
Competition of SARS-CoV-2 Variants in Cell Culture and Tissue: Wins the Fastest Viral Autowave


Abstract
:1. Introduction
- Can these experiments be described by the mathematical model (the simplest possible one)?
- Could the situations of one strain overtaking another strain after some prolonged time interval be observed qualitatively if the spatial effects are taken into account?
- Could spatial effects be very important: for example, could they result in any new information that cannot be obtained in the homogeneous system?

2. Methods
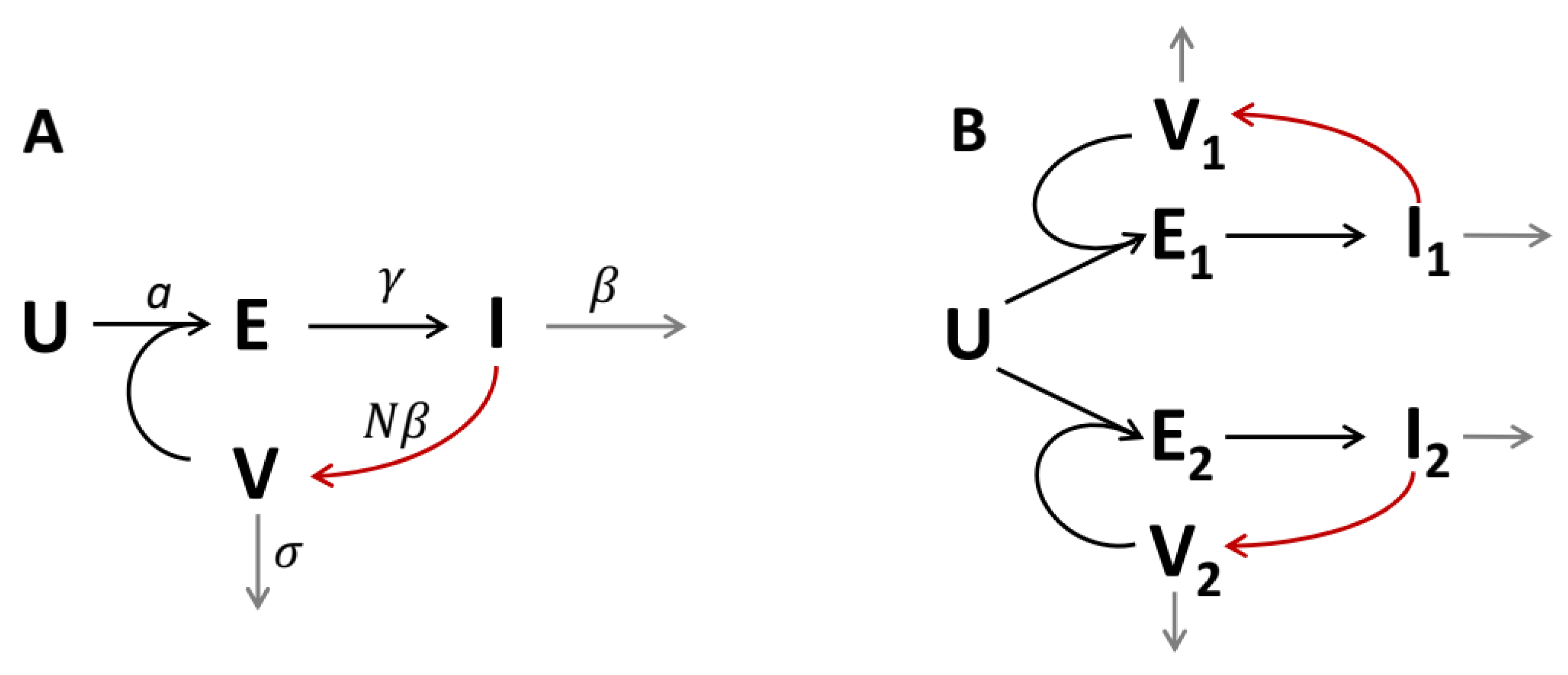
2.1. Model Description and Governing Equations
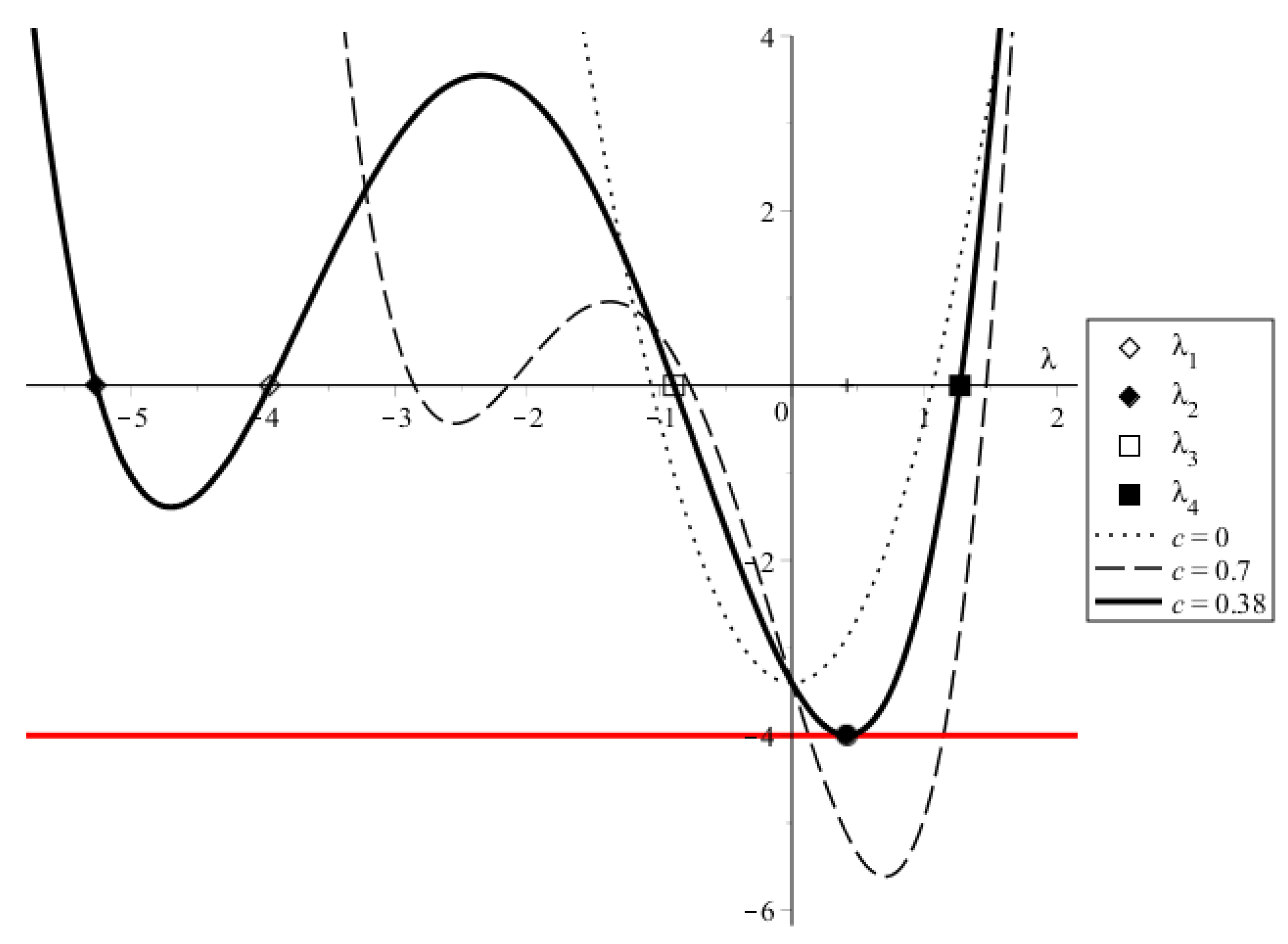
2.2. Steady Travelling Solution
2.3. Two-Strain Model with Competition for Cells
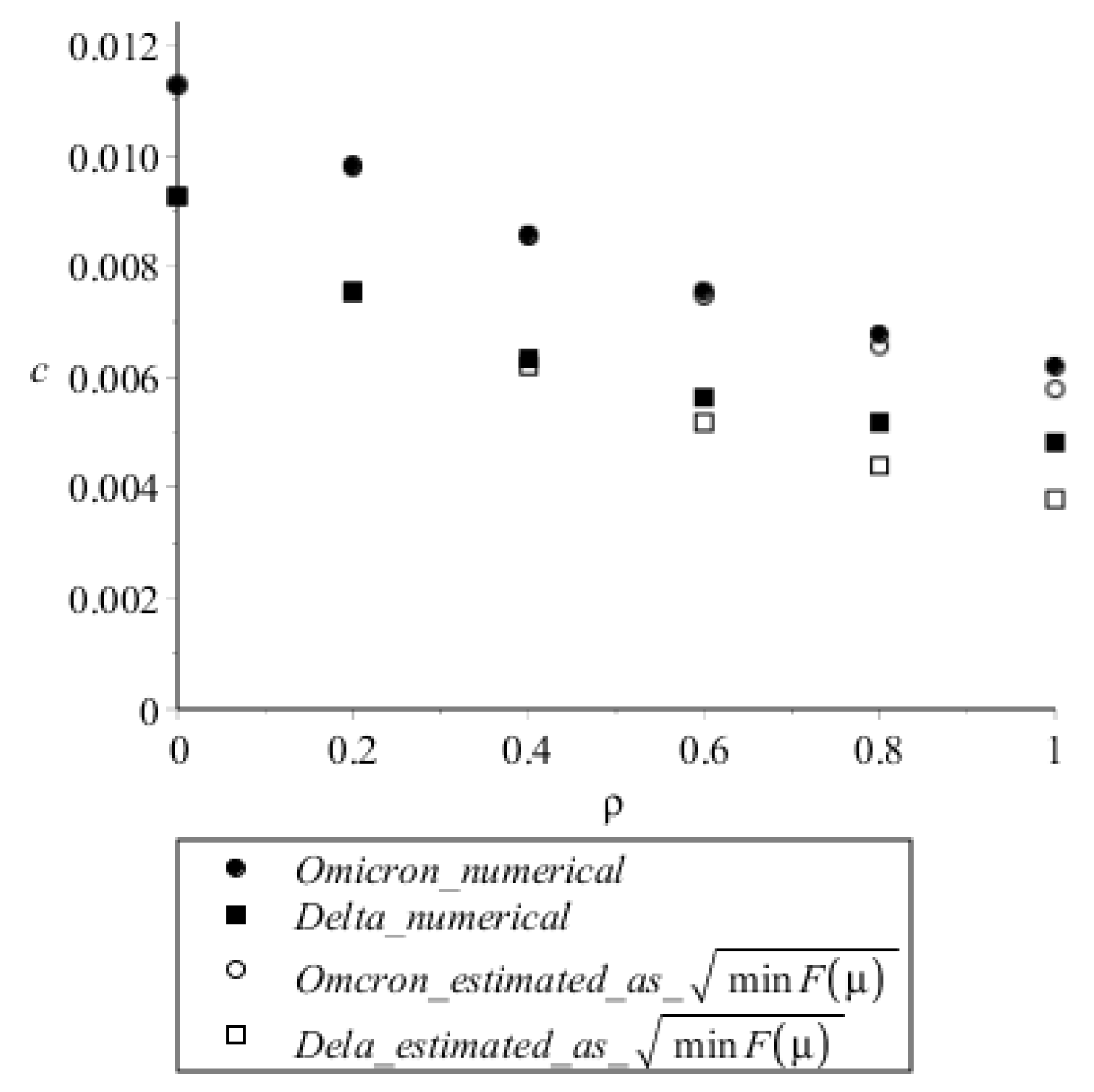
2.4. Model Parameters and Comparison with Experimental Data
- Arbitrary non-dimensional values (typically, of the order of 1) for plotting the explanatory graphs when deriving analytical formulas in the Appendices. For this set, in numerical calculations we take .
- Parameters which correspond to the SARS-CoV-2 variants Delta and Omicron replicating in human nasal epithelial cultures (hNECs) in the in vitro experiment [2] (see Table 1). We estimated them from physical reasons (e.g., D), derived/estimated from experimental results and kept fixed (e.g., , ), or varied under the strict limitations as described below:
- The diffusion coefficient D was estimated using the Stockes–Einstein formula at 300 K assuming virus diameter 100 nm [16] and water viscosity.
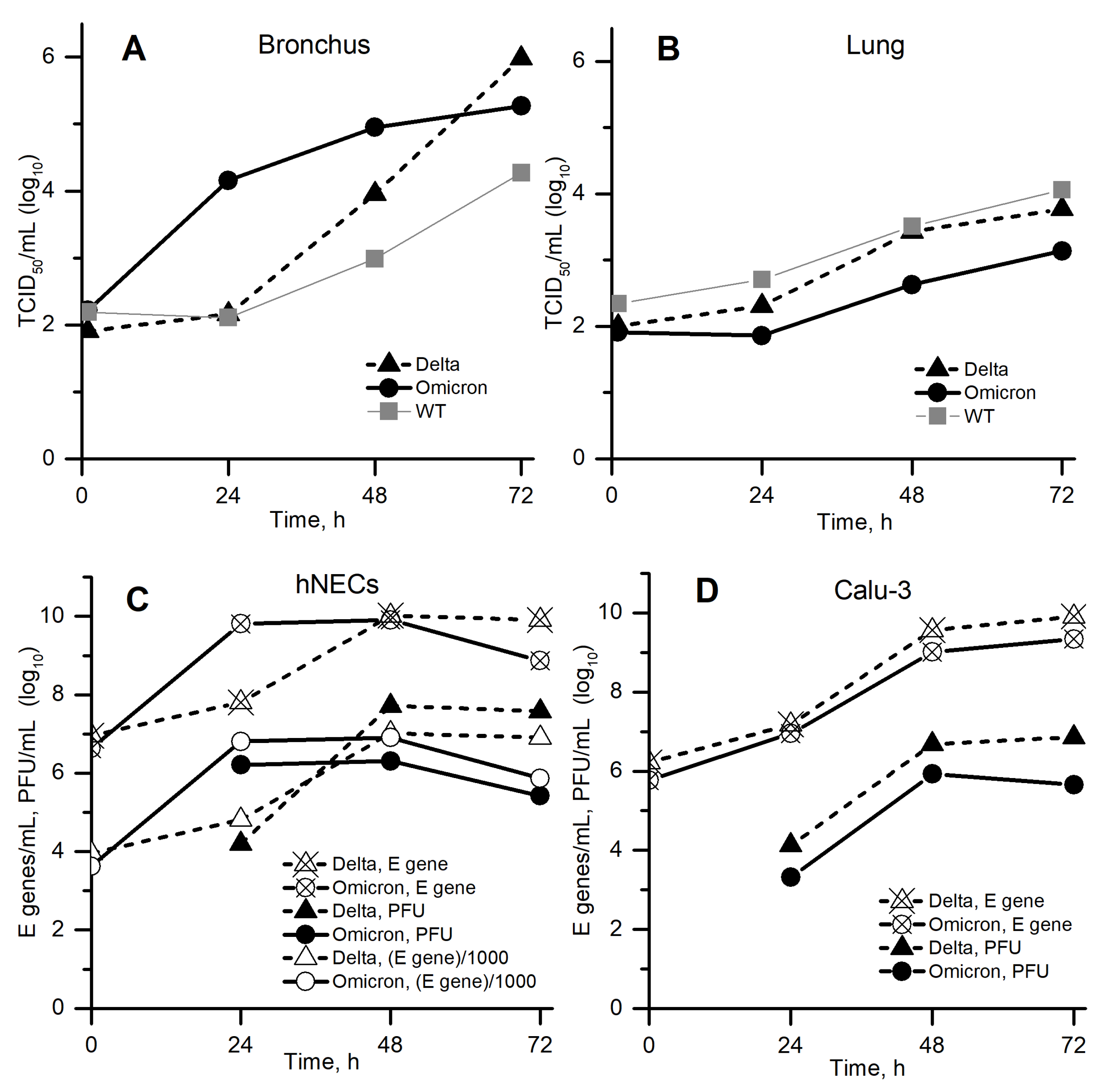
- The experimental in vitro system in [2] is considered as homogeneous, because (a) only average concentrations are presented in that article, (b) the full size of the experimental system is 6.5 mm [17], which is comparable to the width of the front in our numerical calculations (about 1–3 mm, see Figure 5 below), and (c) substantial convection should be expected during inoculation and everyday sampling.
- Since the virus concentration determined by RT-qPCR for the E gene in [2] (see Figure 2C,D) was approximately 1000 times greater than the PFU concentration for all available time points and cell lineages (crossed vs. filled markers), that is only 1 out of 1000 viral particles was able to effectively infect a cell and reproduce with its help, we compared the model variable with the united data for (filled markers) and (open markers). In particular, we estimated as .
- Initial cell concentration was limited in the range .
- The characteristic time of infection () was assumed to be 1 h for both strains. Characteristic times if transition () and cell death () were limited as 2 h and 0.1 h from below, respectively. Virus death times () were limited in the range [0.1, 100] h.
- To avoid large differences between the corresponding parameters of Delta and Omicron variants, the rations , , and were limited to the range of from to 10.
- After the fitting, all parameters were rounded up to 1 decimal digit, and those that were close to each other were set as equal.
- For the obtained set of kinetic parameters, in spatial numerical calculations (with diffusion) we set L = 2 cm or more to be able to track the transition of autowave to the steady propagation regime.
2.5. Numerical Methods
3. Results
3.1. Virus Replication Number Provides the Condition for the Infection Progression,
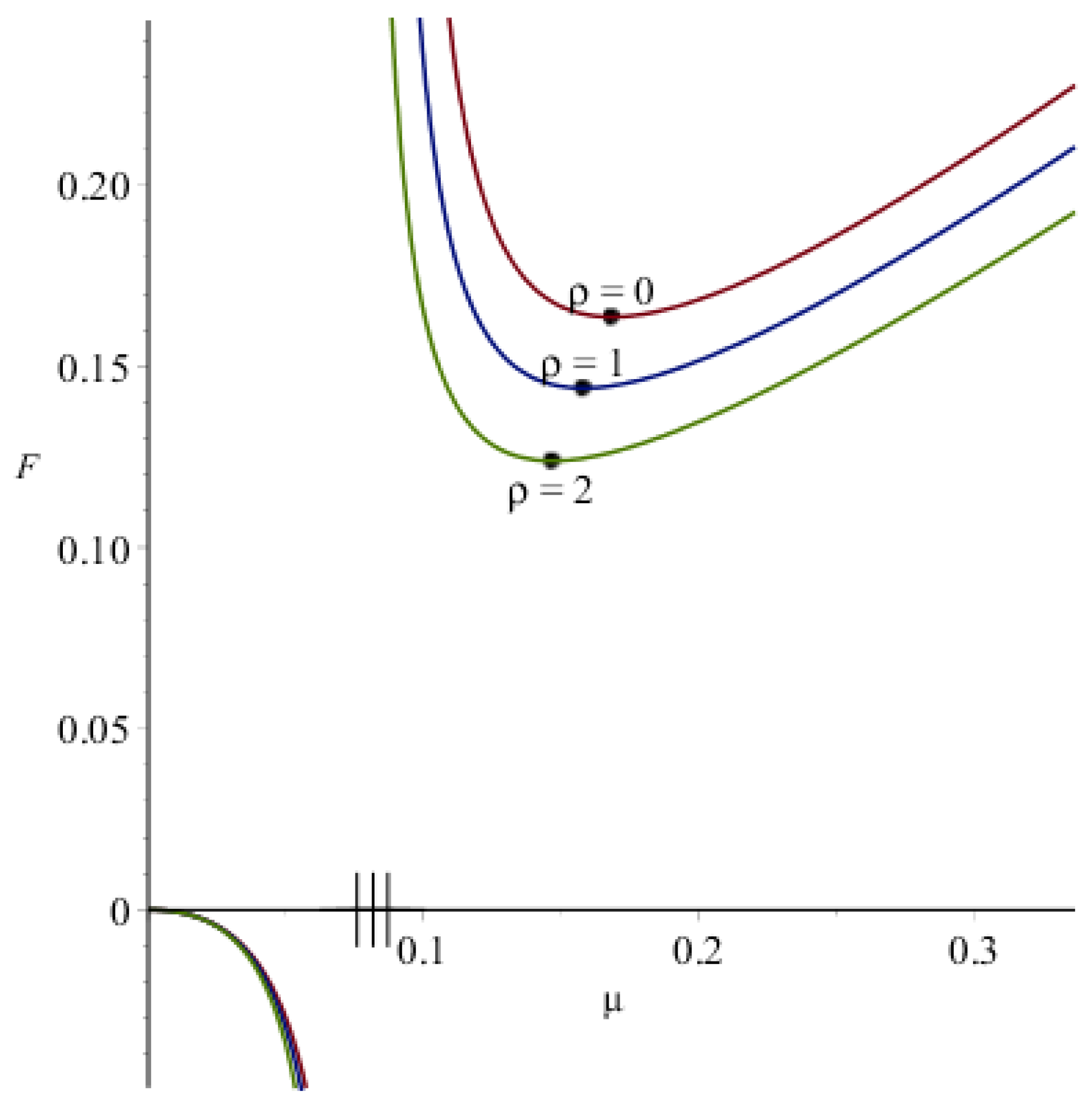
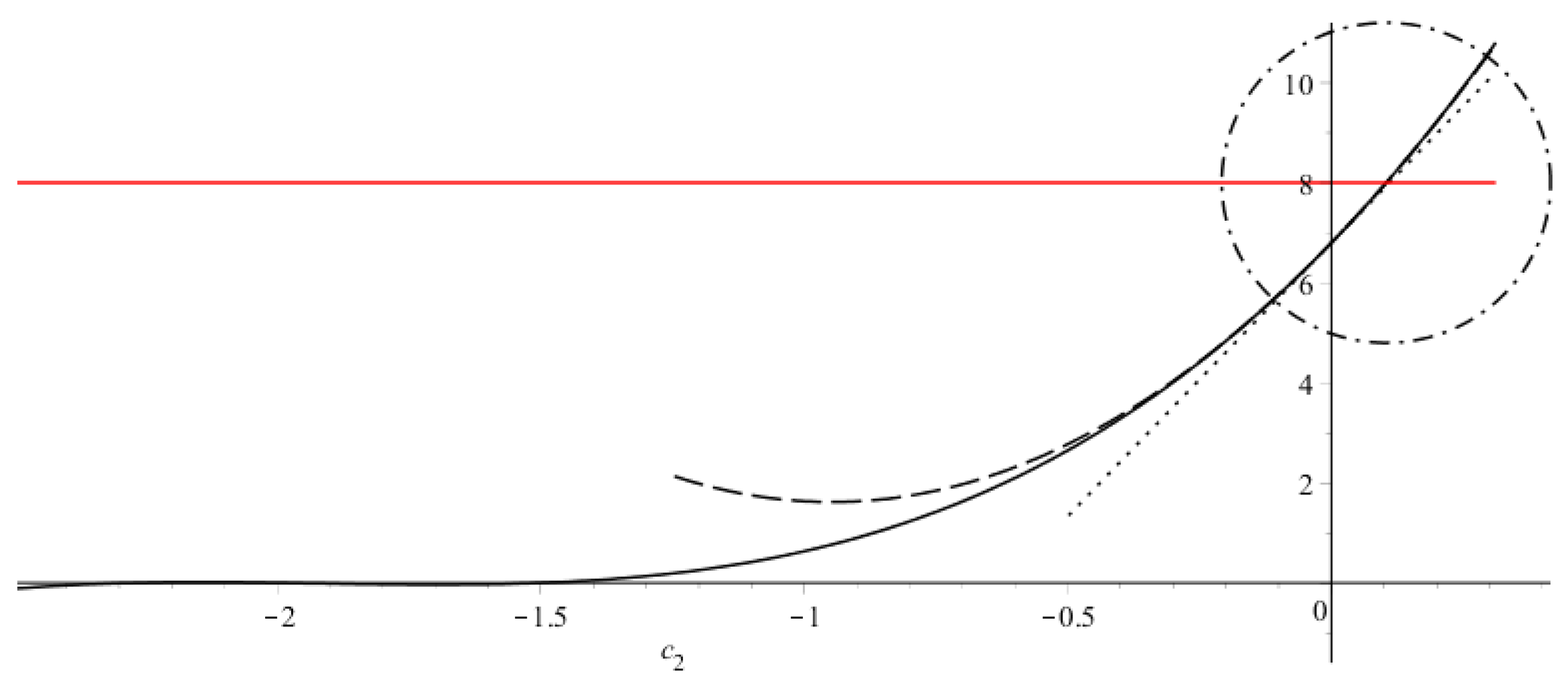
3.2. Estimates of the Steady Wave Speed
3.3. Equations for the Final Concentration of Intact Cells and the Total Spatial Viral Load
3.4. Homogeneous Case without Competition: Omicron Is “Quick” and Wins the Start but Delta Can Overtake It after 1–2 Days’ Lag
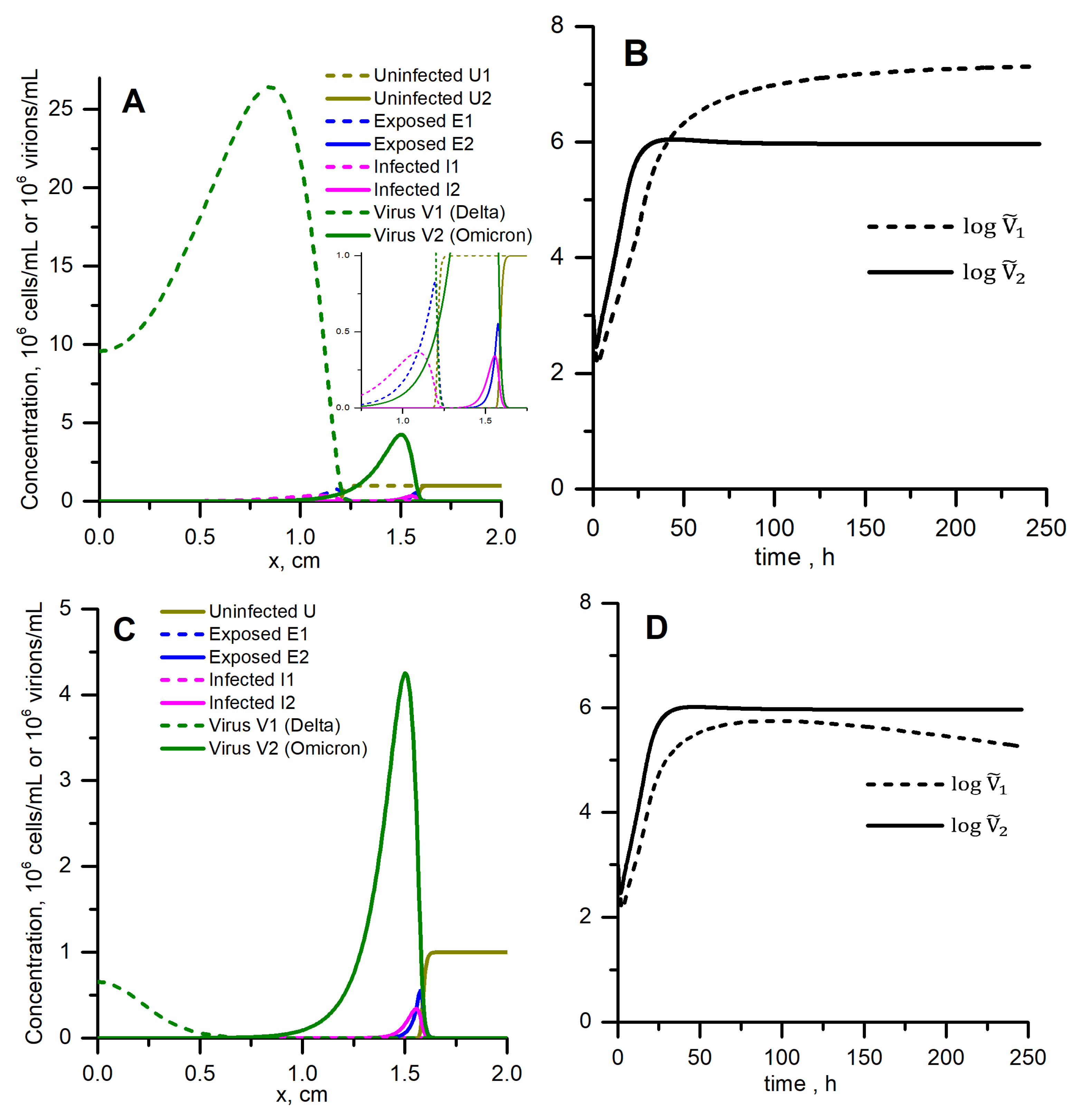
3.5. Spatially Distributed Case without Competition: Omicron Can Win the Race despite Low Concentration and
3.6. Spatially Distributed Case with Competition: Omicron Can Win and Completely Suppress Delta
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| URT | upper respiratory tract |
| LRT | lower respiratory tract |
| hNECs | culture of human nasal epithelial cells |
| KPP or KPPF equation | Kolmogorov–Petrovskii–Piskunov–Fisher equation |
Appendix A
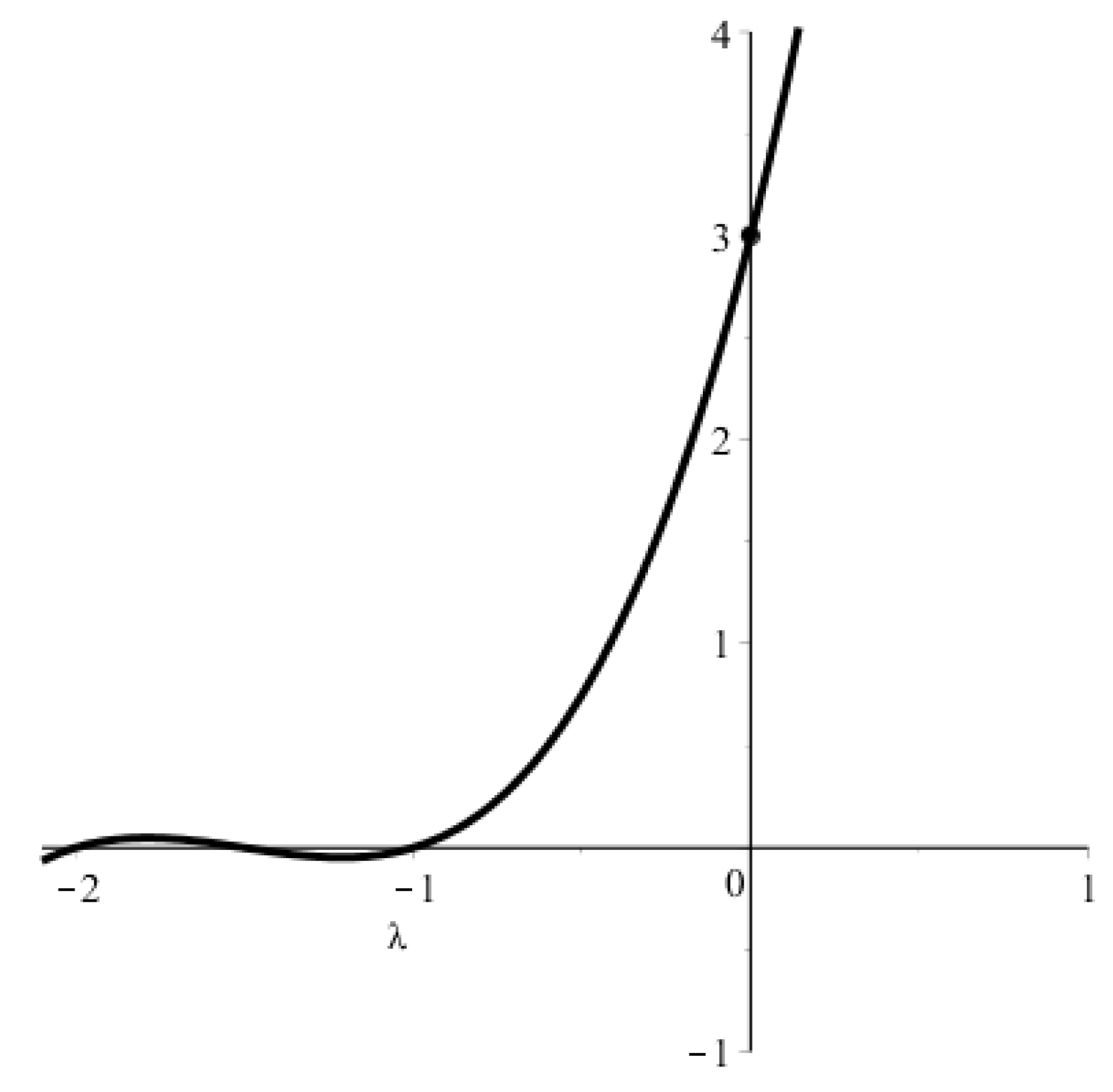
Appendix A.1. Condition for Existence of a Positive Solution to Equation (5)

Appendix A.2. Estimates of the Steady Wave Speed




Appendix A.3. Derivation of Equations for the Final Concentration of Intact Cells and the Total Spatial Viral Load
References
- Hui, K.P.; Ho, J.C.; Cheung, M.c.; Ng, K.c.; Ching, R.H.; Lai, K.l.; Kam, T.T.; Gu, H.; Sit, K.Y.; Hsin, M.K.; et al. SARS-CoV-2 Omicron variant replication in human bronchus and lung ex vivo. Nature 2022, 603, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Peacock, T.P.; Brown, J.C.; Zhou, J.; Thakur, N.; Newman, J.; Kugathasan, R.; Sukhova, K.; Kaforou, M.; Bailey, D.; Barclay, W.S. The SARS-CoV-2 variant, Omicron, shows rapid replication in human primary nasal epithelial cultures and efficiently uses the endosomal route of entry. bioRxiv (version posted January 3) 2022. preprint. [Google Scholar] [CrossRef]
- Murray, J. Mathematical Biology. I. An Introduction, 3rd ed.; Springer: New York, NY, USA, 2002. [Google Scholar] [CrossRef]
- Murray, J. Mathematical Biology. II. Spatial Models and Biomedical Applications, 3rd ed.; Springer: New York, NY, USA, 2003. [Google Scholar] [CrossRef]
- Flerlage, T.; Boyd, D.F.; Meliopoulos, V.; Thomas, P.G.; Schultz-Cherry, S. Influenza virus and SARS-CoV-2: Pathogenesis and host responses in the respiratory tract. Nat. Rev. Microbiol. 2021, 19, 425–441. [Google Scholar] [CrossRef] [PubMed]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.J.; Okuda, K.; Edwards, C.E.; Martinez, D.R.; Asakura, T.; Dinnon, K.H.; Kato, T.; Lee, R.E.; Yount, B.L.; Mascenik, T.M.; et al. SARS-CoV-2 Reverse Genetics Reveals a Variable Infection Gradient in the Respiratory Tract. Cell 2020, 182, 429–446.e14. [Google Scholar] [CrossRef] [PubMed]
- Ga̧secka, A.; Borovac, J.A.; Guerreiro, R.A.; Giustozzi, M.; Parker, W.; Caldeira, D.; Chiva-Blanch, G. Thrombotic Complications in Patients with COVID-19: Pathophysiological Mechanisms, Diagnosis, and Treatment. Cardiovasc. Drugs Ther. 2021, 35, 215–229. [Google Scholar] [CrossRef] [PubMed]
- WHO. Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants (accessed on 13 June 2022).
- Dashkevich, N.M.; Ovanesov, M.V.; Balandina, A.N.; Karamzin, S.S.; Shestakov, P.I.; Soshitova, N.P.; Tokarev, A.A.; Panteleev, M.A.; Ataullakhanov, F.I. Thrombin activity propagates in space during blood coagulation as an excitation wave. Biophys. J. 2012, 103, 2233–2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ataullakhanov, F.; Guriia, G. Spatial aspects of the dynamics of blood coagulation. I. Hypothesis. Biophysics 1994, 39, 89–96. [Google Scholar]
- Tokarev, A.; Ratto, N.; Volpert, V. Mathematical modeling of thrombin generation and wave propagation: From simple to complex models and backwards. In BIOMAT 2018 International Symposium on Mathematical and Computational Biology; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Bocharov, G.; Meyerhans, A.; Bessonov, N.; Trofimchuk, S.; Volpert, V. Interplay between reaction and diffusion processes in governing the dynamics of virus infections. J. Theor. Biol. 2018, 457, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Mahiout, L.; Mozokhina, A.; Tokarev, A.; Volpert, V. Virus replication and competition in a cell culture: Application to the SARS-CoV-2 variants. Appl. Math. Lett. 2022, 113, 108217. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Bar-On, Y.M.; Gleizer, S.; Bernshtein, B.; Flamholz, A.; Phillips, R.; Milo, R. The total number and mass of SARS-CoV-2 virions. Proc. Natl. Acad. Sci. USA 2021, 118, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Epithelix. FAQ, Epithelix’s Tissues. Available online: https://www.epithelix.com/faq (accessed on 13 June 2022).
- Hoops, S.; Sahle, S.; Gauges, R.; Lee, C.; Pahle, J.; Simus, N.; Singhal, M.; Xu, L.; Mendes, P.; Kummer, U.; et al. COPASI—A COmplex PAthway SImulator. Bioinformatics 2006, 22, 3067–3074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokarev, A.A. Velocity–Amplitude Relationship in the Gray–Scott Autowave Model in Isolated Conditions. ACS Omega 2019, 4, 14430–14438. [Google Scholar] [CrossRef] [PubMed]
- Hairer, E.; Nørsett, S.; Wanner, G. Solving Ordinary Differential Equations I. Nonstiff Problems (Russian Translation, 1990); Mir: Moscow, Russia; Springer: Berlin/Heidelberg, Germany; New York, NY, USA; London, UK; Paris, France; Tokyo, Japan, 1987. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Dimension | Delta | Omicron |
|---|---|---|---|
| Equal for both strains: | |||
| h | 1 | ||
| D | |||
| N | |||
| 1 | |||
| Delta | Omicron | Delta | Omicron | |
|---|---|---|---|---|
| Numerical c | 0.0049 | 0.0062 | 0.0094 | 0.011 |
| as , Equation (6) | 0.0038 | 0.0058 | 0.0093 | 0.011 |
| c from Equation (7b) | 0.0069 | 0.0088 | 0.016 | 0.015 |
| Numerical | 24 | 0.93 | 46 | 1.88 |
| , Equations (6) and (9b) | 19 | 0.87 | 45 | 1.65 |
| Numerical | 26 | 4.3 | 27 | 4.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokarev, A.; Mozokhina, A.; Volpert, V. Competition of SARS-CoV-2 Variants in Cell Culture and Tissue: Wins the Fastest Viral Autowave. Vaccines 2022, 10, 995. https://doi.org/10.3390/vaccines10070995
Tokarev A, Mozokhina A, Volpert V. Competition of SARS-CoV-2 Variants in Cell Culture and Tissue: Wins the Fastest Viral Autowave. Vaccines. 2022; 10(7):995. https://doi.org/10.3390/vaccines10070995
Chicago/Turabian StyleTokarev, Alexey, Anastasia Mozokhina, and Vitaly Volpert. 2022. "Competition of SARS-CoV-2 Variants in Cell Culture and Tissue: Wins the Fastest Viral Autowave" Vaccines 10, no. 7: 995. https://doi.org/10.3390/vaccines10070995