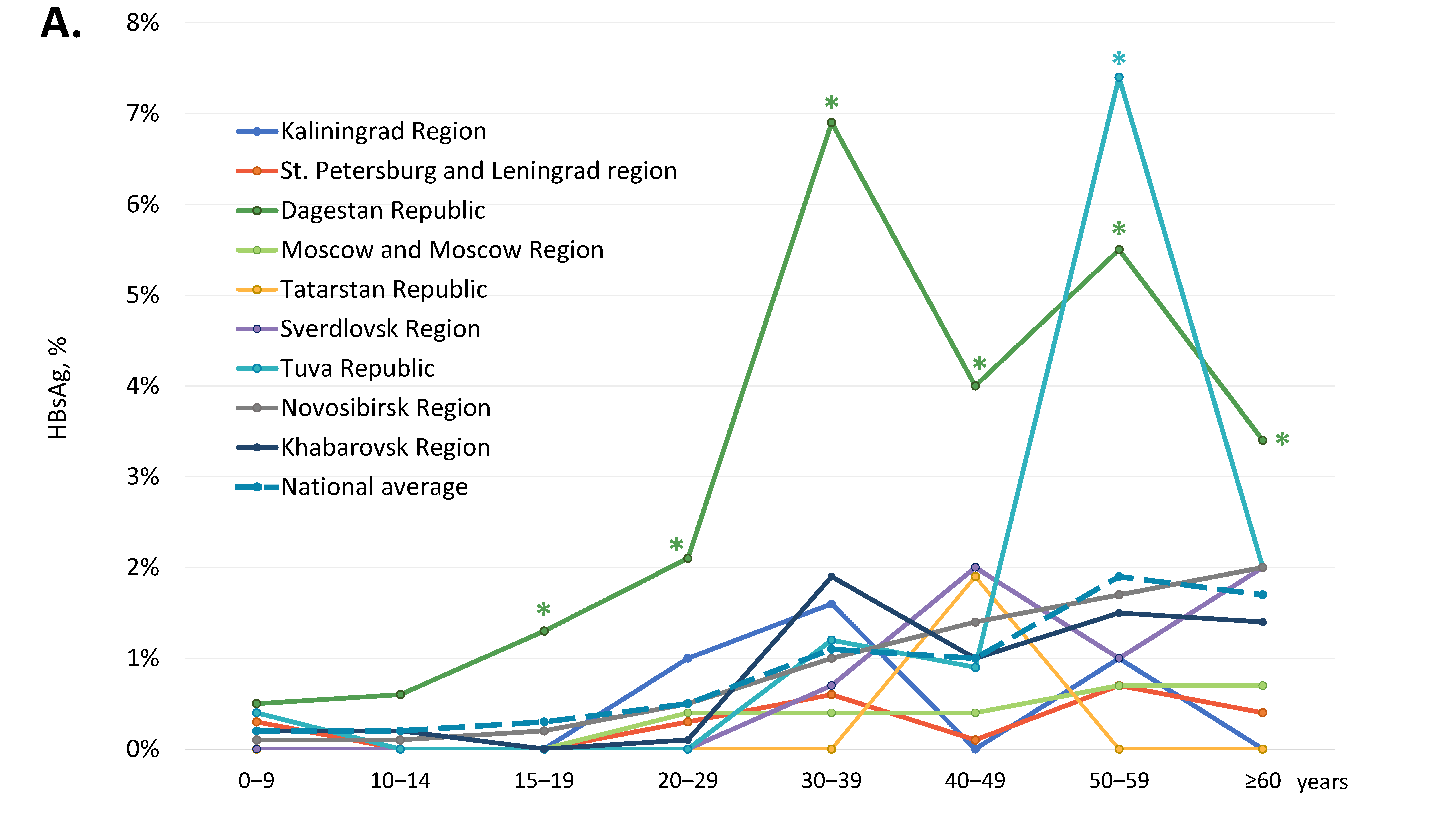
3.1. Frequency of HBV Infection in General Population
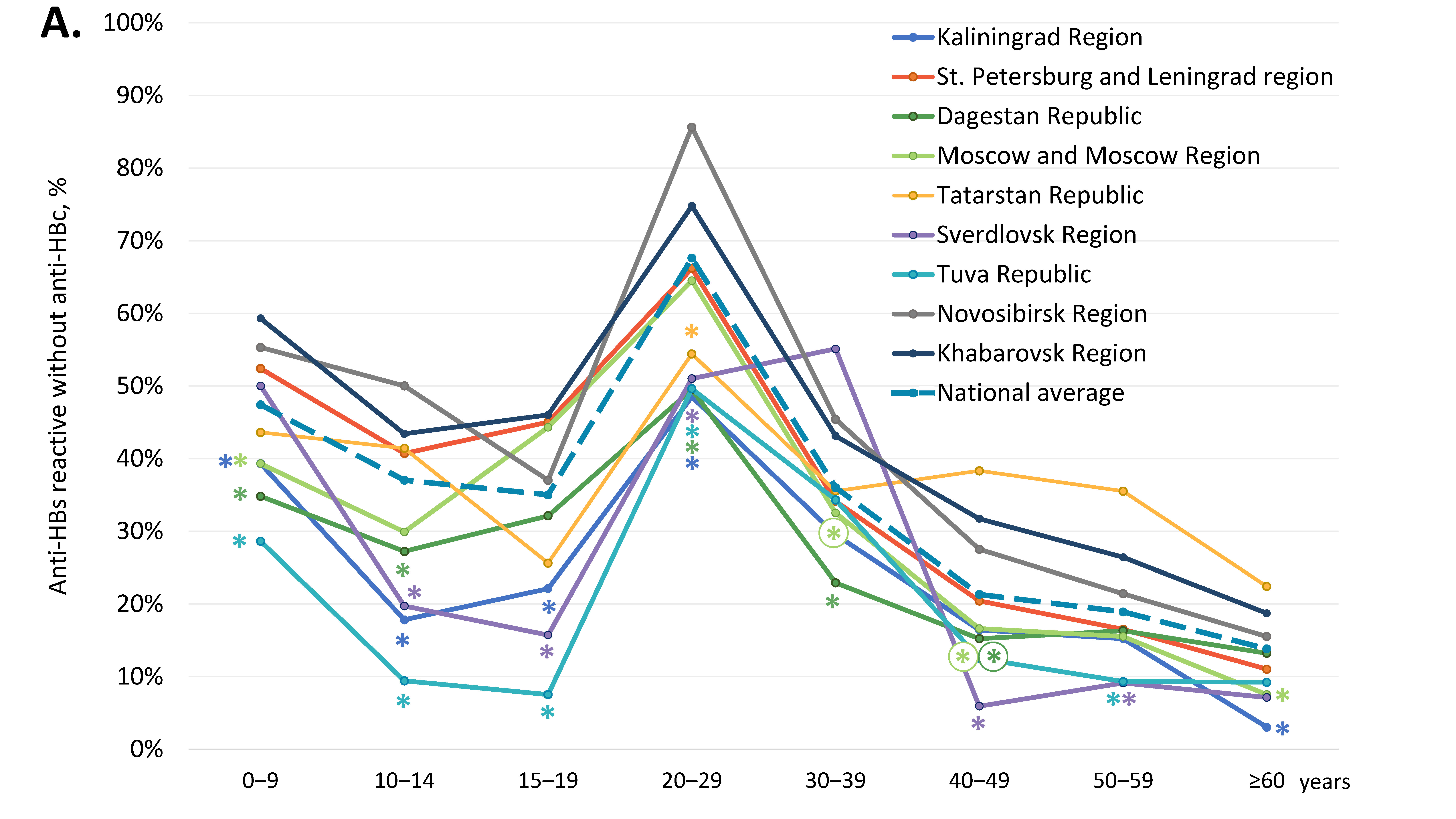
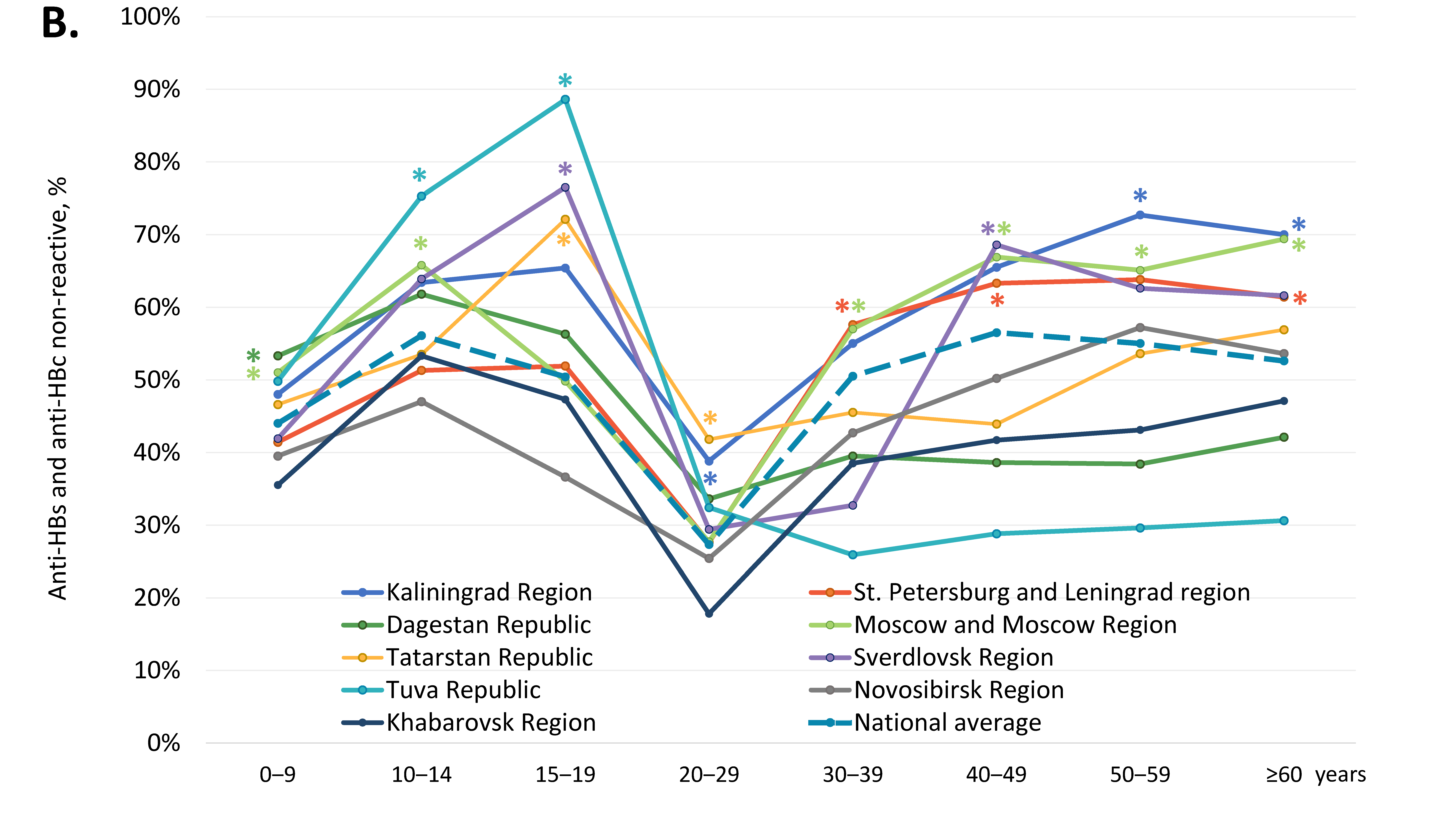
3.2. Vaccine-Induced Humoral HBV Immunity
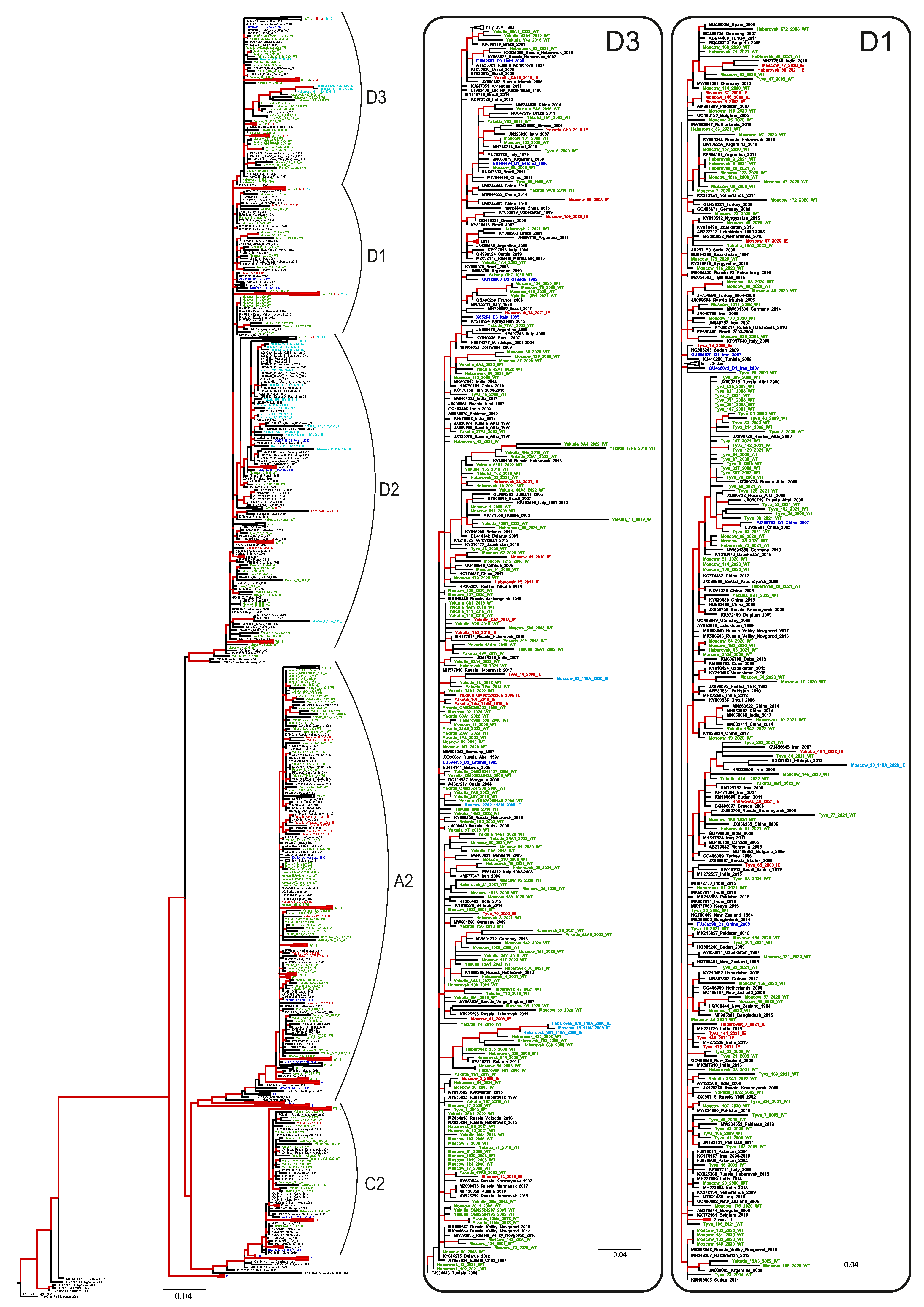
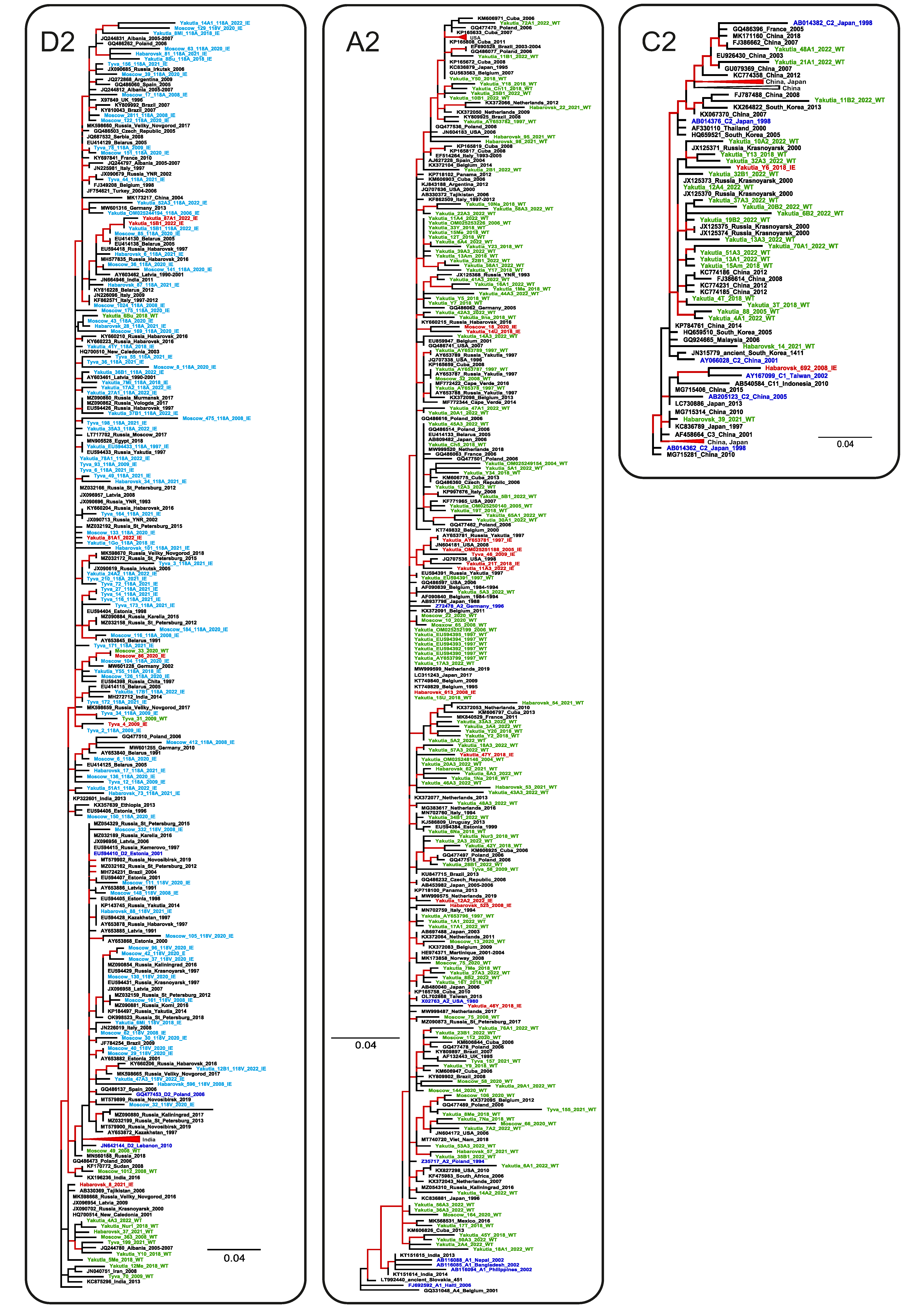
3.3. Prevalence of HBV Genotypes, Serotypes, and Vaccine-Escape Variants
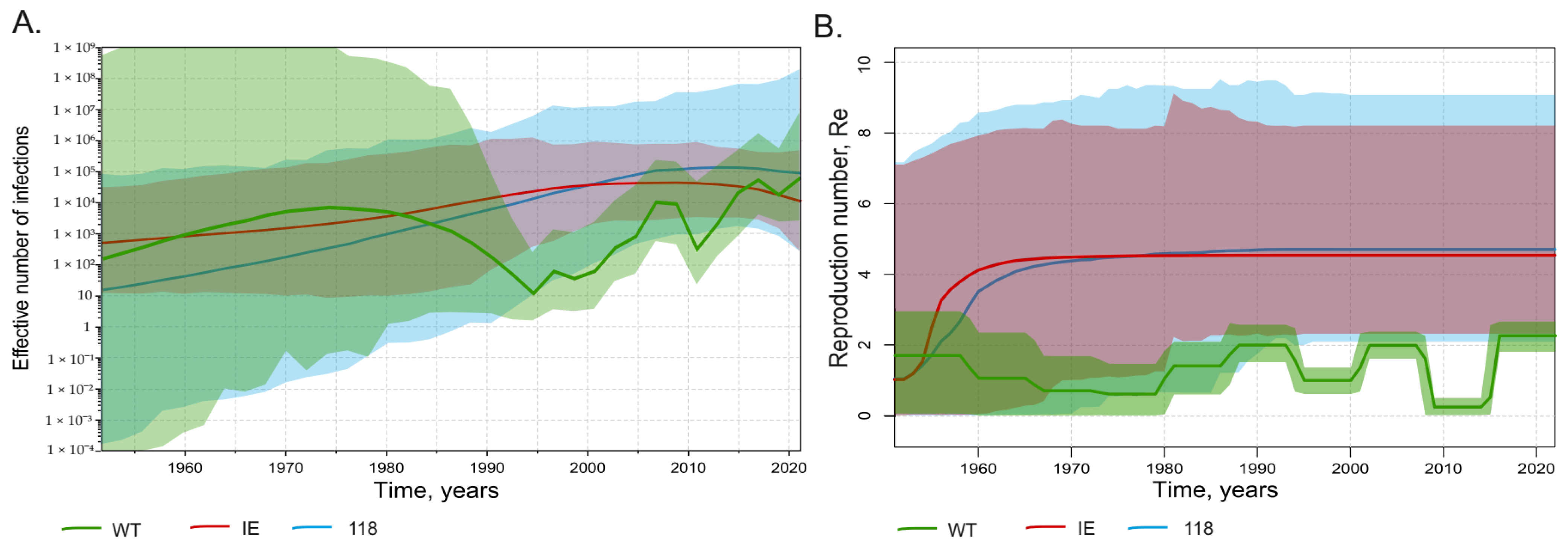
3.4. Comparison of Population Dynamics of Wild-Type HBV and Immune Escape Variants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Global Health Sector Strategy on Viral Hepatitis 2016–2021. Towards Ending Viral Hepatitis; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Prevention of Mother-to-Child Transmission of Hepatitis B Virus: Guidelines on Antiviral Prophylaxis in Pregnancy. Available online: https://www.who.int/publications-detail-redirect/978-92-4-000270-8 (accessed on 11 December 2022).
- Resolution No. 9 Dated 03.11.2013 of the Chief Sanitary Doctor of the Russian Federation. Available online: https://www.rospotrebnadzor.ru/documents/details.php?ELEMENT_ID=227 (accessed on 6 January 2023). (In Russian).
- Federal Center of Hygiene and Epidemiology. Federal Statistical Form 1. Infectious and Parasitic Diseases Morbidity in Russian Federation; Federal Center of Hygiene and Epidemiology: Moscow, Russia, 2021. (In Russian) [Google Scholar]
- Kyuregyan, K.K.; Kichatova, V.S.; Isaeva, O.V.; Potemkin, I.A.; Malinnikova, E.Y.; Lopatukhina, M.A.; Karlsen, A.A.; Asadi Mobarhan, F.A.; Mullin, E.V.; Slukinova, O.S.; et al. Coverage with Timely Administered Vaccination against Hepatitis B Virus and Its Influence on the Prevalence of HBV Infection in the Regions of Different Endemicity. Vaccines 2021, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Cooreman, M.P.; Leroux-Roels, G.; Paulij, W.P. Vaccine- and Hepatitis B Immune Globulin-Induced Escape Mutations of Hepatitis B Virus Surface Antigen. J. Biomed. Sci. 2001, 8, 237–247. [Google Scholar] [CrossRef]
- Gao, S.; Duan, Z.-P.; Coffin, C.S. Clinical Relevance of Hepatitis B Virus Variants. World J. Hepatol. 2015, 7, 1086–1096. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Chang, M.-H.; Ni, Y.-H.; Chen, H.-L. Survey of Hepatitis B Surface Variant Infection in Children 15 Years after a Nationwide Vaccination Programme in Taiwan. Gut 2004, 53, 1499–1503. [Google Scholar] [CrossRef]
- Klushkina, V.V.; Kyuregyan, K.K.; Kozhanova, T.V.; Popova, O.E.; Dubrovina, P.G.; Isaeva, O.V.; Gordeychuk, I.V.; Mikhailov, M.I. Impact of Universal Hepatitis B Vaccination on Prevalence, Infection-Associated Morbidity and Mortality, and Circulation of Immune Escape Variants in Russia. PLoS ONE 2016, 11, e0157161. [Google Scholar] [CrossRef] [PubMed]
- Hajian-Tilaki, K. Sample Size Estimation in Epidemiologic Studies. Casp. J. Intern. Med. 2011, 2, 289–298. [Google Scholar]
- Basuni, A.A.; Carman, W.F. HBV Vaccine-Escape Variants. Methods Mol. Med. 2004, 95, 115–124. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Norder, H.; Couroucé, A.-M.; Magnius, L.O.Y. 1992 Molecular Basis of Hepatitis B Virus Serotype Variations within the Four Major Subtypes. J. Gen. Virol. 1992, 73, 3141–3145. [Google Scholar] [CrossRef]
- Magnius, L.; Mason, W.S.; Taylor, J.; Kann, M.; Glebe, D.; Dény, P.; Sureau, C.; Norder, H. Ictv Report Consortium, null ICTV Virus Taxonomy Profile: Hepadnaviridae. J. Gen. Virol. 2020, 101, 571–572. [Google Scholar] [CrossRef]
- Kramvis, A. Genotypes and Genetic Variability of Hepatitis B Virus. Intervirology 2014, 57, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Karlsen, A.A.; Kyuregyan, K.K.; Isaeva, O.V.; Kichatova, V.S.; Asadi Mobarkhan, F.A.; Bezuglova, L.V.; Netesova, I.G.; Manuylov, V.A.; Pochtovyi, A.A.; Gushchin, V.A.; et al. Different Evolutionary Dynamics of Hepatitis B Virus Genotypes A and D, and Hepatitis D Virus Genotypes 1 and 2 in an Endemic Area of Yakutia, Russia. BMC Infect. Dis. 2022, 22, 452. [Google Scholar] [CrossRef] [PubMed]
- Peto, T.J.; Mendy, M.E.; Lowe, Y.; Webb, E.L.; Whittle, H.C.; Hall, A.J. Efficacy and effectiveness of infant vaccination against chronic hepatitis B in the Gambia Hepatitis Intervention Study (1986-90) and in the nationwide immunisation program. BMC Infect. Dis. 2014, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.H.; Huang, L.M.; Chang, M.H.; Yen, C.J.; Lu, C.Y.; You, S.L.; Kao, J.H.; Lin, Y.C.; Chen, H.L.; Hsu, H.Y.; et al. Two decades of universal hepatitis B vaccination in Taiwan: Impact and implication for future strategies. Gastroenterology 2007, 132, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Zacharakis, G.; Kotsiou, S.; Papoutselis, M.; Vafiadis, N.; Tzara, F.; Pouliou, E.; Maltezos, E.; Koskinas, J.; Papoutselis, K. Changes in the epidemiology of hepatitis B virus infection following the implementation of immunisation programmes in northeastern Greece. Eurosurveillance 2009, 14, 19297. [Google Scholar] [CrossRef]
- Nelson, N.P.; Easterbrook, P.J.; McMahon, B.J. Epidemiology of Hepatitis B Virus Infection and Impact of Vaccination on Disease. Clin. Liver Dis. 2016, 20, 607–628. [Google Scholar] [CrossRef]
- Lu, X.; Quinn, H.E.; Menzies, R.I.; Hueston, L.; Gilbert, L.; McIntyre, P.B. Hepatitis B Seroepidemiology in Australia, One Decade After Universal Vaccination of Infants and Adolescents. Infect. Disord. Drug Targets 2020, 20, 341–347. [Google Scholar] [CrossRef]
- Romano, L.; Zanetti, A.R. Hepatitis B Vaccination: A Historical Overview with a Focus on the Italian Achievements. Viruses. 2022, 14, 1515. [Google Scholar] [CrossRef]
- Le, M.H.; Yeo, Y.H.; Cheung, R.; Henry, L.; Lok, A.S.; Nguyen, M.H. Chronic Hepatitis B Prevalence Among Foreign-Born and U.S.-Born Adults in the United States, 1999–2016. Hepatology 2020, 71, 431–443. [Google Scholar] [CrossRef]
- Melardo, C.; Foglia, F.; Della Rocca, M.T.; Zaino, A.; Morone, M.V.; De Filippis, A.; Finamore, E.; Galdiero, M. Hepatitis B Virus prevalence and serological profiles in a hospital in Southern Italy. New Microbiol. 2022, 45, 243–248. [Google Scholar]
- Huang, L.F.; Lin, Z.Q.; Yang, X.H.; Zhang, H.R.; Wang, F.Z.; Wang, F.; Wu, J.N.; Zhou, Y. Association of vaccine policy with virus infection and awareness of hepatitis B in Fujian, China. Hum. Vaccin Immunother. 2022, 18, 2153535. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Liu, C.; Chen, T.; Wang, Y.; Fan, C.; Lu, L.; Lu, F.; Qu, C. Neonatal Hepatitis B Vaccination Protects Mature Adults from Occult Virus Infection. Hepatol. Int. 2021, 15, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Li, T.; Allain, J.-P.; Zhou, B.; Zhang, Y.; Zhong, M.; Fu, Y.; Li, C. Low Occurrence of HBsAg but High Frequency of Transient Occult HBV Infection in Vaccinated and HBIG-Administered Infants Born to HBsAg Positive Mothers. J. Med. Virol. 2017, 89, 2130–2137. [Google Scholar] [CrossRef] [PubMed]
- Makvandi, M. Update on Occult Hepatitis B Virus Infection. World J. Gastroenterol. 2016, 22, 8720–8734. [Google Scholar] [CrossRef] [PubMed]
- Allain, J.-P. Occult Hepatitis B Virus Infection: Implications in Transfusion. Vox Sang. 2004, 86, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Takuissu, G.R.; Kenmoe, S.; Amougou Atsama, M.; Atenguena Okobalemba, E.; Mbaga, D.S.; Ebogo-Belobo, J.T.; Bowo-Ngandji, A.; Oyono, M.G.; Magoudjou-Pekam, J.N.; Kame-Ngasse, G.I.; et al. Global Epidemiology of Occult Hepatitis B Virus Infections in Blood Donors, a Systematic Review and Meta-Analysis. PLoS ONE 2022, 17, e0272920. [Google Scholar] [CrossRef] [PubMed]
- Raheel, M.; Choga, W.T.; Blackard, J.T. The Distribution of Hepatitis B Virus Surface Antigen Polymorphisms at Positions Associated with Vaccine Escape. J. Med. Virol. 2020, 92, 3336–3343. [Google Scholar] [CrossRef]
- Mozhgani, S.H.; Malekpour, S.A.; Norouzi, M.; Ramezani, F.; Rezaee, S.A.; Poortahmasebi, V.; Sadeghi, M.; Alavian, S.M.; Zarei-Ghobadi, M.; Ghaziasadi, A.; et al. Molecular evolution and phylodynamics of hepatitis B virus infection circulating in Iran. Arch. Virol. 2018, 163, 1479–1488. [Google Scholar] [CrossRef]
- Lin, S.Y.; Toyoda, H.; Kumada, T.; Liu, H.F. Molecular Evolution and Phylodynamics of Acute Hepatitis B Virus in Japan. PLoS ONE 2016, 11, e0157103. [Google Scholar] [CrossRef]
- Zehender, G.; Svicher, V.; Gabanelli, E.; Ebranati, E.; Veo, C.; Lo Presti, A.; Cella, E.; Giovanetti, M.; Bussini, L.; Salpini, R.; et al. Reliable timescale inference of HBV genotype A origin and phylodynamics. Infect. Genet. Evol. 2015, 32, 361–369. [Google Scholar] [CrossRef]
- Li, T.; Chen, H.; Sun, L.; Liu, J. Dating the origin and dispersal of global hepatitis B virus genotype C in humans. Drug Discov. Ther. 2022, 16, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zuo, R.; Wang, J.; Shen, T. Characteristics and evolutionary history of hepatitis B virus quasi-subgenotype B3 in Southeast Asia. Virus Res. 2019, 273, 197762. [Google Scholar] [CrossRef] [PubMed]
- Carman, W.F. The Clinical Significance of Surface Antigen Variants of Hepatitis B Virus. J. Viral Hepat. 1997, 4 (Suppl. 1), 11–20. [Google Scholar] [CrossRef] [PubMed]
- Golsaz-Shirazi, F.; Asadi-Asadabad, S.; Sarvnaz, H.; Mehdi Amiri, M.; Hojjat-Farsangi, M.; Chudy, M.; Jeddi-Tehrani, M.; Shokri, F. Immunoreactivity Pattern of Monoclonal Antibodies against Hepatitis B Vaccine with Global Hepatitis B Virus Genotypes. Clin. Chim. Acta 2020, 510, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Alavian, S.M.; Carman, W.F.; Jazayeri, S.M. HBsAg Variants: Diagnostic-Escape and Diagnostic Dilemma. J. Clin. Virol. 2013, 57, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Tanaka, Y. Cross-Protection of Hepatitis B Vaccination among Different Genotypes. Vaccines 2020, 8, 456. [Google Scholar] [CrossRef]
- List of Vaccines for the Prevention of Infectious Diseases Registered and Approved for Use in the Russian Federation; Federal Service for Surveillance on Consumer Rights Protection and HumanWellbeing: Moscow, Russia. 2020. Available online: http://www.28.rospotrebnadzor.ru/activity/?p=1145&show_year=2008 (accessed on 12 December 2022). (In Russian).
- Lin, Y.-C.; Chang, M.-H.; Ni, Y.-H.; Hsu, H.-Y.; Chen, D.-S. Long-Term Immunogenicity and Efficacy of Universal Hepatitis B Virus Vaccination in Taiwan. J. Infect. Dis. 2003, 187, 134–138. [Google Scholar] [CrossRef]
- Le, M.H.; Yeo, Y.H.; So, S.; Gane, E.; Cheung, R.C.; Nguyen, M.H. Prevalence of Hepatitis B Vaccination Coverage and Serologic Evidence of Immunity Among US-Born Children and Adolescents From 1999 to 2016. JAMA Netw. Open 2020, 3, e2022388. [Google Scholar] [CrossRef]
- Huynh, C.; Minuk, G.Y.; Uhanova, J.; Baikie, M.; Wong, T.; Osiowy, C. Serological and Molecular Epidemiological Outcomes after Two Decades of Universal Infant Hepatitis B Virus (HBV) Vaccination in Nunavut, Canada. Vaccine 2017, 35, 4515–4522. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, Q.; Xu, B.; Zhou, Z.; Wang, Z.; Zhou, Y.H. Influence of maternal antibody against hepatitis B surface antigen on active immune response to hepatitis B vaccine in infants. Vaccine 2008, 26, 6064–6067. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, S.; Luo, C.; Wu, Q.; Liu, Q.; Zhou, Y.H.; Hu, Y. Transplacentally acquired maternal antibody against hepatitis B surface antigen in infants and its influence on the response to hepatitis B vaccine. PLoS ONE 2011, 6, e25130. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-K.; Nie, J.-J.; Li, J.; Zhuang, H. The Effect of HLA on Immunological Response to Hepatitis B Vaccine in Healthy People: A Meta-Analysis. Vaccine 2013, 31, 4355–4361. [Google Scholar] [CrossRef] [PubMed]
- West, D.J.; Calandra, G.B. Vaccine Induced Immunologic Memory for Hepatitis B Surface Antigen: Implications for Policy on Booster Vaccination. Vaccine 1996, 14, 1019–1027. [Google Scholar] [CrossRef] [PubMed]
- Hepatitis B Vaccines: WHO Position Paper—July 2017. Available online: https://www.who.int/publications-detail-redirect/WER9227 (accessed on 12 December 2022).
- Stramer, S.L.; Wend, U.; Candotti, D.; Foster, G.A.; Hollinger, F.B.; Dodd, R.Y.; Allain, J.-P.; Gerlich, W. Nucleic Acid Testing to Detect HBV Infection in Blood Donors. N. Engl. J. Med. 2011, 364, 236–247. [Google Scholar] [CrossRef]
- Sanitary Rules and Norms SanPiN 3.3686-21 “Sanitary and Epidemiological Requirements for the Prevention of Infectious Diseases”. Available online: https://docs.cntd.ru/document/573660140 (accessed on 12 December 2022). (In Russian).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Region | Reactive for HBsAg, % (95% CI) | Reactive for Anti-HBc, % (95% CI) | Reactive for Anti-HBs, but Non-Reactive for Anti-HBc, % (95% CI) | Non-Reactive for Either Anti-HBs or Anti-HBc, % (95% CI) |
|---|---|---|---|---|
| Kaliningrad Region | 0.4% (0.1–1.0%) | 17.1% (15.0–19.5%) * | 27.0% (24.4–29.7%) * | 57.5% (54.5–60.5%) * |
| St. Petersburg and Leningrad Region | 0.4% (0.2–0.6%) | 10.2% (9.4–11.1%) | 40.4% (39.1–41.7%) | 49.4% (48.1–50.7%) * |
| Republic of Dagestan | 2.4% (2.0–2.9%) * | 23.1% (21.9–24.3%) * | 29.8% (28.5–31.1%) * | 47.1% (45.7–48.5%) |
| Moscow and Moscow Region | 0.4% (0.3–0.5%) | 11.3% (10.6–12.0%) | 37.5% (36.5–38.5%) | 51.1% (50.0–52.2%) * |
| Republic of Tatarstan | 0.2% (0.1–0.9%) | 11.5% (9.5–13.9%) | 37.1% (33.8–40.4%) | 51.0% (47.6–54.4%) * |
| Sverdlovsk Region | 0.6% (0.3–1.4%) | 16.5% (14.2–19.0%) | 27.7% (24.9–30.7%) * | 55.9% (52.7–59.1%) * |
| Tuva Republic | 1.2% (0.7–2.1%) | 31.7% (29.1–34.5%) * | 22.0% (19.6–24.5%) * | 46.0% (43.1–49.0%) |
| Novosibirsk Region | 0.8% (0.6–1.0%) | 11.8% (11.1–12.5%) | 46.7% (45.6–47.8%) | 41.5% (40.4–42.6%) |
| Khabarovsk Region | 0.7% (0.5–0.9%) | 14.1% (13.2–15.0%) | 47.3% (45.9–48.7%) | 38.7% (37.4–40.0%) |
| National average | 0.8% (0.7–0.9%) | 14.2% (13.8–14.5%) | 39.4% (38.9–39.9%) | 46.5% (46.0–47.0%) |
| Study Region | Year of Sample Collection and Number of HBV Sequences, n | HBV Genotypes/ Serotypes, n (%) | aa Substitutions in HBsAg Associated with Immune Escape | |||||
|---|---|---|---|---|---|---|---|---|
| aa Substitution * | Number of Sequences Carrying aa Substitution | The Proportion of Sequences with aa Substitution | ||||||
| Per HBV Genotype ** | Total per Region ** | Total per HBV Genotype, Excluding aa Substitutions in Position 118 ** | Total per Region, Excluding aa Substitutions in Position 118 ** | |||||
| Moscow Region | 2008–2010, n = 55 | A/ad, n = 3 (5.5%) | n.d. | 0 | 0/3 | 0/3 | ||
| D/ay, n = 52 (94.5%) | T118A/V/M | 6/5/1 | 18/52 (34.6%) | 18/55 (32.7%) | 6/52 (11.5%) | 6/55 (10.1%) | ||
| P120Q | 1 | |||||||
| T131I/P | 3/1 | |||||||
| D144E | 1 | |||||||
| 2020, n = 136 | A/ad, n = 11 (8.1%) | G130N | 1 | 1/11 | 1/11 | |||
| D/ay, n = 125 (91.9%) | T118A/V | 21/11 | 44/125 (35.2%) | 45/136 (33.1%) | 12/125 (9.6%) | 13/136 (9.6%) | ||
| P120Q | 1 | |||||||
| Q129H/L/P | 1/2 | |||||||
| G130R/S | 1/1 | |||||||
| T131I/N | 1/1 | |||||||
| M133T/I | 3 | |||||||
| G145R | 1 | |||||||
| Tuva Republic | 2008, n = 76 | A/ad, n = 2 (2.6%) | M133T | 1 | 1/2 | 1/2 | ||
| D/ay, n = 74 (97.4%) | T118A | 13 | 19/74 (25.7%) | 20/76 (26.3%) ▪ | 6/74 (8.1%) | 7/76 (9.2%) | ||
| P120T | 1 | |||||||
| T131N | 1 | |||||||
| M133L/K/I | 1/2 | |||||||
| G145S | 1 | |||||||
| 2021, n = 49 | A/ad, n = 2 (4.1%) | n.d. | 0 | 0/2 | 0/2 | |||
| D/ay, n = 47 (95.9%) | T118A | 19 | 22/47 (46.8%) | 22/49 (44.9%)▪ | 3/47 (6.4%) | 3/49 (6.1%) | ||
| P120S | 3 | |||||||
| Sakha Republic (Yakutia) | 1997–2009, n = 58 | A/ad, n = 33 (56.9%) | T126S | 2 | 4/33 (12.1%) | 4/33 (12.1%) | ||
| M133T | 2 | |||||||
| C/ad, n = 2 (3.4%) | n.d. | 0 | 0/2 | 0/2 | ||||
| D/ay, n = 23 (39.7%) | T118A | 2 | 7/23 (30.4%) | 11/58 (19.0%) | 5/23 (21.7%) | 9/58 (15.5%) | ||
| T118A+P120I | 1 | |||||||
| P120S/T | 2 | |||||||
| M133T | 1 | |||||||
| K141Q | 1 | |||||||
| 2018–2022, n = 231 | A/ad, n = 94 (40.7%) | P120T | 1 | 7/94 (7.4%) ▪▪ | 36/231 (15.6%) | 7/94 (7.4%) | 14/231 (6.1%) | |
| Q129R | 1 | |||||||
| Q129P | 1 | |||||||
| G130N | 1 | |||||||
| M133T | 2 | |||||||
| C/ad, n = 21 (9.1%) | M133I | 1 | 1/21 (4.8%) | 1/21 (4.8%) | ||||
| D/ay, n = 116 (50.2%) | T118A/V | 19/4 | 29/116 (25.0%) ▪▪ | 6/116 (5.2%) | ||||
| T118A+Q130R | 1 | |||||||
| T118M+ P120S | 1 | |||||||
| P120S | 1 | |||||||
| G130N+T131H | 1 | |||||||
| T131I/P | 1/1 | |||||||
| Khabarovsk Region | 2008, n = 16 | D/ay, n = 16 (100%) | T118A/V | 1/2 | 4/16 (25%) | 4/16 (25%) | 0/16 (0%) | 0/16 (0%) |
| 2021, n = 65 | A/ad, n = 7 (10.8%) | n.d. | 0 | 0/7 | 0/7 | |||
| C/ad, n = 2 (3.1%) | n.d. | 0 | 0/2 | 0/2 | ||||
| D/ay, n = 56 (86.1%) | T118A/V | 7/2 | 18/56 (32.1%) | 18/65 (27.7%) | 9/56 (16.1%) | 9/65 (13.8%) | ||
| T118A+G130A | 1 | |||||||
| P120S | 1 | |||||||
| T123I | 1 | |||||||
| T126N+D144E | 1 | |||||||
| G130S | 1 | |||||||
| T131N | 1 | |||||||
| M133S/T | 3 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asadi Mobarkhan, F.A.; Manuylov, V.A.; Karlsen, A.A.; Kichatova, V.S.; Potemkin, I.A.; Lopatukhina, M.A.; Isaeva, O.V.; Mullin, E.V.; Mazunina, E.P.; Bykonia, E.N.; et al. Post-Vaccination and Post-Infection Immunity to the Hepatitis B Virus and Circulation of Immune-Escape Variants in the Russian Federation 20 Years after the Start of Mass Vaccination. Vaccines 2023, 11, 430. https://doi.org/10.3390/vaccines11020430
Asadi Mobarkhan FA, Manuylov VA, Karlsen AA, Kichatova VS, Potemkin IA, Lopatukhina MA, Isaeva OV, Mullin EV, Mazunina EP, Bykonia EN, et al. Post-Vaccination and Post-Infection Immunity to the Hepatitis B Virus and Circulation of Immune-Escape Variants in the Russian Federation 20 Years after the Start of Mass Vaccination. Vaccines. 2023; 11(2):430. https://doi.org/10.3390/vaccines11020430
Chicago/Turabian StyleAsadi Mobarkhan, Fedor A., Victor A. Manuylov, Anastasia A. Karlsen, Vera S. Kichatova, Ilya A. Potemkin, Maria A. Lopatukhina, Olga V. Isaeva, Eugeniy V. Mullin, Elena P. Mazunina, Evgeniia N. Bykonia, and et al. 2023. "Post-Vaccination and Post-Infection Immunity to the Hepatitis B Virus and Circulation of Immune-Escape Variants in the Russian Federation 20 Years after the Start of Mass Vaccination" Vaccines 11, no. 2: 430. https://doi.org/10.3390/vaccines11020430