Post-Treatment with Erinacine A, a Derived Diterpenoid of H. erinaceus, Attenuates Neurotoxicity in MPTP Model of Parkinson’s Disease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
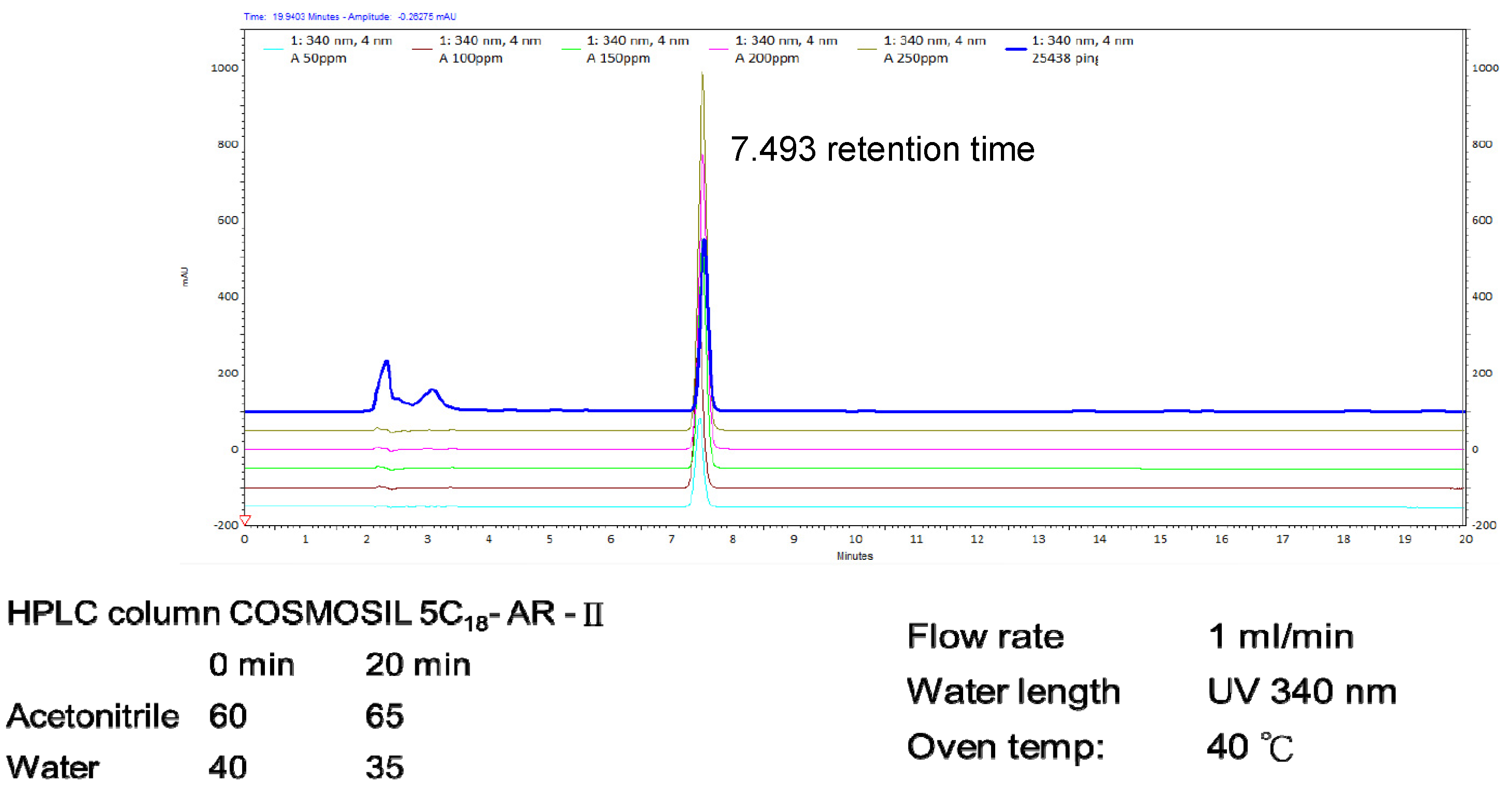
2.1. Hericium erinaceus Extracts and Analysis of Erinacine A
2.2. Animals
2.3. Chemical Reagents and Antibodies
2.4. Immunohistochemistry (IHC)
2.5. Cell Culture
2.6. Assessment of Cell Viability and Apoptosis Assay
2.7. Preparation of Total Cell Extracts and Immunoblotting Analysis
2.8. Statistical Analyses
3. Results
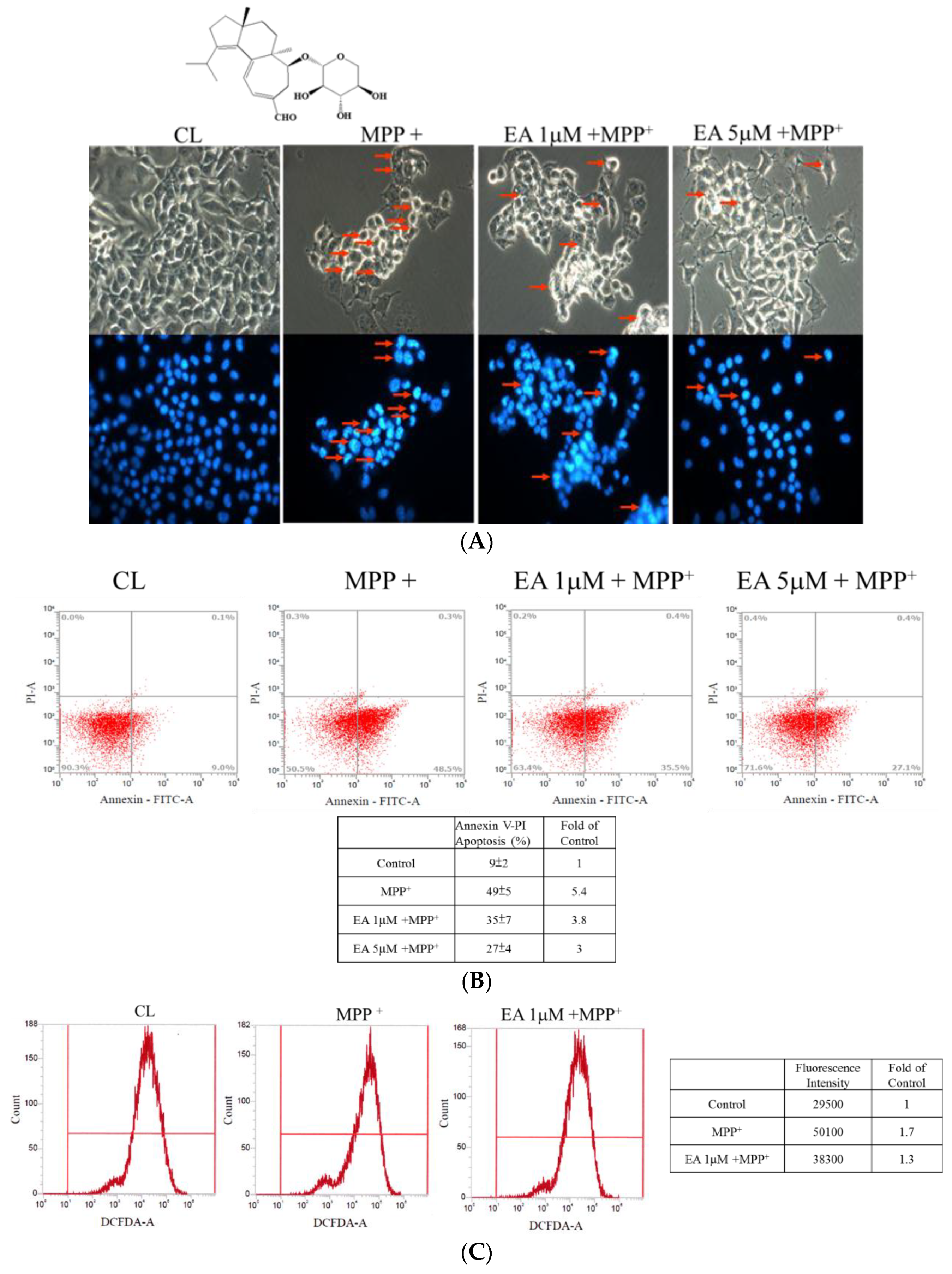
3.1. Erinacine A Prevention of MPP+-Induced Cell Death and ROS Generation of N2a Cells
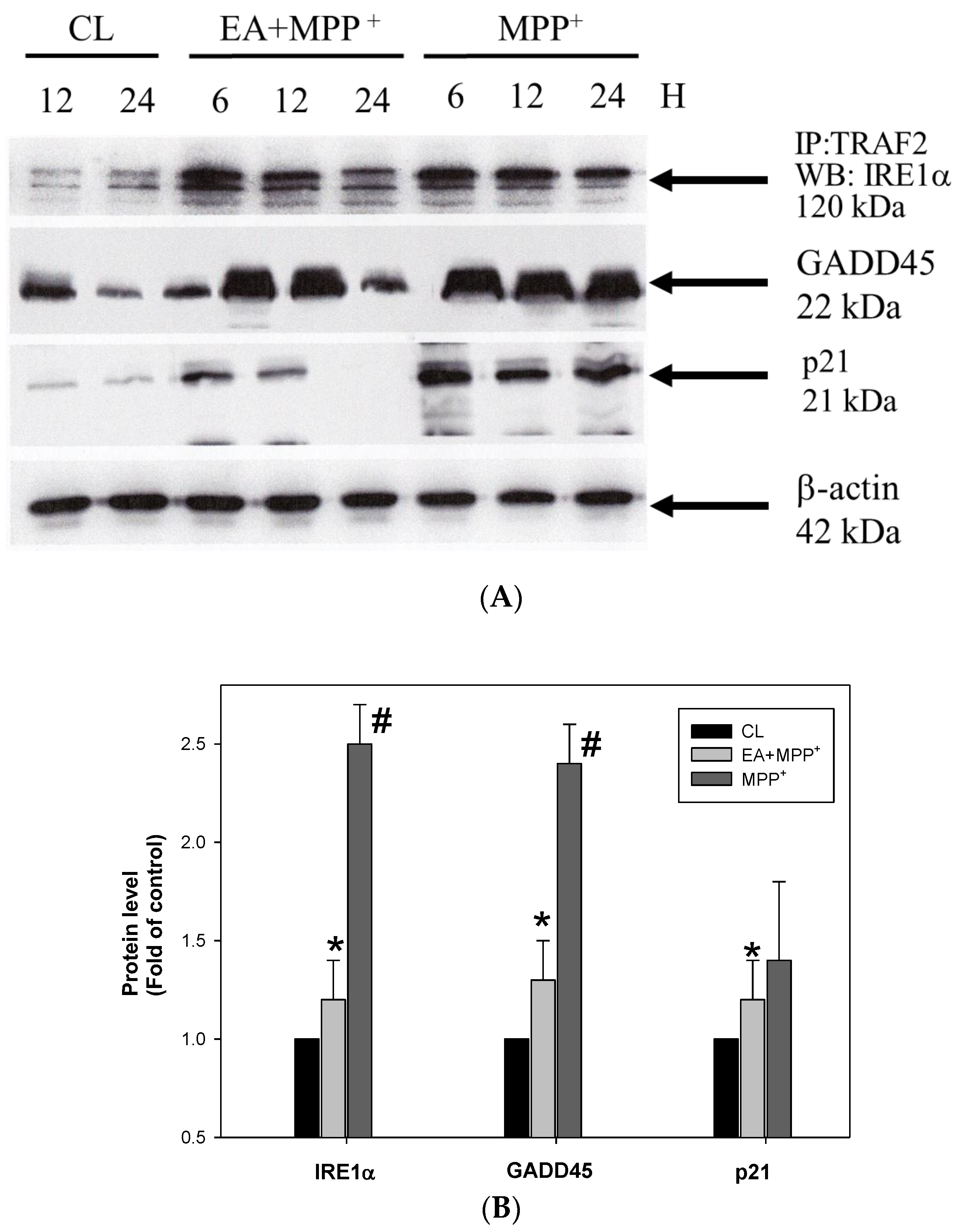
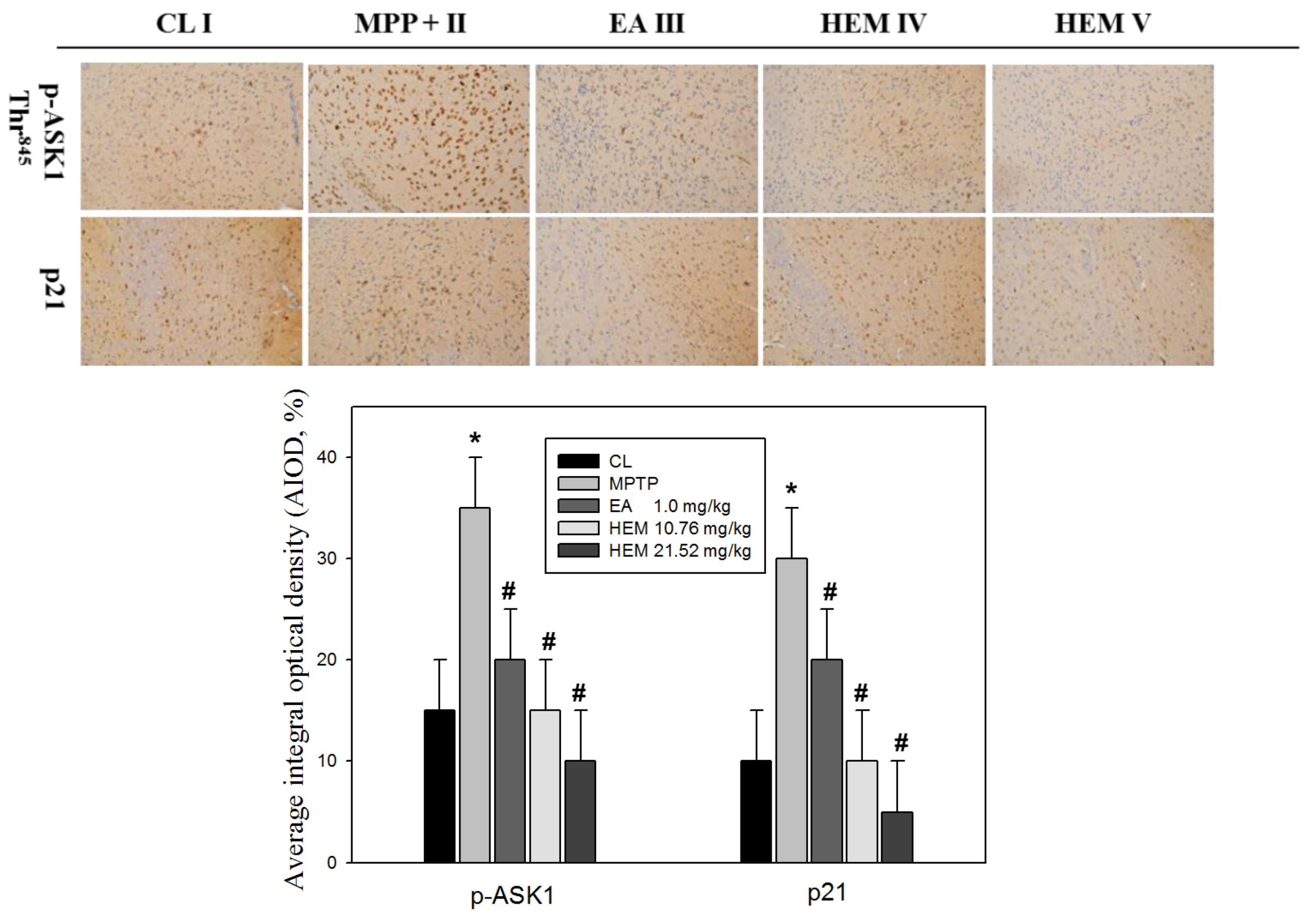
3.2. Erinacine A Reduction of MMP+ and Increased Interaction of TRAF2 and IRE1, as well as Expression of GADD45 and p21
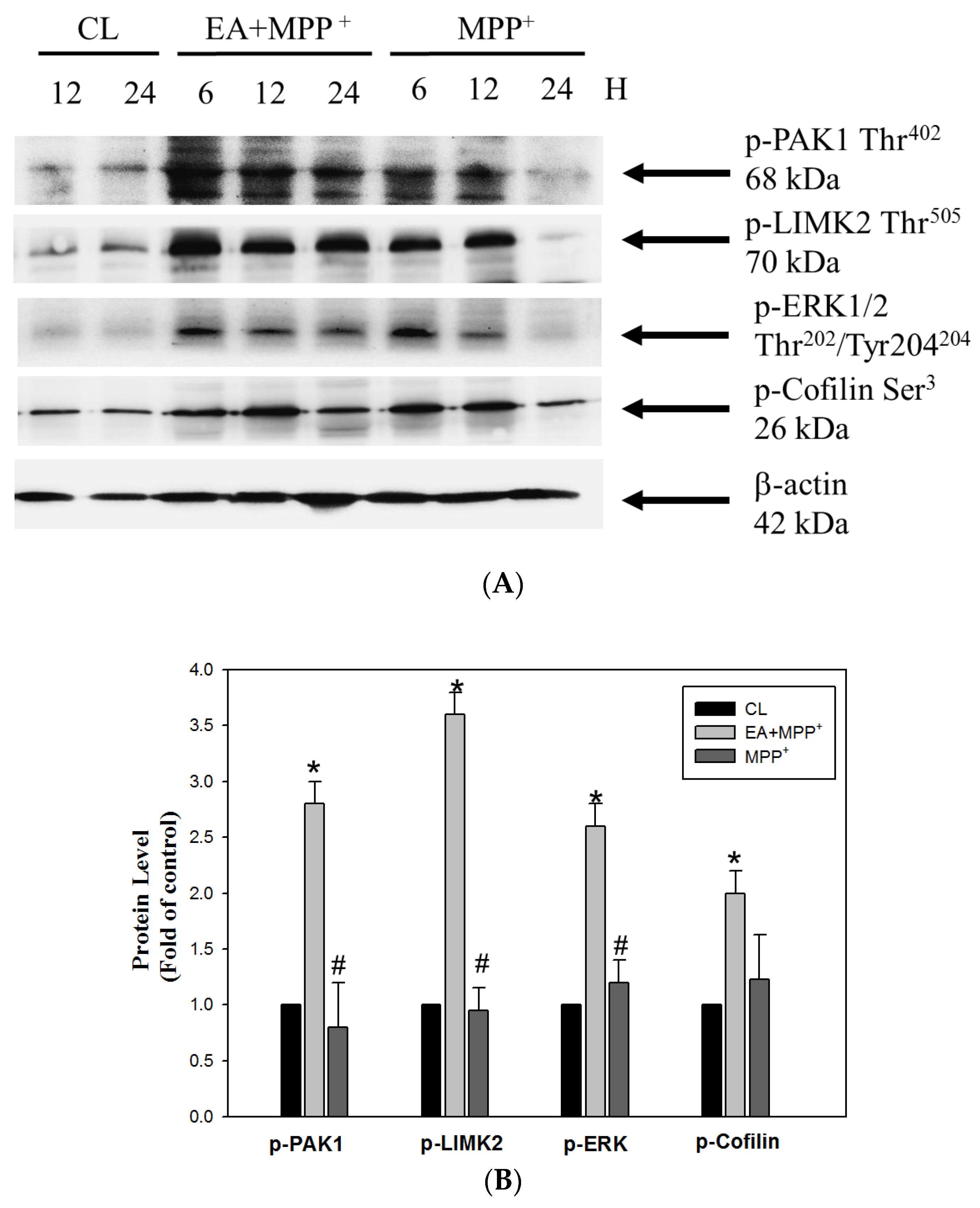
3.3. Erinacine A Enhanced Phosphorylation of PAK1/LIMK2/MEK/Cofilin under MPP+ Challenge
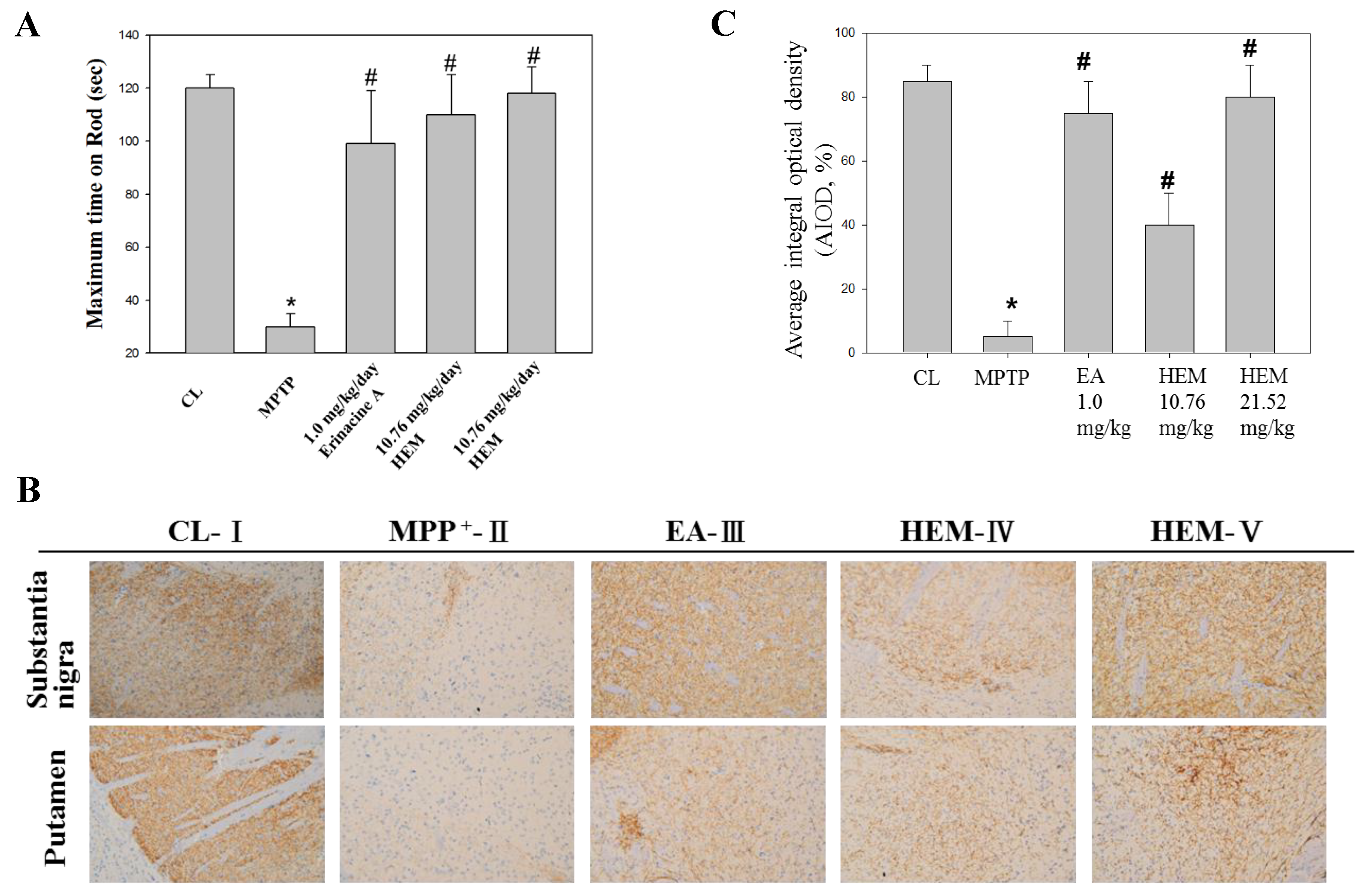
3.4. Post-Treatment with H. Erinaceus Mycelium and Its Structural Analog Erinacine A Weakened MPTP-Induced Neurotoxins and Movement Deficits in Mice
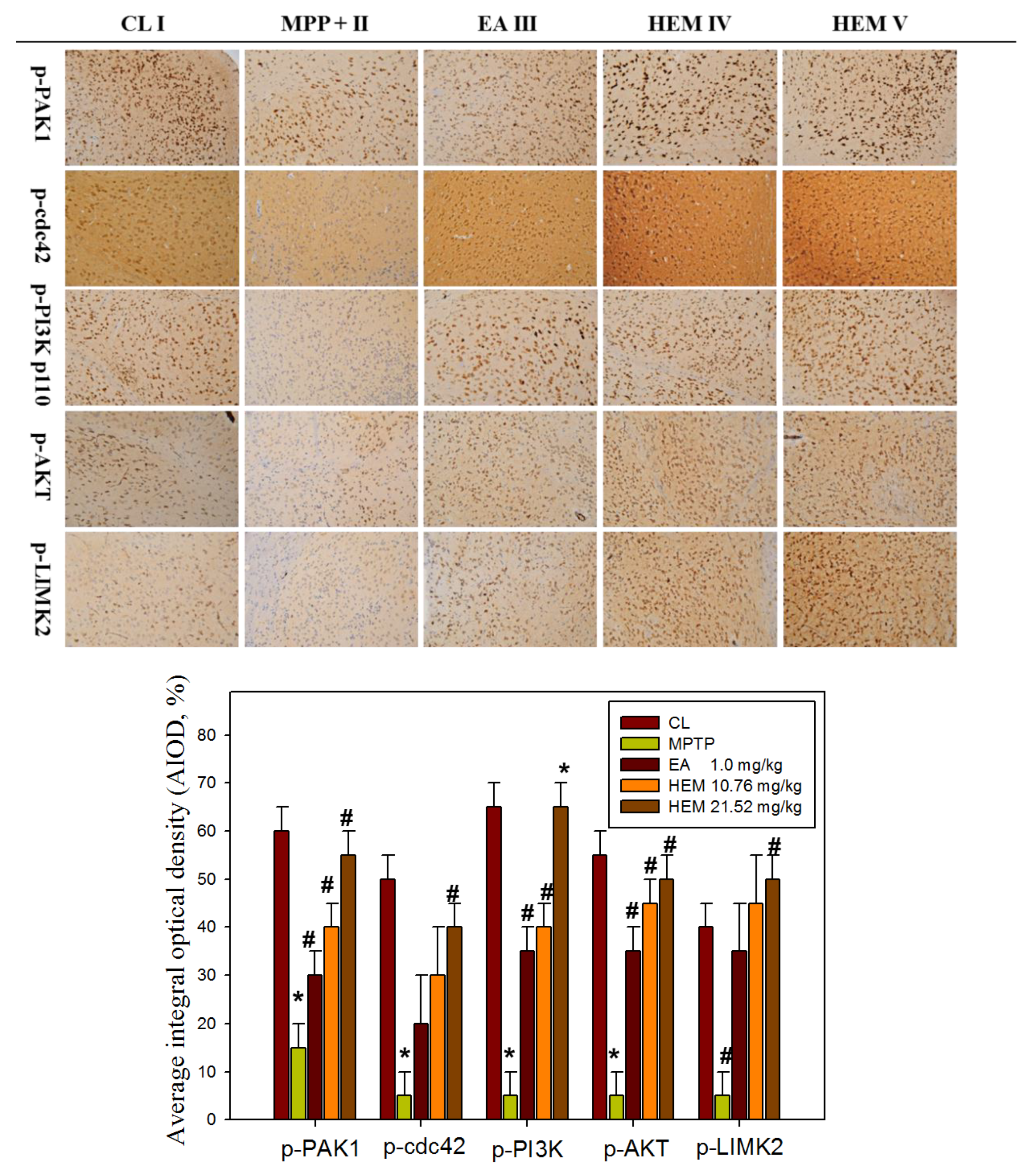
3.5. Post-Treatment with Erinacine A Reversed the PAK1/AKT/LIMK2/ Pathways in MPTP-Treated Animals
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Ethics Statement
Conflicts of Interest
Consent for Publication
Abbreviations
| HEM | Hericium erinaceus mycelium |
| MPTP | 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| ROS | Reactive oxygen species |
| PD | Parkinson’s disease |
| RNS | Reactive nitrogen species |
| DA | Dopamine |
| AD | Alzheimer’s disease |
| HD | Huntington’s disease |
| LIMK2 | LIM domain kinase 2 |
| ER | Endoplasmic reticulum |
References
- Kieburtz, K.; Wunderle, K.B. Parkinson’s Disease: Evidence for Environmental Risk Factors. Mov. Disord. 2013, 28, 8–13. [Google Scholar] [CrossRef]
- Olanow, C.W.; Tatton, W.G. Etiology and Pathogenesis of Parkinson’s Disease. Annu. Rev. Neurosci. 1999, 22, 123–144. [Google Scholar] [CrossRef] [Green Version]
- Salawu, F.; Olokoba, A.; Danburam, A. Current Management of Parkinson’s Disease. Ann. Afr. Med. 2010, 9, 55–61. [Google Scholar] [CrossRef]
- Schulz, J.B.; Falkenburger, B.H. Neuronal Pathology in Parkinson’s Disease. Cell Tissue Res. 2004, 318, 135–147. [Google Scholar] [CrossRef]
- Singer, T.P.; Ramsay, R.R. Mechanism of the Neurotoxicity of MPTP: An update. FEBS Lett. 1990, 274, 1–8. [Google Scholar]
- Zawada, W.M.; Banninger, G.P.; Thornton, J.; Marriott, B.; Cantu, D.; Rachubinski, A.L.; Das, M.; Griffin, W.S.; Jones, S.M. Generation of Reactive Oxygen Species in 1-methyl-4-phenylpyridinium (mpp+) Treated Dopaminergic Neurons Occurs as an Nadph Oxidase-Dependent Two-Wave Cascade. J. Neuroinflamm. 2011, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Dias, V.; Junn, E.; Mouradian, M.M. The Role of Oxidative Stress in Parkinson’s Disease. J. Park. Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef] [Green Version]
- Rizor, A.; Pajarillo, E.; Johnson, J.; Aschner, M.; Lee, E. Lee Astrocytic Oxidative/Nitrosative Stress Contributes to Parkinson’s Disease Pathogenesis: The Dual Role of Reactive Astrocytes. Antioxidants 2019, 8, 265. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Kanthasamy, A.; Ghosh, A.; Anantharam, V.; Kalyanaraman, B.; Kanthasamy, A.G. Mitochondria-Targeted Antioxidants for Treatment of Parkinson’s Disease: Preclinical and Clinical Outcomes. Biochim. Biophys. Acta 2014, 1842, 1282–1294. [Google Scholar] [CrossRef] [Green Version]
- Urano, F.; Wang, X.; Bertolotti, A.; Zhang, Y.; Chung, P.; Harding, H.P.; Ron, D. Coupling of Stress in the ER to Activation of JNK Protein Kinases by Transmembrane Protein Kinase IRE1. Science 2000, 287, 664–666. [Google Scholar] [CrossRef] [Green Version]
- Arias-Romero, L.E.; Chernoff, J. A Tale of Two Paks. Boil. Cell 2008, 100, 97–108. [Google Scholar] [CrossRef]
- Tang, Y.; Zhou, H.; Chen, A.; Pittman, R.N.; Field, J. The Akt Proto-Oncogene Links Ras to Pak and Cell Survival Signals. J. Boil. Chem. 2000, 275, 9106–9109. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, J.S.; Wu, C.-C.; Redwine, J.M.; Comery, T.A.; Arias, R.; Bowlby, M.; Martone, R.; Morrison, J.H.; Pangalos, M.N.; Reinhart, P.H.; et al. Early-Onset Behavioral and Synaptic Deficits in a Mouse Model of Alzheimer’s Disease. Proc. Natl. Acad. Sci. 2006, 103, 5161–5166. [Google Scholar] [CrossRef] [Green Version]
- Kreis, P.; Barnier, J.-V. PAK Signalling in Neuronal Physiology. Cell. Signal. 2009, 21, 384–393. [Google Scholar] [CrossRef]
- Zhou, G.-L.; Zhuo, Y.; King, C.C.; Fryer, B.H.; Bokoch, G.M.; Field, J. Akt Phosphorylation of Serine 21 on Pak1 Modulates Nck Binding and Cell Migration. Mol. Cell. Boil. 2003, 23, 8058–8069. [Google Scholar] [CrossRef] [Green Version]
- Edwards, D.C.; Sanders, L.C.; Bokoch, G.M.; Gill, G.N. Activation of LIM-Kinase by Pak1 Couples Rac/Cdc42 GTPase Signalling to Actin Cytoskeletal Dynamics. Nature 1999, 1, 253–259. [Google Scholar] [CrossRef]
- Lee, K.-F.; Chen, J.-H.; Teng, C.-C.; Shen, C.-H.; Hsieh, M.-C.; Lu, C.-C.; Lee, K.-C.; Lee, L.-Y.; Chen, W.-P.; Chen, C.-C.; et al. Protective Effects of Hericium erinaceus Mycelium and Its Isolated Erinacine A against Ischemia-Injury-Induced Neuronal Cell Death via the Inhibition of iNOS/p38 MAPK and Nitrotyrosine. Int. J. Mol. Sci. 2014, 15, 15073–15089. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M. Chemistry, Nutrition, and Health-Promoting Properties of Hericium Erinaceus (Lion’s Mane) Mushroom Fruiting Bodies and Mycelia and Their Bioactive Compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef]
- Malinowska, E.; Krzyczkowski, W.; Herold, F.; Łapienis, G.; Slusarczyk, J.; Suchocki, P.; Kuraś, M.; Turło, J. Biosynthesis of Selenium-Containing Polysaccharides with Antioxidant Activity in Liquid Culture of Hericium Erinaceum. Enzym. Microb. Technol. 2009, 44, 334–343. [Google Scholar] [CrossRef]
- Yang, B.-K.; Park, J.-B.; Song, C.-H. Hypolipidemic Effect of an Exo-Biopolymer Produced from a Submerged Mycelial Culture of Hericium Erinaceus. Biosci. Biotechnol. Biochem. 2003, 67, 1292–1298. [Google Scholar] [CrossRef] [Green Version]
- Gong, M.; An, J.; Lü, H.-Z.; Wu, C.-F.; Li, Y.-J.; Cheng, J.-Q.; Bao, J.-K. Effects of Denaturation and Amino Acid Modification on Fluorescence Spectrum and Hemagglutinating Activity of Hericium Erinaceum Lectin. Acta Biochim. et Biophys. Sin. 2004, 36, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Yim, M.-H.; Shin, J.-W.; Son, J.-Y.; Oh, S.-M.; Han, S.-H.; Cho, J.-H.; Cho, C.-K.; Yoo, H.-S.; Lee, Y.-W.; Son, C.-G.; et al. Soluble Components of Hericium Erinaceum Induce NK Cell Activation via Production of Interleukin-12 in Mice Splenocytes. Acta Pharmacol. Sin. 2007, 28, 901–907. [Google Scholar] [CrossRef]
- Shimbo, M.; Kawagishi, H.; Yokogoshi, H. Erinacine A Increases Catecholamine and Nerve Growth Factor Content in the Central Nervous System of Rats. Nutr. Res. 2005, 25, 617–623. [Google Scholar] [CrossRef]
- Lee, K.C.; Kuo, H.C.; Shen, C.H.; Lu, C.C.; Huang, W.S.; Hsieh, M.C.; Huang, C.Y.; Kuo, Y.H.; Hsieh, Y.Y.; Teng, C.C.; et al. A Proteomics Approach to Identifying Novel Protein Targets Involved in Erinacine a-Mediated Inhibition of Colorectal Cancer Cells’ Aggressiveness. J. Cell. Mol. Med. 2017, 21, 588–599. [Google Scholar] [CrossRef] [Green Version]
- Kuo, H.-C.; Lu, C.-C.; Shen, C.-H.; Tung, S.-Y.; Hsieh, M.C.; Lee, K.-C.; Lee, L.-Y.; Chen, C.-C.; Teng, C.-C.; Huang, W.-S.; et al. Hericium Erinaceus Mycelium and its Isolated Erinacine A Protection from MPTP-Induced Neurotoxicity through the ER Stress, Triggering an Apoptosis Cascade. J. Transl. Med. 2016, 14, 78. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Webb, N.J.; Asmussen, H.; Niu, S.; Horwitz, A.F. A GIT1/PIX/Rac/PAK Signaling Module Regulates Spine Morphogenesis and Synapse Formation through MLC. J. Neurosci. 2005, 25, 3379–3388. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.-C.; Huang, W.-S.; Lee, K.-F.; Lee, K.-C.; Hsieh, M.-C.; Huang, C.-Y.; Lee, L.-Y.; Lee, B.-O.; Teng, C.-C.; Shen, C.-H.; et al. Inhibitory Effect of Erinacines A on the Growth of DLD-1 Colorectal Cancer Cells is Induced by Generation of Reactive Oxygen Species and Activation of p70S6K and p21. J. Funct. Foods 2016, 21, 474–484. [Google Scholar] [CrossRef]
- Tsai, J.-J.; Kuo, H.-C.; Lee, K.-F.; Tsai, T.-H. Proteomic Analysis of Plasma from Rats Following Total Parenteral Nutrition-Induced Liver Injury. Proteomics 2015, 15, 3865–3874. [Google Scholar] [CrossRef]
- Tseng, T.-H.; Shen, C.-H.; Huang, W.-S.; Chen, C.-N.; Liang, W.-H.; Lin, T.-H.; Kuo, H.-C. Activation of Neutral-Sphingomyelinase, MAPKs, and p75 NTR-Mediating Caffeic Acid Phenethyl Ester–Induced Apoptosis in C6 Glioma Cells. J. Biomed. Sci. 2014, 21, 61. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.-H.; Tung, S.-Y.; Huang, W.-S.; Lu, C.-C.; Lee, K.-C.; Hsieh, Y.-Y.; Chang, P.-J.; Liang, H.-F.; Chen, J.-H.; Lin, T.-H.; et al. Exploring the Effects of Tert-butylhydroperoxide Induced Liver Injury using Proteomic Approach. Toxicology 2014, 316, 61–70. [Google Scholar] [CrossRef]
- Huang, W.S.; Hsieh, M.C.; Huang, C.Y.; Kuo, Y.H.; Tung, S.Y.; Shen, C.H.; Hsieh, Y.Y.; Teng, C.C.; Lee, K.F.; Chen, T.C.; et al. The Association of Cxc Receptor 4 Mediated Signaling Pathway with Oxaliplatin-Resistant Human Colorectal Cancer Cells. PLoS ONE 2016, 11, e0159927. [Google Scholar] [CrossRef] [PubMed]
- Cannon, J.R.; Greenamyre, J.T. Neurotoxic in vivo Models of Parkinson’s Disease Recent Advances. Prog. Brain. Res. 2010, 184, 17–33. [Google Scholar]
- Venkatesan, R.; Ji, E.; Kim, S.Y. Phytochemicals That Regulate Neurodegenerative Disease by Targeting Neurotrophins: A Comprehensive Review. BioMed Res. Int. 2015, 2015, 1–22. [Google Scholar] [CrossRef]
- Kum, W.F.; Durairajan, S.S.K.; Bian, Z.X.; Man, S.C.; Lam, Y.C.; Xie, L.X.; Lu, J.H.; Wang, Y.; Huang, X.Z.; Li, M. Treatment of Idiopathic Parkinson’s Disease with Traditional Chinese Herbal Medicine: A Randomized Placebo-Controlled Pilot Clinical Study. Evidence-Based Complement. Altern. Med. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Li, I.-C.; Lee, L.-Y.; Tzeng, T.-T.; Chen, W.-P.; Chen, Y.-P.; Shiao, Y.-J.; Chen, C.-C. Neurohealth Properties of Hericium Erinaceus Mycelia Enriched with Erinacines. Behav. Neurol. 2018, 2018, 1–10. [Google Scholar]
- Mori, K.; Obara, Y.; Hirota, M.; Azumi, Y.; Kinugasa, S.; Inatomi, S.; Nakahata, N. Nerve Growth Factor-Inducing Activity of Hericium Erinaceus in 1321N1 Human Astrocytoma Cells. Boil. Pharm. Bull. 2008, 31, 1727–1732. [Google Scholar] [CrossRef] [Green Version]
- Kichina, J.V.; Goc, A.; Al-Husein, B.; Somanath, P.R.; Kandel, E.S. PAK1 as a Therapeutic Target. Expert Opin. Ther. Targets 2010, 14, 703–725. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wu, Z.; Gao, X.; Zhu, Q.; Jin, Y.; Wu, A.; Huang, A.C.J. Anchanling Reduces Pathology in a Lactacystin- Induced Parkinson’s Disease Model☆. Neural Regen. Res. 2012, 7, 165–170. [Google Scholar]
- Lin, Y.-R.; Chen, H.-H.; Ko, C.-H.; Chan, M.-H. Differential Inhibitory Effects of Honokiol and Magnolol on Excitatory Amino Acid-Evoked Cation Signals and NMDA-Induced Seizures. Neuropharmacology 2005, 49, 542–550. [Google Scholar] [CrossRef]
- Wang, M.; Gao, Y.; Xu, D.; Konishi, T.; Gao, Q. Hericium Erinaceus (Yamabushitake): A Unique Resource for Developing Functional Foods and Medicines. Food Funct. 2014, 5, 3055–3064. [Google Scholar] [CrossRef]
- Mori, K.; Inatomi, S.; Ouchi, K.; Azumi, Y.; Tuchida, T. Improving Effects of the Mushroom Yamabushitake (Hericium erinaceus) on Mild Cognitive Impairment: A Double-Blind Placebo-Controlled Clinical Trial. Phytotherapy Res. 2009, 23, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Kremer, L.; Taleb, O.; Boehm, N.; Mensah-Nyagan, A.G.; Trifilieff, E.; De Seze, J.; Brun, S. FTY720 Controls Disease Severity and Attenuates Sciatic Nerve Damage in Chronic Experimental Autoimmune Neuritis. J. Neuroinflamm. 2019, 16, 54. [Google Scholar] [CrossRef] [PubMed]
- Henrich, M.T.; Geibl, F.F.; Lee, B.; Chiu, W.H.; Koprich, J.B.; Brotchie, J.M.; Timmermann, L.; Decher, N.; Matschke, L.A.; Oertel, W.H. A53t-alpha-synuclein Overexpression in Murine Locus Coeruleus Induces Parkinson’s Disease-Like Pathology in Neurons and Glia. Acta Neuropathol. Commun. 2018, 6, 39. [Google Scholar] [CrossRef] [PubMed]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.-F.; Tung, S.-Y.; Teng, C.-C.; Shen, C.-H.; Hsieh, M.C.; Huang, C.-Y.; Lee, K.-C.; Lee, L.-Y.; Chen, W.-P.; Chen, C.-C.; et al. Post-Treatment with Erinacine A, a Derived Diterpenoid of H. erinaceus, Attenuates Neurotoxicity in MPTP Model of Parkinson’s Disease. Antioxidants 2020, 9, 137. https://doi.org/10.3390/antiox9020137
Lee K-F, Tung S-Y, Teng C-C, Shen C-H, Hsieh MC, Huang C-Y, Lee K-C, Lee L-Y, Chen W-P, Chen C-C, et al. Post-Treatment with Erinacine A, a Derived Diterpenoid of H. erinaceus, Attenuates Neurotoxicity in MPTP Model of Parkinson’s Disease. Antioxidants. 2020; 9(2):137. https://doi.org/10.3390/antiox9020137
Chicago/Turabian StyleLee, Kam-Fai, Shui-Yi Tung, Chih-Chuan Teng, Chien-Heng Shen, Meng Chiao Hsieh, Cheng-Yi Huang, Ko-Chao Lee, Li-Ya Lee, Wan-Ping Chen, Chin-Chu Chen, and et al. 2020. "Post-Treatment with Erinacine A, a Derived Diterpenoid of H. erinaceus, Attenuates Neurotoxicity in MPTP Model of Parkinson’s Disease" Antioxidants 9, no. 2: 137. https://doi.org/10.3390/antiox9020137