

Antibodies Induced by Homologous or Heterologous Inactivated (CoronaVac/BBIBP-CorV) and Recombinant Protein Subunit Vaccines (ZF2001) Dramatically Enhanced Inhibitory Abilities against B.1.351, B.1.617.2, and B.1.1.529 Variants

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Cell Culture, Expression and Purification of SARS-CoV-2 Strains RBD, and hACE2 Recombinant Proteins
2.3. Enzyme-Linked Immunoassay of Competition
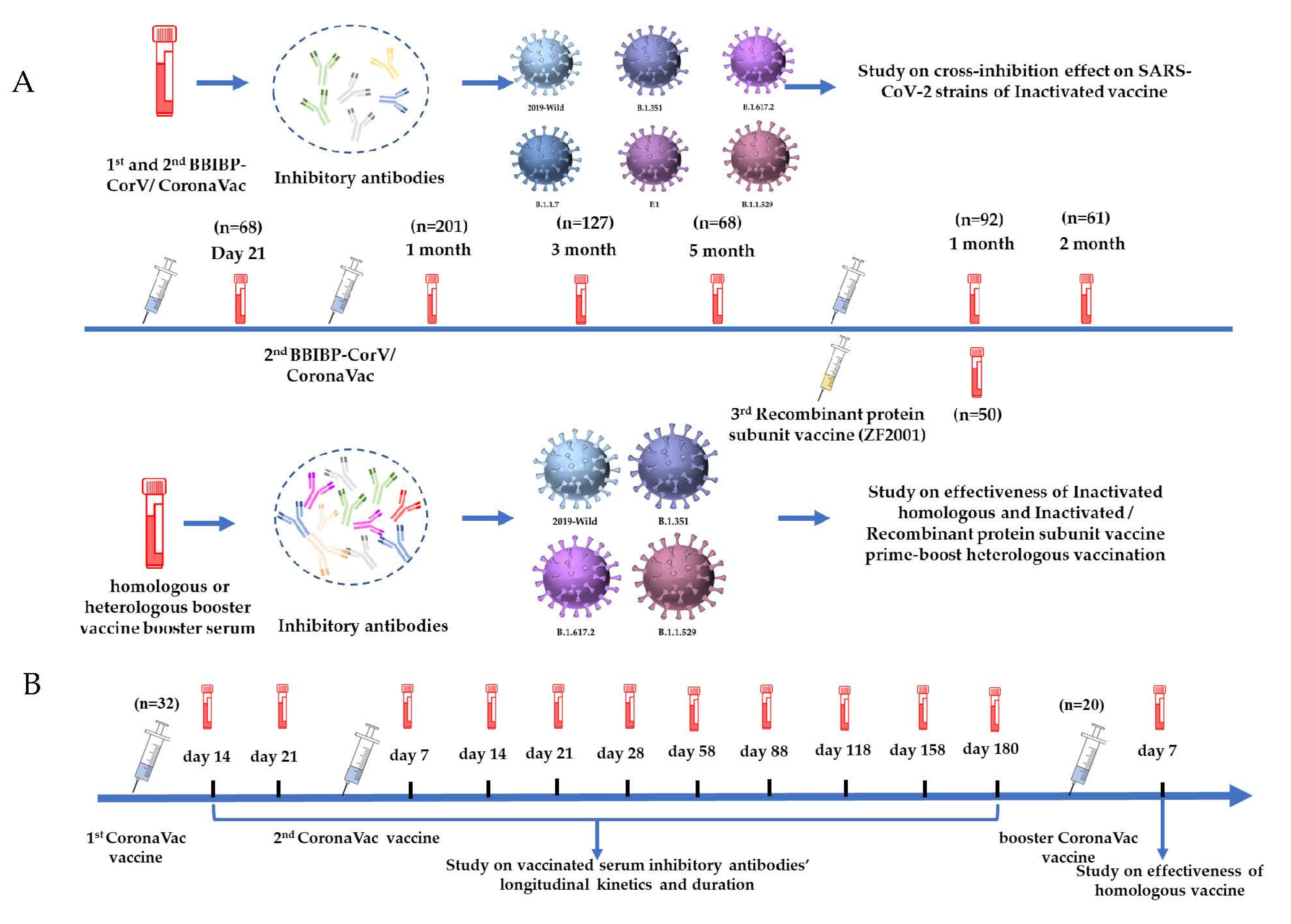
2.4. Detailed Studies on the Primary Inactivated Vaccine-Induced Inhibitory Antibodies Cross-Inhibition against Wild-Type and B.1.1.7, B.1.351, P.1, B.1.617.2, and B.1.1.529 Mutant Strains
2.5. Longitudinal Kinetics and Duration of Inactivated Vaccine-Induced Inhibitory Antibodies
2.6. Evaluation of Antibodies Inhibitory Ability with Inactivated Homologous and Inactivated/Recombinant Protein Subunit Vaccine Prime-Boost Heterologous Vaccination
2.7. Data Statistics
3. Results
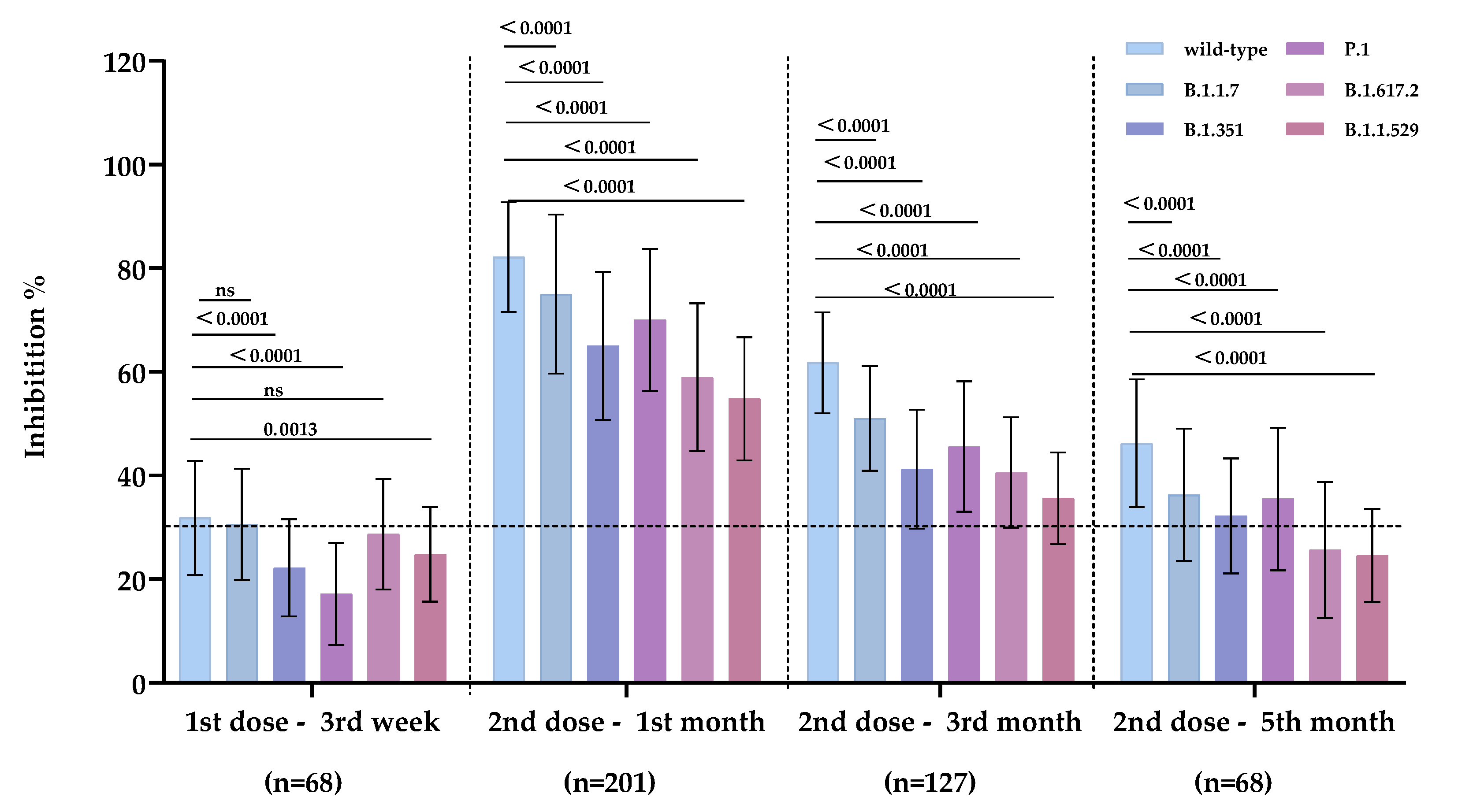
3.1. Inhibitory Antibody Induced by CoronaVac/BBIBP-CorV Exhibited the Highest Inhibition Capacity against SARS-CoV-2 Wild-Type and Cross-Inhibiting Activities against SARS-CoV-2 Mutant Strains
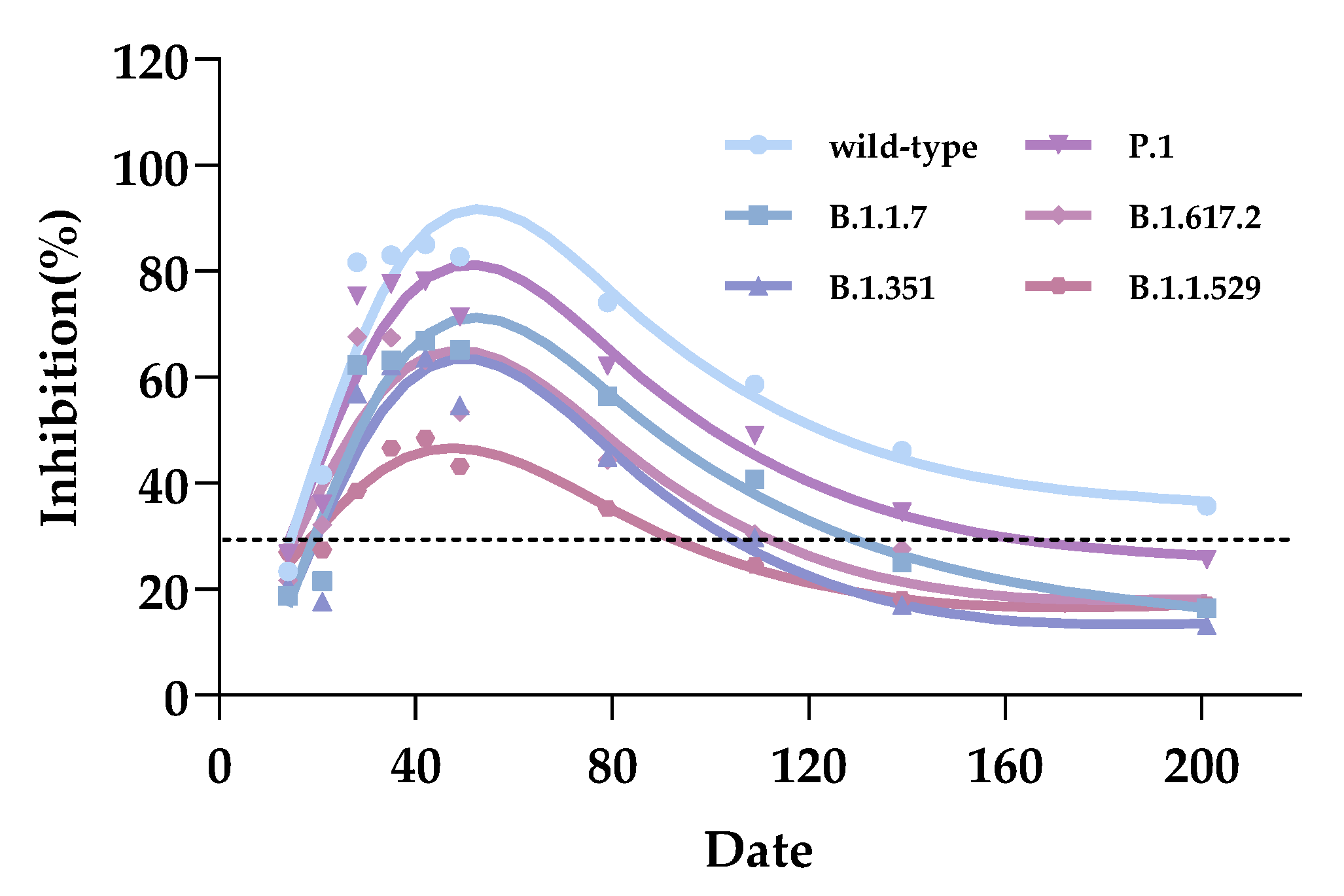
3.2. Longitudinal Kinetics and Duration Effectiveness of Vaccinated Serum Inhibiting Antibodies
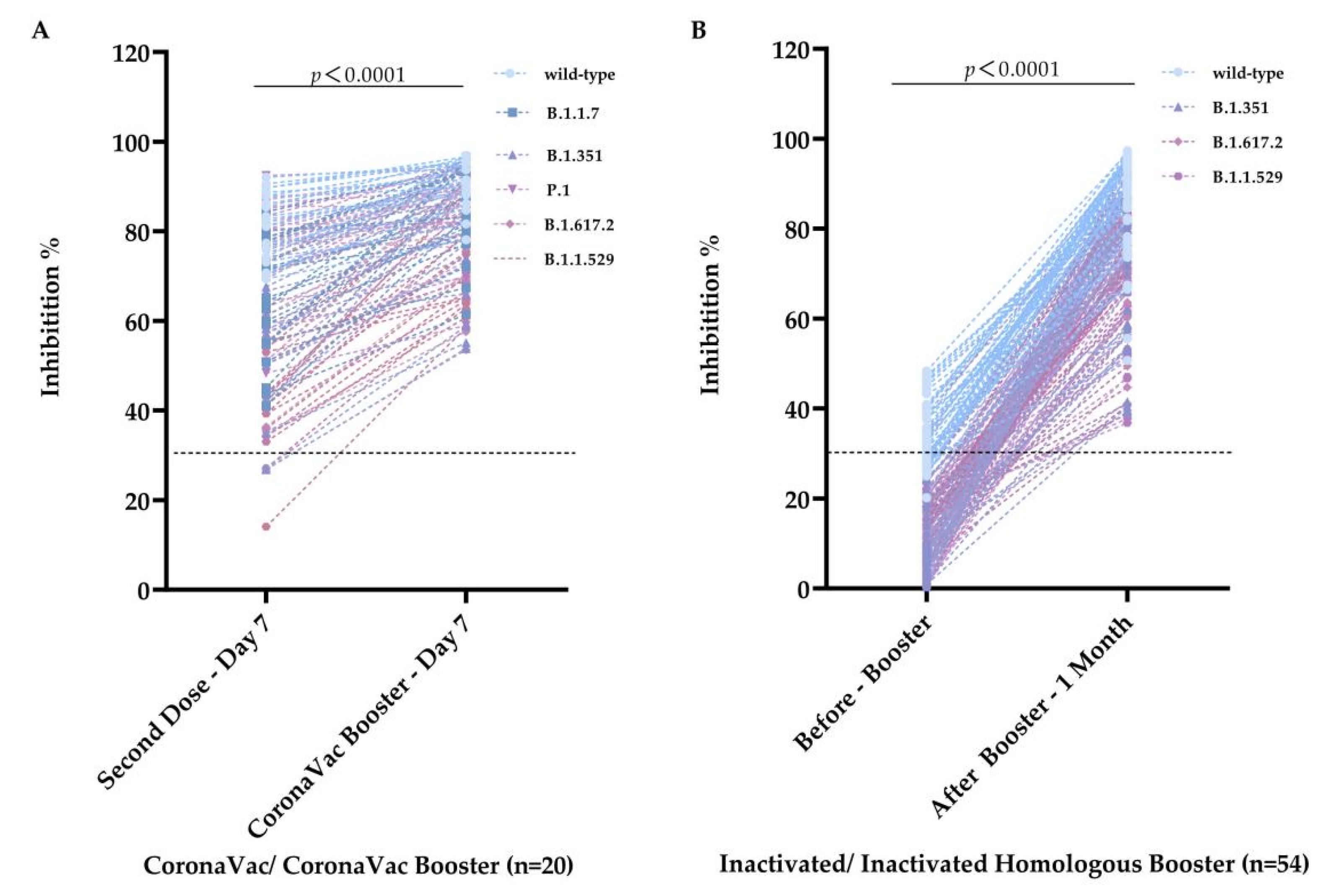
3.3. Vaccinated Serum Antibodies Showed Extensive Inhibitory Activities following Homologous Booster Vaccination
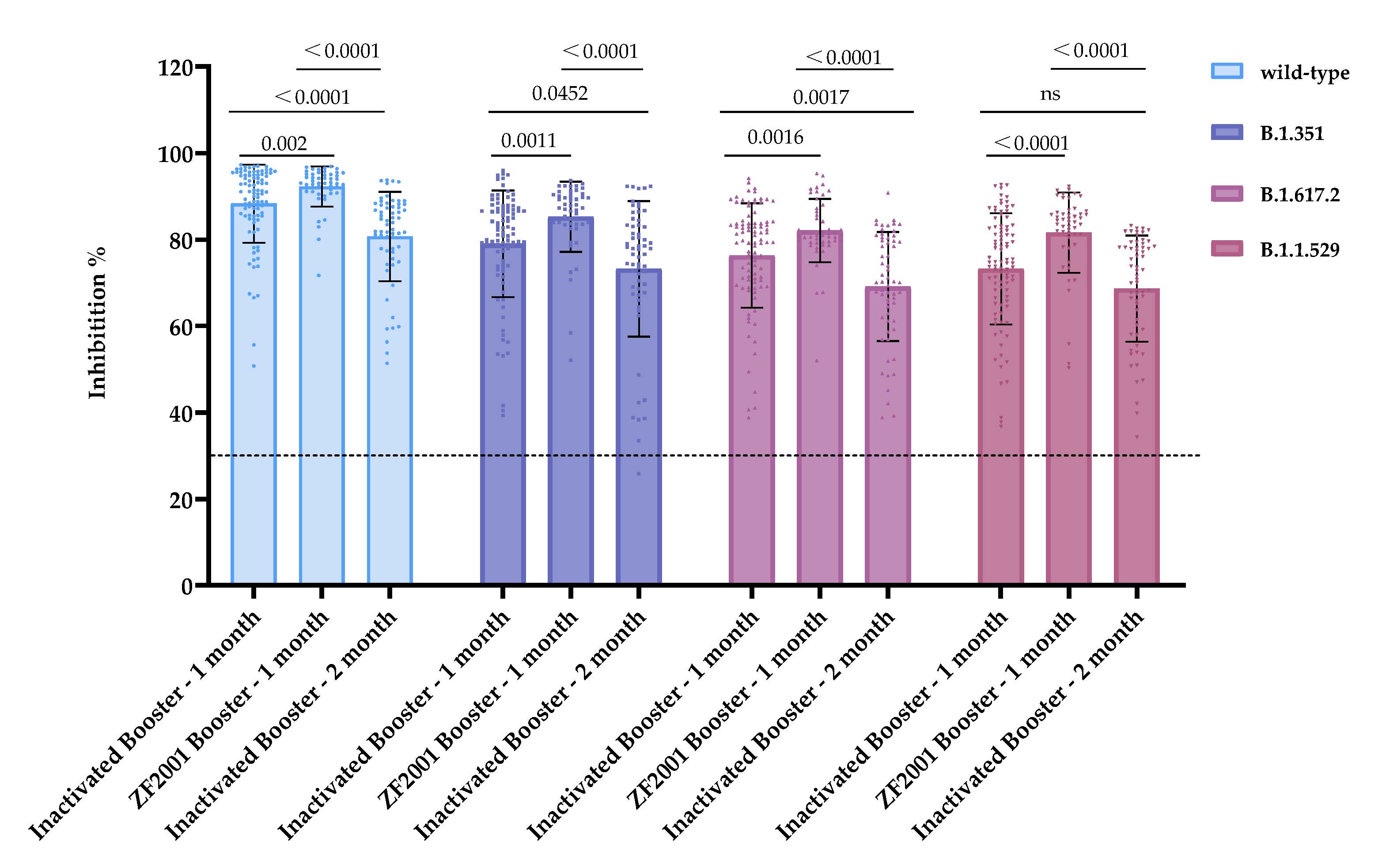
3.4. Heterologous Vaccination with the Recombinant Protein Subunit Vaccine Significantly Recalled and Increased the Humoral Immune Responses against Wild-Type, B.1.351, B.1.617.2, and B.1.1.529 Mutant Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 16 November 2022).
- Callaway, E. The race for coronavirus vaccines: A graphical guide. Nature 2020, 580, 576–577. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.-C.; Li, Y.-H.; Guan, X.-H.; Hou, L.-H.; Wang, W.-J.; Li, J.-X.; Wu, S.-P.; Wang, B.-S.; Wang, Z.; Wang, L.; et al. Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: A dose-escalation, open-label, non-randomised, first-in-human trial. Lancet 2020, 395, 1845–1854. [Google Scholar] [CrossRef]
- Piccoli, L.; Park, Y.-J.; Tortorici, M.A.; Czudnochowski, N.; Walls, A.C.; Beltramello, M.; Silacci-Fregni, C.; Pinto, D.; Rosen, L.E.; Bowen, J.E.; et al. Mapping Neutralizing and Immunodominant Sites on the SARS-CoV-2 Spike Receptor-Binding Domain by Structure-Guided High-Resolution Serology. Cell 2020, 183, 1024–1042.e21. [Google Scholar] [CrossRef] [PubMed]
- Karim, S.S.A.; de Oliveira, T. New SARS-CoV-2 Variants—Clinical, Public Health, and Vaccine Implications. N. Engl. J. Med. 2021, 384, 1866–1868. [Google Scholar] [CrossRef] [PubMed]
- Wibmer, C.K.; Ayres, F.; Hermanus, T.; Madzivhandila, M.; Kgagudi, P.; Oosthuysen, B.; Lambson, B.E.; de Oliveira, T.; Vermeulen, M.; van der Berg, K.; et al. SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. Nat. Med. 2021, 27, 622–625. [Google Scholar] [CrossRef]
- Tada, T.; Dcosta, B.M.; Samanovic-Golden, M.; Herati, R.S.; Cornelius, A.; Mulligan, M.J.; Landau, N.R. Neutralization of viruses with European, South African, and United States SARS-CoV-2 variant spike proteins by convalescent sera and BNT162b2 mRNA vaccine-elicited antibodies. bioRxiv 2021. preprint. [Google Scholar] [CrossRef]
- Wu, K.; Werner, A.P.; Moliva, J.I.; Koch, M.; Choi, A.; Stewart-Jones, G.B.E.; Bennett, H.; Boyoglu-Barnum, S.; Shi, W.; Graham, B.S.; et al. mRNA-1273 vaccine induces neutralizing antibodies against spike mutants from global SARS-CoV-2 variants. bioRxiv 2021. preprint. [Google Scholar] [CrossRef]
- Mallapaty, S. WHO approval of Chinese CoronaVac COVID vaccine will be crucial to curbing pandemic. Nature 2021, 594, 161–162. [Google Scholar] [CrossRef]
- Ura, T.; Okuda, K.; Shimada, M. Developments in Viral Vector-Based Vaccines. Vaccines 2014, 2, 624–641. [Google Scholar] [CrossRef] [Green Version]
- Cele, S.; Gazy, I.; Jackson, L.; Hwa, S.-H.; Tegally, H.; Lustig, G.; Giandhari, J.; Pillay, S.; Wilkinson, E.; Naidoo, Y.; et al. Escape of SARS-CoV-2 501Y.V2 from neutralization by convalescent plasma. Nature 2021, 593, 142–146. [Google Scholar] [CrossRef]
- Zhao, H.; Lu, L.; Peng, Z.; Chen, L.-L.; Meng, X.; Zhang, C.; Ip, J.D.; Chan, W.-M.; Chu, A.W.-H.; Chan, K.-H.; et al. SARS-CoV-2 Omicron variant shows less efficient replication and fusion activity when compared with Delta variant in TMPRSS2-expressed cells. Emerg. Microbes. Infect. 2022, 11, 277–283. [Google Scholar] [CrossRef]
- Cele, S.; Jackson, L.; Khoury, D.S.; Khan, K.; Moyo-Gwete, T.; Tegally, H.; San, J.E.; Cromer, D.; Scheepers, C.; Amoako, D.; et al. SARS-CoV-2 Omicron has extensive but incomplete escape of Pfizer BNT162b2 elicited neutralization and requires ACE2 for infection. medRxiv 2021. preprint. [Google Scholar] [CrossRef]
- Abu-Raddad, L.J.; Chemaitelly, H.; Butt, A.A. Effectiveness of the BNT162b2 COVID-19 vaccine against the B.1.1.7 and B.1.351 variants. N. Engl. J. Med. 2021, 385, 187–189. [Google Scholar] [CrossRef]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J.; et al. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Li, S.; Jin, X.; Han, J.-B.; Xu, K.; Xu, S.; Han, Y.; Liu, C.; Zheng, T.; Liu, M.; et al. A tandem-repeat dimeric RBD protein-based COVID-19 vaccine zf2001 protects mice and nonhuman primates. Emerg. Microbes. Infect. 2022, 11, 1058–1071. [Google Scholar] [CrossRef]
- Precision Vaccinations. Zifivax (ZF2001) COVID-19 Vaccine. Available online: https://www.precisionvaccinations.com/vaccines/zifivax-zf2001-covid-19-vaccine (accessed on 13 April 2022).
- Dai, L.; Gao, L.; Tao, L.; Hadinegoro, S.R.; Erkin, M.; Ying, Z.; He, P.; Girsang, R.T.; Vergara, H.; Akram, J.; et al. Efficacy and Safety of the RBD-Dimer-Based COVID-19 Vaccine ZF2001 in Adults. N. Engl. J. Med. 2022, 386, 2097–2111. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, R.; Qiao, S.T.; Wang, X.; Zhang, W.; Ruan, W.; Dai, L.; Han, P.; Gao, G.F. Omicron SARS-CoV-2 Neutralization from Inactivated and ZF2001 Vaccines. N. Engl. J. Med. 2022, 387, 277–280. [Google Scholar] [CrossRef] [PubMed]
- National Health Commission of the People’s Republic of China. Questions and Answers about COVID-19 Vaccine Booster Immunization. Available online: http://www.nhc.gov.cn/xcs/yqfkdt/202111/67a59e40580d4b4687b3ed738333f6a9.shtml (accessed on 15 November 2021).
- Bright Uzbekistan. Uzbek Chinese Vaccine is Approved for Emergency Use in China. Available online: https://brightuzbekistan.uz/en/uzbek-chinese-vaccine-is-approved-for-emergency-use-in-china/ (accessed on 7 December 2022).
- Leng, S. Anhui Zhifei Longcom’s COVID-19 Vaccine Approved for Emergency Use in Colombia. Available online: https://www.globaltimes.cn/page/202201/1246703.shtml (accessed on 24 January 2022).
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.; Tiu, C.; Hu, Z.; Chen, V.C.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2-spike protein-protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Reimerink, J.; Torriani, G.; Brouwer, F.; Godeke, G.-J.; Yerly, S.; Hoogerwerf, M.; Vuilleumier, N.; Kaiser, L.; Eckerle, I.; et al. Validation and clinical evaluation of a SARS-CoV-2 surrogate virus neutralisation test (sVNT). Emerg. Microbes. Infect. 2020, 9, 2394–2403. [Google Scholar] [CrossRef] [PubMed]
- Dispinseri, S.; Secchi, M.; Pirillo, M.F.; Tolazzi, M.; Borghi, M.; Brigatti, C.; De Angelis, M.L.; Baratella, M.; Bazzigaluppi, E.; Venturi, G.; et al. Neutralizing antibody responses to SARS-CoV-2 in symptomatic COVID-19 is persistent and critical for survival. Nat. Commun. 2021, 12, 2670. [Google Scholar] [CrossRef]
- Sukumaran, A.; Thomas, R.E.; Krishnan, R.A.; Thomas, T.; Thomas, R.; Vijayan, D.K.; Paul, J.K.; Vasudevan, D.M. Sequential Profiling of Anti-SARS CoV-2 IgG Antibody in Post COVID-19 Patients. Indian J. Clin. Biochem. 2022, 37, 349–355. [Google Scholar] [CrossRef]
- Bagno, F.F.; Andrade, L.A.F.; Sergio, S.A.R.; Parise, P.L.; Toledo-Teixeira, D.A.; Gazzinelli, R.T.; Fernandes, A.P.S.M.; Teixeira, S.M.R.; Granja, F.; Proenca-Modena, J.L.; et al. Previous Infection with SARS-CoV-2 Correlates with Increased Protective Humoral Responses after a Single Dose of an Inactivated COVID-19 Vaccine. Viruses 2022, 14, 510. [Google Scholar] [CrossRef]
- Medeiros-Ribeiro, A.C.; Aikawa, N.E.; Saad, C.G.S.; Yuki, E.F.N.; Pedrosa, T.; Fusco, S.R.G.; Rojo, P.T.; Pereira, R.M.R.; Shinjo, S.K.; Andrade, D.C.O.; et al. Immunogenicity and safety of the CoronaVac inactivated vaccine in patients with autoimmune rheumatic diseases: A phase 4 trial. Nat. Med. 2021, 27, 1744–1751. [Google Scholar] [CrossRef]
- Clemens, S.A.C.; Weckx, L.; Clemens, R.; Mendes, A.V.A.; Souza, A.R.; Silveira, M.B.V.; de Guarda, S.N.F.; de Nobrega, M.M.; de Moraes Pinto, M.I.; Gonzalez, I.G.S.; et al. Heterologous versus homologous COVID-19 booster vaccination in previous recipients of two doses of CoronaVac COVID-19 vaccine in Brazil (RHH-001): A phase 4, non-inferiority, single blind, randomised study. Lancet 2022, 399, 521–529. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; Lam, E.C.; St. Denis, K.; Nitido, A.D.; Garcia, Z.H.; Hauser, B.M.; Feldman, J.; Pavlovic, M.N.; Gregory, D.J.; Poznansky, M.C.; et al. Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell 2021, 184, 2372–2383.e9. [Google Scholar] [CrossRef] [PubMed]
- WHO. Interim Recommendations for Use of the Inactivated COVID-19 Vaccine, CoronaVac, Developed by Sinovac. Available online: https://apps.who.int/iris/bitstream/handle/10665/341454/WHO-2019-nCoV-vaccines-SAGE-recommendation-Sinovac-CoronaVac-2021.1-eng.pdf (accessed on 15 March 2022).
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An mRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Corbett, K.S.; Flynn, B.; Foulds, K.E.; Francica, J.R.; Boyoglu-Barnum, S.; Werner, A.P.; Flach, B.; O’Connell, S.; Bock, K.W.; Minai, M.; et al. Evaluation of the mRNA-1273 Vaccine against SARS-CoV-2 in Nonhuman Primates. N. Engl. J. Med. 2020, 383, 1544–1555. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ginn, H.M.; Dejnirattisai, W.; Supasa, P.; Wang, B.; Tuekprakhon, A.; Nutalai, R.; Zhou, D.; Mentzer, A.J.; Zhao, Y.; et al. Reduced neutralization of SARS-CoV-2 B.1.617 by vaccine and convalescent serum. Cell 2021, 184, 4220–4236.e13. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kruger, N.; Schulz, S.; Cossmann, A.; Rocha, C.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Moldenhauer, A.-S.; Winkler, M.S.; et al. The Omicron variant is highly resistant against antibody-mediated neutralization: Implications for control of the COVID-19 pandemic. Cell 2022, 185, 447–456.e11. [Google Scholar] [CrossRef]
- Figueiredo-Campos, P.; Blankenhaus, B.; Mota, C.; Gomes, A.; Serrano, M.; Ariotti, S.; Costa, C.; Nunes-Cabaco, H.; Mendes, A.M.; Gaspar, P.; et al. Seroprevalence of anti-SARS-CoV-2 antibodies in COVID-19 patients and healthy volunteers up to 6 months post disease onset. Eur. J. Immunol. 2020, 50, 2025–2040. [Google Scholar] [CrossRef]
- Collier, D.A.; De Marco, A.; Ferreira, I.A.T.M.; Meng, B.; Datir, R.P.; Walls, A.C.; Kemp, S.A.; Bassi, J.; Pinto, D.; Silacci-Fregni, C.; et al. Sensitivity of SARS-CoV-2 B.1.1.7 to mRNA vaccine-elicited antibodies. Nature 2021, 593, 136–141. [Google Scholar] [PubMed]
- Cao, Y.; Hao, X.; Wang, X.; Wu, Q.; Song, R.; Zhao, D.; Song, W.; Wang, Y.; Yisimayi, A.; Wang, W.; et al. Humoral immunogenicity and reactogenicity of CoronaVac or ZF2001 booster after two doses of inactivated vaccine. Cell Res. 2022, 32, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Buathong, R.; Hunsawong, T.; Wacharapluesadee, S.; Guharat, S.; Jirapipatt, R.; Ninwattana, S.; Thippamom, N.; Jitsatja, A.; Jones, A.R.; Rungrojchareonkit, K.; et al. Homologous or Heterologous COVID-19 Booster Regimens Significantly Impact Sero-Neutralization of SARS-CoV-2 Virus and Its Variants. Vaccines 2022, 10, 1321. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.; Zhang, H.; Zhang, Q.; Zhang, Y.; Lin, K.; Fu, Z.; Song, J.; Zhao, Y.; Fan, M.; Wang, H.; et al. Recombinant protein subunit vaccine booster following two-dose inactivated vaccines dramatically enhanced anti-RBD responses and neutralizing titers against SARS-CoV-2 and Variants of Concern. Cell Res. 2022, 32, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Ju, B.; Zhang, Q.; Ge, J.; Wang, R.; Sun, J.; Ge, X.; Yu, J.; Shan, S.; Zhou, B.; Song, S.; et al. Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 2020, 584, 115–119. [Google Scholar] [CrossRef]
- Yang, R.; Huang, B.; Ruhan, A.; Li, W.; Wang, W.; Deng, Y.; Tan, W. Development and effectiveness of pseudotyped SARS-CoV-2 system as determined by neutralizing efficiency and entry inhibition test in vitro. Biosaf. Health 2020, 2, 226–231. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccination Strategy | The First Dose | The Second Dose | Booster | |||
|---|---|---|---|---|---|---|
| 3rd Week | 1st Month | 3rd Month | 5th Month | 1st Month | 2nd Month | |
| Inactivated Vaccine | n = 68 | n = 201 | n = 127 | n = 68 | n = 92 | n = 61 |
| ZF2001 | n = 50 | |||||
| Date | Inhibition (%) | |||||
|---|---|---|---|---|---|---|
| Wild-Type | B.1.1.7 | B.1.351 | P.1 | B.1.617.2 | B.1.1.529 | |
| 1–3rd week | 31.81 | 30.56 | 22.18 | 17.14 | 28.67 | 24.81 |
| 2–1st month | 82.17 | 75.00 | 65.04 | 69.99 | 58.97 | 54.79 |
| 2–3rd month | 61.76 | 51.03 | 41.21 | 45.59 | 40.61 | 35.61 |
| 2–5th month | 46.25 | 36.28 | 32.23 | 35.48 | 25.64 | 24.58 |
| Vaccination Strategy | Inhibition (%) ± SD | |||
|---|---|---|---|---|
| Wild-Type | B.1.351 | B.1.617.2 | B.1.1.529 | |
| Inactivated Booster—1 month | 88.30 ± 9.05 | 79.01 ± 12.32 | 76.34 ± 12.07 | 72.07 ± 12.88 |
| ZF2001 Booster—1 month | 92.28 ± 4.62 | 85.31 ± 8.15 | 82.10 ± 7.36 | 81.27 ± 9.30 |
| Inactivated Booster—2 month | 80.74 ± 10.35 | 73.24 ± 15.71 | 69.12 ± 12.61 | 71.54 ± 12.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Hong, Y.; Chen, E.; Wang, Y.; Ma, B.; Li, J.; Su, W.; Zhou, Y.; Zhang, M. Antibodies Induced by Homologous or Heterologous Inactivated (CoronaVac/BBIBP-CorV) and Recombinant Protein Subunit Vaccines (ZF2001) Dramatically Enhanced Inhibitory Abilities against B.1.351, B.1.617.2, and B.1.1.529 Variants. Vaccines 2022, 10, 2110. https://doi.org/10.3390/vaccines10122110
Xu X, Hong Y, Chen E, Wang Y, Ma B, Li J, Su W, Zhou Y, Zhang M. Antibodies Induced by Homologous or Heterologous Inactivated (CoronaVac/BBIBP-CorV) and Recombinant Protein Subunit Vaccines (ZF2001) Dramatically Enhanced Inhibitory Abilities against B.1.351, B.1.617.2, and B.1.1.529 Variants. Vaccines. 2022; 10(12):2110. https://doi.org/10.3390/vaccines10122110
Chicago/Turabian StyleXu, Xuesong, Yi Hong, Erjing Chen, Yaping Wang, Biao Ma, Jiali Li, Wei Su, Yuxin Zhou, and Mingzhou Zhang. 2022. "Antibodies Induced by Homologous or Heterologous Inactivated (CoronaVac/BBIBP-CorV) and Recombinant Protein Subunit Vaccines (ZF2001) Dramatically Enhanced Inhibitory Abilities against B.1.351, B.1.617.2, and B.1.1.529 Variants" Vaccines 10, no. 12: 2110. https://doi.org/10.3390/vaccines10122110