Spontaneous Formation of Melanin from Dopamine in the Presence of Iron

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and General Procedures
2.2. Preparation of Dopamine Solutions
2.3. UV-Vis Spectrophotometry and the 480 nm Assay
2.4. Animal Subjects
2.5. Brain Slice Preparation
2.6. Carbon Fiber Electrodes, Calibration, and Fast Scan Cyclic Voltammetry
2.7. NMR Studies
2.8. Statistical Analyses
3. Results
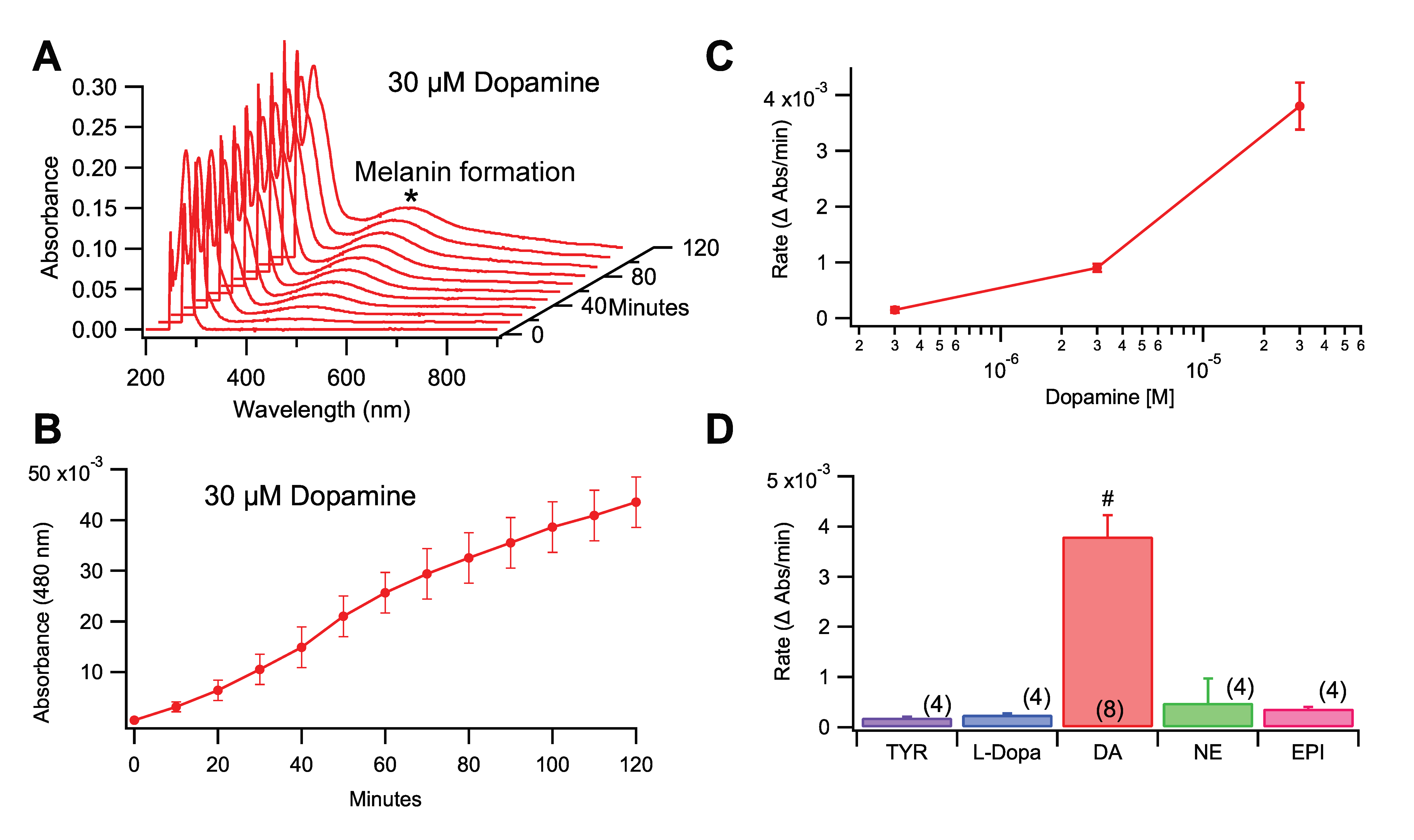
3.1. Role of Initial Substrate in Rate of Melanin Formation
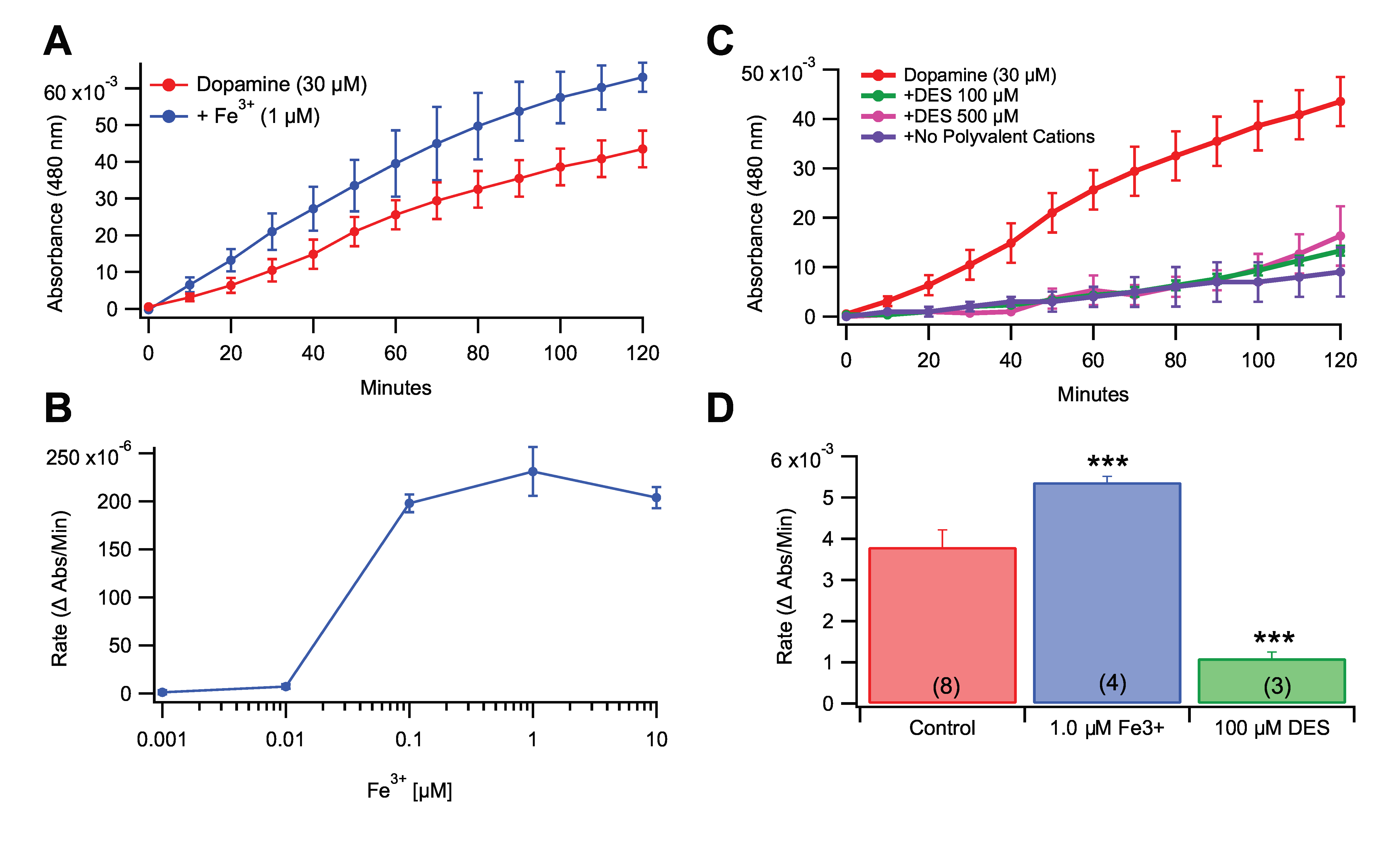
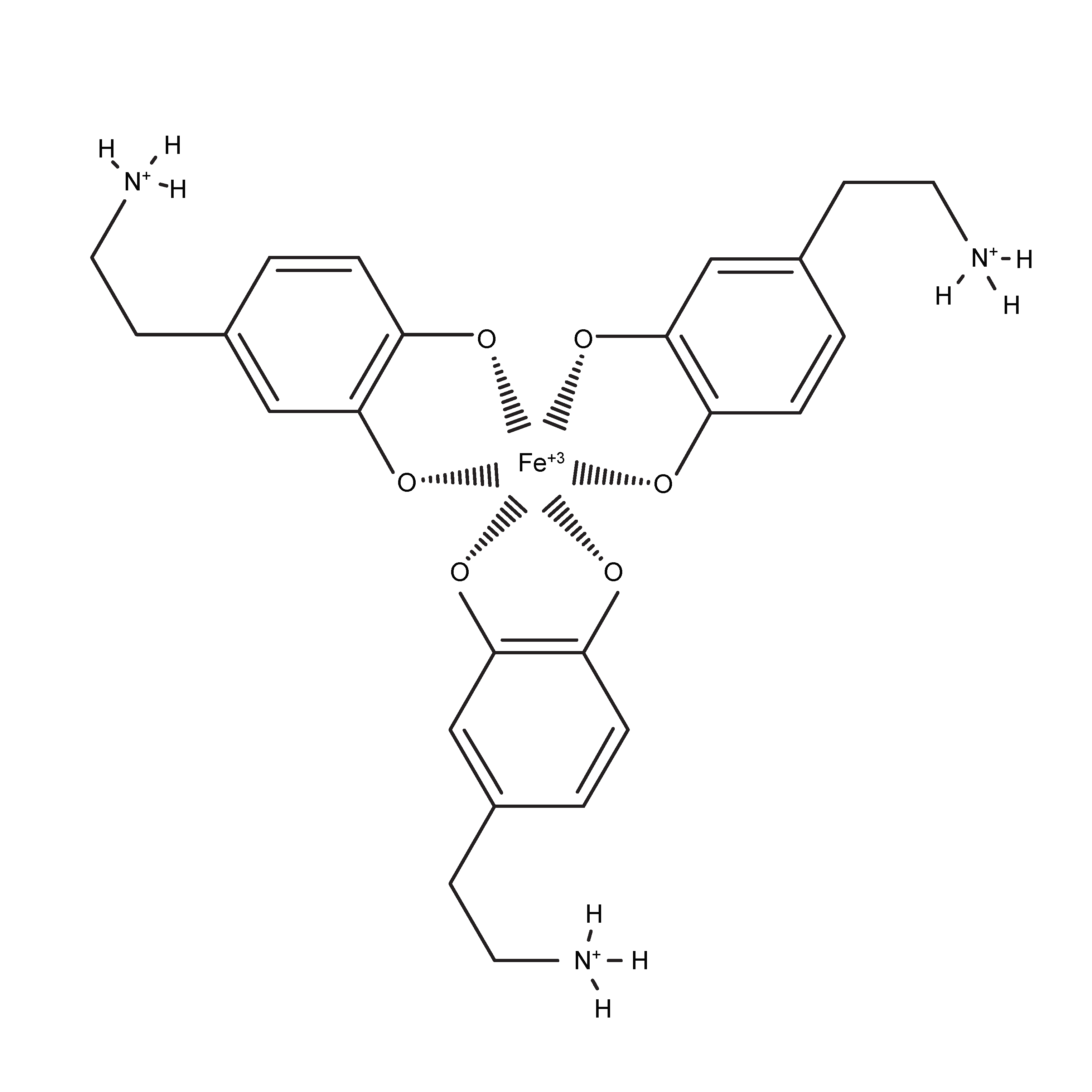
3.2. Role of Fe3+ in Rate of Dopamine Melanogenesis
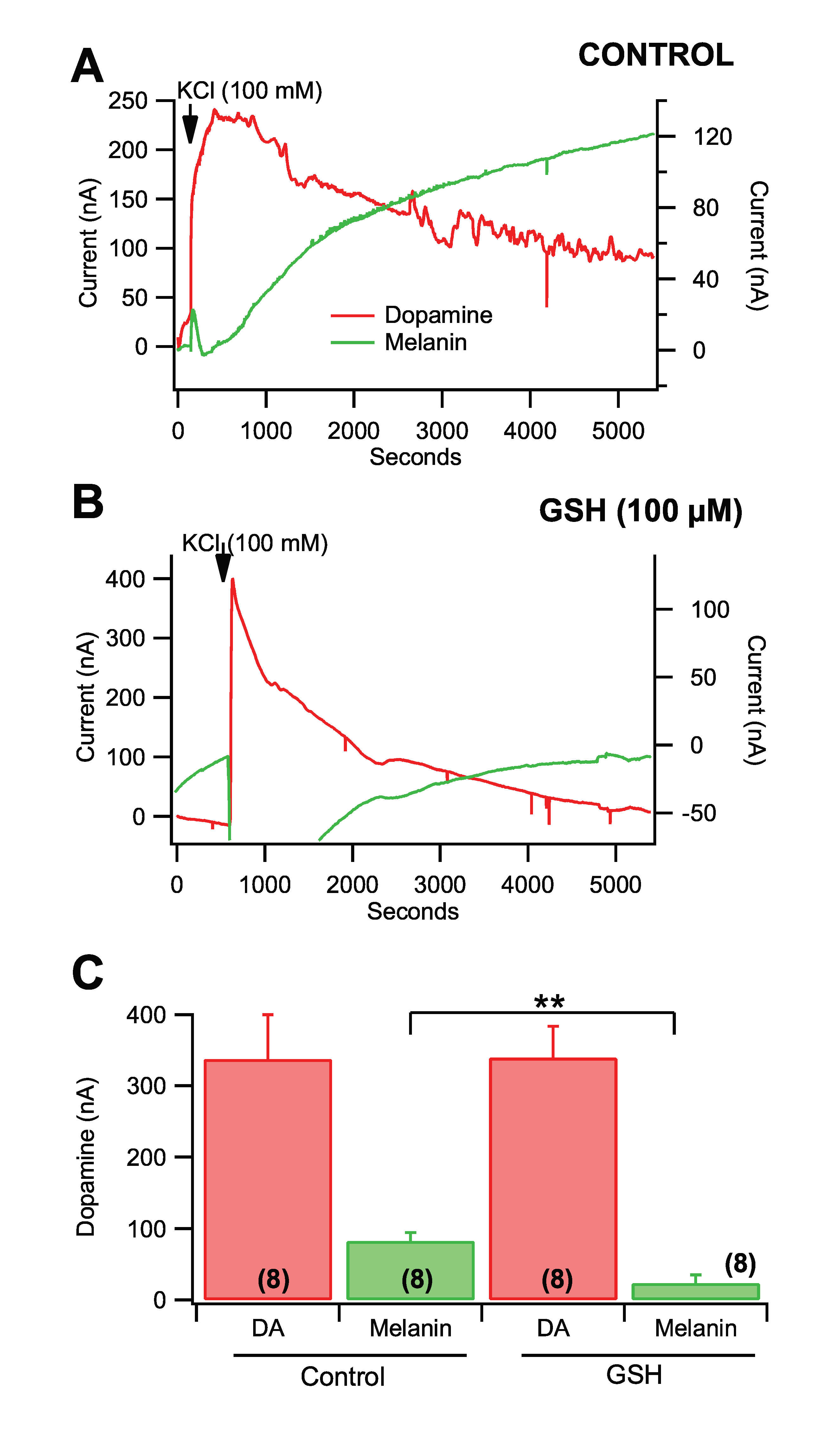
3.3. Role of Ca2+ in Rate of Dopamine Melanogenesis
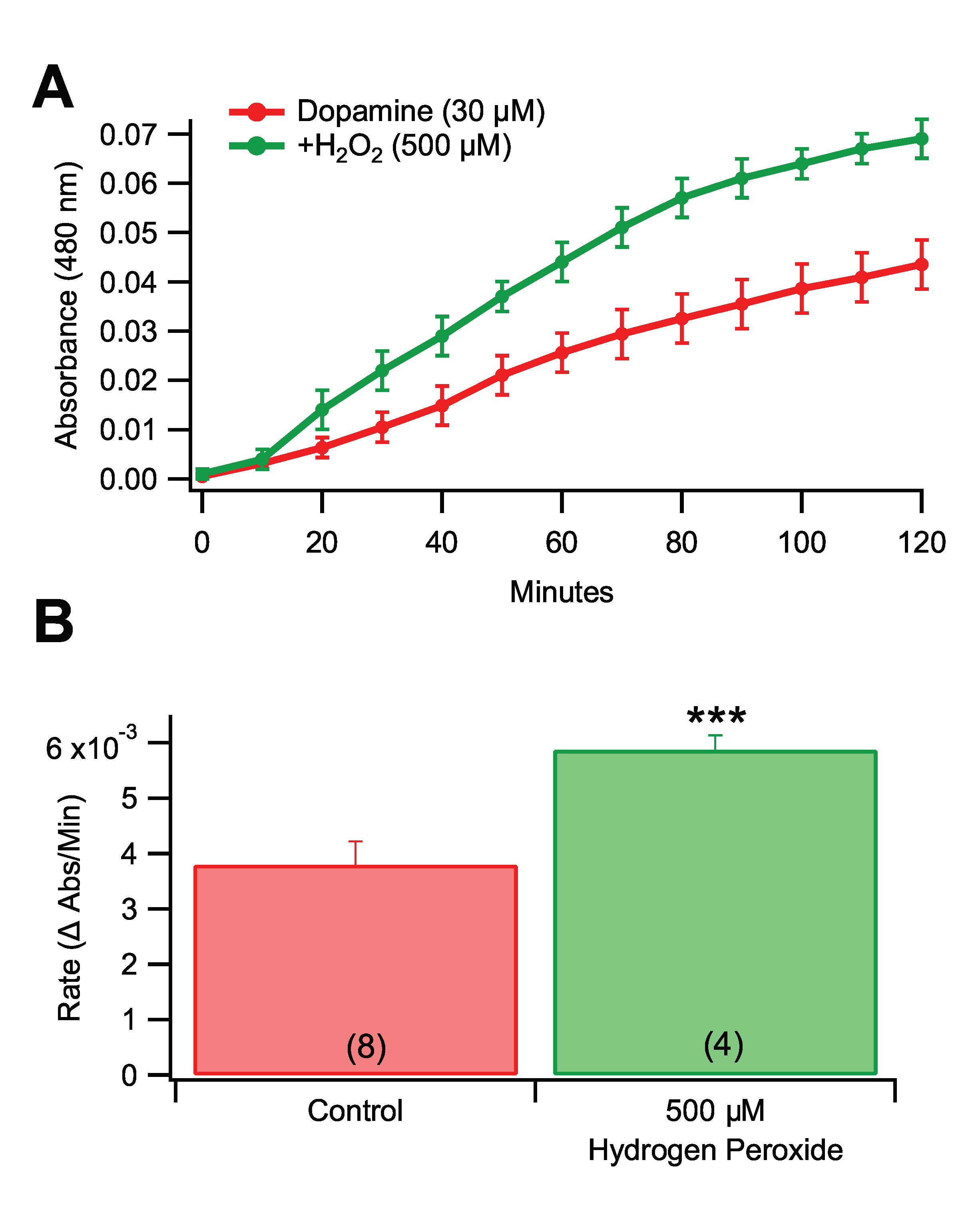
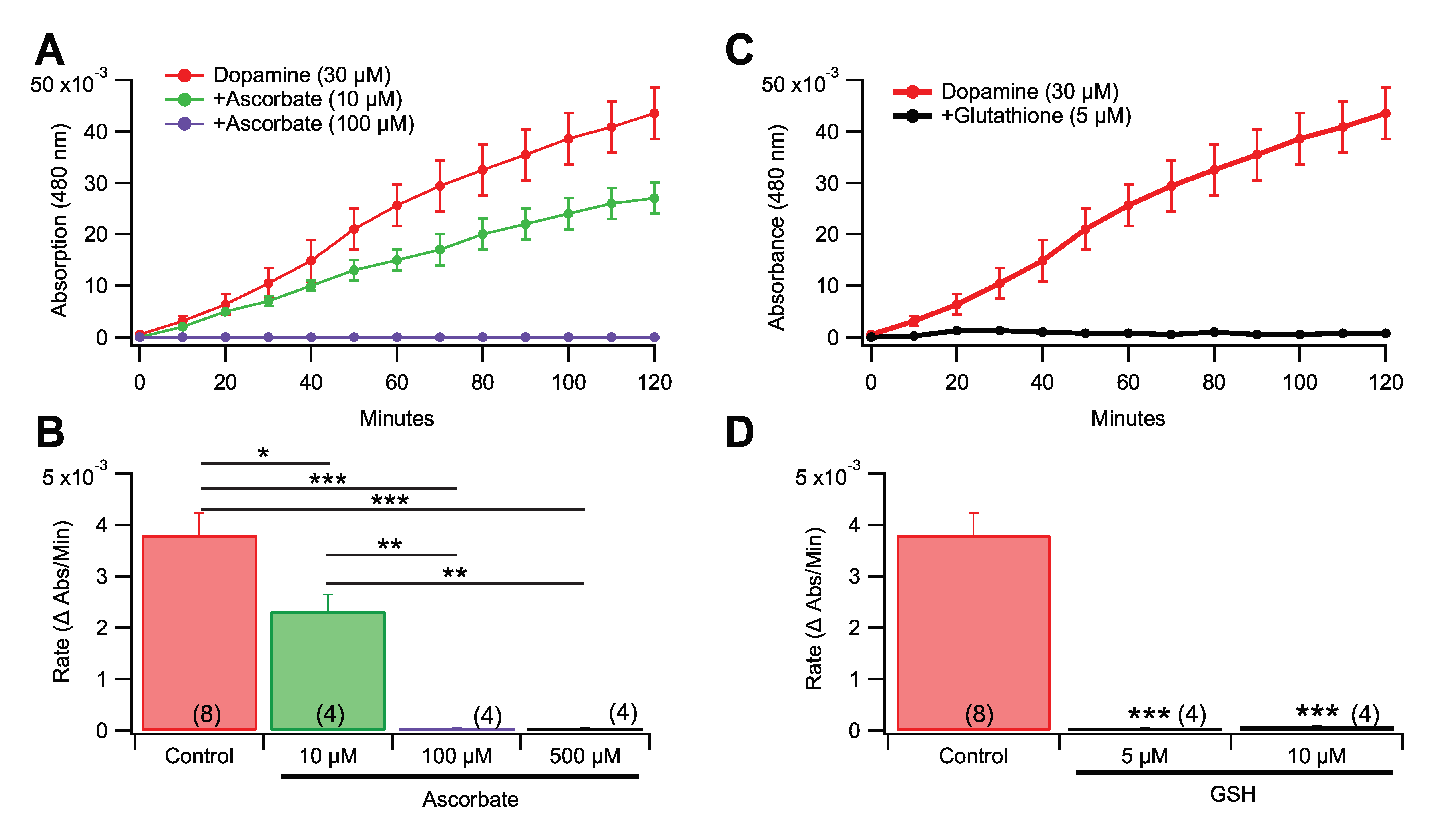
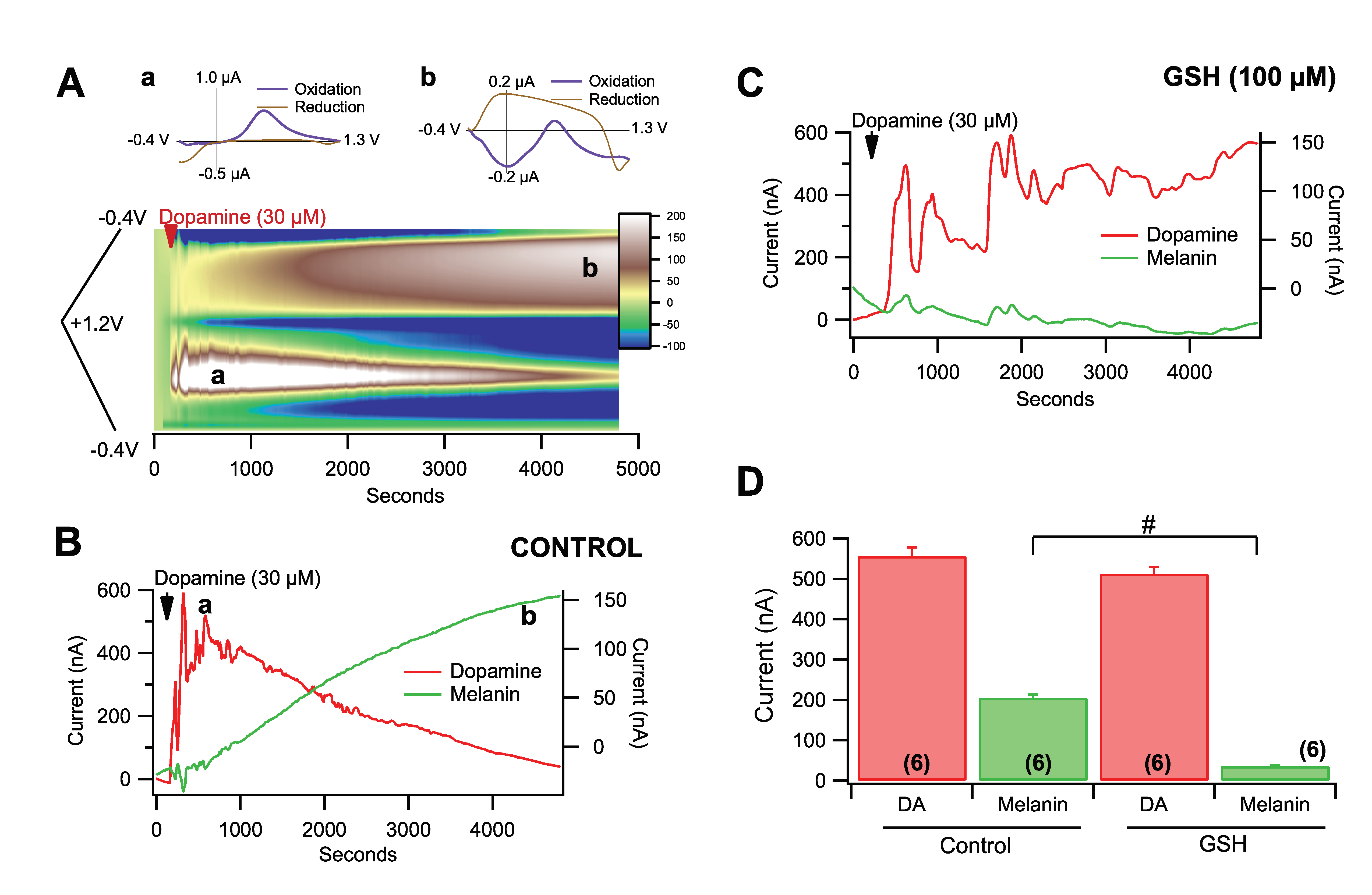
3.4. Effects of Pro-Oxidants and Antioxidants in Rate of Dopamine Melanogenesis
3.5. Fast-Scan Cyclic Voltammetry of Dopamine Melanogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DA | Dopamine |
| DES | Desferoxamine |
| DOPAL | 3,4-dihydroxyphenylacetaldehyde |
| ECF | Extracellular Fluid Buffer |
| GSH | Glutathione |
| H-NMR | Proton Nuclear Magnetic Resonance |
| ICF | Intracellular Fluid Buffer |
| NM | Neuromelanin |
| PD | Parkinson’s Disease |
| SNc | Substantia Nigra Pars Compacta |
| VTA | Ventral Tegmental Area |
References
- Godwin, L.S.; Castle, J.T.; Kohli, J.S.; Goff, P.S.; Cairney, C.J.; Keith, W.N.; Sviderskaya, E.V.; Bennett, D.C. Isolation, culture, and transfection of melanocytes. Curr. Protoc. Cell Biol. 2014, 63, 1.8.1–1.8.20. [Google Scholar] [CrossRef] [PubMed]
- Rohrbach, J.M.; Süsskind, D.; Grüb, M. The melanocyte and the eye: A review with special emphasis on the cornea. Klin. Monbl. Augenheilkd. 2011, 229, 42–47. [Google Scholar] [CrossRef]
- Lin, C.-S.; Zak, F.G. Studies on Melanocytes: VI. Melanocytes in the Middle Ear. Arch. Otolaryngol. Neck Surg. 1982, 108, 489–490. [Google Scholar] [CrossRef]
- Fritz, M.A.; Roehm, P.C.; Bannan, M.A.; Lalwani, A.K. Extracellular Melanin in Inflammatory Middle Ear Disease. Laryngoscope 2014, 124, E241–E244. [Google Scholar] [CrossRef]
- Curtis, J.L.; Rubinstein, L.J. Pigmented olfactory neuroblastoma. A new example of melanotic neuroepithelial neoplasm. Cancer 1982, 49, 2136–2143. [Google Scholar] [CrossRef]
- Rzepka, Z.; Buszman, E.; Beberok, A.; Wrześniok, D. From tyrosine to melanin: Signaling pathways and factors regulating melanogenesis. Postepy Hig. Med. Dosw. 2016, 70, 695–708. [Google Scholar] [CrossRef]
- Meyskens, F.L.; Farmer, P.J.; Anton-Culver, H. Etiologic Pathogenesis of Melanoma: A Unifying Hypothesis for the Missing Attributable Risk. Clin. Cancer Res. 2004, 10, 2581–2583. [Google Scholar] [CrossRef] [Green Version]
- Fedorow, H.; Tribl, F.; Halliday, G.; Gerlach, M.; Riederer, P.; Double, K.L. Neuromelanin in human dopamine neurons: Comparison with peripheral melanins and relevance to Parkinson’s disease. Prog. Neurobiol. 2005, 75, 109–124. [Google Scholar] [CrossRef]
- Jankovic, J. Parkinson′s disease: Clinical features and diagnosis. J. Neurol. Neurosurg. Psychiatry 2008, 79, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Dauer, W.; Przedborski, S. Parkinson′s Disease. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef] [Green Version]
- Mann, D.M.; Yates, P.O. Possible role of neuromelanin in the pathogenesis of Parkinson′s disease. Mech. Ageing Dev. 1983, 21, 193–203. [Google Scholar] [CrossRef]
- Gibb, W.R.; Lees, A.J. Anatomy, pigmentation, ventral and dorsal subpopulations of the substantia nigra, and differential cell death in Parkinson′s disease. J. Neurol. Neurosurg. Psychiatry 1991, 54, 388–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarȩba, M.; Bober, A.; Korytowski, W.; Zecca, L.; Sarna, T.; Zareba, M. The effect of a synthetic neuromelanin on yield of free hydroxyl radicals generated in model systems. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 1995, 1271, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Gaki, G.S.; Papavassiliou, A.G. Oxidative Stress-Induced Signaling Pathways Implicated in the Pathogenesis of Parkinson’s Disease. Neuromol. Med. 2014, 16, 217–230. [Google Scholar] [CrossRef]
- Dias, V.; Junn, E.; Mouradian, M.M. The Role of Oxidative Stress in Parkinson′s Disease. J. Park. Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.M.; Main, B.S.; Crack, P.J. Neuroinflammation and oxidative stress: Co-conspirators in the pathology of Parkinson’s disease. Neurochem. Int. 2013, 62, 803–819. [Google Scholar] [CrossRef]
- Zecca, L.; Wilms, H.; Geick, S.; Claasen, J.-H.; Brandenburg, L.-O.; Holzknecht, C.; Panizza, M.L.; Zucca, F.A.; Deuschl, G.; Sievers, J.; et al. Human neuromelanin induces neuroinflammation and neurodegeneration in the rat substantia nigra: Implications for Parkinson′s disease. Acta Neuropathol. 2008, 116, 47–55. [Google Scholar] [CrossRef]
- Zecca, L.; Zucca, F.; Albertini, A.; Rizzio, E.; Fariello, R.G. A proposed dual role of neuromelanin in the pathogenesis of Parkinson′s disease. Neurology 2006, 67, S8–S11. [Google Scholar] [CrossRef] [Green Version]
- Bogerts, B. A brainstem atlas of catecholaminergic neurons in man, using melanin as a natural marker. J. Comp. Neurol. 1981, 197, 63–80. [Google Scholar] [CrossRef]
- Ju, K.-Y.; Lee, Y.; Lee, S.; Park, S.B.; Lee, J.-K. Bioinspired Polymerization of Dopamine to Generate Melanin-Like Nanoparticles Having an Excellent Free-Radical-Scavenging Property. Biomacromolecules 2011, 12, 625–632. [Google Scholar] [CrossRef]
- Hilton, R.J.; Seare, M.C.; Andros, N.D.; Kenealey, Z.; Orozco, C.M.; Webb, M.; Watt, R.K. Phosphate inhibits in vitro Fe3+ loading into transferrin by forming a soluble Fe(III)–phosphate complex: A potential non-transferrin bound iron species. J. Inorg. Biochem. 2012, 110, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Aisen, P.; Leibman, A.; Zweier, J. Stoichiometric and site characteristics of the binding of iron to human transferrin. J. Biol. Chem. 1978, 253, 1930–1937. [Google Scholar] [PubMed]
- Graham, D.G. Oxidative pathways for catecholamines in the genesis of neuromelanin and cytotoxic quinones. Mol. Pharmacol. 1978, 14, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Mosca, L.; Blarzino, C.; Coccia, R.; Foppoli, C.; Rosei, M.A. Melanins From Tetrahydroisoquinolines. Free. Radic. Biol. Med. 1998, 24, 161–167. [Google Scholar] [CrossRef]
- Schilaty, N.D.; Hedges, D.M.; Jang, E.Y.; Folsom, R.J.; Yorgason, J.T.; McIntosh, J.M.; Steffensen, S.C. Acute Ethanol Inhibits Dopamine Release in the Nucleus Accumbens via α6 Nicotinic Acetylcholine Receptors. J. Pharmacol. Exp. Ther. 2014, 349, 559–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yorgason, J.T.; España, R.A.; Jones, S.R. Demon Voltammetry and Analysis software: Analysis of cocaine-induced alterations in dopamine signaling using multiple kinetic measures. J. Neurosci. Methods 2011, 202, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Wightman, R.M.; Jankowski, J.A.; Kennedy, R.T.; Kawagoe, K.T.; Schroeder, T.J.; Leszczyszyn, D.J.; Near, J.A.; DiLiberto, E.J.; Viveros, O.H. Temporally resolved catecholamine spikes correspond to single vesicle release from individual chromaffin cells. Proc. Natl. Acad. Sci. USA 1991, 88, 10754–10758. [Google Scholar] [CrossRef] [Green Version]
- Pothos, E.N.; Davila, V.; Sulzer, D. Presynaptic Recording of Quanta from Midbrain Dopamine Neurons and Modulation of the Quantal Size. J. Neurosci. 1998, 18, 4106–4118. [Google Scholar] [CrossRef]
- Zecca, L.; Tampellini, D.; Gerlach, M.; Riederer, P.; Fariello, R.G.; Sulzer, D. Substantia nigra neuromelanin: Structure, synthesis, and molecular behaviour. Mol. Pathol. 2001, 54, 414–418. [Google Scholar] [CrossRef]
- LZecca, L.; Stroppolo, A.; Gatti, A.; Tampellini, D.; Toscani, M.; Gallorini, M.; Giaveri, G.; Arosio, P.; Santambrogio, P.; Fariello, R.G.; et al. The role of iron and copper molecules in the neuronal vulnerability of locus coeruleus and substantia nigra during aging. Proc. Natl. Acad. Sci. USA 2004, 101, 9843–9848. [Google Scholar] [CrossRef] [Green Version]
- Butler, M.; Mañez, P.A.; Cabrera, G.M.; Maitre, P. Gas Phase Structure and Reactivity of Doubly Charged Microhydrated Calcium(II)–Catechol Complexes Probed by Infrared Spectroscopy. J. Phys. Chem. 2014, 118, 4942–4954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulzer, D.; Bogulavsky, J.; Larsen, K.E.; Behr, G.; Karatekin, E.; Kleinman, M.H.; Turro, N.; Krantz, D.; Edwards, R.H.; Greene, L.A.; et al. Neuromelanin biosynthesis is driven by excess cytosolic catecholamines not accumulated by synaptic vesicles. Proc. Natl. Acad. Sci. USA 2000, 97, 11869–11874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hider, R.C.; Howlin, B.J.; Miller, J.R.; Mohd-Nor, A.; Silver, J. Model compounds for microbial iron-transport compounds. Part IV. Further solution chemistry and Mössbauer studies on iron(II) and iron(III) catechol complexes. Inorg. Chim. Acta 1983, 80, 51–56. [Google Scholar] [CrossRef]
- Zecca, L.; Casella, L.; Albertini, A.; Bellei, C.; Zucca, F.A.; Engelen, M.; Zadlo, A.C.; Szewczyk, G.; Zareba, M.; Sarna, T. Neuromelanin can protect against iron-mediated oxidative damage in system modeling iron overload of brain aging and Parkinson′s disease. J. Neurochem. 2008, 106, 1866–1875. [Google Scholar] [CrossRef]
- Koeppen, A.H. The history of iron in the brain. J. Neurol. Sci. 1995, 134, 1–9. [Google Scholar] [CrossRef]
- Mccord, J.M.; Day, E.D. Superoxide-dependent production of hydroxyl radical catalyzed by iron-EDTA complex. FEBS Lett. 1978, 86, 139–142. [Google Scholar] [CrossRef] [Green Version]
- Kessel, S.L.; Emberson, R.M.; Debrunner, P.G.; Hendrickson, D.N. Iron(III), manganese(III), and cobalt(III) complexes with single chelating o-semiquinone ligands. Inorg. Chem. 1980, 19, 1170–1178. [Google Scholar] [CrossRef]
- Kawabata, T.; Schepkin, V.D.; Haramaki, N.; Phadke, R.S.; Packer, L. Iron coordination by catechol derivative antioxidants. Biochem. Pharmacol. 1996, 51, 1569–1577. [Google Scholar] [CrossRef] [Green Version]
- Li, S.W.; Lin, T.-S.; Minteer, S.; Ruggiero, D.A. 3,4-Dihydroxyphenylacetaldehyde and hydrogen peroxide generate a hydroxyl radical: Possible role in Parkinson’s disease pathogenesis. Mol. Brain Res. 2001, 93, 1–7. [Google Scholar] [CrossRef]
- Burke, W.J.; Li, S.W.; Williams, E.A.; Nonneman, R.; Zahm, D.S. 3,4-Dihydroxyphenylacetaldehyde is the toxic dopamine metabolite in vivo: Implications for Parkinson’s disease pathogenesis. Brain Res. 2003, 989, 205–213. [Google Scholar] [CrossRef]
- Oh, E.S.; Heo, C.; Kim, J.-S.; Suh, M.; Lee, Y.H.; Kim, J.-M. Hyperspectral fluorescence imaging for cellular iron mapping in thein vitromodel of Parkinson’s disease. J. Biomed. Opt. 2013, 19, 051207. [Google Scholar] [CrossRef] [Green Version]
- Winterbourn, C.C. Toxicity of iron and hydrogen peroxide: The Fenton reaction. Toxicol. Lett. 1995, 82–83, 969–974. [Google Scholar] [CrossRef]
- Naoi, M.; Maruyama, W. Type B Monoamine Oxidase and Neurotoxins. Eur. Neurol. 1993, 33, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Enochs, W.S.; Nilges, M.J.; Swartz, H.M. Purified Human Neuromelanin, Synthetic Dopamine Melanin as a Potential Model Pigment, and the Normal Human Substantia Nigra: Characterization by Electron Paramagnetic Resonance Spectroscopy. J. Neurochem. 1993, 61, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Zecca, L.; Swartz, H.M. Total and paramagnetic metals in human substantia nigra and its neuromelanin. J. Neural Transm. 1993, 5, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Double, K.L.; Gerlach, M.; Schünemann, V.; Trautwein, A.X.; Zecca, L.; Gallorini, M.; Youdim, M.B.; Riederer, P.; Ben-Shachar, D. Iron-binding characteristics of neuromelanin of the human substantia nigra. Biochem. Pharmacol. 2003, 66, 489–494. [Google Scholar] [CrossRef]
- Mosharov, E.V.; Larsen, K.E.; Kanter, E.; Phillips, K.A.; Wilson, K.; Schmitz, Y.; Krantz, D.E.; Kobayashi, K.; Edwards, R.H.; Sulzer, D. Interplay between Cytosolic Dopamine, Calcium, and α-Synuclein Causes Selective Death of Substantia Nigra Neurons. Neuron 2009, 62, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Lebedev, A.V.; Ivanova, M.V.; Timoshin, A.A.; Ruuge, E.K. Paramagnetic calcium melanins. Biophysics 2013, 58, 37–42. [Google Scholar] [CrossRef]
- Schulz, J.B.; Lindenau, J.; Seyfried, J.; Dichgans, J. Glutathione, oxidative stress and neurodegeneration. Eur. J. Biochem. 2000, 267, 4904–4911. [Google Scholar] [CrossRef]
- Nakamura, K.; Wang, W.; Kang, U.J. The role of glutathione in dopaminergic neuronal survival. J. Neurochem. 1997, 69, 1850–1858. [Google Scholar] [CrossRef]
- Yorgason, J.T.; Hedges, D.M.; Obray, J.D.; Jang, E.Y.; Bills, K.B.; Woodbury, M.; Williams, B.; Parsons, M.J.; Andres, M.A.; Steffensen, S.C. Methamphetamine increases dopamine release in the nucleus accumbens through calcium-dependent processes. Psychopharmacology 2020, 237, 1317–1330. [Google Scholar] [CrossRef] [PubMed]
- Ryu, I.S.; Yoon, S.S.; Choi, M.J.; Lee, Y.E.; Kim, J.S.; Kim, W.H.; Cheong, J.H.; Kim, H.J.; Jang, C.; Lee, Y.S.; et al. The potent psychomotor, rewarding and reinforcing properties of 3-fluoromethamphetamine in rodents. Addict. Biol. 2020, 25, e12846. [Google Scholar] [CrossRef] [PubMed]
- Yorgason, J.T.; Zeppenfeld, D.M.; Williams, J.T. Cholinergic Interneurons Underlie Spontaneous Dopamine Release in Nucleus Accumbens. J. Neurosci. 2017, 37, 2086–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, E.Y.; Ryu, Y.-H.; Lee, B.H.; Chang, S.-C.; Yeo, M.J.; Kim, S.H.; Folsom, R.J.; Schilaty, N.D.; Kim, K.J.; Yang, C.H.; et al. Involvement of reactive oxygen species in cocaine-taking behaviors in rats. Addict. Biol. 2015, 20, 663–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedges, D.M.; Obray, J.D.; Yorgason, J.T.; Jang, E.Y.; Weerasekara, V.K.; Uys, J.D.; Bellinger, F.P.; Steffensen, S.C. Methamphetamine Induces Dopamine Release in the Nucleus Accumbens Through a Sigma Receptor-Mediated Pathway. Neuropsychopharmacol. 2017, 43, 1405–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, E.Y.; Yang, C.H.; Hedges, D.M.; Kim, S.P.; Lee, J.Y.; Ekins, T.G.; Garcia, B.T.; Kim, H.Y.; Nelson, A.C.; Kim, N.J.; et al. The role of reactive oxygen species in methamphetamine self-administration and dopamine release in the nucleus accumbens. Addict. Biol. 2017, 22, 1304–1315. [Google Scholar] [CrossRef]
- Yorgason, J.T.; Jones, S.R.; España, R.A. Low and high affinity dopamine transporter inhibitors block dopamine uptake within 5 sec of intravenous injection. Neuroscience 2011, 182, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Yorgason, J.T.; Calipari, E.S.; Ferris, M.J.; Karkhanis, A.N.; Fordahl, S.C.; Weiner, J.L.; Jones, S.R. Social isolation rearing increases dopamine uptake and psychostimulant potency in the striatum. Neuropharmacology 2016, 101, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Sulzer, D.; Mosharov, E.; Tallóczy, Z.; Zucca, F.A.; Simon, J.D.; Zecca, L. Neuronal pigmented autophagic vacuoles: Lipofuscin, neuromelanin, and ceroid as macroautophagic responses during aging and disease. J. Neurochem. 2008, 106, 24–36. [Google Scholar] [CrossRef]
- Gała̧zka-Friedman, J.; Bauminger, E.R.; Friedman, A.; Barcikowska, M.; Hechel, D.; Nowik, I. Iron in parkinsonian and control substantia nigra—A Mössbauer spectroscopy study. Mov. Disord. 1996, 11, 8–16. [Google Scholar] [CrossRef]
- Gerlach, M.; Trautwein, A.X.; Zecca, L.; Youdim, M.B.H.; Riederer, P. Mössbauer Spectroscopic Studies of Purified Human Neuromelanin Isolated from the Substantia Nigra. J. Neurochem. 2002, 65, 923–926. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hedges, D.M.; Yorgason, J.T.; Perez, A.W.; Schilaty, N.D.; Williams, B.M.; Watt, R.K.; Steffensen, S.C. Spontaneous Formation of Melanin from Dopamine in the Presence of Iron. Antioxidants 2020, 9, 1285. https://doi.org/10.3390/antiox9121285
Hedges DM, Yorgason JT, Perez AW, Schilaty ND, Williams BM, Watt RK, Steffensen SC. Spontaneous Formation of Melanin from Dopamine in the Presence of Iron. Antioxidants. 2020; 9(12):1285. https://doi.org/10.3390/antiox9121285
Chicago/Turabian StyleHedges, David M., Jordan T. Yorgason, Andrew W. Perez, Nathan D. Schilaty, Benjamin M. Williams, Richard K. Watt, and Scott C. Steffensen. 2020. "Spontaneous Formation of Melanin from Dopamine in the Presence of Iron" Antioxidants 9, no. 12: 1285. https://doi.org/10.3390/antiox9121285