
Allelic Variants of HLA-C Upstream Region, PSORS1C3, MICA, TNFA and Genes Involved in Epidermal Homeostasis and Barrier Function Influence the Clinical Response to Anti-IL-12/IL-23 Treatment of Patients with Psoriasis
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Ethics Statement
2.2. SNP Analysis
2.3. Statistical Analysis
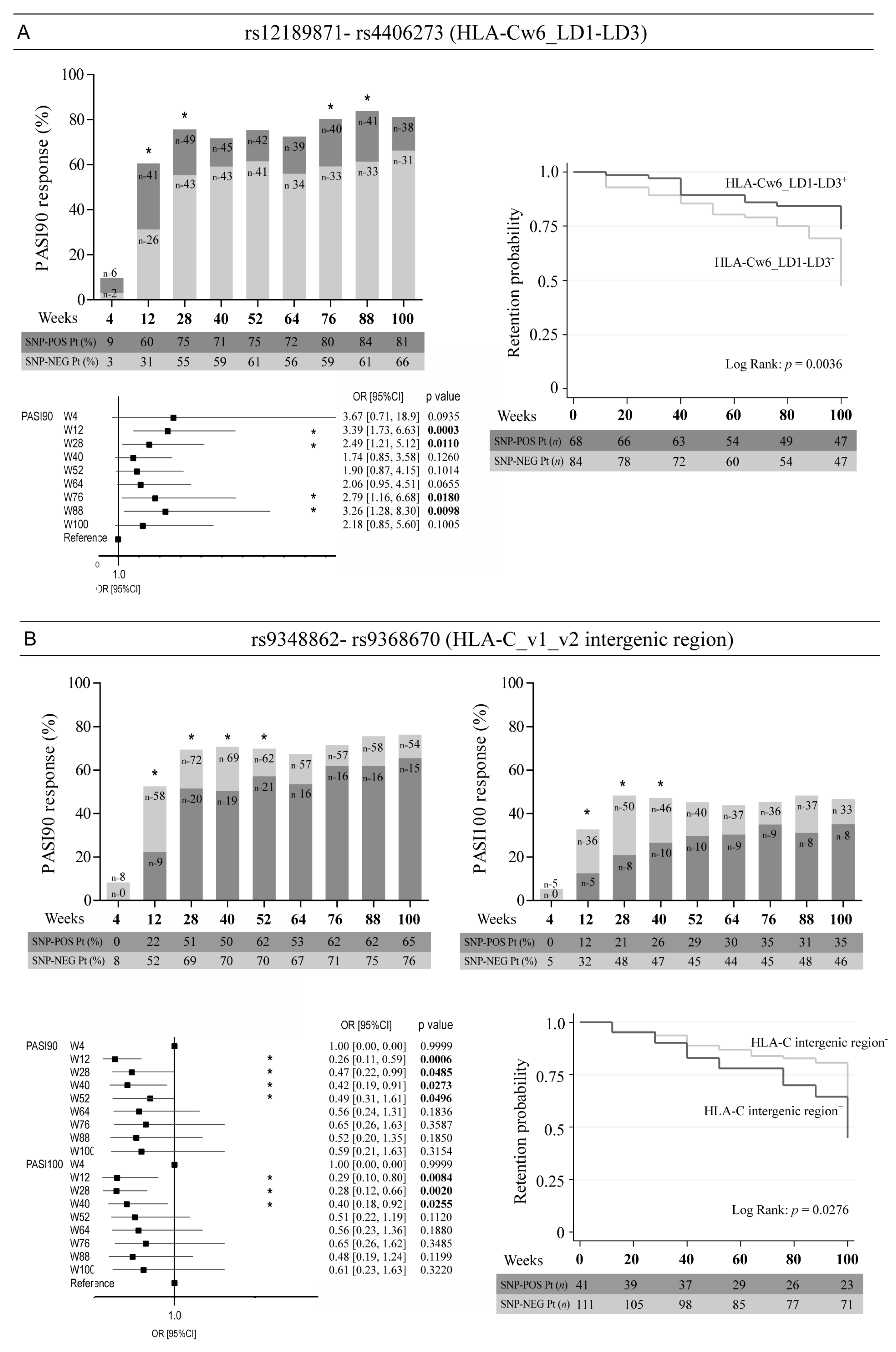
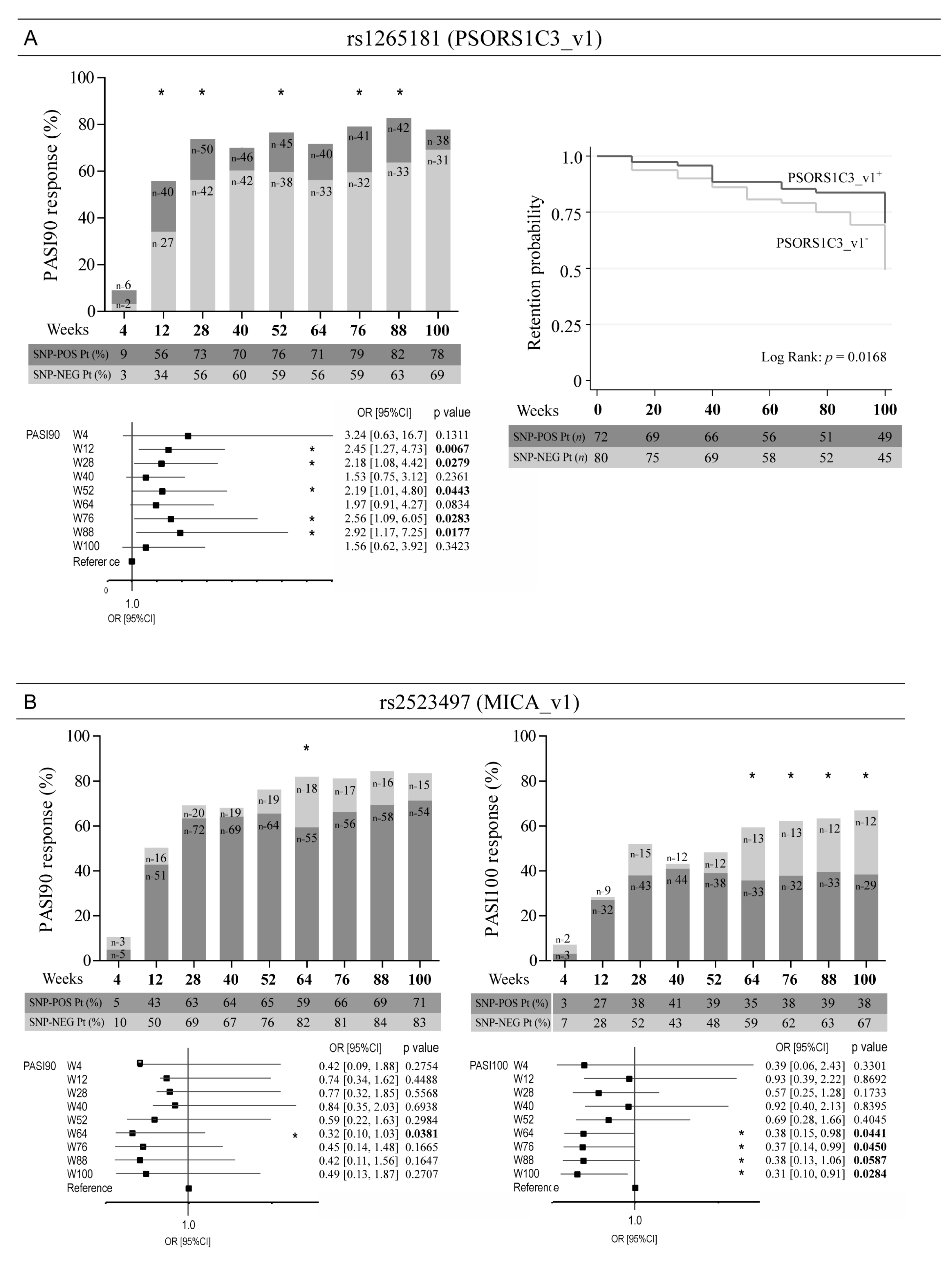
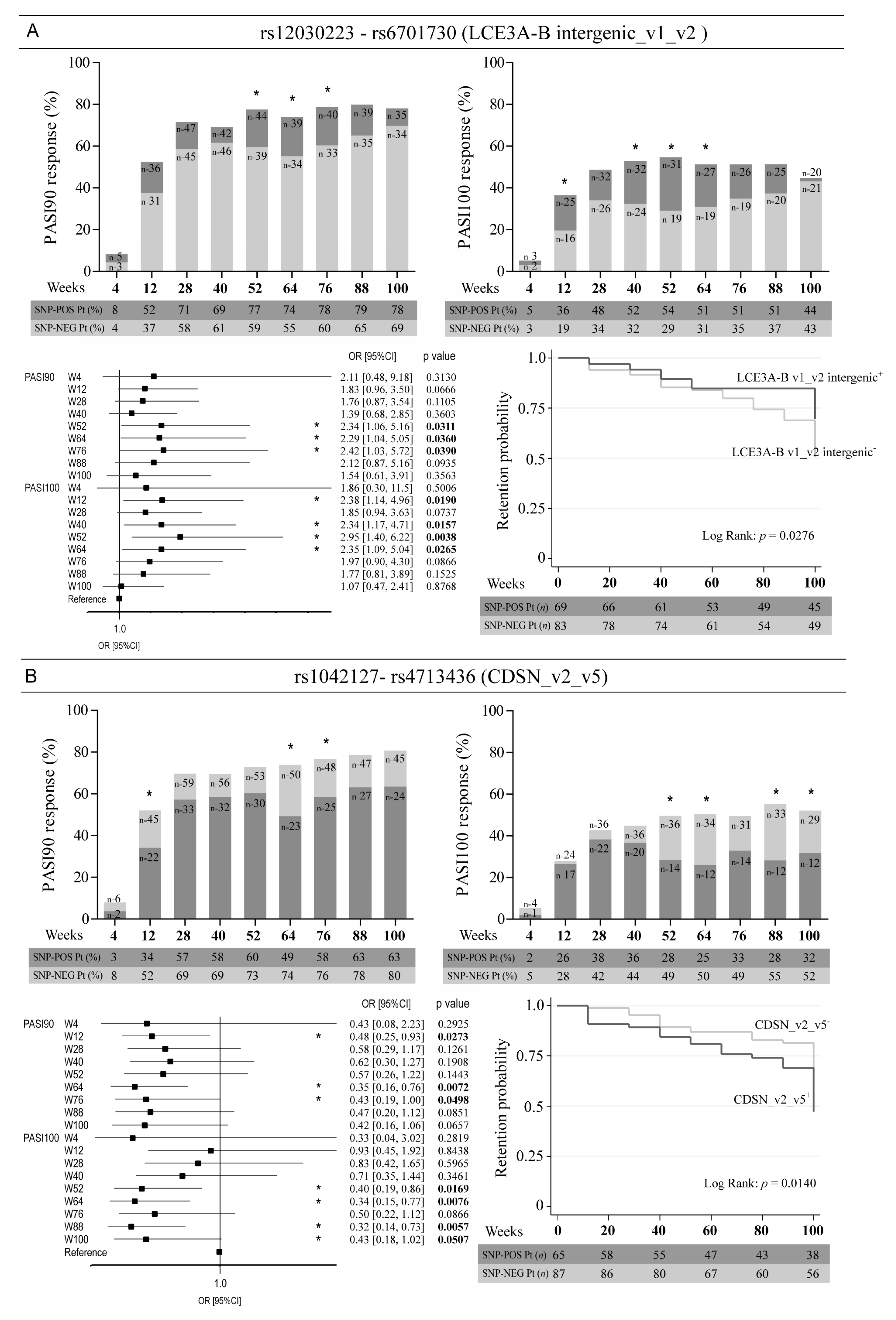
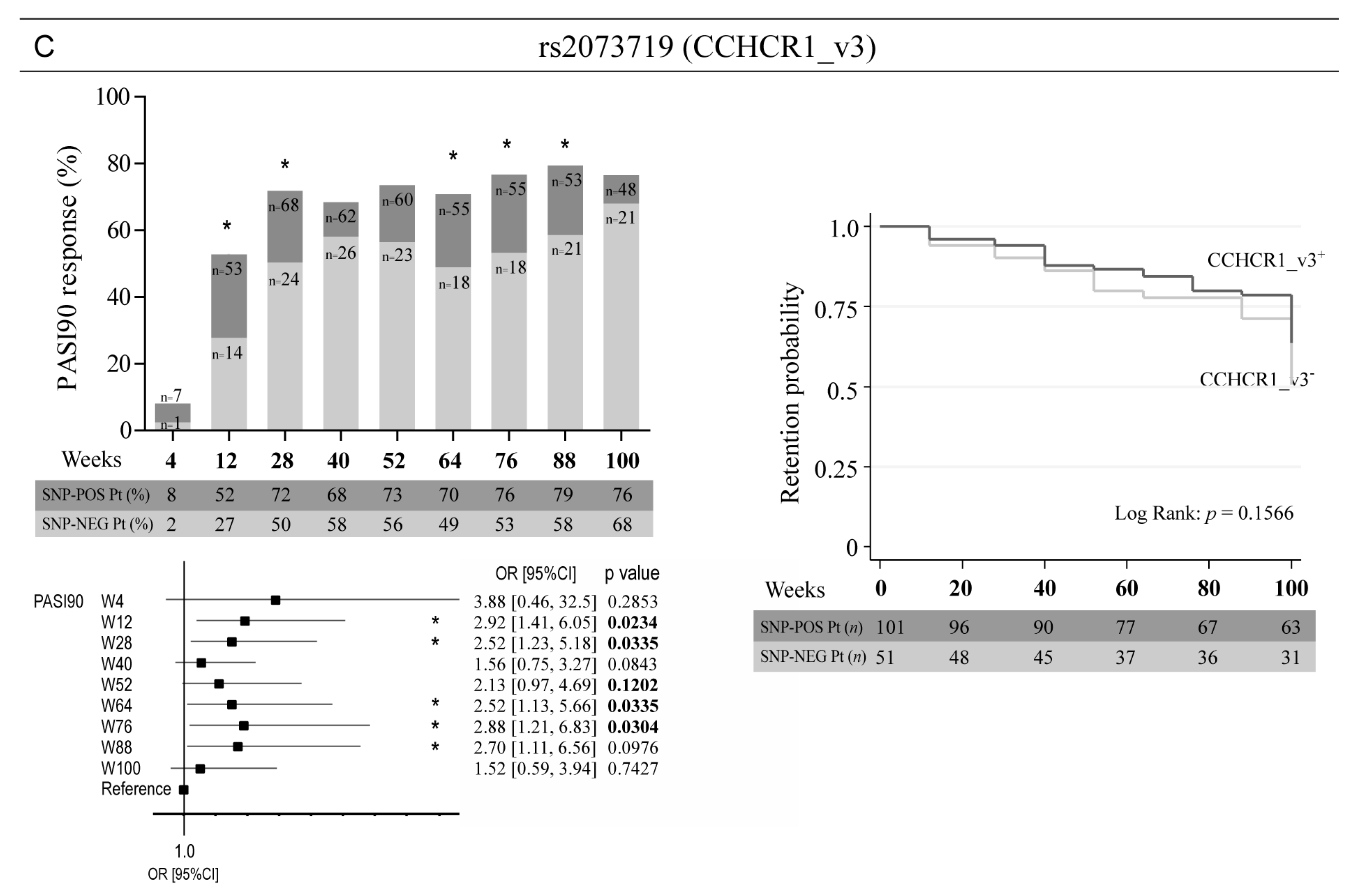
3. Results
3.1. Patients Responding to Ustekinumab Therapy Show Specific SNP Patterns Coherently with HLA-Cw6 Status and BMI Values < 30
3.2. Optimal Response to Ustekinumab Associates with the Allelic Variant Status of SNPs in the Intergenic Region Upstream of HLA-C, as Well as in PSORS1C3 and MICA Genes
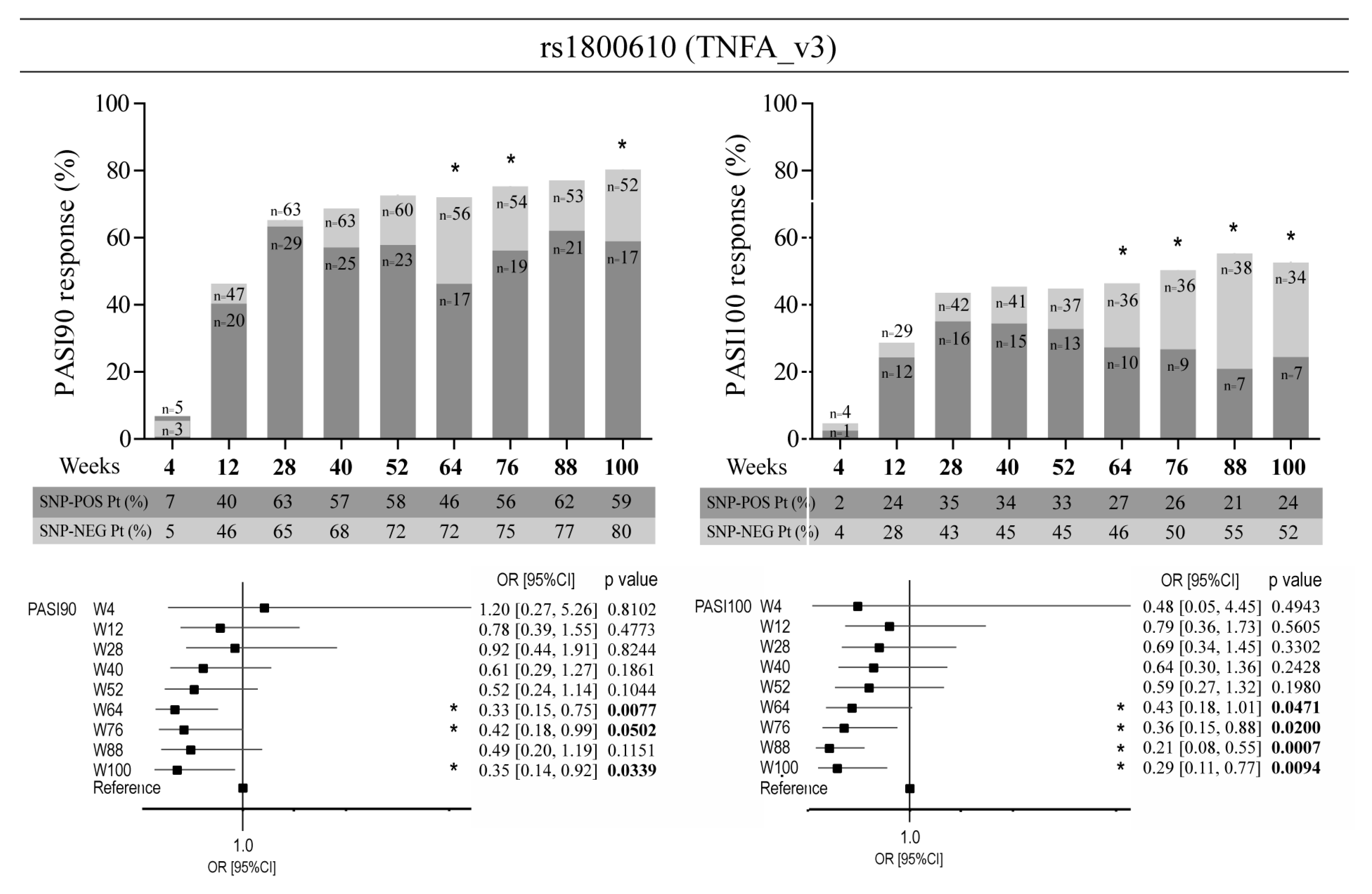
3.3. Allelic Variants of Genes Involved in Epidermal Homeostasis and Barrier Function and TNFA Affect the Clinical Response to Ustekinumab
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Albanesi, C.; Madonna, S.; Gisondi, P.; Girolomoni, G. The Interplay Between Keratinocytes and Immune Cells in the Pathogenesis of Psoriasis. Front. Immunol. 2018, 9, 1549. [Google Scholar] [CrossRef] [Green Version]
- Christophers, E.; Metzler, G.; Röcken, M. Bimodal immune activation in psoriasis. Br. J. Dermatol. 2014, 170, 59–65. [Google Scholar] [CrossRef]
- Dand, N.; Mahil, S.K.; Capon, F.; Smith, C.H.; Simpson, M.A.; Barker, J.N. Psoriasis and Genetics. Acta Derm. Venereol. 2020, 100, adv00030. [Google Scholar] [CrossRef] [Green Version]
- Capon, F. The Genetic Basis of Psoriasis. Int. J. Mol. Sci. 2017, 18, 2526. [Google Scholar] [CrossRef] [Green Version]
- Dand, N.; Mucha, S.; Tsoi, L.C.; Mahil, S.K.; Stuart, P.E.; Arnold, A.; Baurecht, H.; Burden, A.D.; Callis Duffin, K.; Chandran, V.; et al. Exome-wide association study reveals novel psoriasis susceptibility locus at TNFSF15 and rare protective alleles in genes contributing to type I IFN signalling. Hum. Mol. Genet. 2017, 26, 4301–4313. [Google Scholar] [CrossRef] [Green Version]
- Tsoi, L.C.; Stuart, P.E.; Tian, C.; Gudjonsson, J.E.; Das, S.; Zawistowski, M.; Ellinghaus, E.; Barker, J.N.; Chandran, V.; Dand, N.; et al. Large scale meta-analysis characterizes genetic architecture for common psoriasis associated variants. Nat. Commun. 2017, 8, 15382. [Google Scholar] [CrossRef] [Green Version]
- Patrick, M.T.; Stuart, P.E.; Raja, K.; Gudjonsson, J.E.; Tejasvi, T.; Yang, J.; Chandran, V.; Das, S.; Callis-Duffin, K.; Ellinghaus, E.; et al. Genetic signature to provide robust risk assessment of psoriatic arthritis development in psoriasis patients. Nat. Commun. 2018, 9, 4178. [Google Scholar] [CrossRef] [Green Version]
- Nair, R.P.; Duffin, K.C.; Helms, C.; Ding, J.; Stuart, P.E.; Goldgar, D.; Gudjonsson, J.E.; Li, Y.; Tejasvi, T.; Feng, B.J.; et al. Genome-wide scan reveals association of psoriasis with IL-23 and NF-kappaB pathways. Nat. Genet. 2009, 41, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Strange, A.; Capon, F.; Spencer, C.C.; Knight, J.; Weale, M.E.; Allen, M.H.; Barton, A.; Band, G.; Bellenguez, C.; Bergboer, J.G.; et al. A genome-wide association study identifies new psoriasis susceptibility loci and an interaction between HLA-C and ERAP1. Nat. Genet. 2010, 42, 985–990. [Google Scholar] [CrossRef]
- De Cid, R.; Riveira-Munoz, E.; Zeeuwen, P.L.; Robarge, J.; Liao, W.; Dannhauser, E.N.; Giardina, E.; Stuart, P.E.; Nair, R.; Helms, C.; et al. Deletion of the late cornified envelope LCE3B and LCE3C genes as a susceptibility factor for psoriasis. Nat. Genet. 2009, 41, 211–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellinghaus, E.; Ellinghaus, D.; Stuart, P.E.; Nair, R.P.; Debrus, S.; Raelson, J.V.; Belouchi, M.; Fournier, H.; Reinhard, C.; Ding, J.; et al. Genome-wide association study identifies a psoriasis susceptibility locus at TRAF3IP2. Nat. Genet. 2010, 42, 991–995. [Google Scholar] [CrossRef] [PubMed]
- Cargill, M.; Schrodi, S.J.; Chang, M.; Garcia, V.E.; Brandon, R.; Callis, K.P.; Matsunami, N.; Ardlie, K.G.; Civello, D.; Catanese, J.J.; et al. A large-scale genetic association study confirms IL12B and leads to the identification of IL23R as psoriasis-risk genes. Am. J. Hum. Genet. 2007, 80, 273–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinle, A.; Li, P.; Morris, D.L.; Groh, V.; Lanier, L.L.; Strong, R.K.; Spies, T. Interactions of human NKG2D with its ligands MICA, MICB, and homologs of the mouse RAE-1 protein family. Immunogenetics 2001, 53, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Lande, R.; Botti, E.; Jandus, C.; Dojcinovic, D.; Fanelli, G.; Conrad, C.; Chamilos, G.; Feldmeyer, L.; Marinari, B.; Chon, S.; et al. The antimicrobial peptide LL37 is a T-cell autoantigen in psoriasis. Nat. Commun. 2014, 5, 5621. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-Duculan, J.; Bonifacio, K.M.; Hawkes, J.E.; Kunjravia, N.; Cueto, I.; Li, X.; Gonzalez, J.; Garcet, S.; Krueger, J.G. Autoantigens ADAMTSL5 and LL37 are significantly upregulated in active Psoriasis and localized with keratinocytes, dendritic cells and other leukocytes. Exp. Dermatol. 2017, 26, 1075–1082. [Google Scholar] [CrossRef]
- Raposo, I.; Carvalho, C.; Bettencourt, A.; Da Silva, B.M.; Leite, L.; Selores, M.; Torres, T. Psoriasis pharmacogenetics: HLA-Cw*0602 as a marker of therapeutic response to ustekinumab. Eur. J. Dermatol. 2017, 27, 528–530. [Google Scholar] [CrossRef]
- Dand, N.; Duckworth, M.; Baudry, D.; Russell, A.; Curtis, C.J.; Lee, S.H.; Evans, I.; Mason, K.J.; Alsharqi, A.; Becher, G.; et al. HLA-C*06:02 genotype is a predictive biomarker of biologic treatment response in psoriasis. J. Allergy Clin. Immunol. 2019, 143, 2120–2130. [Google Scholar] [CrossRef] [Green Version]
- Morelli, M.; Galluzzo, M.; Madonna, S.; Scarponi, C.; Scaglione, G.L.; Galluccio, T.; Andreani, M.; Pallotta, S.; Girolomoni, G.; Bianchi, L.; et al. and other HLA-C alleles, as well as. Expert Opin. Biol. Ther. 2021, 21, 259–270. [Google Scholar] [CrossRef]
- Langley, R.G.; Lebwohl, M.; Krueger, G.G.; Szapary, P.O.; Wasfi, Y.; Chan, D.; Hsu, M.C.; You, Y.; Poulin, Y.; Korman, N.; et al. Long-term efficacy and safety of ustekinumab, with and without dosing adjustment, in patients with moderate-to-severe psoriasis: Results from the PHOENIX 2 study through 5 years of follow-up. Br. J. Dermatol. 2015, 172, 1371–1383. [Google Scholar] [CrossRef]
- Galluzzo, M.; D’Adamio, S.; Silvaggio, D.; Lombardo, P.; Massaro, A.; Egan, C.G.; Bianchi, L.; Talamonti, M. Ustekinumab treatment for moderate-to-severe plaque psoriasis: Eight-year real-life experience. Expert Opin. Biol. Ther. 2020, 20, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Stuart, P.E.; Nair, R.P.; Ellinghaus, E.; Ding, J.; Tejasvi, T.; Gudjonsson, J.E.; Li, Y.; Weidinger, S.; Eberlein, B.; Gieger, C.; et al. Genome-wide association analysis identifies three psoriasis susceptibility loci. Nat. Genet. 2010, 42, 1000–1004. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, L.C.; Spain, S.L.; Knight, J.; Ellinghaus, E.; Stuart, P.E.; Capon, F.; Ding, J.; Li, Y.; Tejasvi, T.; Gudjonsson, J.E.; et al. Identification of 15 new psoriasis susceptibility loci highlights the role of innate immunity. Nat. Genet. 2012, 44, 1341–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoi, L.C.; Spain, S.L.; Ellinghaus, E.; Stuart, P.E.; Capon, F.; Knight, J.; Tejasvi, T.; Kang, H.M.; Allen, M.H.; Lambert, S.; et al. Enhanced meta-analysis and replication studies identify five new psoriasis susceptibility loci. Nat. Commun. 2015, 6, 7001. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.J.; Willis, T.J. Canonical analysis of principal coordinates: A useful method of constrained ordination for ecology. Ecology 2003, 84, 511. [Google Scholar] [CrossRef]
- Anderson, M.J.; Robinson, J. Generalised discriminant analysis based on distances. Aust. New Zealand J. Stat. 2003, 45, 301. [Google Scholar] [CrossRef]
- Talamonti, M.; Botti, E.; Galluzzo, M.; Teoli, M.; Spallone, G.; Bavetta, M.; Chimenti, S.; Costanzo, A. Pharmacogenetics of psoriasis: HLA-Cw6 but not LCE3B/3C deletion nor TNFAIP3 polymorphism predisposes to clinical response to interleukin 12/23 blocker ustekinumab. Br. J. Dermatol. 2013, 169, 458–463. [Google Scholar] [CrossRef]
- Chiu, H.Y.; Wang, T.S.; Chan, C.C.; Cheng, Y.P.; Lin, S.J.; Tsai, T.F. Human leucocyte antigen-Cw6 as a predictor for clinical response to ustekinumab, an interleukin-12/23 blocker, in Chinese patients with psoriasis: A retrospective analysis. Br. J. Dermatol. 2014, 171, 1181–1188. [Google Scholar] [CrossRef]
- Prieto-Pérez, R.; Solano-López, G.; Cabaleiro, T.; Román, M.; Ochoa, D.; Talegón, M.; Baniandrés, O.; López-Estebaranz, J.L.; de la Cueva, P.; Daudén, E.; et al. Polymorphisms Associated with Age at Onset in Patients with Moderate-to-Severe Plaque Psoriasis. J. Immunol. Res. 2015, 2015, 101879. [Google Scholar] [CrossRef] [Green Version]
- González-Lara, L.; Batalla, A.; Coto, E.; Gómez, J.; Eiris, N.; Santos-Juanes, J.; Queiro, R.; Coto-Segura, P. The TNFRSF1B rs1061622 polymorphism (p.M196R) is associated with biological drug outcome in Psoriasis patients. Arch. Dermatol. Res. 2015, 307, 405–412. [Google Scholar] [CrossRef]
- Galluzzo, M.; Boca, A.N.; Botti, E.; Potenza, C.; Malara, G.; Malagoli, P.; Vesa, S.; Chimenti, S.; Buzoianu, A.D.; Talamonti, M.; et al. IL12B (p40) Gene Polymorphisms Contribute to Ustekinumab Response Prediction in Psoriasis. Dermatology 2016, 232, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Loft, N.D.; Skov, L.; Iversen, L.; Gniadecki, R.; Dam, T.N.; Brandslund, I.; Hoffmann, H.J.; Andersen, M.R.; Dessau, R.B.; Bergmann, A.C.; et al. Associations between functional polymorphisms and response to biological treatment in Danish patients with psoriasis. Pharm. J. 2018, 18, 494–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto-Pérez, R.; Llamas-Velasco, M.; Cabaleiro, T.; Solano-López, G.; Márquez, B.; Román, M.; Ochoa, D.; Talegón, M.; Daudén, E.; Abad-Santos, F. Pharmacogenetics of ustekinumab in patients with moderate-to-severe plaque psoriasis. Pharmacogenomics 2017, 18, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Masouri, S.; Stefanaki, I.; Ntritsos, G.; Kypreou, K.P.; Drakaki, E.; Evangelou, E.; Nicolaidou, E.; Stratigos, A.J.; Antoniou, C. A Pharmacogenetic Study of Psoriasis Risk Variants in a Greek Population and Prediction of Responses to Anti-TNF-α and Anti-IL-12/23 Agents. Mol. Diagn. Ther. 2016, 20, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Connell, W.T.; Hong, J.; Liao, W. Genome-Wide Association Study of Ustekinumab Response in Psoriasis. Front. Immunol. 2022, 12, 815121. [Google Scholar] [CrossRef]
- Holm, S.J.; Sánchez, F.; Carlén, L.M.; Mallbris, L.; Ståhle, M.; O’Brien, K.P. HLA-Cw*0602 associates more strongly to psoriasis in the Swedish population than variants of the novel 6p21.3 gene PSORS1C3. Acta Derm. Venereol. 2005, 85, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Mirzadeh Azad, F.; Malakootian, M.; Mowla, S.J. lncRNA PSORS1C3 is regulated by glucocorticoids and fine-tunes OCT4 expression in non-pluripotent cells. Sci. Rep. 2019, 9, 8370. [Google Scholar] [CrossRef] [Green Version]
- Ovejero-Benito, M.C.; Prieto-Pérez, R.; Llamas-Velasco, M.; Belmonte, C.; Cabaleiro, T.; Román, M.; Ochoa, D.; Talegón, M.; Saiz-Rodríguez, M.; Daudén, E.; et al. Polymorphisms associated with etanercept response in moderate-to-severe plaque psoriasis. Pharmacogenomics 2017, 18, 631–638. [Google Scholar] [CrossRef]
- Qidwai, T.; Khan, F. Tumour necrosis factor gene polymorphism and disease prevalence. Scand. J. Immunol. 2011, 74, 522–547. [Google Scholar] [CrossRef]
- Talaat, R.M.; Abdelkhalek, M.S.; El-Maadawy, E.A.; Abdel-Mageed, W.S.; El-Shenawy, S.Z.; Osman, M.A. Association of TNF-Alpha gene polymorphisms and susceptibility to hepatitis B virus infection in Egyptians. Hum. Immunol. 2017, 78, 739–746. [Google Scholar] [CrossRef]
- Niehues, H.; Tsoi, L.C.; van der Krieken, D.A.; Jansen, P.A.M.; Oortveld, M.A.W.; Rodijk-Olthuis, D.; van Vlijmen, I.M.J.J.; Hendriks, W.J.A.J.; Helder, R.W.; Bouwstra, J.A.; et al. Psoriasis-Associated Late Cornified Envelope (LCE) Proteins Have Antibacterial Activity. J. Invest. Dermatol. 2017, 137, 2380–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Guzman Strong, C.; Conlan, S.; Deming, C.B.; Cheng, J.; Sears, K.E.; Segre, J.A. A milieu of regulatory elements in the epidermal differentiation complex syntenic block: Implications for atopic dermatitis and psoriasis. Hum. Mol. Genet. 2010, 19, 1453–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, N.K.; Dilolli, M.N.; Miller, L.S. Pushing the Envelope in Psoriasis: Late Cornified Envelope Proteins Possess Antimicrobial Activity. J. Invest. Dermatol. 2017, 137, 2257–2259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiśniewski, A.; Matusiak, Ł.; Szczerkowska-Dobosz, A.; Nowak, I.; Kuśnierczyk, P. HLA-C*06:02-independent, gender-related association of PSORS1C3 and PSORS1C1/CDSN single-nucleotide polymorphisms with risk and severity of psoriasis. Mol. Genet. Genom. 2018, 293, 957–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asumalahti, K.; Veal, C.; Laitinen, T.; Suomela, S.; Allen, M.; Elomaa, O.; Moser, M.; de Cid, R.; Ripatti, S.; Vorechovsky, I.; et al. Coding haplotype analysis supports HCR as the putative susceptibility gene for psoriasis at the MHC PSORS1 locus. Hum. Mol. Genet. 2002, 11, 589–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tervaniemi, M.H.; Katayama, S.; Skoog, T.; Siitonen, H.A.; Vuola, J.; Nuutila, K.; Tammimies, K.; Suomela, S.; Kankuri, E.; Kere, J.; et al. Intracellular signalling pathways and cytoskeletal functions converge on the psoriasis candidate gene CCHCR1 expressed at P-bodies and centrosomes. BMC Genom. 2018, 19, 432. [Google Scholar] [CrossRef]
- Nast, A.; Jacobs, A.; Rosumeck, S.; Werner, R.N. Efficacy and Safety of Systemic Long-Term Treatments for Moderate-to-Severe Psoriasis: A Systematic Review and Meta-Analysis. J. Invest. Dermatol. 2015, 135, 2641–2648. [Google Scholar] [CrossRef] [Green Version]
- Corbett, M.; Ramessur, R.; Marshall, D.; Acencio, M.L.; Ostaszewski, M.; Barbosa, I.A.; Dand, N.; Di Meglio, P.; Haddad, S.; Jensen, A.H.M.; et al. Biomarkers of systemic treatment response in people with psoriasis: A scoping review. Br. J. Dermatol. 2022, 187, 494–506. [Google Scholar] [CrossRef]
- Van Vugt, L.J.; van den Reek, J.M.P.A.; Hannink, G.; Coenen, M.J.H.; de Jong, E.M.G.J. Association of HLA-C*06:02 Status with Differential Response to Ustekinumab in Patients with Psoriasis: A Systematic Review and Meta-analysis. JAMA Dermatol. 2019, 155, 708–715. [Google Scholar] [CrossRef] [Green Version]
- Do, L.H.D.; Maibach, H. Pharmacogenomics/updated for precision medicine in dermatology. J. Dermatolog. Treat. 2019, 30, 410–413. [Google Scholar] [CrossRef]
- Derbyshire, M. Patent expiry dates for biologicals: 2018 update. GaBI J. 2019, 8, 24–31. [Google Scholar]
- Barker, J.; Girolomoni, G.; Egeberg, A.; Goncalves, J.; Pieper, B.; Kang, T. Anti-TNF biosimilars in psoriasis: From scientific evidence to real-world experience. J. Dermatol. Treat. 2020, 31, 794–800. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | |
|---|---|
| Male/female (n) | 98/54 |
| Mean ± SD (range) | |
| Age (years) | 50.6 ± 14.4 (18–85) |
| PASI at baseline | 19.1 ± 10.8 (5–62) |
| PASI at follow-up (n) | |
| Week 4 (137) | 10.2 ± 7.83 (0–45) |
| Week 12 (152) | 5.23 ± 6.97 (0–40) |
| Week 28 (143) | 3.35 ± 5.59 (0–40) |
| Week 40 (136) | 3.01 ± 5.23 (0–40) |
| Week 52 (123) | 2.39 ± 3.71 (0–18) |
| Week 64 (115) | 2.56 ± 3.89 (0–20) |
| Week 76 (106) | 2.31 ± 3.91 (0–25) |
| Week 88 (103) | 2.18 ± 3.54 (0–18) |
| Week 100 (94) | 1.93 ± 3.27 (0–18) |
| Duration of disease (years) | 21.1 ± 13.4 (1–67) |
| Age at disease onset (years) | 27.1 ± 13.7 (7–73) |
| Weight (kg) | 81.2 ± 20.9 (23–168) |
| BMI (kg/m2) | 29.1 ± 17.8 (15.2–40.4) |
| Biologics before anti-IL-12/IL-23 therapy | n (%) |
| 0 prior biologics | 85 (55.2) |
| 1 prior biologics | 38 (24.7) |
| 2 prior biologics | 22 (14.3) |
| ≥3 prior biologics | 9 (5.8) |
| Comorbidities | 98 |
| Hypertension | 42 |
| Type 2 diabetes mellitus | 7 |
| Hyperlipidemia | 19 |
| Depression | 3 |
| Obesity (BMI ≥ 30.0–34.9 kg/m2) | 27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morelli, M.; Galluzzo, M.; Scarponi, C.; Madonna, S.; Scaglione, G.L.; Girolomoni, G.; Talamonti, M.; Bianchi, L.; Albanesi, C. Allelic Variants of HLA-C Upstream Region, PSORS1C3, MICA, TNFA and Genes Involved in Epidermal Homeostasis and Barrier Function Influence the Clinical Response to Anti-IL-12/IL-23 Treatment of Patients with Psoriasis. Vaccines 2022, 10, 1977. https://doi.org/10.3390/vaccines10111977
Morelli M, Galluzzo M, Scarponi C, Madonna S, Scaglione GL, Girolomoni G, Talamonti M, Bianchi L, Albanesi C. Allelic Variants of HLA-C Upstream Region, PSORS1C3, MICA, TNFA and Genes Involved in Epidermal Homeostasis and Barrier Function Influence the Clinical Response to Anti-IL-12/IL-23 Treatment of Patients with Psoriasis. Vaccines. 2022; 10(11):1977. https://doi.org/10.3390/vaccines10111977
Chicago/Turabian StyleMorelli, Martina, Marco Galluzzo, Claudia Scarponi, Stefania Madonna, Giovanni Luca Scaglione, Giampiero Girolomoni, Marina Talamonti, Luca Bianchi, and Cristina Albanesi. 2022. "Allelic Variants of HLA-C Upstream Region, PSORS1C3, MICA, TNFA and Genes Involved in Epidermal Homeostasis and Barrier Function Influence the Clinical Response to Anti-IL-12/IL-23 Treatment of Patients with Psoriasis" Vaccines 10, no. 11: 1977. https://doi.org/10.3390/vaccines10111977