Phytochemical Characterization of Dillenia indica L. Bark by Paper Spray Ionization-Mass Spectrometry and Evaluation of Its Antioxidant Potential Against t-BHP-Induced Oxidative Stress in RAW 264.7 Cells

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Extraction
2.2. PSI-MS
2.3. Data Processing
2.4. Radical-Scavenging Activity Assays
2.5. Cell Culture and Cell Viability Assay
2.6. Measurement of Intracellular ROS
2.7. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results and Discussion
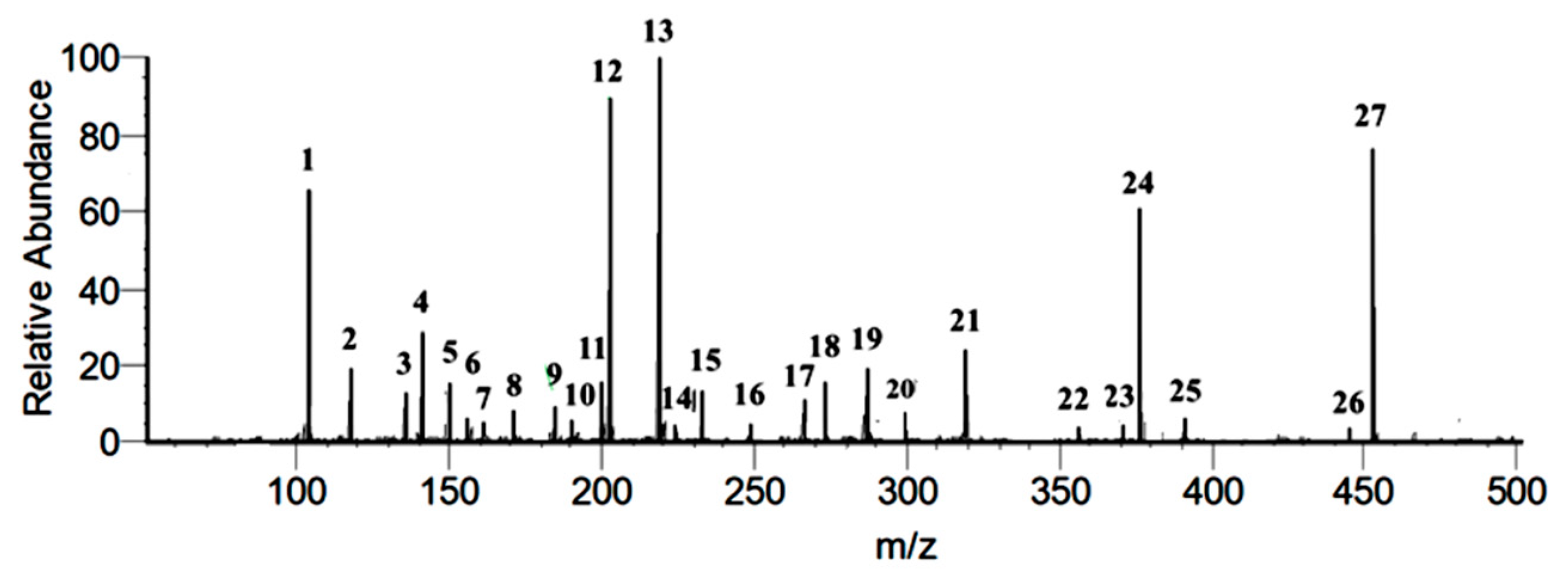
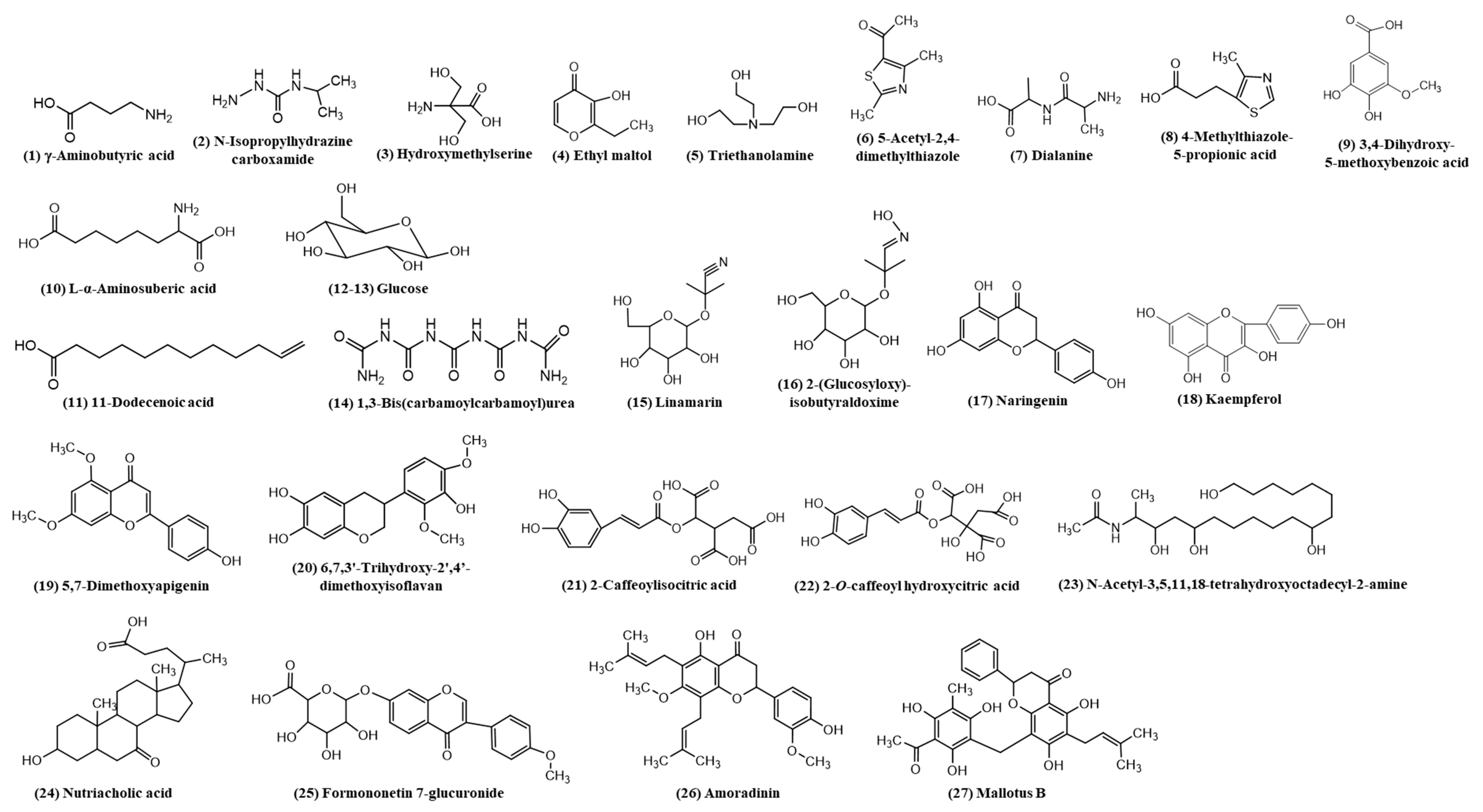
3.1. Identification of Secondary Metabolites of DIBEt
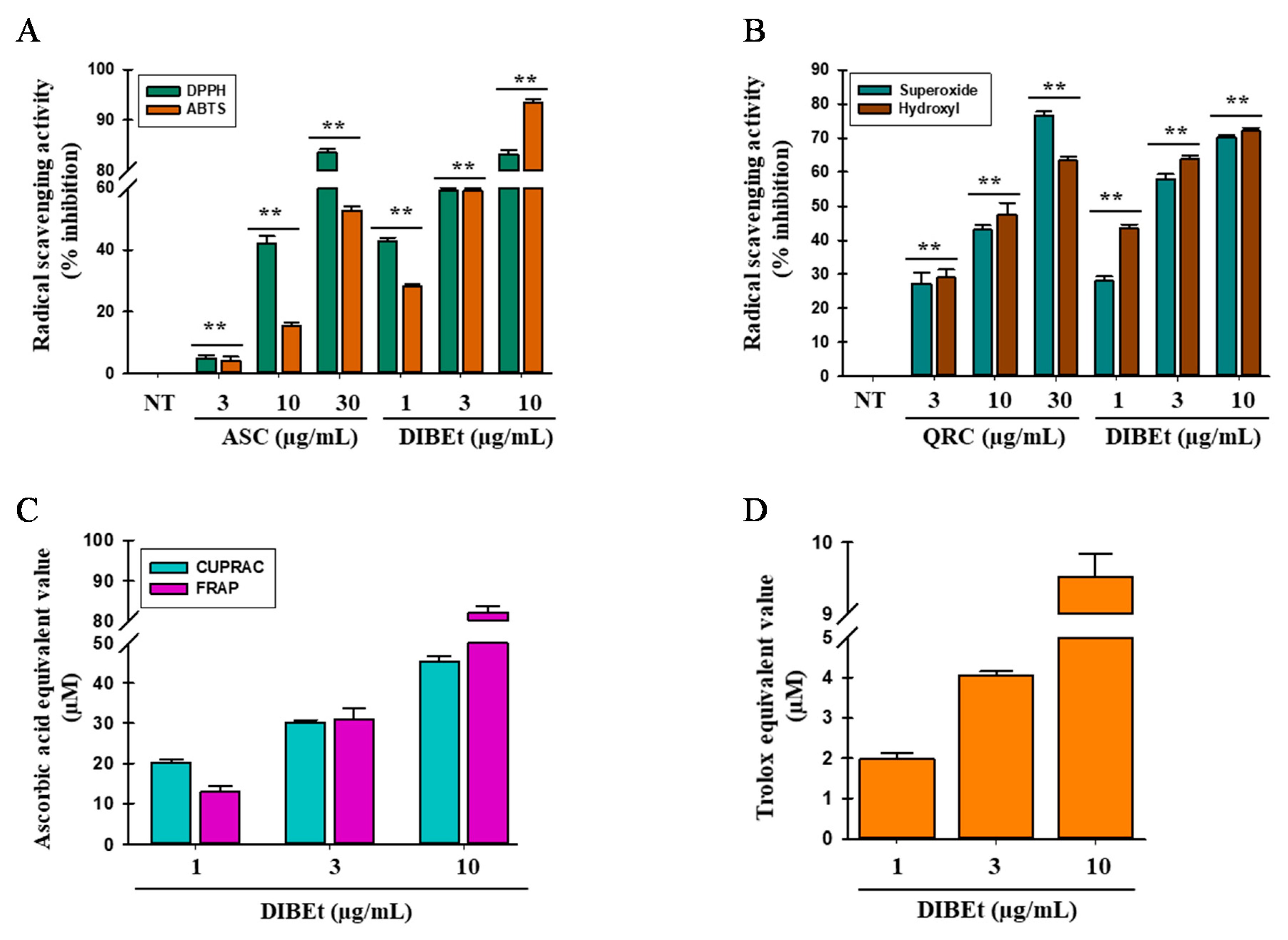
3.2. Radical-Scavenging Activities of DIB Extracts
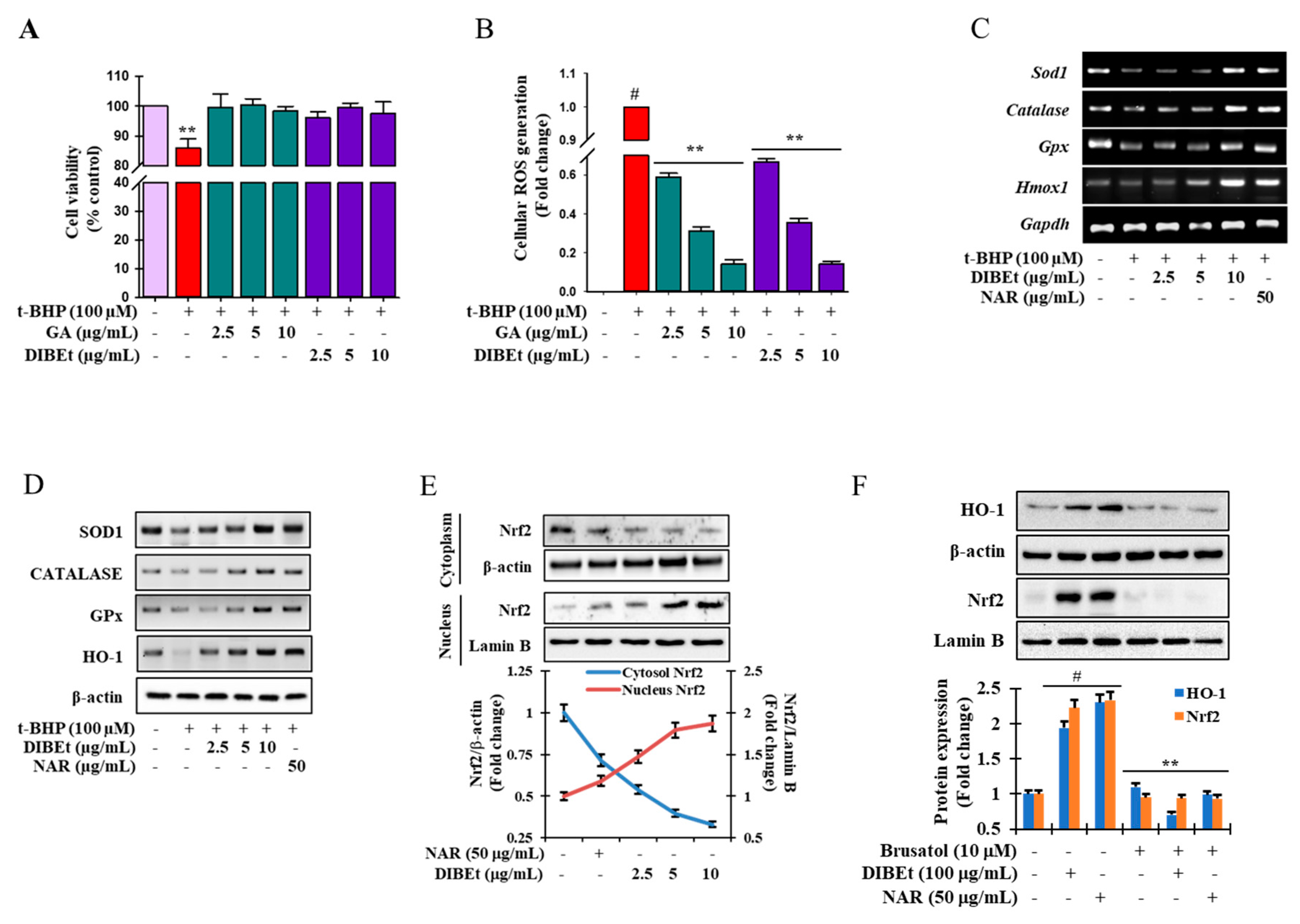
3.3. DIBEt Attenuates t-BHP-Induced Cellular Oxidative Stress
3.4. Effects of DIBEt on Antioxidant Enzyme Expression in RAW 264.7 Cells
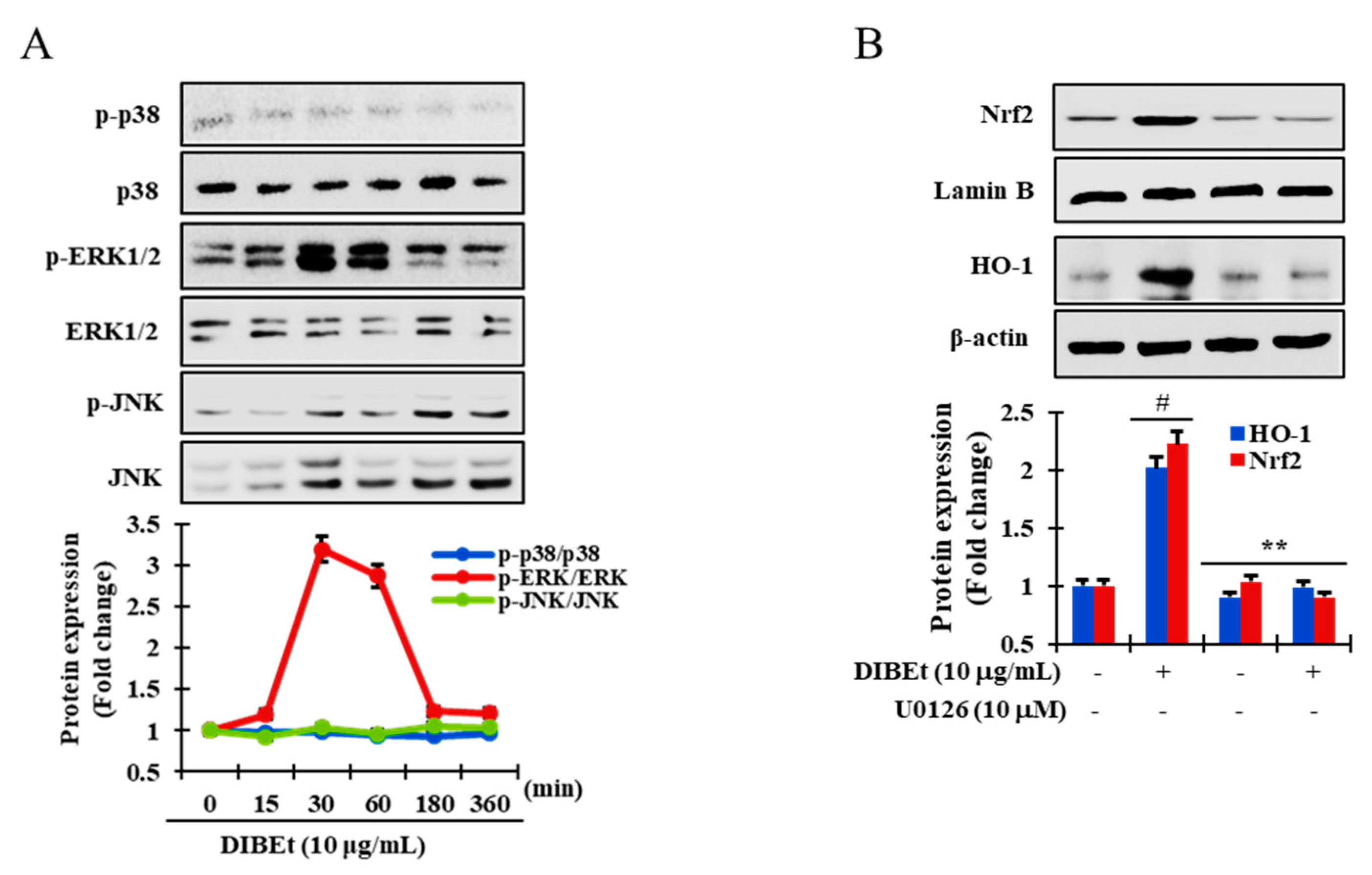
3.5. Effects of DIBEt on Phase II Enzymes Mediated by Nrf2 Nuclear Translocation in RAW 264.7 Cells
3.6. DIBEt Regulates Nrf2 Translocation Via Activation of MAPK to Lessen Oxidative Stress
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Banerjee, J.; Das, A.; Sinha, M.; Saha, S. Biological efficacy of medicinal plant extracts in preventing oxidative damage. Oxidative Med. Cell. Longev. 2018, 2018, 7904349. [Google Scholar] [CrossRef] [PubMed]
- Castaneda, O.A.; Lee, S.C.; Ho, C.T.; Huang, T.C. Macrophages in oxidative stress and models to evaluate the antioxidant function of dietary natural compounds. J. Food Drug Anal. 2017, 25, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.B.; Ju, M.K.; Lee, S.H. DNA protecting activities of Nymphaea nouchali (Burm. f) flower extract attenuate t-BHP-induced oxidative stress cell death through Nrf2-mediated induction of heme oxygenase-1 expression by activating MAP-kinases. Int. J. Mol. Sci. 2017, 18, 2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, K.; Motohashi, H.; Yamamoto, M. Molecular mechanisms of the Keap1-Nrf2 pathway in stress response and cancer evolution. Genes Cells 2011, 16, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Dinkova-Kostova, A.T. The cytoprotective role of the Keap1-Nrf2 pathway. Arch. Toxicol. 2011, 85, 241–272. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, J.; Cooks, R.G.; Ouyang, Z. Paper spray for direct analysis of complex mixtures using mass spectrometry. Angew. Chem. (Int. Ed. Engl.) 2010, 49, 877–880. [Google Scholar] [CrossRef] [PubMed]
- Di Donna, L.; Taverna, D.; Indelicato, S.; Napoli, A.; Sindona, G.; Mazzotti, F. Rapid assay of resveratrol in red wine by paper spray tandem mass spectrometry and isotope dilution. Food Chem. 2017, 229, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Teodoro, J.A.R.; Pereira, H.V.; Sena, M.M.; Piccin, E.; Zacca, J.J.; Augusti, R. Paper spray mass spectrometry and chemometric tools for a fast and reliable identification of counterfeit blended Scottish whiskies. Food Chem. 2017, 237, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Wei, P.; Hsu, H.C.; Cooks, R.G. Direct analysis of 4-methylimidazole in foods using paper spray mass spectrometry. Analyst 2013, 138, 4624–4630. [Google Scholar] [CrossRef] [PubMed]
- Saiful Yazan, L.; Armania, N. Dillenia species: A review of the traditional uses, active constituents and pharmacological properties from pre-clinical studies. Pharm. Biol. 2014, 52, 890–897. [Google Scholar] [CrossRef]
- Kumar, D.; Mallick, S.; Vedasiromoni, J.R.; Pal, B.C. Anti-leukemic activity of Dillenia indica L. fruit extract and quantification of betulinic acid by HPLC. Phytomed. Int. J. Phytother. Phytopharmacol. 2010, 17, 431–435. [Google Scholar] [CrossRef]
- Abdille, M.H.; Singh, R.; Jayaprakasha, G.; Jena, B. Antioxidant activity of the extracts from Dillenia indica fruits. Food Chem. 2005, 90, 891–896. [Google Scholar] [CrossRef]
- Deraedt, R.; Jouquey, S.; Delevallee, F.; Flahaut, M. Release of prostaglandins E and F in an algogenic reaction and its inhibition. Eur. J. Pharmacol. 1980, 61, 17–24. [Google Scholar] [CrossRef]
- Grover, J.K.; Yadav, S.; Vats, V. Medicinal plants of India with anti-diabetic potential. J. Ethnopharmacol. 2002, 81, 81–100. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vazquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef] [PubMed]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E.; et al. METLIN: A technology platform for identifying knowns and unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.B.; Kwon, K.R.; Lee, S.H.; Lee, S.H. Lannea coromandelica (Houtt.) Merr. induces heme oxygenase 1 (HO-1) expression and reduces oxidative stress via the p38/c-Jun N-terminal kinase-nuclear factor erythroid 2-related factor 2 (p38/JNK-NRF2)-mediated antioxidant pathway. Int. J. Mol. Sci. 2017, 18, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.B.; Ju, M.K.; Kwon, Y.G.; Lee, S.H. Protopine attenuates inflammation stimulated by carrageenan and LPS via the MAPK/NF-kappaB pathway. Food Chem. Toxicol. 2019, 131, 110583. [Google Scholar] [CrossRef]
- Rahman, M.S.; Hee Choi, Y.; Seok Choi, Y.; Alam, M.B.; Han Lee, S.; Cheol Yoo, J. A novel antioxidant peptide, purified from Bacillus amyloliquefaciens, showed strong antioxidant potential via Nrf-2 mediated heme oxygenase-1 expression. Food Chem. 2018, 239, 502–510. [Google Scholar] [CrossRef]
- Piraud, M.; Vianey-Saban, C.; Petritis, K.; Elfakir, C.; Steghens, J.P.; Morla, A.; Bouchu, D. ESI-MS/MS analysis of underivatised amino acids: A new tool for the diagnosis of inherited disorders of amino acid metabolism. Fragmentation study of 79 molecules of biological interest in positive and negative ionisation mode. Rapid Commun. Mass Spectr. RCM 2003, 17, 1297–1311. [Google Scholar] [CrossRef] [PubMed]
- Shiojima, K.; Arai, Y.; Masuda, K.; Takase, Y.; Ageta, T.; Ageta, H. Mass spectra of pentacyclic triterpenoids. Chem. Pharmaceut. Bull. 1992, 40, 1683–1690. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Sarma, B.P.; Ahmed, G.; Nirmala, C.B.; Choudhury, M.K. In vitro anti oxidant activity total phenolic content of Dillenia indica Garcinia penducalata, commonly used fruits in Assamese cuisine. Free Radic. Antioxid. 2012, 2, 30–36. [Google Scholar] [CrossRef]
- Kumaran, A.; Karunakaran, R.J. In vitro antioxidant activities of methanol extracts of five Phyllanthus species from India. LWT-Food Sci. Technol. 2007, 40, 344–352. [Google Scholar] [CrossRef]
- Astley, S.B. Dietary antioxidants—Past, present and future? Trends Food Sci. Technol. 2003, 14, 93–98. [Google Scholar] [CrossRef]
- Holasova, M.; Fiedlerova, V.; Smrcinova, H.; Orsak, M.; Lachman, J.; Vavreinova, S. Buckwheat—The source of antioxidant activity in functional foods. Food Res. Int. 2002, 35, 207–211. [Google Scholar] [CrossRef]
- Huyut, Z.; Beydemir, Ş.; Gülçin, İ. Antioxidant and antiradical properties of selected flavonoids and phenolic compounds. Biochem. Res. Int. 2017, 2017, 7616791. [Google Scholar] [CrossRef]
- Kapepula, P.M.; Kabamba Ngombe, N.; Tshisekedi Tshibangu, P.; Tsumbu, C.; Franck, T.; Mouithys-Mickalad, A.; Mumba, D.; Tshala-Katumbay, D.; Serteyn, D.; Tits, M.; et al. Comparison of metabolic profiles and bioactivities of the leaves of three edible Congolese Hibiscus species. Nat. Prod. Res. 2017, 31, 2885–2892. [Google Scholar] [CrossRef]
- Cavia-Saiz, M.; Busto, M.D.; Pilar-Izquierdo, M.C.; Ortega, N.; Perez-Mateos, M.; Muñiz, P. Antioxidant properties, radical scavenging activity and biomolecule protection capacity of flavonoid naringenin and its glycoside naringin: A comparative study. J. Sci. Food Agric. 2010, 90, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fang, X.; Ge, L.; Cao, F.; Zhao, L.; Wang, Z.; Xiao, W. Antitumor, antioxidant and anti-inflammatory activities of kaempferol and its corresponding glycosides and the enzymatic preparation of kaempferol. PLoS ONE 2018, 13, e0197563. [Google Scholar] [CrossRef]
- Canabady-Rochelle, L.L.; Harscoat-Schiavo, C.; Kessler, V.; Aymes, A.; Fournier, F.; Girardet, J.M. Determination of reducing power and metal chelating ability of antioxidant peptides: Revisited methods. Food Chem. 2015, 183, 129–135. [Google Scholar] [CrossRef]
- Baran, M.Y.; Emecen, G.; Simon, A.; Tóth, G.; Kuruuzum-Uz, A. Assessment of the antioxidant activity and genotoxicity of the extracts and isolated glycosides with a new flavonoid from Lotus aegaeus (Gris.) Boiss. Ind. Crops Prod. 2020, 153, 112590. [Google Scholar] [CrossRef]
- Gangwar, M.; Goel, R.K.; Nath, G. Mallotus philippinensis Muell. Arg (Euphorbiaceae): Ethnopharmacology and phytochemistry review. Biomed. Res. Int. 2014, 2014, 213973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Yu, R.; Zhou, Q.; Jiang, S.; Le, G. Protective effects of γ-aminobutyric acid against H2O2-induced oxidative stress in RIN-m5F pancreatic cells. Nutr. Metab. 2018, 15, 60. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Gao, J.; Zheng, Y.; Wang, X.; Chen, C.; Cao, K.; Xu, J.; Li, Y.; Lu, W.; Liu, J. Zeaxanthin induces Nrf2-mediated phase II enzymes in protection of cell death. Cell Death Dis. 2014, 5, e1218. [Google Scholar] [CrossRef] [Green Version]
- Poljsak, B.; Šuput, D.; Milisav, I. Achieving the balance between ROS and antioxidants: When to use the synthetic antioxidants. Oxidative Med. Cell. Longev. 2013, 2013, 956792. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Araujo, J.A.; Zhang, M.; Yin, F. Heme oxygenase-1, oxidation, inflammation, and atherosclerosis. Front. Pharmacol. 2012, 3, 119. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Pachón, M.S.; Berná, G.; Otaolaurruchi, E.; Troncoso, A.M.; Martín, F.; García-Parrilla, M.C. Changes in antioxidant endogenous enzymes (activity and gene expression levels) after repeated red wine intake. J. Agric. Food Chem. 2009, 57, 6578–6583. [Google Scholar] [CrossRef]
- Crespo, I.; García-Mediavilla, M.V.; Almar, M.; González, P.; Tuñón, M.J.; Sánchez-Campos, S.; González-Gallego, J. Differential effects of dietary flavonoids on reactive oxygen and nitrogen species generation and changes in antioxidant enzyme expression induced by proinflammatory cytokines in Chang liver cells. Food Chem. Toxicol. 2008, 46, 1555–1569. [Google Scholar] [CrossRef] [PubMed]
- Schröter, D.; Neugart, S.; Schreiner, M.; Grune, T.; Rohn, S.; Ott, C. Amaranth’s 2-caffeoylisocitric acid—An anti-inflammatory caffeic acid derivative that impairs NF-κB signaling in LPS-challenged RAW 264.7 macrophages. Nutrients 2019, 11, 571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, W.; An, Y.; He, X.; Zhang, D.; He, W. Protection of kaempferol on oxidative stress-induced retinal pigment epithelial cell damage. Oxidative Med. Cell. Longev. 2018, 2018, 1610751. [Google Scholar] [CrossRef]
- Wang, K.; Chen, Z.; Huang, L.; Meng, B.; Zhou, X.; Wen, X.; Ren, D. Naringenin reduces oxidative stress and improves mitochondrial dysfunction via activation of the Nrf2/ARE signaling pathway in neurons. Int. J. Mol. Med. 2017, 40, 1582–1590. [Google Scholar] [CrossRef] [Green Version]
- Aladaileh, S.H.; Hussein, O.E.; Abukhalil, M.H.; Saghir, S.A.M.; Bin-Jumah, M.; Alfwuaires, M.A.; Germoush, M.O.; Almaiman, A.A.; Mahmoud, A.M. Formononetin upregulates Nrf2/HO-1 signaling and prevents oxidative stress, inflammation, and kidney injury in methotrexate-induced rats. Antioxidants (Basel Switz.) 2019, 8, 430. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Wang, J.; Zhao, G.; Lin, M.; Lang, Y.; Zhang, D.; Feng, D.; Tu, C. Apigenin protects human melanocytes against oxidative damage by activation of the Nrf2 pathway. Cell Stress Chaperones 2020, 25, 277–285. [Google Scholar] [CrossRef]
- Vomund, S.; Schäfer, A.; Parnham, M.J.; Brüne, B.; von Knethen, A. Nrf2, the master regulator of anti-oxidative responses. Int. J. Mol. Sci. 2017, 18, 2772. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-P.; Goldring, C.E.; Copple, I.M.; Wang, H.-Y.; Wei, W.; Kitteringham, N.R.; Park, B.K. Extract of Ginkgo biloba induces phase 2 genes through Keap1-Nrf2-ARE signaling pathway. Life Sci. 2007, 80, 1586–1591. [Google Scholar] [CrossRef]
- Shen, G.; Hebbar, V.; Nair, S.; Xu, C.; Li, W.; Lin, W.; Keum, Y.S.; Han, J.; Gallo, M.A.; Kong, A.N. Regulation of Nrf2 transactivation domain activity. The differential effects of mitogen-activated protein kinase cascades and synergistic stimulatory effect of Raf and CREB-binding protein. J. Biol. Chem. 2004, 279, 23052–23060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Yoon, J.; Ham, H.; Sung, J.; Kim, Y.; Choi, Y.; Lee, J.-S.; Jeong, H.-S.; Lee, J.; Kim, D. Black rice extract protected HepG2 cells from oxidative stress-induced cell death via ERK1/2 and Akt activation. Nutr. Res. Pract. 2014, 8, 125–131. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound Name | MW | MM | EF | (M + H) m/z a | (M + H) m/z b | PSI-MS/MS |
|---|---|---|---|---|---|---|---|
| (Positive Ionization) | |||||||
| 1 | γ-Aminobutyric acid (GABA) | 103.121 | 103.063 | C4H9NO2 | 104.108 | 104.071 | 87.04, 60.02, 58.07 |
| 2 | N-Isopropylhydrazinecarboxamide | 117.152 | 117.09 | C4H11N3O | 118.087 | 118.098 | 101.06, 59.07, 58.06 |
| 3 | Hydroxymethylserine | 135.119 | 135.053 | C4H9NO4 | 136.062 | 136.061 | 119.035 |
| 4 | Ethyl maltol | 140.138 | 140.047 | C7H8O3 | 141.055 | 141.055 | 140.06, 126.03, 123.04, 113.06, 108.02, 81.04 |
| 5 | Triethanolamine | 149.188 | 149.105 | C6H15NO3 | 150.113 | 150.113 | 132.10, 120.10, 103.06, 88.07 |
| 6 | 5-Acetyl-2,4-dimethylthiazole | 155.22 | 155.04 | C7H10NOS | 156.043 | 155.048 | 140.03, 122.07, 81.04 |
| 7 | Dialanine | 160.171 | 160.085 | C6H12N2O3 | 161.097 | 161.092 | 145.06, 144.07, 131.06, 118.05, 101.07, 88.04, 72.04 |
| 8 | 4-Methylthiazole-5-propionic acid | 171.214 | 171.035 | C7H9NO2S | 172.043 | 172.043 | 156.06, 141.08, 128.07, 113.07 |
| 9 | 3,5-Dihydroxy-4-methoxybenzoic acid | 184.157 | 184.037 | C8H8O5 | 185.045 | 185.045 | 170.02, 155.03, 126.02, 109.03, 95.05, 91.02, 77.04 |
| 10 | L-α-Aminosuberic acid | 189.211 | 189.1 | C8H15NO4 | 190.108 | 190.108 | 173.08, 130.09, 128.07, 113.06, 101.06, 70.01 |
| 11 | 11-Dodecenoic acid | 198.306 | 198.162 | C12H22O2 | 199.169 | 199.169 | 181.16, 127.08, 95.09, 85.03 |
| 12 | Glucose Na adduct | 203.053 | 203.052 | ||||
| 13 | Glucose K adduct | 219.027 | 219.026 | ||||
| 14 | 1,3-Bis(carbamoylcarbamoyl)urea (Carbonyldibiuret) | 232.156 | 232.056 | C5H8N6O5 | 233.063 | 233.063 | 216.08, 188.09, 145.03, 119.01, 102.05 |
| 15 | Linamarin | 247.247 | 247.106 | C10H17NO6 | 248.114 | 248.113 | 230.10, 182.08, 128.07, 115.04, 98.04 |
| 16 | 2-(Glucosyloxy) isobutyraldoxime | 265.262 | 265.112 | C10H19NO7 | 266.123 | 266.123 | 248.11, 230.10, 182.08, 128.07, 115.04, 98.04 |
| 17 | Naringenin | 272.256 | 272.068 | C15H12O5 | 273.076 | 273.076 | 153.01, 147.04, 119.05 |
| 18 | Kaempferol | 286.239 | 286.048 | C15H10O6 | 287.056 | 287.055 | 269.04, 141.05, 213.05, 165.02, 153.02, 137.02, 121.02 |
| 19 | 5,7-Dimethoxyapigenin | 298.29 | 298.084 | C17H14O5 | 299.056 | 299.091 | 271.10, 253.09, 179.03, 137.06, 123.04 |
| 20 | 6,7,3′-Trihydroxy-2′,4′-dimethoxyisoflavan (Bryaflavan) | 318.325 | 318.11 | C17H18O6 | 319.116 | 319.11 | 301.11, 245.08, 195.10, 167.07, 153.05, 149.05, 137.02 |
| 21 | 2-Caffeoylisocitric acid | 354.267 | 354.059 | C15H14O10 | 355.07 | 355.066 | 310.07, 121.03, 203.02, 192.03, 177.04 |
| 22 | 2-O-caffeoylhydroxycitric acid | 370.26 | 370.053 | C15H14O11 | 371.075 | 371.061 | 311.04, 279.05, 267.05, 237.04 |
| 23 | N-Acetyl-3,5,11,18-tetrahydroxyoctadecyl-2-amine | 375.52 | 375.295 | C20H41NO5 | 376.26 | 376.303 | 358.30, 340.28, 226.18, 161.15, 147.14, 137.06, 123.04, 109.10 |
| 24 | Nutriacholic acid | 390.62 | 390.277 | C24H38O4 | 391.284 | 391.284 | 361.27, 207.14, 189.13, 161.13, 149.13 |
| 25 | Formononetin 7-glucoronide | 444.38 | 444.105 | C22H20O10 | 445.12 | 445.113 | 413.12, 251.07, 137.02, 123.04 |
| 26 | Amoradinin | 452.521 | 452.219 | C27H32O6 | 453.231 | 453.231 | 391.15, 373.14, 361.14, 207.07, 191.07 |
| 27 | Mallotus B (Isoallorottlerin) | 518.554 | 518.194 | C30H30O8 | 519.205 | 519.202 | 339.17, 237.11 |
| Compound Name | DPPH a | ABTS a | CUPRAC b | FRAP b | |
|---|---|---|---|---|---|
| 1 | GABA | >100 | >100 | 0.12 ± 1.51 | 0.91 ± 1.25 |
| 2 | Ethyl maltol | 29.42 ± 0.85 | 34.52 ± 0.35 | 21.05 ± 1.15 | 28.52 ± 0.55 |
| 3 | Dialanine | 85.69 ± 0.25 | 92.57 ± 0.53 | 09.55 ± 0.25 | 11.02 ± 1.52 |
| 4 | 3,4-Dihydroxy-5-methoxybenzoic acid | 18.25 ± 1.52 | 12.05 ± 0.45 | 25.15 ± 0.85 | 35.25 ± 0.89 |
| 5 | Linamarin | >100 | >100 | 0.32 ± 0.94 | 0.71 ± 0.65 |
| 6 | Naringenin | 4.52 ± 0.12 | 3.98 ± 0.78 | 33.32 ± 0.31 | 54.34 ± 0.29 |
| 7 | Kaempferol | 10.32 ± 0.54 | 5.21 ± 0.45 | 31.02 ± 0.57 | 45.53 ± 0.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alam, M.B.; Ahmed, A.; Islam, S.; Choi, H.-J.; Motin, M.A.; Kim, S.; Lee, S.-H. Phytochemical Characterization of Dillenia indica L. Bark by Paper Spray Ionization-Mass Spectrometry and Evaluation of Its Antioxidant Potential Against t-BHP-Induced Oxidative Stress in RAW 264.7 Cells. Antioxidants 2020, 9, 1099. https://doi.org/10.3390/antiox9111099
Alam MB, Ahmed A, Islam S, Choi H-J, Motin MA, Kim S, Lee S-H. Phytochemical Characterization of Dillenia indica L. Bark by Paper Spray Ionization-Mass Spectrometry and Evaluation of Its Antioxidant Potential Against t-BHP-Induced Oxidative Stress in RAW 264.7 Cells. Antioxidants. 2020; 9(11):1099. https://doi.org/10.3390/antiox9111099
Chicago/Turabian StyleAlam, Md Badrul, Arif Ahmed, Syful Islam, Hee-Jeong Choi, Md Abdul Motin, Sunghwan Kim, and Sang-Han Lee. 2020. "Phytochemical Characterization of Dillenia indica L. Bark by Paper Spray Ionization-Mass Spectrometry and Evaluation of Its Antioxidant Potential Against t-BHP-Induced Oxidative Stress in RAW 264.7 Cells" Antioxidants 9, no. 11: 1099. https://doi.org/10.3390/antiox9111099