1. Introduction
Squalene (SQ) is a lipophilic biomolecule that belongs to the chemical class of triterpenes. It is formed by six isoprene units, so, it is a 30-carbon isoprenoid compound with six double bonds (C
30H
50). SQ is an odorless, colorless, liquid oil. The molecule was named squalene because it was initially isolated by Tsujimoto from shark (
Squalus milsukurii and other squaloids) liver oil [
1,
2,
3], but subsequent studies have also found this compound in several plant extracts [
4].
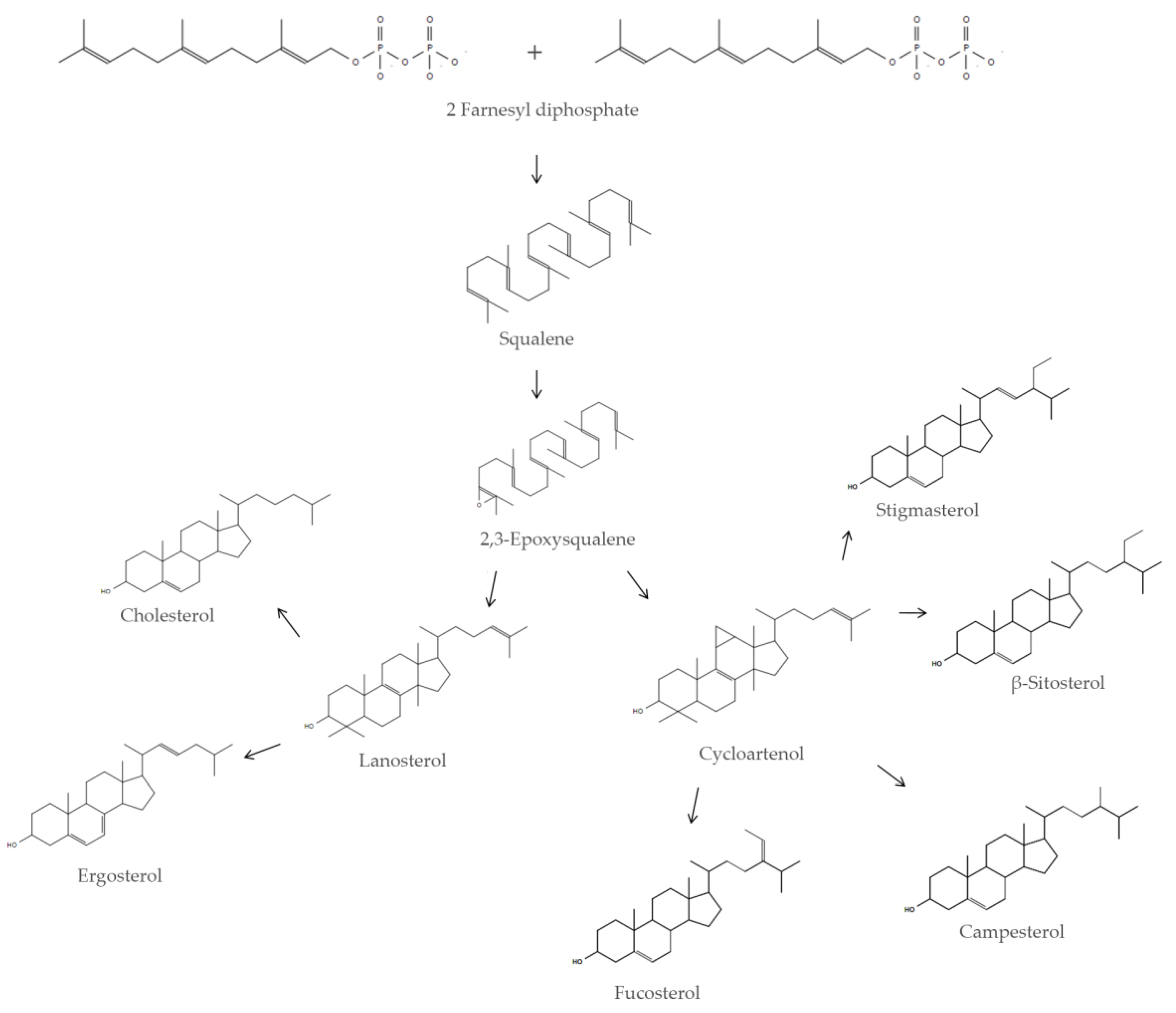
In eukaryotic cells SQ is a metabolic intermediate in the sterol biosynthetic pathway [
5]; squalene synthase, an enzyme associated with the membrane of the endoplasmic reticulum, catalyzes the biosynthesis of SQ in a two-step reaction starting from two units of farnesyl diphosphate [
6]. SQ is the precursor of cholesterol in animal cells and the precursor of phytosterols in plant cells (
Figure 1).
As an intermediate in sterol biosynthesis, SQ plays a crucial role in plant and animal organisms [
1,
7]. Several in vitro and in vivo studies have already underlined the antioxidant effects of SQ, probably due to its molecular properties directly linked to its activity towards free radicals [
1,
2,
8]. In fact, due to its conformation, it can easily move through cellular and subcellular membranes, allowing rapid distribution in every cell compartment [
9]. In addition, it acts as scavenger and quencher against some free radicals. In particular, SQ breaks lipid peroxidation chains both in the initial phase and in the propagation phase [
8].
However, SQ’s role in the prevention or treatment of oxidative stress-related cardiovascular diseases (CVDs) is still controversial and further studies are needed to investigate the correlation between high plasma SQ concentrations and CVDs.
This work focuses on current knowledge about SQ roles in humans, its endogenous and exogenous sources, and its beneficial and negative impact on health, with particular attention to metabolic and cardiovascular disorders.
2. Sources of Squalene
2.1. Endogenous Sources of Squalene
In human body, squalene is present in multiple organs and tissues. As expected, the liver is the main player of its biosynthesis and this organ represents the primary way through which SQ enters the systemic circulation. Animal studies demonstrate that interference in cholesterol biosynthesis in liver could influence squalene concentration both at the local (in hepatocytes) and systemic levels [
10].
SQ is present in plasma embedded into lipoproteins: its concentration is particularly high in very low density lipoproteins (VLDL), but significant concentrations of SQ are also found in low density lipoproteins (LDL) and high density lipoproteins (HDL), and its amount seems to be directly correlated with triglyceride abundance in these components [
11,
12].
Because of SQ hydrophobic nature, it could be found in a concentration of 275 µg/g in adipocytes [
13]. Squalene constitutes about 12% of the human skin surface lipids [
14], probably because, compared with other mammals, human skin is not so hairy, and therefore it needs more strategies to counteract the photo-oxidative action of UV rays [
15]. Moreover, several research studies have pointed out SQ high concentrations (about 475 µg/g) in epithelial tissue, suggesting a possible in loco synthesis of this compound, where sebaceous glands figure as probable sources of SQ [
4,
11,
13].
2.2. Exogenous Sources of Squalene
Recent studies have highlighted exogenous natural or synthetic sources of squalene. Indeed, SQ can be introduced through food products, food supplements and drugs [
2]. As previously mentioned, SQ was first isolated from shark liver oil in which its concentration reaches nearly 40% of the total oil [
2,
11,
13]. A shark’s liver can constitute up to 25% of its body mass, and the role of squalene in the liver is known to contribute together with diacyl glycerol esters to the control of animal flotation [
16]. Several works point out and support its dermatological use [
15]. Fortunately, today cosmetic industry is strictly regulated by laws which protect these animal species and SQ has to be isolated from other natural or synthetic sources.
In this scenario plants, where it is a precursor of phytosterols and other secondary plant metabolites, figure as alternative natural sources of squalene. SQ is highly concentrated in the unsaponifiable part of several plant oils. Furthermore, several studies demonstrate that plant levels of squalene are strictly correlated to the plant source, the location and period of plant growth and the extraction method used to separate SQ from other metabolites [
11]. Considering all these factors, it could be necessary to use high quantities of raw materials to extract SQ from plant rather than animal sources. For these reasons several methods have been applied in order to reach higher concentrations of SQ in plant extracts. In addition to traditional mechanical and chemical extraction, using ethanol [
17], new approaches have been developed with successful results, such as supercritical CO
2 extraction [
17,
18,
19].
The most relevant plant sources of SQ are oils extracted from amaranth, olive, rice, wheat germ, grape seed, peanut and soybean [
4,
11,
20]. As previously mentioned the abundance of SQ in the extract is influenced by several factors, but promising results obtained in this research field offer positive data on the possible use of plant-derived SQ in pharmaceutical and cosmetic sectors and in the nutraceutical field.
Regarding exogenous SQ fate in the digestive tract, animal and human studies have already tested the bioavailability of dietary SQ, underlining that nearly 60% of the ingested compound is absorbed. Non-absorbed SQ is in part excreted with feces and the rest probably metabolized by the gut microbiota, however further studies are needed to elucidate this pathway [
21,
22].
3. ROS/RNS and Squalene
3.1. ROS/RNS and Squalene
Oxidative stress leading to lipid peroxidation is triggered by different oxidizing molecules and, among them, O
2● is the first reactive oxygen species (ROS) involved in the damage of cell membranes. The presence of SQ seems to reduce this radical by releasing an electron, through reaction (1), at the moment observed only in ex vivo models and not yet demonstrated in cellular systems:
The stabilization of the radical occurs through the formation of a squalene hydroperoxide (HOO-SQ) and the electronic rearrangement of SQ by resonance, a typical phenomenon related to molecules containing a number of double bonds [
23]. This reaction leads to the formation of different isomers, which are specifically: 2-OOH-SQ, 3-OOH-SQ, 6-OOH-SQ, 7-OOH-SQ, 10-OOH-SQ, 11-OOH-SQ. In particular, the study by Shimizu et al. [
14], examined this behavior by oxidizing SQ with 3-(1,4-epidioxy-4-methyl-1,4-dihydro-1-naphytyl)propionic acid (EP), a compound used to release O
2 by thermal degradation. SQ oxidation occurs in a dose dependent fashion and the different isomers are produced in the same percentages. During this reaction O
2● acts as an oxidizing agent and reacts with a double bond to form hydroperoxides through the ene-reaction [
24].
The rate of this reaction is expressed through a kinetic constant: the quenching constant (Kq). Krasnovsky and colleagues calculated this value as the sum of three other kinetic constants [
25]: K
ph: expresses physical quenching; K
1: expresses kinetics reaction occurring between an oxidizing agent and an allylic hydrogen atom of the reducing agent, which is part of a methylene group between C=C and C–C; K
2: expresses kinetics reaction occurring between an oxidizing agent and an allylic hydrogen atom of the reducing agent, which is part of a methylene group between C=C and C=C. Since SQ does not have conjugated double bonds, this value is equal to 0.
Squalene Kq (Kq
SQ) has been measured by Kohno et al. [
8], in an environment saturated with butanol at 35 °C. Kq
SQ (2.66 × 10
6 M
−1 s
−1) is greater than the Kq of some membrane lipids, such as ethyl linolenate, eicosapentaenoic acid ethyl ester and docosahexaenoic acid ethyl ester. Since the Kq of saturated and monounsaturated fatty acids is lower than that of polyunsaturated acids [
25], and Kq
SQ shows the highest value among membrane polyunsaturated acids, the stabilization reaction will take place faster in presence of SQ rather than other membrane lipids. Therefore, SQ oxidation occurs earlier than other lipids, blocking the peroxidative reactions and thus protecting cell membranes. Furthermore, Kq
SQ has a lower value than Kq of α-Tocopherol, but similar to the Kq of dibutylhydroxytoluene (BHT) molecules, widely used as antioxidants in food industry.
Moreover, SQ is inserted in cell membrane and contributes to its stabilization [
26,
27]. In particular, in the plasma membrane, due to its hydrophobicity, it is oriented parallel to the plane of symmetry of the lipid bilayer, embedded between fatty acids terminal methyl groups of the two layers. The strategic position of SQ forms an intermediate barrier between the two phospholipidic layers and allows the molecule to act as a quencher that blocks uncontrolled electron flows. The role of SQ in stabilizing cell membranes underlines its important role also in cardiac cell membrane reconstitution and in the regulation of membrane proteins and ion movement [
26].
3.2. Oxidative Stress in Skin and Squalene
Antioxidant properties of SQ were primarily studied in epithelial tissue. In fact, the molecule is highly abundant in skin, that is the largest tissue exposed to different environmental stressors leading to oxidative stress, such as pollutants, photo-oxidation and UV-light. In particular, Aioi and colleagues show the scavenging activity of SQ on superoxide anion formation in keratinocytes exposed to oxidative stressors [
28], suggesting a protective role of the molecule that acts in combination to superoxide dismutase.
3.3. Cardiovascular Antioxidant Systems and Squalene
Recent studies, aiming to evaluate the effect of SQ on antioxidant enzyme levels and activity in cardiovascular and hepatic tissues, showed a positive correlation between SQ and glutathione and non-glutathione-dependent enzymatic defenses. In fact, even if the first antioxidant activity of SQ is linked to lipid peroxidation blockade, as already described, growing evidence is emerging on its involvement in the regulation of the expression and activation of glutathione peroxidase (GPx), catalase (CAT), superoxide dismutase (SOD) and glutathione S-transferase (GST) [
29,
30]. SQ prevents the alterations of SOD and CAT by detoxifying the cytosolic environment. Moreover, it acts reconstituting GSH and safeguarding the enzymatic activity of GPx. Furthermore, the GSSG/GSH ratio, which is considered as a marker of oxidative stress status, decreases after SQ treatment [
30].
The positive influence of SQ on antioxidant enzymes has been demonstrated in stress situations, in particular, in animal models of isoproterenol-induced myocardial infarction [
29]. Summarizing these results, it could be reasonable thinking the possible use of SQ in maintaining antioxidant enzyme activity in the heart and vascular system.
Table 1 summarizes different works in which the antioxidant properties of SQ were evaluated in cardiovascular system and in liver.
4. Oxidative Stress Associated CVDs and Squalene
The endothelium lining the lumen of blood vessels is pivotal in maintaining vascular function and its regulation has an important redox component. In fact, physiological ROS levels contribute to vasodilatation, angiogenesis, vascular remodeling and maintenance of lower blood pressure [
34]. The main source of ROS in the vasculature is the NADPH oxidase (NOX). NOX-derived ROS act as essential second messenger molecules contributing to the regulation of normal cell function [
35]. NOX4 and NOX2 isoforms are the major ROS producers in vascular smooth muscle cells and endothelial cells. In particular, H
2O
2 produced by NOX4 activates eNOS leading to the generation of NO, which exerts vasoprotective effects on the endothelium keeping blood vessels dilated and controlling blood pressure [
36,
37]. Moreover, vascular endothelial growth factor (VEGF), a physiological regulator of angiogenesis, induces endothelial ROS production through NOX4 and NOX2 activation and promotes endothelial cell proliferation, migration and survival [
38]. In addition, normal levels of ROS are critical for the physiological response of vascular smooth muscle cells. In particular NOX4-induced H
2O
2 promotes cell differentiation and the phenotypic switch from a proliferative state to a contractile state, regulating the vascular tone [
39].
Moreover, NOX4 is expressed not only in endothelial and vascular smooth muscle cells, but also in cardiomyocytes, in which it regulates autophagy, leading to its steady activation, and cytoprotective and antioxidant pathways [
40].
Adverse cardiocirculatory events induced by oxidative stress are primarily linked to endothelial dysfunction [
41,
42,
43,
44], characterized by a complex multifactorial pathway, including eNOS uncoupling and increased concentration of inflammatory cytokines, both exacerbating the damaging effect induced by ROS/RNS boost [
41]. Among risk factors that contribute to the development of endothelial dysfunction, some are classified as non-modifiable factors, as aging and family history of CVDs, other are classified as modifiable risk factors and are related to lifestyle [
41].
The following paragraphs describe the role of SQ in different oxidative stress associated CVDs, that are strictly linked to endothelial dysfunction development, and underline the possible use of SQ as a natural supplement to support standard drug therapy.
4.1. Hypercholesterolemia
Hypercholesterolemia is a metabolic disorder characterized by high levels of circulating low density lipoproteins (LDL) and related adverse effects. According to clinical health guidelines, reference values are: total cholesterol < 200 mg/dL, LDL < 100 mg/dL and HDL > 50 mg/dL; higher circulating values of total cholesterol and LDL have to be considered as hypercholesterolemic conditions [
45].
Presence of high levels of LDL and ROS can contribute to the development of atherosclerosis: oxidized LDL (oxLDL) are taken up by macrophages that become foam cells; macrophages also produce cytokines that recruit T cells that are involved in amplifying the inflammation. Endothelial cells produce cytokines that stimulate smooth muscle cell proliferation and migration; foam cells and smooth muscle cells form a lipid core surrounded by fibrotic tissue, the atherosclerotic plaque, highly susceptible to rupture and thrombus formation [
42]. Given the high instability of the plaque and the possible negative effects derived from its rupture, it is important to interfere with both its onset and progression with hypocholesterolemic agents.
SQ, as a precursor of cholesterol, could be reasonably considered as an exacerbating factor for hypercholesterolemia. Despite that, Khor et al. demonstrated that addition of SQ to a high fat diet in hamsters does not affect levels of LDL, HDL and triglycerides and that the increased cholesterol ester accumulation in SQ fed hamster liver could account for the hypocholesterolemic effects of the supplemented chow [
46,
47]. Furthermore, Kritchevsky and colleagues demonstrated that a SQ based diet for 7 weeks in rabbits does not induce atheroma formation neither is directly involved in atherosclerosis development [
46,
48]. Other studies underline sex-related responses to SQ supplementation in ApolipoproteinE (ApoE
-/-) KO mice, a useful animal model for atherosclerosis: males show a reduction of atherosclerotic lesion area independently from plasmatic lipid panel, while females do not show a reduction of atherosclerotic lesion area, but have significantly reduced plasma levels of cholesterol and triglycerides [
49]. Other studies pointed out the molecular mechanism involved in SQ effects, some of them highlighting the activation of liver X receptors (LXRs), nuclear receptors engaged in homeostasis of cholesterol, fatty acids and glucose. In particular, a recent study shows that SQ acts as a selective modulator of LXRs in macrophages, inhibiting cholesterol accumulation and promoting its efflux; moreover, in hepatocytes SQ activates PPARα, thus upregulating the expression of genes related to fatty acid uptake, fatty acid oxidation, ketogenesis and reverse cholesterol transport metabolism. Based on these considerations, SQ represents a natural compound that can be used in dyslipidemic patients without negative side effects induced by standard administered drugs [
50,
51].
Granados-Principal and colleagues also studied SQ role in atherosclerotic plaque formation: they observed a dose-dependent reduction of surface expression of the oxLDL receptor CD36, in monocytes and macrophages and a consequent reduction in oxLDL uptake; furthermore, SQ exerted antiproliferative effect on monocytes and macrophages, thus contributing in atherosclerosis decline [
52].
SQ also seems to be directly involved in cholesterol synthesis break, through a negative feedback mechanism: SQ alone or in combination with hypocholesterolemic drugs, such as statins, promotes both the down-regulation of HMG-CoA reductase, a key enzyme in cholesterol biosynthesis, and the cholesterol and bile acid excretion [
53,
54].
4.2. Hypertension
Hypertension is a complex, multifactorial, and multisystem disorder. Several studies have shown that endothelial dysfunction is an early and common trigger of hypertension and it is characterized by an impairment of the physiological pathways controlling vascular tone, primarily the NO pathway. Burst of ROS in dysfunctional endothelial cells lead to the irreversible production of peroxynitrite (ONOO
−); ONOO
− can rapidly diffuse trough the cell and oxidize proteins, inducing eNOS uncoupling and thus decreasing NO production [
55]. Furthermore, high ROS levels enhance angiotensin II signaling in resistance arteries and induce vascular smooth muscle cell hypertrophy [
38].
New antihypertensive approaches based on nutraceutical properties of some food and plant extracts and, among them, squalene has shown positive effects in lowering blood pressure. Martirosyan and colleagues reported that SQ-rich amaranth oil reduces systolic blood pressure in a concentration dependent way [
56]. Furthermore, Liu et al. showed that orally administered SQ for 4 weeks in rats diminishes both plasma lipids and blood pressure, probably through a reduction in circulating levels of leptin [
46,
57].
4.3. Hyperglycemia-Dependent Endothelial Dysfunction
Hyperglycemia is characterized by high fasting glucose levels in plasma, >200 mg/dL, due to genetic factors or wrong lifestyle habits. Chronic high glycemia causes enhancement of different molecules, in particular, polyol, advanced glycation end products, protein kinase C and exosamine, all involved in multiorgan damages. ROS rising, due to NOX enhanced activity and eNOS uncoupling, is strictly related to endothelial dysfunction associated to hyperglycemia [
58]. Whereby dietary interventions in reducing glycemia are scant, hypoglycemic drugs are prescribed.
In recent years, many studies have suggested the use of plant extracts in order to lower plasma glucose and to prevent the development of ROS-induced endothelial dysfunction: among them, squalene is proposed as an antioxidant molecule capable of lowering both endothelial ROS and glycemia in hyperglycemic patients. At this regard, Liu and colleagues observed a reduction in plasma glucose in male Wistar rats fed with a high SQ diet in a time-dependent trend [
57], while, Widyawati and collaborators highlight an hypoglycemic effect of a SQ-rich extract of
Sygyzium polyanthum, mediated by a reduced glucose absorption in the gut and by an increased glucose uptake in the skeletal muscle [
59]. Conversely, Valdes et al. showed no significant blood glucose reduction by SQ in streptozotocin-induced diabetic mice, pointing out the need of further investigation to define the hypoglycemic role of SQ [
60].
5. Pathological Implications of Squalene
As a precursor of cholesterol biosynthesis, SQ both from endogenous or exogenous sources [
61,
62], the roles of SQ as a marker or a risk factor in the development of CVDs are essential. In order to define SQ implication in cardiovascular pathology, a few studies underline the possible strategic role of the molecule in coronary artery disease (CAD) [
63] and in the development of visceral obesity and metabolic syndrome [
64,
65]. Moreover, emerging therapies for dyslipidemia are addressed to squalene synthase as a possible pharmacological target [
21,
66,
67].
5.1. Coronary Artery Disease
Coronary artery disease (CAD) is a pathological condition characterized by atherosclerotic plaque formation in coronary vessels and it is strictly related to oxidative stress status. CAD is a multifactorial pathology in which both genetic and environmental factors contribute to its development. Studies underline different risk factors that are involved in the progression of this condition, among others, high LDL and low HDL cholesterol in plasma, hypertension, diabetes, smoking, sex and age. CAD could open to more severe conditions such as myocardial infarction and heart failure, so, it is important to stop the progression of the pathology lowering existing risk factors [
68]. Rajaratnam and colleagues underline that in menopausal women high plasma concentration of squalene could represent a risk factor for CAD: they found higher plasma SQ related to serum cholesterol in women with cardiovascular predisposition to CAD than in controls. Moreover, elevated concentration of SQ were found in atherosclerotic plaques delivered by LDL [
63]. The authors ascribed the high serum concentration of SQ to the decreased cholesterol synthesis resulting from hypercholesterolemic diet, and suggest SQ as a marker for CAD diagnosis [
63,
69,
70,
71].
5.2. Metabolic Syndrome
Metabolic syndrome is a multifactorial disease characterized by dyslipidemia, hypertension, diabetes and obesity [
72,
73]. Several interventions are needed to reverse this condition: life style changes are the first line modifications, and, to overcome critical situations, drug administration could be necessary.
In this scenario, abdominal adiposity and obesity are evident phenotypes of metabolic syndrome, when other latent symptoms are not yet manifested. Peltola and colleagues highlighted that high levels of serum SQ are linked to visceral obesity; as adipose tissue synthesizes and stores SQ, which is only in part converted to cholesterol, the authors suggest that SQ in adipose tissue could have detrimental effects in abdominal obesity, thus suggesting SQ as marker for metabolic syndrome [
65]. According to these results, Lupattelli and collaborators demonstrated that in metabolic syndrome low synthesis of cholesterol and high absorption of SQ occur, thus explaining its high plasma concentrations [
64]. Only a few data are available defining the critical role of SQ in metabolic syndrome and in the control of the resulting systemic oxidative stress status. They open essential questions about its direct or simply marker role in the onset of the pathological picture typical of metabolic syndrome.
5.3. Squalene Synthase Inhibitors
In order to assess a complete description of current knowledge about squalene and CVDs. it could be necessary also to define new advances in pharmacological therapies for dyslipidemia and the role of SQ in this pathological condition, strictly associated with the rise of circulating free radicals. Today most used drugs to treat dyslipidemia are statins, that block cholesterol biosynthesis inhibiting HMG-CoA reductase activity, fibrates and bile-acid sequestrants [
27,
61,
66]. New frontiers are under investigation because these drugs have several adverse side effects when chronically administered.
For this reason, other enzymes in the cholesterol biosynthetic pathway are under investigation as putative pharmacological targets; among them, squalene synthase (SQS) represents a good candidate because it catalyzes one of the last reactions in cholesterol synthesis; indeed, its blockade does not interfere with the synthesis of other fundamental intermediates in the pathway, such as geranylgeranyl diphosphate or ubiquinone, and because only moderate side effect has been highlighted using this approach [
67].
As a consequence of inhibition of SQS, low levels of endogenous squalene are reached and this may suggest a possible role of the molecule in the onset of hypercholesterolemia that have to be confirmed with further investigations [
61,
62,
66].
6. Nutraceutical Properties of SQ
Nutraceuticals are defined as foods or ingredients or natural extracts that have a role in promoting health, thus preventing or treating some pathologies [
74,
75]. The previously described multiple beneficial effects of SQ support its potential nutraceutical properties and its possible use as food additive to prevent or treat some pathological conditions [
74,
75]. Toxicological studies show that SQ is well tolerated when consumed orally and that it has faster time of absorbance when compared to cholesterol [
4,
26].
Even if some nutraceutical properties of SQ have already been mentioned, recent animal studies underline in a more detailed manner antioxidant and cardioprotective effects of SQ- enriched diets. Indeed, SQ enriched diets show beneficial effects on mitochondrial energy status, lipid peroxidation and membrane stability. A number of research groups observed a stronger scavenging activity of SQ compared to vitamin E in elderly people [
31]. Administration of SQ seems to have several positive effects on cardiovascular system, reducing atherosclerosis through induction of paraoxonase 1 (PON1) protective against the development of atherosclerosis [
33], enhancing peripheral blood flow and improving central arterial elasticity [
76]. Moreover, chemoprotective effects of SQ are currently under investigation [
32,
77,
78,
79,
80,
81].
7. Squalene: Healthy or Harmful in CVDs? Summary and Future Directions
The aim of this paragraph is to summarize both positive and negative effects of SQ in CVDs, as emerged from the previous sections, and to try to dissolve the vagueness around its pathophysiological role.
As pointed out in
Table 2, all the results suggesting a role of SQ in the onset of CVDs arise from correlation studies, while a number of research articles reporting positive role of SQ, properly deepen the molecular mechanisms involved and this is in agreement with Ibrahim and colleagues [
46]. Among them, activation of LXRs in macrophages [
50], activation of PPARα in hepatocytes [
51], reduction of CD36 expression in monocytes and macrophages [
52] and downregulation of HMG-CoA reductase in the liver [
53], strongly support the hypolipidemic role of SQ and its resulting anti-atherogenic effect. Based on these observations, future studies in SQ and CVDs should focus on cellular and molecular pathways directly affected by the triterpene, thus integrating clinical, cellular, molecular and biochemical analysis.
8. Conclusions
Squalene is a key metabolite in the sterol pathway and it is pivotal in regulating cellular and systemic physiology in eukaryotic organisms. Its role as a metabolic intermediate underlines its involvement in the regulation of different biochemical pathways, such as the regulation of sterol homeostasis (
Figure 1). Despite the wide distribution of SQ in animal organisms, its direct role in regulating both pathologic and non-pathologic conditions is an important frontier of investigation.
In particular, its involvement in oxidative stress responses in human is still debated. Several studies indicate a dual role of the compound primarily linked to its chemical structure: the possible implication of SQ in antioxidant cellular response, and, in the other hand, the possible contribution of SQ in exacerbating detrimental effects of oxidative stress associated diseases.
This review focuses on the role of SQ in the development of oxidative stress associated CVDs. Treatment of most common CVDs, such as hypercholesterolemia, hypertension and hyperglycemia, with SQ enriched-diet shows promising results, underlining both quenching activity and enhancement of antioxidant systems induced by SQ against ROS/RNS burst [
46]. Moreover, giving the emerging pivotal role of LXRs as a beneficial modulator of cardiovascular physiology [
82], and the recent findings of SQ as an agonist of LXRs [
50], future studies could be aimed at investigating the potential link between squalene and LXRs in cardiomyocytes and endothelial cells and in deepen the resulting intracellular pathway.
Even if chemical characterization and ex vivo results suggest a possible protective role of SQ against oxidative stress and free radicals, further human trials are needed to fully elucidate the cellular pathways activated by SQ in the regulation of cardiovascular ROS/RNS production.
Author Contributions
Conceptualization, M.P.G. and C.M.B.; writing—original draft preparation, G.Q., M.M., F.G.; writing—review and editing, M.P.G., R.L., S.A., C.M.B.; supervision, M.P.G., C.M.B.; project administration, M.P.G., C.M.B., A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the local funding of the University of Turin and by funding from Regione Piemonte (Fondo Sociale Europeo—Apprendistato di Alta Formazione e di Ricerca, 2016–2020) for the research activity of MM and assigned to CMB.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, S.-K.; Karadeniz, F. Chapter 14—Biological Importance and Applications of Squalene and Squalane. In Advances in Food and Nutrition Research; Kim, S.-K., Ed.; Marine Medicinal Foods; Academic Press: Cambridge, MA, USA, 2012; Volume 65, pp. 223–233. [Google Scholar]
- Bhilwade, H.N.; Tatewaki, N.; Nishida, H.; Konishi, T. Squalene as novel food factor. Curr. Pharm. Biotechnol. 2010, 11, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, M. A highly unsaturated hydrocarbon in shark liver oil. J. Ind. Eng. Chem. 1916, 8, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Reddy, L.H.; Couvreur, P. Squalene: A natural triterpene for use in disease management and therapy. Adv. Drug Deliv. Rev. 2009, 61, 1412–1426. [Google Scholar] [CrossRef] [PubMed]
- Rezen, T.; Debeljak, N.; Kordis, D.; Rozman, D. New aspects on lanosterol 14alpha-demethylase and cytochrome P450 evolution: Lanosterol/cycloartenol diversification and lateral transfer. J. Mol. Evol. 2004, 59, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Tansey, T.R.; Shechter, I. Structure and regulation of mammalian squalene synthase. Biochimica et Biophysica Acta (BBA)—Mol. Cell Biol. Lipids 2000, 1529, 49–62. [Google Scholar] [CrossRef]
- Spanova, M.; Daum, G. Squalene—Biochemistry, molecular biology, process biotechnology, and applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1299–1320. [Google Scholar] [CrossRef]
- Kohno, Y.; Egawa, Y.; Itoh, S.; Nagaoka, S.; Takahashi, M.; Mukai, K. Kinetic study of quenching reaction of singlet oxygen and scavenging reaction of free radical by squalene in n-butanol. Biochim. Biophys. Acta 1995, 1256, 52–56. [Google Scholar] [CrossRef]
- O’Sullivan, L.; Woods, J.A.; O’Brien, N.M. Squalene but not n-3 fatty acids protect against hydrogen peroxide-induced sister chromatid exchanges in Chinese hamster V79 cells. Nutr. Res. 2002, 22, 847–857. [Google Scholar] [CrossRef]
- Ott, D.B.; Lachance, P.A. Biochemical controls of liver cholesterol biosynthesis. Am. J. Clin. Nutr. 1981, 34, 2295–2306. [Google Scholar] [CrossRef] [Green Version]
- Popa, I.; Băbeanu, N.; Niță, S.; Popa, O. Squalene-Natural resources and applications. Farmacia 2014, 62, 840–862. [Google Scholar]
- Saudek, C.D.; Frier, B.M.; Liu, G.C. Plasma squalene: Lipoprotein distribution and kinetic analysis. J. Lipid Res. 1978, 19, 827–835. [Google Scholar] [PubMed]
- Liu, G.C.; Ahrens, E.H.; Schreibman, P.H.; Crouse, J.R. Measurement of squalene in human tissues and plasma: Validation and application. J. Lipid Res. 1976, 17, 38–45. [Google Scholar] [PubMed]
- Shimizu, N.; Ito, J.; Kato, S.; Otoki, Y.; Goto, M.; Eitsuka, T.; Miyazawa, T.; Nakagawa, K. Oxidation of squalene by singlet oxygen and free radicals results in different compositions of squalene monohydroperoxide isomers. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.-R.; Lin, Y.-K.; Fang, J.-Y. Biological and pharmacological activities of squalene and related compounds: Potential uses in cosmetic dermatology. Molecules 2009, 14, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Gleiss, A.C.; Potvin, J.; Goldbogen, J.A. Physical trade-offs shape the evolution of buoyancy control in sharks. Proc. Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraujalis, P.; Venskutonis, P.R. Supercritical carbon dioxide extraction of squalene and tocopherols from amaranth and assessment of extracts antioxidant activity. J. Supercrit. Fluids 2013, 80, 78–85. [Google Scholar] [CrossRef]
- Wejnerowska, G.; Heinrich, P.; Gaca, J. Separation of squalene and oil from Amaranthus seeds by supercritical carbon dioxide. Sep. Purif. Technol. 2013, 110, 39–43. [Google Scholar] [CrossRef]
- De Vita, D.; Messore, A.; Toniolo, C.; Frezza, C.; Scipione, L.; Bertea, C.M.; Micera, M.; Di Sarno, V.; Madia, V.N.; Pindinello, I.; et al. Towards a new application of amaranth seed oil as an agent against Candida albicans. Nat. Prod. Res. 2019, 1–6. [Google Scholar] [CrossRef]
- Tikekar, R.V.; Ludescher, R.D.; Karwe, M.V. Processing stability of squalene in amaranth and antioxidant potential of amaranth extract. J. Agric. Food Chem. 2008, 56, 10675–10678. [Google Scholar] [CrossRef]
- Relas, H.; Gylling, H.; Miettinen, T.A. Dietary squalene increases cholesterol synthesis measured with serum non-cholesterol sterols after a single oral dose in humans. Atherosclerosis 2000, 152, 377–383. [Google Scholar] [CrossRef]
- Strandberg, T.E.; Tilvis, R.S.; Miettinen, T.A. Metabolic variables of cholesterol during squalene feeding in humans: Comparison with cholestyramine treatment. J. Lipid Res. 1990, 31, 1637–1643. [Google Scholar] [PubMed]
- Belitz, H.-D.; Grosch, W.; Schieberle, P. Food Chemistry, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-3-540-69933-0. [Google Scholar]
- Yardımcı, Ş.D.; Kaya, N.; Balci, M. Regioselectivity in the ene-reaction of singlet oxygen with cyclic alkenes: Photooxygenation of methyl-substituted 1,4-cyclohexadiene derivatives. Tetrahedron 2006, 62, 10633–10638. [Google Scholar] [CrossRef]
- Krasnovsky, A.A.; Kagan, V.E.; Minin, A.A. Quenching of singlet oxygen luminescence by fatty acids and lipids: Contribution of physical and chemical mechanisms. FEBS Lett. 1983, 155, 233–236. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Grande, M.A.; Gorinstein, S.; Espitia-Rangel, E.; Dávila-Ortiz, G.; Martínez-Ayala, A.L. Plant Sources, Extraction Methods, and Uses of Squalene. Available online: https://www.hindawi.com/journals/ija/2018/1829160/ (accessed on 12 March 2020).
- Nathan, J.A. Squalene and cholesterol in the balance at the ER membrane. Proc. Natl. Acad. Sci. USA 2020, 117, 8228–8230. [Google Scholar] [CrossRef] [PubMed]
- Aioi, A.; Shimizu, T.; Kuriyama, K. Effect of squalene on superoxide anion generation induced by a skin irritant, lauroylsarcosine. Int. J. Pharm. 1995, 113, 159–164. [Google Scholar] [CrossRef]
- Sabeena Farvin, K.H.; Anandan, R.; Kumar, S.H.S.; Shiny, K.S.; Sankar, T.V.; Thankappan, T.K. Effect of squalene on tissue defense system in isoproterenol-induced myocardial infarction in rats. Pharmacol. Res. 2004, 50, 231–236. [Google Scholar] [CrossRef]
- Ravi Kumar, S.; Narayan, B.; Sawada, Y.; Hosokawa, M.; Miyashita, K. Combined effect of astaxanthin and squalene on oxidative stress in vivo. Mol. Cell. Biochem. 2016, 417, 57–65. [Google Scholar] [CrossRef]
- Buddhan, S.; Sivakumar, R.; Dhandapani, N.; Ganesan, B.; Anandan, R. Protective effect of dietary squalene supplementation on mitochondrial function in liver of aged rats. Prostaglandins Leukot. Essent. Fatty Acids 2007, 76, 349–355. [Google Scholar] [CrossRef]
- Motawi, T.M.K.; Sadik, N.A.H.; Refaat, A. Cytoprotective effects of DL-alpha-lipoic acid or squalene on cyclophosphamide-induced oxidative injury: An experimental study on rat myocardium, testicles and urinary bladder. Food Chem. Toxicol. 2010, 48, 2326–2336. [Google Scholar] [CrossRef]
- Gabás Rivera, C.; Barranquero, C.; Martínez-Beamonte, R.; Navarro Ferrando, M.Á.; Surra, J.; Osada, J. Dietary Squalene Increases High Density Lipoprotein-Cholesterol and Paraoxonase 1 and Decreases Oxidative Stress in Mice. PLoS ONE 2014, 9, e104224. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Knock, G.A. NADPH oxidase in the vasculature: Expression, regulation and signalling pathways; role in normal cardiovascular physiology and its dysregulation in hypertension. Free Radic. Biol. Med. 2019, 145, 385–427. [Google Scholar] [CrossRef] [PubMed]
- Bretón-Romero, R.; González de Orduña, C.; Romero, N.; Sánchez-Gómez, F.J.; de Álvaro, C.; Porras, A.; Rodríguez-Pascual, F.; Laranjinha, J.; Radi, R.; Lamas, S. Critical role of hydrogen peroxide signaling in the sequential activation of p38 MAPK and eNOS in laminar shear stress. Free Radic. Biol. Med. 2012, 52, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, Q.; Zhu, J.; Xiao, Q.; Zhang, L. Reactive oxygen species: Key regulators in vascular health and diseases. Br. J. Pharmacol. 2018, 175, 1279–1292. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Luo, Z.; Pepe, A.E.; Margariti, A.; Zeng, L.; Xu, Q. Embryonic stem cell differentiation into smooth muscle cells is mediated by Nox4-produced H2O2. Am. J. Physiol. Cell Physiol. 2009, 296, C711–C723. [Google Scholar] [CrossRef] [Green Version]
- Rajaram, R.D.; Dissard, R.; Jaquet, V.; de Seigneux, S. Potential benefits and harms of NADPH oxidase type 4 in the kidneys and cardiovascular system. Nephrol. Dial. Transplant. 2019, 34, 567–576. [Google Scholar] [CrossRef]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [Green Version]
- Cervantes Gracia, K.; Llanas-Cornejo, D.; Husi, H. CVD and Oxidative Stress. J. Clin. Med. 2017, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, P.; Rengarajan, T.; Thangavel, J.; Nishigaki, Y.; Sakthisekaran, D.; Sethi, G.; Nishigaki, I. The Vascular Endothelium and Human Diseases. Int. J. Biol. Sci. 2013, 9, 1057–1069. [Google Scholar] [CrossRef] [Green Version]
- Endemann, D.H.; Schiffrin, E.L. Endothelial dysfunction. J. Am. Soc. Nephrol. 2004, 15, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Institute for Quality and Efficiency in Health Care (IQWiG). High Cholesterol: Overview; IQWiG: Cologne, Germany, 2017. [Google Scholar]
- Ibrahim, N.I.; Fairus, S.; Zulfarina, M.S.; Naina Mohamed, I. The Efficacy of Squalene in Cardiovascular Disease Risk-A Systematic Review. Nutrients 2020, 12, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khor, H.T.; Chieng, D.Y. Lipidaemic effects of tocotrienols, tocopherols and squalene: Studies in the hamster. Asia Pac. J. Clin. Nutr. 1997, 6, 36–40. [Google Scholar] [PubMed]
- Kritchevsky, D.; Moyer, A.W.; Tesar, W.C.; Logan, J.B.; Brown, R.A.; Richmond, G. Squalene feeding in experimental atherosclerosis. Circ. Res. 1954, 2, 340–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillén, N.; Acín, S.; Navarro, M.A.; Perona, J.S.; Arbonés-Mainar, J.M.; Arnal, C.; Sarría, A.J.; Surra, J.C.; Carnicer, R.; Orman, I.; et al. Squalene in a sex-dependent manner modulates atherosclerotic lesion which correlates with hepatic fat content in apoE-knockout male mice. Atherosclerosis 2008, 197, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Hien, H.T.M.; Ha, N.C.; Thom, L.T.; Hong, D.D. Squalene promotes cholesterol homeostasis in macrophage and hepatocyte cells via activation of liver X receptor (LXR) α and β. Biotechnol. Lett. 2017, 39, 1101–1107. [Google Scholar] [CrossRef]
- Hoang, T.M.H.; Nguyen, C.H.; Le, T.T.; Hoang, T.H.Q.; Ngo, T.H.T.; Hoang, T.L.A.; Dang, D.H. Squalene isolated from Schizochytrium mangrovei is a peroxisome proliferator-activated receptor-α agonist that regulates lipid metabolism in HepG2 cells. Biotechnol. Lett. 2016, 38, 1065–1071. [Google Scholar] [CrossRef]
- Granados-Principal, S.; Quiles, J.L.; Ramirez-Tortosa, C.L.; Ochoa-Herrera, J.; Perez-Lopez, P.; Pulido-Moran, M.; Ramirez-Tortosa, M.C. Squalene ameliorates atherosclerotic lesions through the reduction of CD36 scavenger receptor expression in macrophages. Mol. Nutr. Food Res. 2012, 56, 733–740. [Google Scholar] [CrossRef]
- Chan, P.; Tomlinson, B.; Lee, C.B.; Lee, Y.S. Effectiveness and safety of low-dose pravastatin and squalene, alone and in combination, in elderly patients with hypercholesterolemia. J. Clin. Pharmacol. 1996, 36, 422–427. [Google Scholar] [CrossRef]
- Shin, D.H.; Heo, H.J.; Lee, Y.J.; Kim, H.K. Amaranth squalene reduces serum and liver lipid levels in rats fed a cholesterol diet. Br. J. Biomed. Sci. 2004, 61, 11–14. [Google Scholar] [CrossRef]
- Pinheiro, L.C.; Oliveira-Paula, G.H. Sources and effects of oxidative stress in hypertension. Curr. Hypertens. Rev. 2019. [Google Scholar] [CrossRef] [PubMed]
- Martirosyan, D.M.; Miroshnichenko, L.A.; Kulakova, S.N.; Pogojeva, A.V.; Zoloedov, V.I. Amaranth oil application for coronary heart disease and hypertension. Lipids Health Dis. 2007, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xu, X.; Bi, D.; Wang, X.; Zhang, X.; Dai, H.; Chen, S.; Zhang, W. Influence of squalene feeding on plasma leptin, testosterone & blood pressure in rats. Indian J. Med. Res. 2009, 129, 150–153. [Google Scholar] [PubMed]
- Fiorentino, T.V.; Prioletta, A.; Zuo, P.; Folli, F. Hyperglycemia-induced oxidative stress and its role in diabetes mellitus related cardiovascular diseases. Curr. Pharm. Des. 2013, 19, 5695–5703. [Google Scholar] [CrossRef] [PubMed]
- Widyawati, T.; Yusoff, N.A.; Asmawi, M.Z.; Ahmad, M. Antihyperglycemic Effect of Methanol Extract of Syzygium polyanthum (Wight.) Leaf in Streptozotocin-Induced Diabetic Rats. Nutrients 2015, 7, 7764–7780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdes, M.; Calzada, F.; Mendieta-Wejebe, J. Structure–Activity Relationship Study of Acyclic Terpenes in Blood Glucose Levels: Potential α-Glucosidase and Sodium Glucose Cotransporter (SGLT-1) Inhibitors. Molecules 2019, 24, 4020. [Google Scholar] [CrossRef] [Green Version]
- Visser, M.E.; Jakulj, L.; Kastelein, J.J.P.; Stroes, E.S.G. LDL-C-lowering therapy: Current and future therapeutic targets. Curr. Cardiol. Rep. 2008, 10, 512–520. [Google Scholar] [CrossRef]
- Wierzbicki, A.S. New lipid-lowering agents. Expert Opin. Emerg. Drugs 2003, 8, 365–376. [Google Scholar] [CrossRef]
- Rajaratnam, R.A.; Gylling, H.; Miettinen, T.A. Independent association of serum squalene and noncholesterol sterols with coronary artery disease in postmenopausal women. J. Am. Coll. Cardiol. 2000, 35, 1185–1191. [Google Scholar] [CrossRef] [Green Version]
- Lupattelli, G.; De Vuono, S.; Mannarino, E. Patterns of cholesterol metabolism: Pathophysiological and therapeutic implications for dyslipidemias and the metabolic syndrome. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 620–627. [Google Scholar] [CrossRef]
- Peltola, P.; Pihlajamäki, J.; Koutnikova, H.; Ruotsalainen, E.; Salmenniemi, U.; Vauhkonen, I.; Kainulainen, S.; Gylling, H.; Miettinen, T.A.; Auwerx, J.; et al. Visceral obesity is associated with high levels of serum squalene. Obesity (Silver Spring) 2006, 14, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Kourounakis, A.P.; Katselou, M.G.; Matralis, A.N.; Ladopoulou, E.M.; Bavavea, E. Squalene synthase inhibitors: An update on the search for new antihyperlipidemic and antiatherosclerotic agents. Curr. Med. Chem. 2011, 18, 4418–4439. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Chu, C.-L.; Chen, L.; Shui, D. Evaluation of potential inhibitors of squalene synthase based on virtual screening and in vitro studies. Comput. Biol. Chem. 2019, 80, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Wang, J.; Tian, J.; Tang, Y. Coronary Artery Disease: From Mechanism to Clinical Practice. Adv. Exp. Med. Biol. 2020, 1177, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Rajaratnam, R.A.; Gylling, H.; Miettinen, T.A. Serum squalene in postmenopausal women without and with coronary artery disease. Atherosclerosis 1999, 146, 61–64. [Google Scholar] [CrossRef]
- Rajaratnam, R.A.; Gylling, H.; Miettinen, T.A. Impaired postprandial clearance of squalene and apolipoprotein B-48 in post-menopausal women with coronary artery disease. Clin. Sci. 1999, 97, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Gylling, H.; Hallikainen, M.; Rajaratnam, R.A.; Simonen, P.; Pihlajamäki, J.; Laakso, M.; Miettinen, T.A. The metabolism of plant sterols is disturbed in postmenopausal women with coronary artery disease. Metab. Clin. Exp. 2009, 58, 401–407. [Google Scholar] [CrossRef]
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef]
- Samson, S.L.; Garber, A.J. Metabolic syndrome. Endocrinol. Metab. Clin. N. Am. 2014, 43, 1–23. [Google Scholar] [CrossRef]
- Das, L.; Bhaumik, E.; Raychaudhuri, U.; Chakraborty, R. Role of nutraceuticals in human health. J. Food Sci. Technol. 2012, 49, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Nasri, H.; Baradaran, A.; Shirzad, H.; Rafieian-Kopaei, M. New concepts in nutraceuticals as alternative for pharmaceuticals. Int. J. Prev. Med. 2014, 5, 1487–1499. [Google Scholar] [PubMed]
- Hamadate, N.; Matsumoto, Y.; Seto, K.; Yamamoto, T.; Yamaguchi, H.; Nakagawa, T.; Yamamoto, E.; Fukagawa, M.; Yazawa, K. Vascular effects and safety of supplementation with shark liver oil in middle-aged and elderly males. Exp. Ther. Med. 2015, 10, 641–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, T.J. Squalene: Potential chemopreventive agent. Expert Opin. Investig. Drugs 2000, 9, 1841–1848. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.V.; Newmark, H.L.; Reddy, B.S. Chemopreventive effect of squalene on colon cancer. Carcinogenesis 1998, 19, 287–290. [Google Scholar] [CrossRef] [Green Version]
- Maksimenko, A.; Dosio, F.; Mougin, J.; Ferrero, A.; Wack, S.; Reddy, L.H.; Weyn, A.-A.; Lepeltier, E.; Bourgaux, C.; Stella, B.; et al. A unique squalenoylated and nonpegylated doxorubicin nanomedicine with systemic long-circulating properties and anticancer activity. Proc. Natl. Acad. Sci. USA 2014, 111, E217–E226. [Google Scholar] [CrossRef] [Green Version]
- Senthilkumar, S.; Devaki, T.; Manohar, B.M.; Babu, M.S. Effect of squalene on cyclophosphamide-induced toxicity. Clin. Chim. Acta 2006, 364, 335–342. [Google Scholar] [CrossRef]
- Narayan, B.H.; Tatewaki, N.; Giridharan, V.V.; Nishida, H.; Konishi, T. Modulation of doxorubicin-induced genotoxicity by squalene in Balb/c mice. Food Funct. 2010, 1, 174–179. [Google Scholar] [CrossRef]
- Cannon, M.V.; van Gilst, W.H.; de Boer, R.A. Emerging role of liver X receptors in cardiac pathophysiology and heart failure. Basic Res. Cardiol. 2016, 111, 3. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).

 ,
, 


{kind=link}
{kind=link}