Reactive Sulfur Species Interact with Other Signal Molecules in Root Nodule Symbiosis in Lotus japonicus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Materials and Growth Conditions
2.2. Nitrogenase Activity
2.3. Endogenous Signal Molecule Production in Roots and Nodules
2.4. Treatment of Roots and Nodules with the Sulfane Sulfur Donor Na2S3
3. Results
3.1. Detection of Polysulfides in the Roots of Lotus japonicus by using SSP4
3.2. Effect of Na2S3 on the Amount of NO, ROS, and H2S in Roots
3.3. Localization of RSS and Signal Molecules during Rhizobial Infection
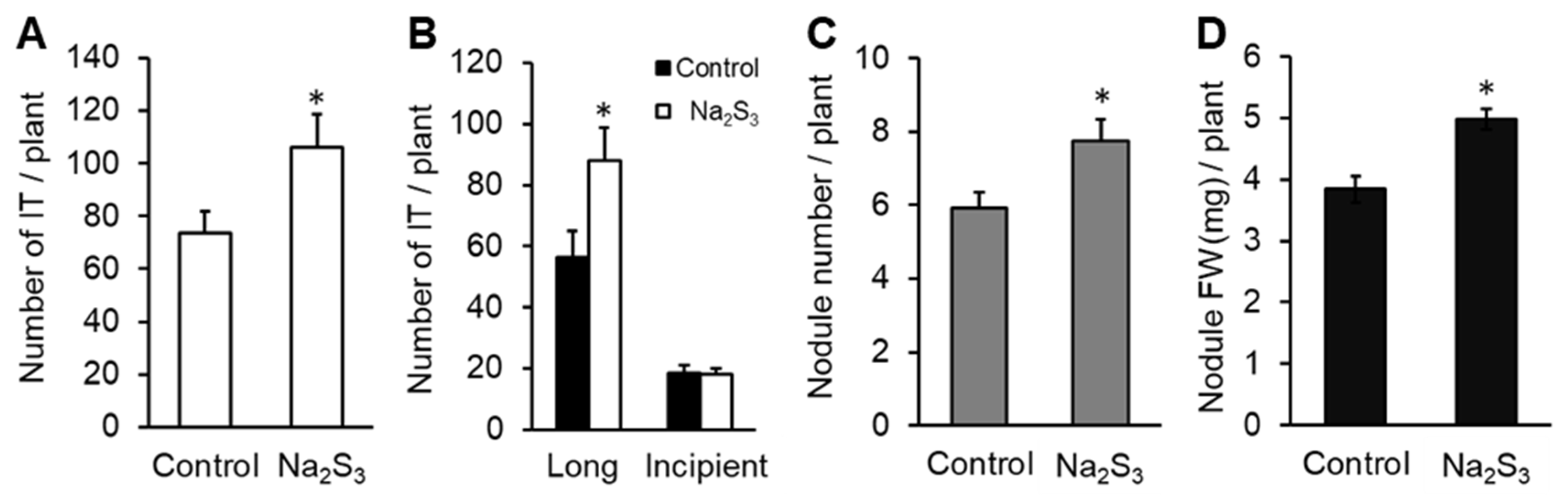
3.4. Effect of RSS on Rhizobial Infection
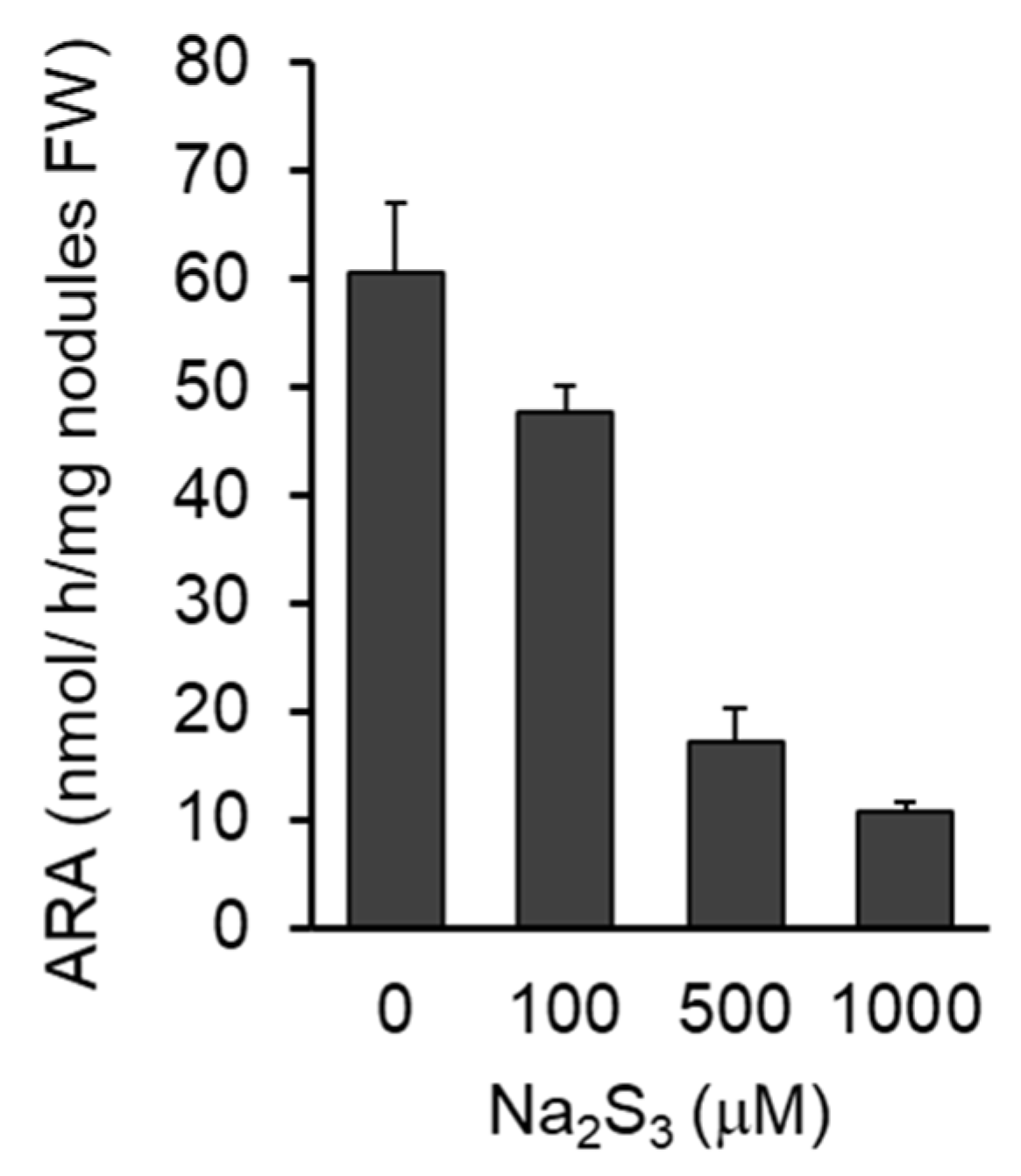
3.5. Effects of RSS on Nitrogenase Activity of Nodules
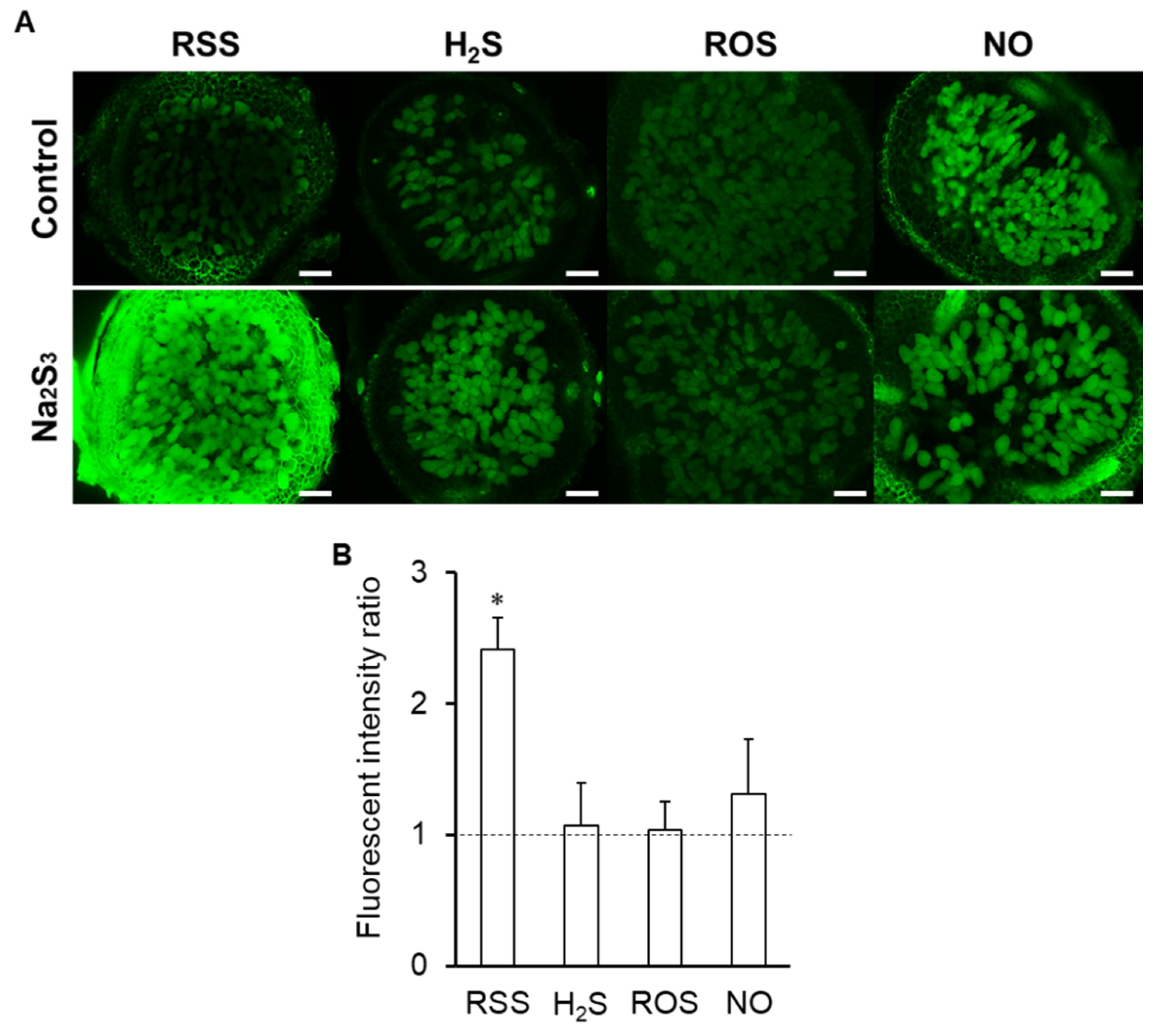
3.6. Effects of RSS on Signal Molecules in Infected Cells
4. Discussion
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Besson-Bard, A.; Pugin, A.; Wendehenne, D. New Insights into Nitric Oxide Signaling in Plants. Annu. Rev. Plant Boil. 2008, 59, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.A. ROS in biotic interactions. Physiol. Plant. 2010, 138, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, A.; Kopriva, S. Hydrogen Sulfide in Plants: From Dissipation of Excess Sulfur to Signaling Molecule. Nitric Oxide 2014, 41, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Hancock, J.; Whiteman, M. Hydrogen Sulfide and Cell Signaling: Team Player or Referee? Plant Physiol. Biochem. 2014, 78, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Delledonne, M.; Zeier, J.; Marocco, A.; Lamb, C. Signal Interactions between Nitric Oxide and Reactive Oxygen Intermediates in the Plant Hypersensitive Disease Resistance Response. Proc. Natl. Acad. Sci. 2001, 98, 13454–13459. [Google Scholar] [CrossRef] [Green Version]
- Espey, M.G.; Miranda, K.M.; Thomas, U.D.; Xavier, S.; Citrin, D.; Vitek, M.P.; Wink, D.A. A Chemical Perspective on the Interplay between NO, Reactive Oxygen Species, and Reactive Nitrogen Oxide Species. Ann. New York Acad. Sci. 2002, 962, 195–206. [Google Scholar] [CrossRef]
- Kirsch, M.; De Groot, H. Formation of Peroxynitrite from Reaction of Nitroxyl Anion with Molecular Oxygen. J. Boil. Chem. 2002, 277, 13379–13388. [Google Scholar] [CrossRef] [Green Version]
- Grün, S.; Lindermayr, C.; Sell, S.; Durner, J. Nitric Oxide and Gene Regulation in Plants. J. Exp. Bot. 2006, 57, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Ahlfors, R.; Brosché, M.; Kollist, H.; Kangasjärvi, J. Nitric Oxide Modulates Ozone-Induced Cell Death, Hormone Biosynthesis and Gene Expression in Arabidopsis Thaliana. Plant J. 2009, 58, 1–12. [Google Scholar] [CrossRef]
- Moreau, M.; Lindermayr, C.; Durner, J.; Klessig, D.F. NO Synthesis and Signaling in Plants - where do we stand? Physiol. Plant. 2010, 138, 372–383. [Google Scholar] [CrossRef]
- García-Mata, C.; LaMattina, L. Hydrogen Sulphide, a Novel Gasotransmitter Involved in Guard Cell Signalling. New Phytol. 2010, 188, 977–984. [Google Scholar] [CrossRef]
- Zhang, H.; Jones, R.L.; Tan, Z.-Q.; Hu, L.-Y.; Wang, S.-H.; Luo, J.-P. Hydrogen Sulfide Alleviates Aluminum Toxicity in Germinating Wheat Seedlings. J. Integr. Plant Boil. 2010, 52, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-G.; Yang, S.-Z.; Long, W.-B.; Yang, G.-X.; Shen, Z.-Z. Hydrogen Sulphide may be a Novel Downstream Signal Molecule in Nitric Oxide-Induced Heat Tolerance of Maize ( Zea mays L.) Seedlings. Plant, Cell Environ. 2013, 36, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Scuffi, D.; Álvarez, C.; LaSpina, N.; Gotor, C.; LaMattina, L.; García-Mata, C. Hydrogen Sulfide Generated by L-Cysteine Desulfhydrase Acts Upstream of Nitric Oxide to Modulate Abscisic Acid-Dependent Stomatal Closure. Plant Physiol. 2014, 166, 2065–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanatsiou, M.; Scuffi, D.; Blatt, M.R.; García-Mata, C. Hydrogen Sulfide Regulates Inward-Rectifying K+ Channels in Conjunction with Stomatal Closure. Plant Physiol. 2015, 168, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Hancock, J.T.; Whiteman, M. Hydrogen Sulfide Signaling: Interactions with Nitric Oxide and Reactive Oxygen Species. Ann N. Y. Acad. Sci. 2016, 1365, 5–14. [Google Scholar] [CrossRef]
- De Gara, L.; De Pinto, M.C.; Tommasi, F. The Antioxidant Systems Vis-à-vis Reactive Oxygen Species during Plant–Pathogen Interaction. Plant Physiol. Biochem. 2003, 41, 863–870. [Google Scholar] [CrossRef]
- Bellin, D.; Asai, S.; Delledonne, M.; Yoshioka, H. Nitric Oxide as a Mediator for Defense Responses. Mol. Plant-Microbe Interactions 2013, 26, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Mur, L.A.J.; Hebelstrup, K.H.; Gupta, K.J. Striking a Balance: Does Nitrate Uptake and Metabolism Regulate both NO Generation and Scavenging? Front. Plant Sci. 2013, 4, 288. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Loake, G.J.; Chu, C. Cross-Talk of Nitric Oxide and Reactive Oxygen Species in Plant Programed Cell Death. Front. Plant Sci. 2013, 4, 314. [Google Scholar] [CrossRef] [Green Version]
- Pauly, N.; Pucciariello, C.; Mandon, K.; Innocenti, G.; Jamet, A.; Baudouin, E.; Hérouart, D.; Frendo, P.; Puppo, A. Reactive Oxygen and Nitrogen Species and Glutathione: Key Players in the Legume-Rhizobium Symbiosis. J. Exp. Bot. 2006, 57, 1769–1776. [Google Scholar] [CrossRef] [Green Version]
- Sasakura, F.; Uchiumi, T.; Shimoda, Y.; Suzuki, A.; Takenouchi, K.; Higashi, S.; Abe, M. A Class 1 Hemoglobin Gene fromAlnus firmaFunctions in Symbiotic and Nonsymbiotic Tissues to Detoxify Nitric Oxide. Mol. Plant-Microbe Interact. 2006, 19, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Puppo, A.; Pauly, N.; Boscari, A.; Mandon, K.; Brouquisse, R. Hydrogen Peroxide and Nitric Oxide: Key Regulators of the Legume—Rhizobium and Mycorrhizal Symbioses. Antioxid. Redox Signal. 2013, 18, 2202–2219. [Google Scholar] [CrossRef]
- Damiani, I.; Pauly, N.; Puppo, A.; Brouquisse, R.; Boscari, A. Reactive Oxygen Species and Nitric Oxide Control Early Steps of the Legume—Rhizobium Symbiotic Interaction. Front. Plant Sci. 2016, 7, 454. [Google Scholar] [CrossRef]
- Del Giudice, J.; Cam, Y.; Damiani, I.; Fung-Chat, F.; Meilhoc, E.; Bruand, C.; Brouquisse, R.; Puppo, A.; Boscari, A. Nitric Oxide Is Required for an Optimal Establishment of the Medicago Truncatula–Sinorhizobium Meliloti Symbiosis. New Phytol. 2011, 191, 405–417. [Google Scholar] [CrossRef] [Green Version]
- Trinchant, J.-C.; Rigaud, J. Nitrite and Nitric Oxide as Inhibitors of Nitrogenase from Soybean Bacteroids. Appl. Environ. Microbiol. 1982, 44, 1385–1388. [Google Scholar] [CrossRef] [Green Version]
- Fukudome, M.; Calvo-Begueria, L.; Kado, T.; Osuki, K.-I.; Rubio, M.C.; Murakami, E.-I.; Nagata, M.; Kucho, K.-I.; Sandal, N.; Stougaard, J.; et al. Hemoglobin LjGlb1-1 Is Involved in Nodulation and Regulates the Level of Nitric Oxide in the Lotus Japonicus-Mesorhizobium Loti Symbiosis. J. Exp. Bot. 2016, 67, 5275–5283. [Google Scholar] [CrossRef] [Green Version]
- Shimoda, Y.; Shimoda-Sasakura, F.; Kanamori, N.; Nagata, M.; Suzuki, A.; Abe, M.; Higashi, S.; Uchiumi, T.; Kucho, K.-I. Overexpression of Class 1 Plant Hemoglobin Genes Enhances Symbiotic Nitrogen Fixation Activity between Mesorhizobium lotiand Lotus japonicus. Plant J. 2009, 57, 254–263. [Google Scholar] [CrossRef]
- Meilhoc, E.; Cam, Y.; Skapski, A.; Bruand, C. The Response to Nitric Oxide of the Nitrogen-Fixing Symbiont Sinorhizobium meliloti. Mol. Plant-Microbe Interact. 2010, 23, 748–759. [Google Scholar] [CrossRef] [Green Version]
- Cam, Y.; Pierre, O.; Boncompagni, E.; Hérouart, D.; Meilhoc, E.; Bruand, C. Nitric oxide (NO): A Key Player in the Senescence of Medicago Truncatula Root Nodules. New Phytol. 2012, 196, 548–560. [Google Scholar] [CrossRef]
- Fukudome, M.; Watanabe, E.; Osuki, K.; Imaizumi, R.; Aoki, T.; Becana, M.; Uchiumi, T. Stably-Transformed Lotus Japonicus Plants Overexpressing Phytoglobin Ljglb1-1 Show Decreased Nitric Oxide Levels in Roots and Nodules as well as Delayed Nodule Senescence. Plant Cell Physiol. 2019, 60, 816–825. [Google Scholar] [CrossRef]
- Peleg-Grossman, S.; Melamed-Book, N.; Levine, A. ROS Production during Symbiotic Infection Suppresses Pathogenesis-Related Gene Expression. Plant Signal. Behav. 2012, 7, 409–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthikala, M.-K.; Montiel, J.; Sánchez-López, R.; Nava, N.; Cárdenas, L.; Quinto, C. Respiratory Burst Oxidase Homolog Gene A Is Crucial for Rhizobium Infection and Nodule Maturation and Function in Common Bean. Front. Plant Sci. 2017, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Hanyu, M.; Fujimoto, H.; Tejima, K.; Saeki, K. Functional Differences of Two Distinct Catalases in Mesorhizobium loti MAFF303099 under Free-Living and Symbiotic Conditions. J Bacteriol. 2009, 191, 1463–1471. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Tang, J.; Liu, X.-P.; Wang, Y.; Yu, W.; Peng, W.-Y.; Fang, F.; Ma, D.-F.; Wei, Z.-J.; Hu, L.-Y. Hydrogen Sulfide Promotes Root Organogenesis in Ipomoea batatas, Salix matsudana and Glycine Max. J. Integr. Plant Boil. 2009, 51, 1086–1094. [Google Scholar] [CrossRef]
- Laureano-Marín, A.M.; Moreno, I.; Romero, L.C.; Gotor, C. Negative Regulation of Autophagy by Sulfide Is Independent of Reactive Oxygen Species1. Plant Physiol. 2016, 171, 1378–1391. [Google Scholar]
- Li, Z.-G.; Min, X.; Zhou, Z.-H. Hydrogen Sulfide: A Signal Molecule in Plant Cross-Adaptation. Front. Plant Sci. 2016, 7, 334. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.; Zhang, N.-N.; Pan, Q.; Zhang, J.-H.; Chen, J.; Wei, G.-H. Hydrogen Sulfide Promotes Nodulation and Nitrogen Fixation in Soybean-Rhizobia Symbiotic System. Mol. Plant-Microbe Interact. 2019, 32, 972–985. [Google Scholar] [CrossRef]
- Gruhlke, M.C.; Slusarenko, A.J. The Biology of Reactive Sulfur Species (RSS). Plant Physiol. Biochem. 2012, 59, 98–107. [Google Scholar] [CrossRef]
- Aroca, A.; Gotor, C.; Romero, L.C. Hydrogen Sulfide Signaling in Plants: Emerging Roles of Protein Persulfidation. Front. Plant Sci. 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Nishida, M.; Sawa, T.; Kitajima, N.; Ono, K.; Inoue, H.; Ihara, H.; Motohashi, H.; Yamamoto, M.; Suematsu, M.; Kurose, H.; et al. Hydrogen Sulfide Anion Regulates Redox Signaling via Electrophile Sulfhydration. Nat. Methods 2012, 8, 714–724. [Google Scholar] [CrossRef]
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive Cysteine Persulfides and S-polythiolation Regulate Oxidative Stress and Redox Signaling. Proc. Natl. Acad. Sci. 2014, 111, 7606–7611. [Google Scholar] [CrossRef] [Green Version]
- Ono, K.; Akaike, T.; Sawa, T.; Kumagai, Y.; Wink, D.A.; Tantillo, D.J.; Hobbs, A.J.; Nagy, P.; Xian, M.; Lin, J.; et al. Redox Chemistry and Chemical Biology of H2S, Hydropersulfides, and Derived Species: Implications of Their Possible Biological Activity and Utility. Free. Radic. Boil. Med. 2014, 77, 82–94. [Google Scholar] [CrossRef] [Green Version]
- Flavin, M. Microbial Transsulfuration: The Mechanism of an Enzymatic Disulfide Elimination Reaction. J. Boil. Chem. 1962, 237, 768–777. [Google Scholar]
- Yamanishi, T.; Tuboi, S. The Mechanism of the L-Cystine Cleavage Reaction Catalyzed by Rat Liver γ-Cystathionase1. J. Biochem. 1981, 89, 1913–1921. [Google Scholar] [CrossRef]
- Stipanuk, M.H. Metabolism of Sulfur-Containing Amino Acids. Annu Rev Nutr. 1986, 6, 179–209. [Google Scholar] [CrossRef]
- Watanabè, M.; Osada, J.; Aratani, Y.; Kluckman, K.; Reddick, R.; Malinow, M.R.; Maeda, N. Mice Deficient in Cystathionine Beta-synthase: Animal Models for Mild and Severe Homocyst(e)inemia. Proc. Natl. Acad. Sci. 1995, 92, 1585–1589. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Jhee, K.-H.; Kruger, W.D. Production of the Neuromodulator H2S by Cystathionine β-Synthase via the Condensation of Cysteine and Homocysteine. J. Boil. Chem. 2004, 279, 52082–52086. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a Physiologic Vasorelaxant: Hypertension in Mice with Deletion of Cystathionine Gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef] [Green Version]
- Ishii, I.; Akahoshi, N.; Yamada, H.; Nakano, S.; Izumi, T.; Suematsu, M. Cystathionine γ-Lyase-Deficient Mice Require Dietary Cysteine to Protect against Acute Lethal Myopathy and Oxidative Injury*. J. Boil. Chem. 2010, 285, 26358–26368. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, H. The NO World for Plants: Achieving Balance in an Open System. Plant, Cell Environ. 2005, 28, 78–84. [Google Scholar] [CrossRef]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of Antioxidant Potential of Plants and its Relevance to Therapeutic Applications. Int. J. Boil. Sci. 2015, 11, 982–991. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, H.; Cohen, M.F. Biological Consilience of Hydrogen Sulfide and Nitric Oxide in Plants: Gases of Primordial Earth Linking Plant, Microbial and Animal Physiologies. Nitric Oxide 2016, 55, 91–100. [Google Scholar] [CrossRef]
- Nagata, M.; Murakami, E.-I.; Shimoda, Y.; Shimoda-Sasakura, F.; Kucho, K.-I.; Suzuki, A.; Abe, M.; Higashi, S.; Uchiumi, T. Expression of a Class 1 Hemoglobin Gene and Production of Nitric Oxide in Response to Symbiotic and Pathogenic Bacteria in Lotus japonicus. Mol. Plant-Microbe Interact. 2008, 21, 1175–1183. [Google Scholar] [CrossRef] [Green Version]
- Fåhraues, G. The Infection of Clover Root Hair by Nodule Bacteria Studied by a Single Glass Slide Technique. J. Gen. Microbiol. 1957, 16, 374–381. [Google Scholar]
- Kaneko, T.; Nakamura, Y.; Sato, S.; Asamizu, E.; Kato, T.; Sasamoto, S.; Watanabe, A.; Idesawa, K.; Ishikawa, A.; Kawashima, K.; et al. Complete Genome Structure of the Nitrogen-Fixing Symbiotic Bacterium Mesorhizobium loti. Curr. Neuropharmacol. 2000, 7, 331–338. [Google Scholar]
- Maekawa, T.; Maekawa-Yoshikawa, M.; Takeda, N.; Imaizumi-Anraku, H.; Murooka, Y.; Hayashi, M. Gibberellin Controls the Nodulation Signaling Pathway in Lotus japonicus. Plant J. 2009, 58, 183–194. [Google Scholar] [CrossRef]
- Małolepszy, A.; Urbański, D.F.; James, E.K.; Sandal, N.; Isono, E.; Stougaard, J.; Andersen, S.U. The Deubiquitinating Enzyme AMSH1 Is Required for Rhizobial Infection and Nodule Organogenesis in Lotus japonicus. Plant J. 2015, 83, 719–731. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B. Reactive Sulfur Species (RSS): Possible New Players in the Oxidative Metabolism of Plant Peroxisomes. Front. Plant Sci. 2015, 6, 116. [Google Scholar] [CrossRef] [Green Version]
- Cortese-Krott, M.M.; Fernandez, B.O.; Santos, J.L.; Mergia, E.; Grman, M.; Nagy, P.; Kelm, M.; Butler, A.; Feelisch, M. Nitrosopersulfide (SSNO (-)) Accounts for sustained NO Bioactivity of S-nitrosothiols Following Reaction with Sulfide. Redox Boil. 2014, 2, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Shen, J.; Edmonds, K.A.; Luebke, J.L.; Hickey, A.K.; Palmer, L.D.; Chang, F.-M.J.; Bruce, K.A.; Kehl-Fie, T.E.; Skaar, E.P.; et al. Sulfide Homeostasis and Nitroxyl Intersect via Formation of Reactive Sulfur Species in Staphylococcus aureus. mSphere 2017, 2, e00082-17. [Google Scholar] [CrossRef] [Green Version]
- Marutani, E.; Sakaguchi, M.; Chen, W.; Sasakura, K.; Liu, J.; Xian, M.; Hanaoka, K.; Nagano, T.; Ichinose, F. Cytoprotective effects of Hydrogen Sulfide-Releasing N-methyl-D-aspartate Receptor Antagonists Are Mediated by Intracellular Sulfane Sulfur. Med. Chem. Comm. 2014, 5, 1577–1583. [Google Scholar] [CrossRef] [Green Version]
- Misak, A.; Kurakova, L.; Goffa, E.; Brezova, V.; Grman, M.; Ondriasova, E.; Chovanec, M.; Ondrias, K. Sulfide (Na₂S) and Polysulfide (Na₂S₂) Interacting with Doxycycline Produce/Scavenge Superoxide and Hydroxyl Radicals and Induce/Inhibit DNA Cleavage. Molecules. 2019, 24, 1148. [Google Scholar] [CrossRef] [Green Version]
- Hérouart, D.; Baudouin, E.; Frendo, P.; Harrison, J.; Santos, R.; Jamet, A.; Van De Sype, G.; Touati, D.; Puppo, A. Reactive Oxygen Species, Nitric Oxide and Glutathione: A Key Role in the Establishment of the Legume–Rhizobium Symbiosis? Plant Physiol. Biochem. 2002, 40, 619–624. [Google Scholar] [CrossRef]
- Santos, R.; Hérouart, D.; Sigaud, S.; Touati, D.; Puppo, A. Oxidative Burst in Alfalfa-Sinorhizobium Meliloti Symbiotic Interaction. Mol. Plant-Microbe Interactions 2001, 14, 86–89. [Google Scholar] [CrossRef] [Green Version]
- Sigaud, S.; Becquet, V.; Frendo, P.; Puppo, A.; Hérouart, D. Differential Regulation of Two Divergent Sinorhizobium meliloti Genes for HPII-Like Catalases during Free-Living Growth and Protective Role of Both Catalases during Symbiosis. J. Bacteriol. 1999, 181, 2634–2639. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.; Herouart, D.; Puppo, A.; Touati, D. Critical Protective Role of Bacterial Superoxide Dismutase in Rhizobium-Legume Symbiosis. Mol. Microbiol. 2000, 38, 750–759. [Google Scholar] [CrossRef] [Green Version]
- Melo, P.M.; Silva, L.S.; Ribeiro, I.; Seabra, A.R.; Carvalho, H.G. Glutamine Synthetase is a Molecular Target of Nitric Oxide in Root Nodules of Medicago Truncatula and is Regulated by Tyrosine Nitration. Plant Physiol. 2011, 157, 1505–1517. [Google Scholar] [CrossRef] [Green Version]
- Maiti, D.; Sarkar, T.S.; Ghosh, S. Detection of S-Nitrosothiol and Nitrosylated Proteins in Arachis hypogaea Functional Nodule: Response of the Nitrogen Fixing Symbiont. PLOS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Navascués, J.; Perez-Rontome, C.; Gay, M.; Marcos, M.; Yang, F.; Walker, F.A.; Desbois, A.; Abian, J.; Becana, M. Leghemoglobin Green Derivatives with Nitrated Hemes Evidence Production of Highly Reactive Nitrogen Species during Aging of Legume Nodules. Proc. Natl. Acad. Sci. 2012, 109, 2660–2665. [Google Scholar] [CrossRef] [Green Version]
- Takata, T.; Ihara, H.; Hatano, N.; Tsuchiya, Y.; Akaike, T.; Watanabe, Y. Reactive Sulfur Species Inactivate Ca2+/Calmodulin-Dependent Protein Kinase IV via S-polysulfidation of Its Active-Site Cysteine Residue. Biochem J. 2017, 474, 2547–2562. [Google Scholar] [CrossRef] [PubMed]
- Van Brussel, A.A.N.; Planqué, K.; Quispel, A. The Wall of Rhizobium leguminosarum in Bacteroid and Free-living Forms. J. Gen. Microbiol. 1977, 101, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Mikuláss, K.R.; Nagy, K.; Bogos, B.; Szegletes, Z.; Kovács, E.; Farkas, A.; Váró, G.; Kondorosi, É.; Kereszt, A. Antimicrobial Nodule-Specific Cysteine-Rich Peptides Disturb the Integrity of Bacterial Outer and Inner Membranes and Cause Loss of Membrane Potential. Ann. Clin. Microbiol. Antimicrob. 2016, 15, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Ye, Z.; Zhao, J. RbrA, a Cyanobacterial rubrerythrin, Functions as a FNR-Dependent Peroxidase in Heterocysts in Protection of Nitrogenase from Damage by Hydrogen Peroxide in Anabaena sp. PCC 7120. Mol. Microbiol. 2007, 66, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Kanahama, K.; Kanayama, Y. Involvement of Nitric Oxide in the Inhibition of Nitrogenase Activity by Nitrate in Lotus Root Nodules. J. Plant Physiol. 2010, 167, 238–241. [Google Scholar] [CrossRef]
- Krusell, L.; Krause, K.; Ott, T.; Desbrosses, G.; Krämer, U.; Sato, S.; Nakamura, Y.; Tabata, S.; James, E.K.; Sandal, N.; et al. The Sulfate Transporter SST1 Is Crucial for Symbiotic Nitrogen Fixation in Lotus japonicus Root Nodules. Plant Cell 2005, 17, 1625–1636. [Google Scholar] [CrossRef] [Green Version]
- Becana, M.; Wienkoop, S.; Matamoros, M.A. Sulfur Transport and Metabolism in Legume Root Nodules. Front. Plant Sci. 2018, 9, 1434. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukudome, M.; Shimada, H.; Uchi, N.; Osuki, K.-i.; Ishizaki, H.; Murakami, E.-i.; Kawaguchi, M.; Uchiumi, T. Reactive Sulfur Species Interact with Other Signal Molecules in Root Nodule Symbiosis in Lotus japonicus. Antioxidants 2020, 9, 145. https://doi.org/10.3390/antiox9020145
Fukudome M, Shimada H, Uchi N, Osuki K-i, Ishizaki H, Murakami E-i, Kawaguchi M, Uchiumi T. Reactive Sulfur Species Interact with Other Signal Molecules in Root Nodule Symbiosis in Lotus japonicus. Antioxidants. 2020; 9(2):145. https://doi.org/10.3390/antiox9020145
Chicago/Turabian StyleFukudome, Mitsutaka, Hazuki Shimada, Nahoko Uchi, Ken-ichi Osuki, Haruka Ishizaki, Ei-ichi Murakami, Masayoshi Kawaguchi, and Toshiki Uchiumi. 2020. "Reactive Sulfur Species Interact with Other Signal Molecules in Root Nodule Symbiosis in Lotus japonicus" Antioxidants 9, no. 2: 145. https://doi.org/10.3390/antiox9020145