

Low-Dose Acetylsalicylic Acid and Mitochondria-Targeted Antioxidant Mitoquinone Attenuate Non-Alcoholic Steatohepatitis in Mice

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. Histopathologic Evaluation
2.3. Quantitative Real-Time Polymerase Chain Reaction
2.4. Western Blot Analysis
2.5. Analysis of 15-Epi-Lipoxin A4 in Liver
2.6. Statistical Analysis
3. Results
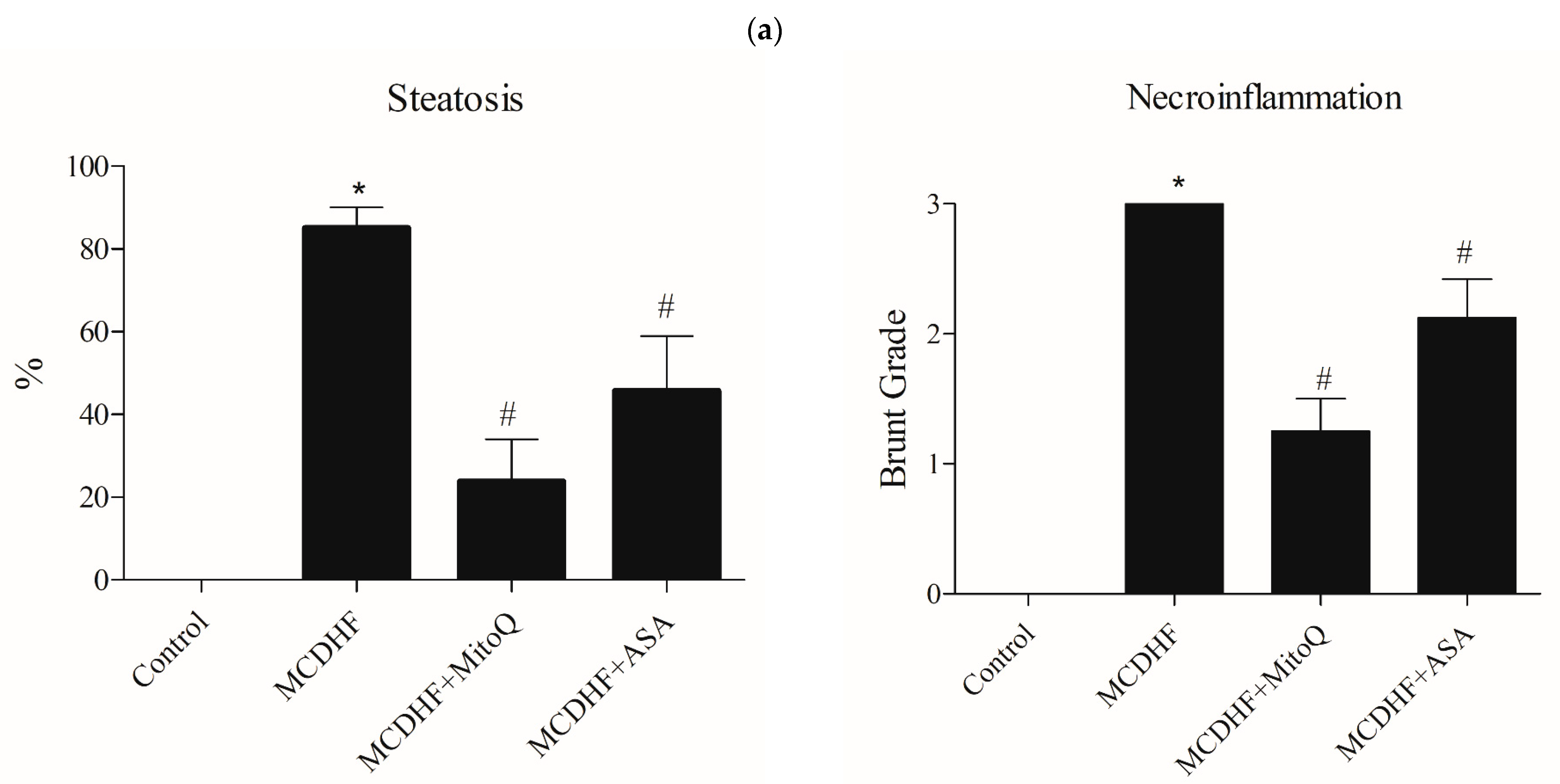
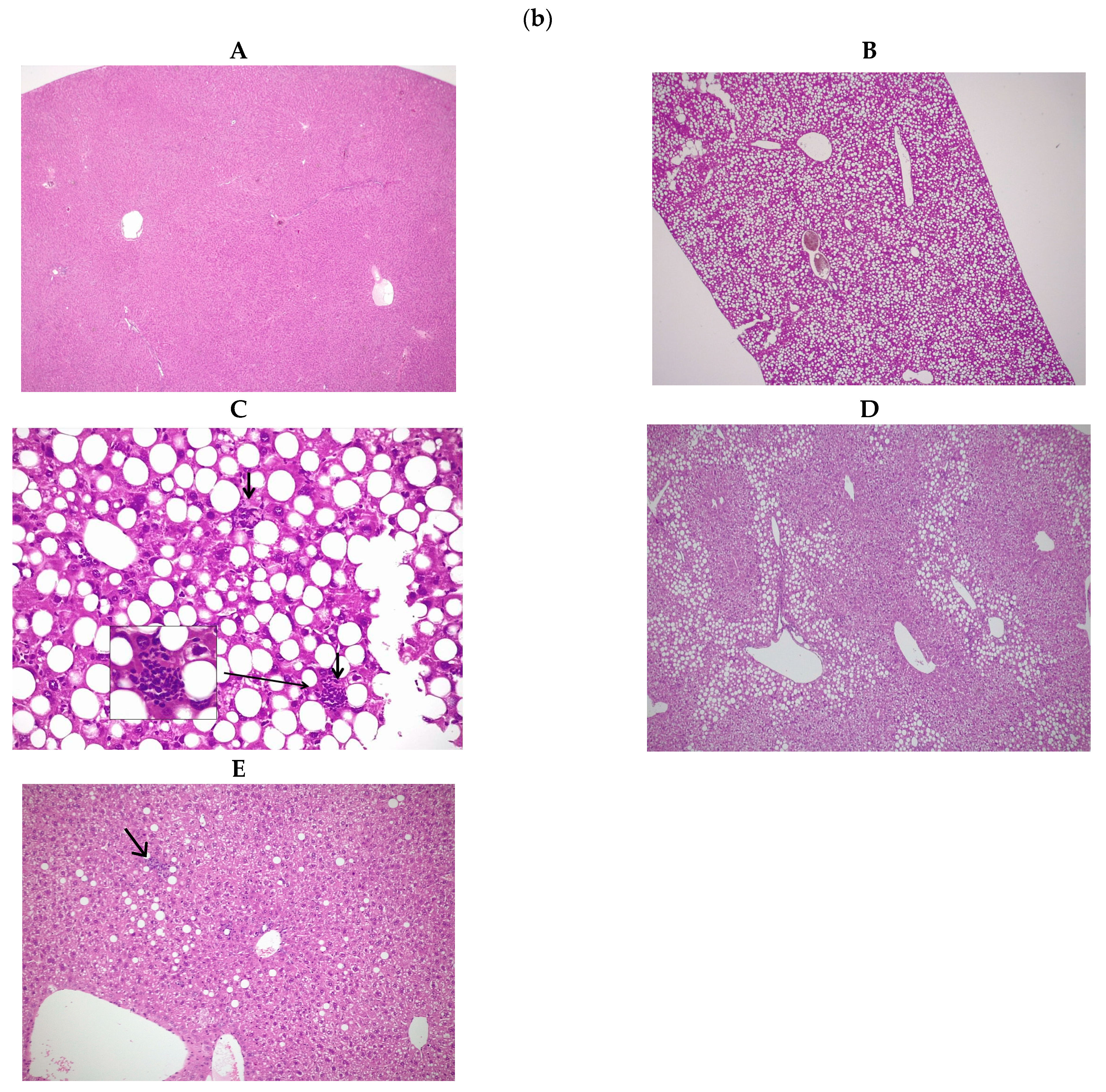
3.1. Effects of ASA and MitoQ on Liver Steatosis and Necroinflammation through Histopathology
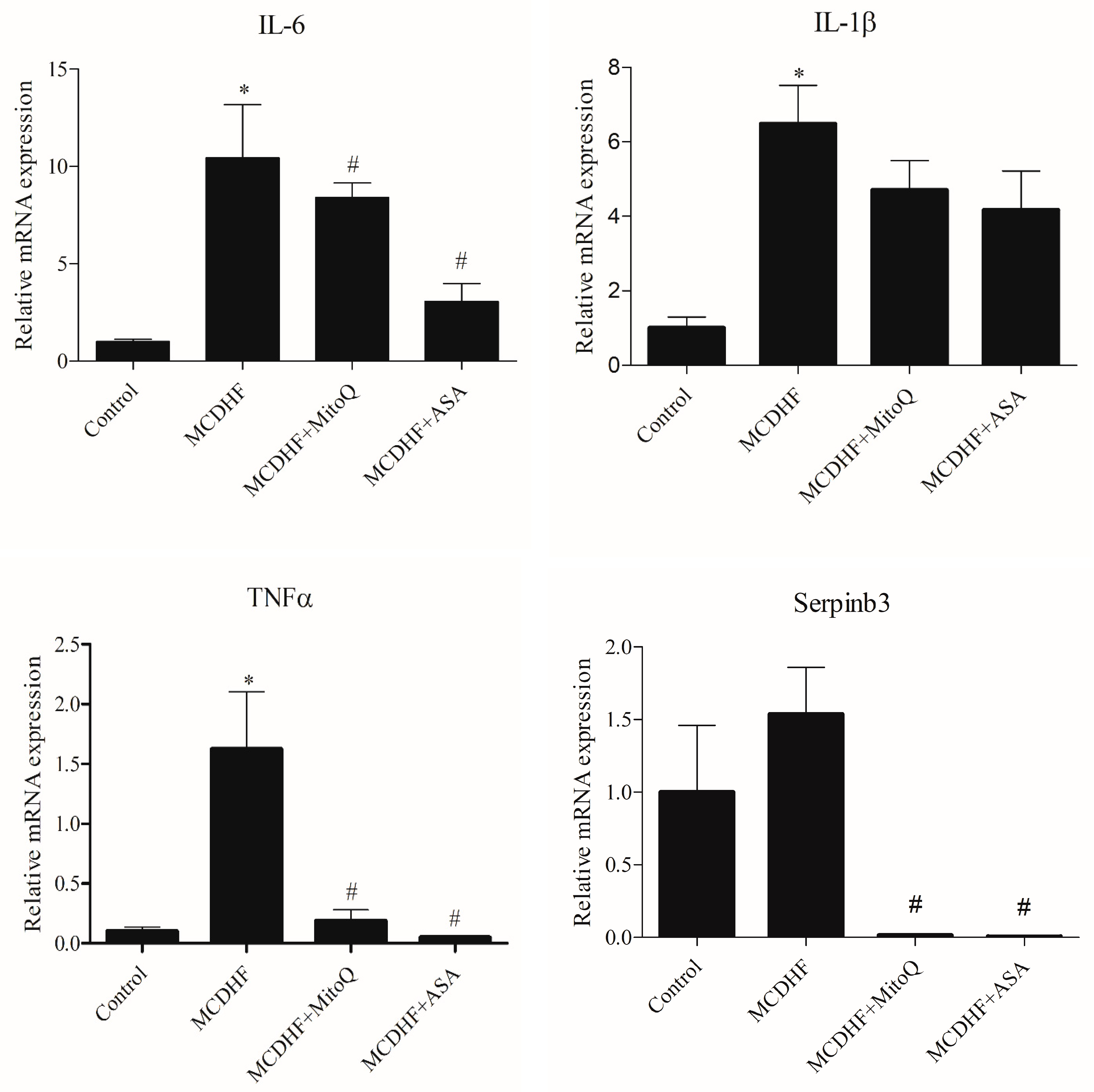
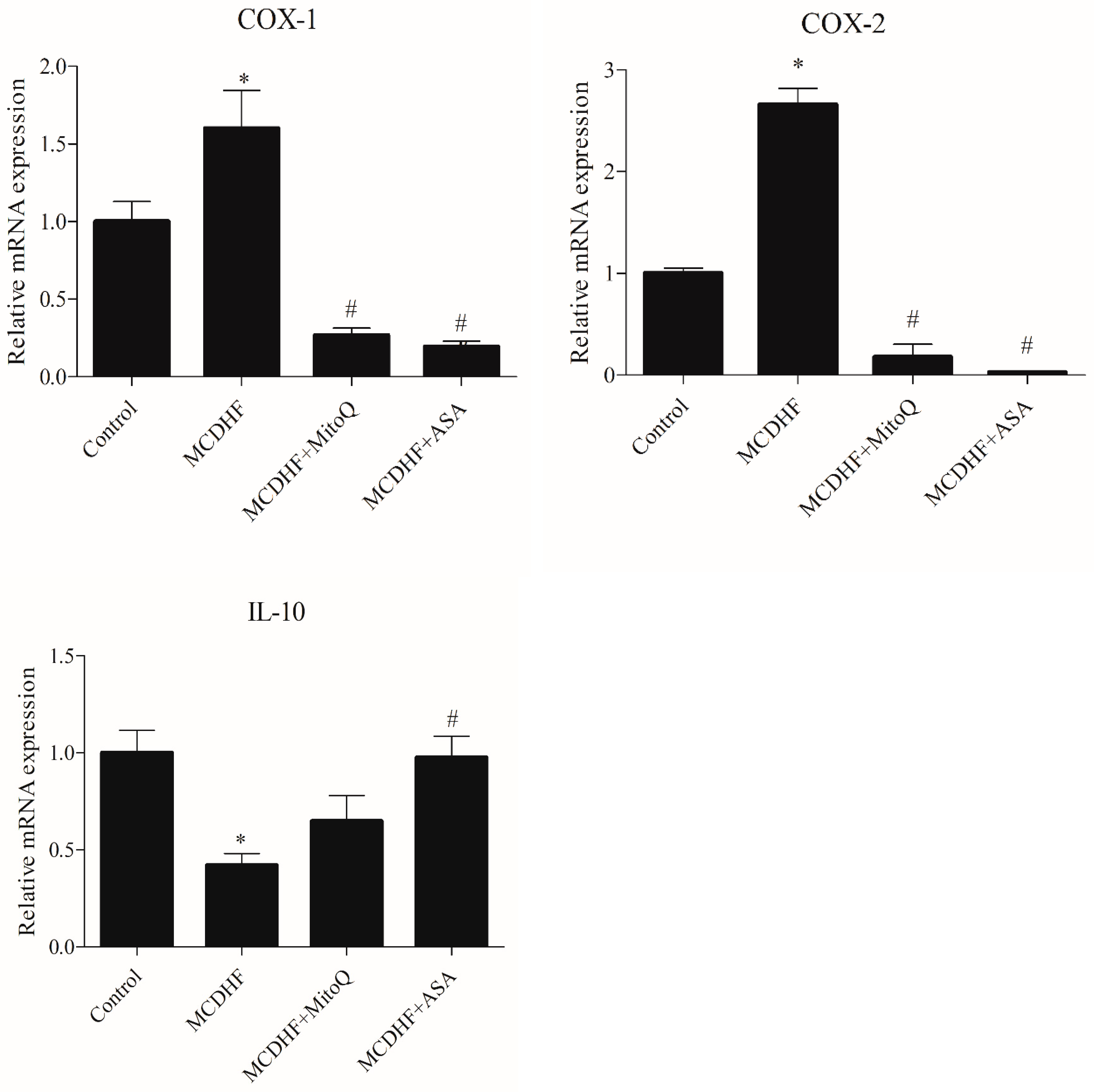
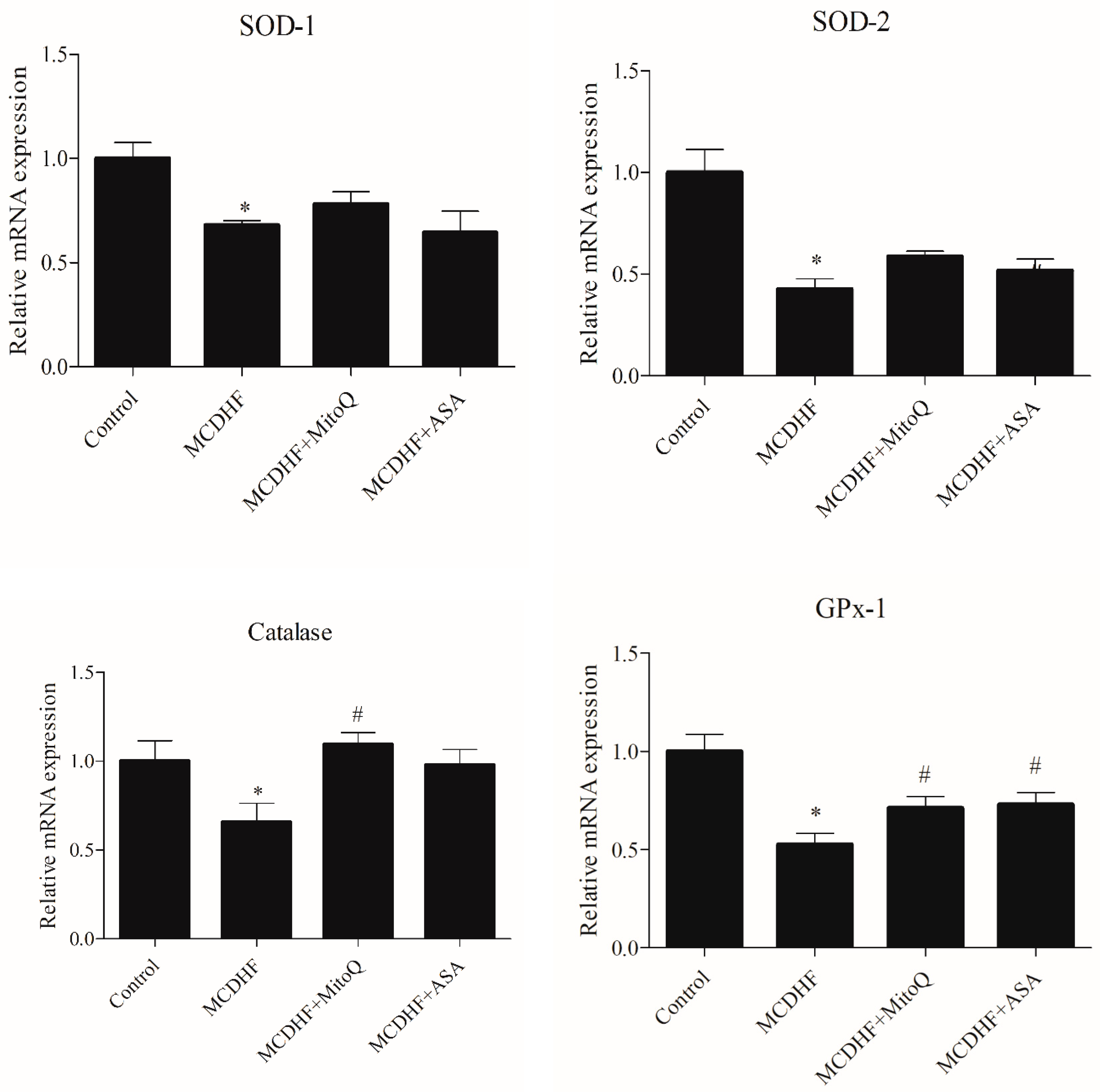
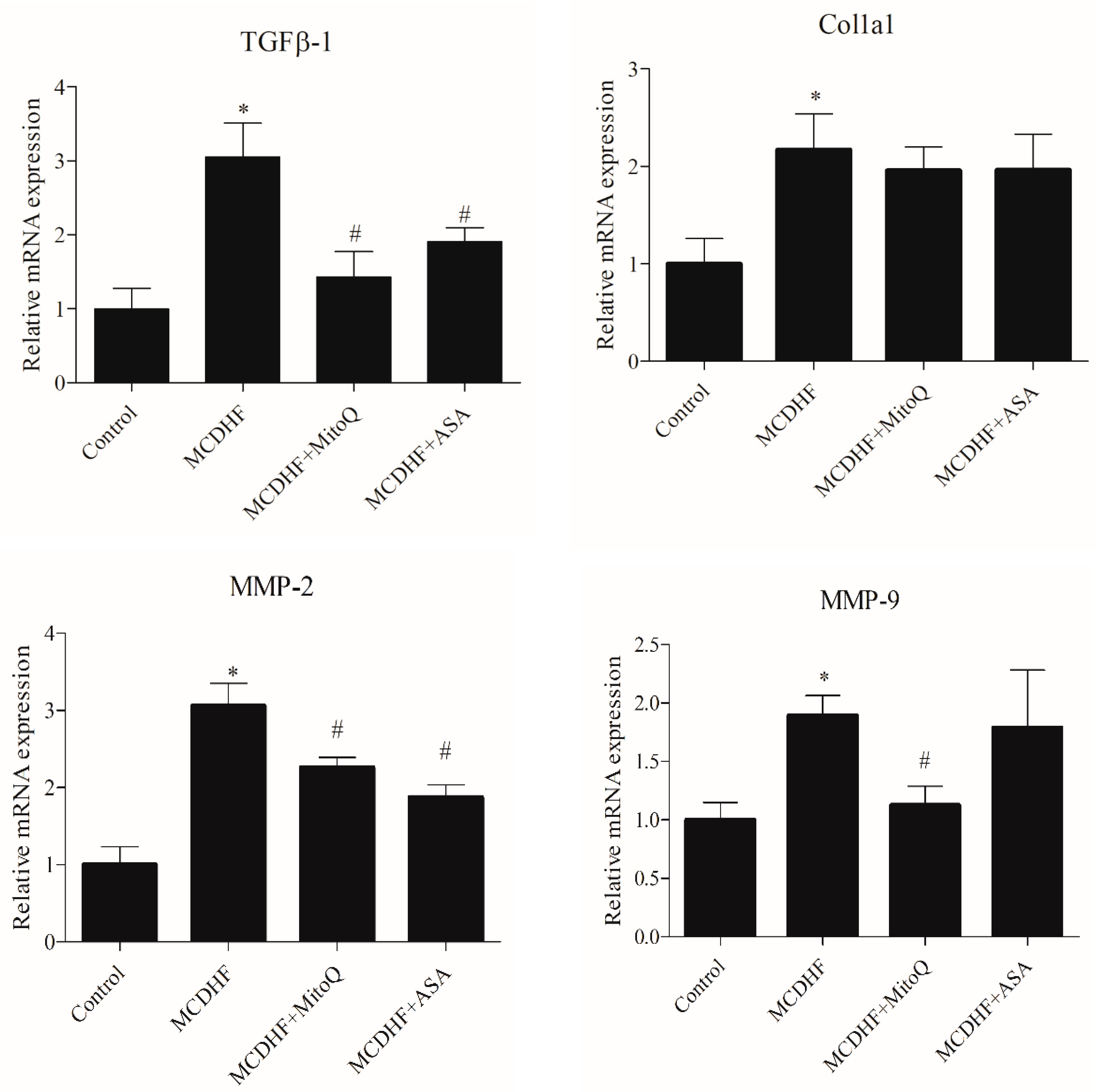
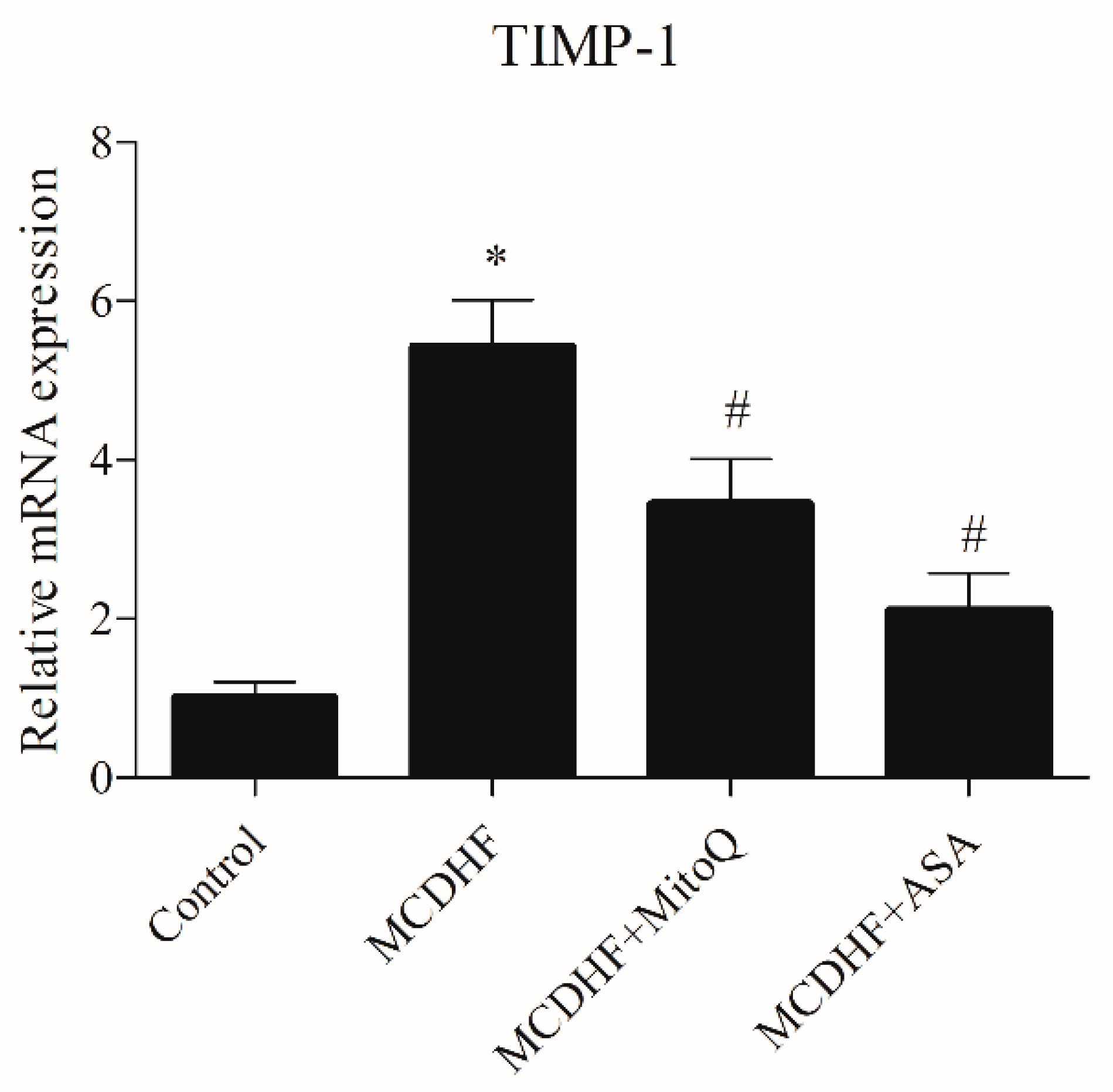
3.2. Effects of ASA and MitoQ on Liver Expression of Genes Associated with Inflammation, Oxidative Stress, and Fibrosis
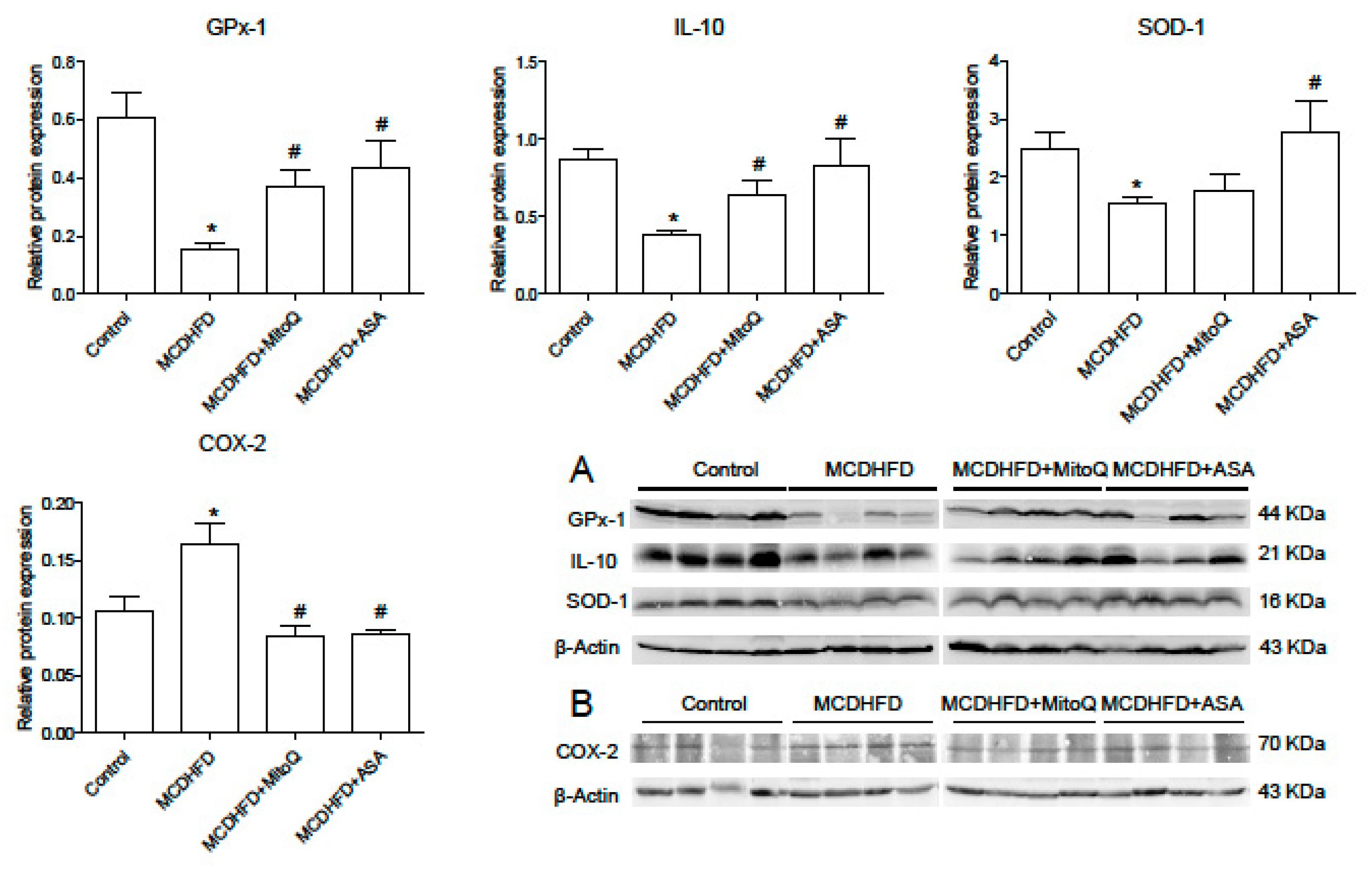
3.3. Effects of ASA and MitoQ on Protein Expression of SOD-1, GPx-1, IL-10, and COX-2 in Liver
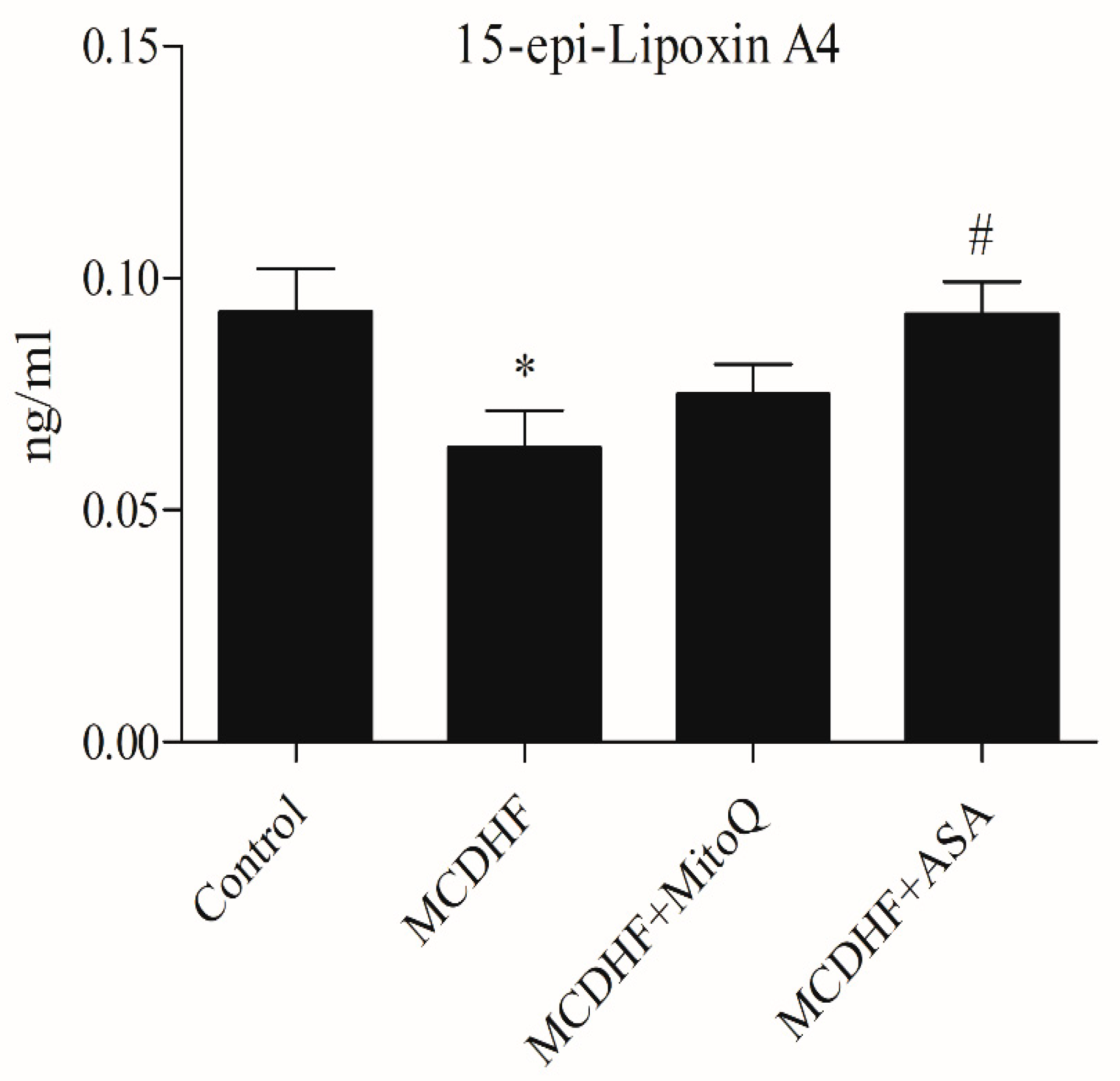
3.4. Effects of ASA and MitoQ on Hepatic and Serum 15-epi-Lipoxin A4
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Day, C.P.; James, O.F. Steatohepatitis: A tale of two “hits”? Gastroenterology 1998, 114, 842–845. [Google Scholar] [CrossRef] [PubMed]
- Tiniakos, D.G.; Vos, M.B.; Brunt, E.M. Nonalcoholic Fatty Liver Disease: Pathology and Pathogenesis. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 145–171. [Google Scholar] [CrossRef] [PubMed]
- deAlmeida, I.T.; Cortez-Pinto, H.; Fidalgo, G.; Rodrigues, D.; Camilo, M.E. Plasma total and free fatty acids composition in human non-alcoholic steatohepatitis. Clin. Nutr. 2002, 21, 219–223. [Google Scholar]
- Nassir, F.; Ibdah, J.A. Role of Mitochondria in Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2014, 15, 8713–8742. [Google Scholar] [CrossRef]
- Ibdah, J.A.; Perlegas, P.; Zhao, Y.; Angdisen, J.; Borgerink, H.; Shadoan, M.K.; Wagner, J.D.; Matern, D.; Rinaldo, P.; Cline, J.M. Mice Heterozygous for a Defect in Mitochondrial Trifunctional Protein Develop Hepatic Steatosis and Insulin Resistance. Gastroenterology 2005, 128, 1381–1390. [Google Scholar] [CrossRef]
- Rector, R.S.; Thyfault, J.P.; Uptergrove, G.M.; Morris, E.M.; Naples, S.P.; Borengasser, S.J.; Mikus, C.R.; Laye, M.J.; Laughlin, M.H.; Booth, F.W.; et al. Mitochondrial dysfunction precedes insulin resistance and hepatic steatosis and contributes to the natural history of non-alcoholic fatty liver disease in an obese rodent model. J. Hepatol. 2010, 52, 727–736. [Google Scholar] [CrossRef]
- Karkucinska-Wieckowska, A.; Simoes, I.C.M.; Kalinowski, P.; Lebiedzinska-Arciszewska, M.; Zieniewicz, K.; Milkiewicz, P.; Górska-Ponikowska, M.; Pinton, P.; Malik, A.N.; Krawczyk, M.; et al. Mitochondria, oxidative stress and nonalcoholic fatty liver disease: A complex relationship. Eur. J. Clin. Investig. 2021, 52, e13622. [Google Scholar] [CrossRef]
- Mao, G.; Kraus, G.A.; Kim, I.; Spurlock, M.E.; Bailey, T.B.; Beitz, D.C. Effect of a mitochondria-targeted vitamin E derivative on mitochondrial alteration and systemic oxidative stress in mice. Br. J. Nutr. 2011, 106, 87–95. [Google Scholar] [CrossRef]
- Menzies, D.; Nair, A.; Meldrum, K.T.; Hopkinson, P.; Lipworth, B.J. Effect of aspirin on airway inflammation and pulmonary function in patients with persistent asthma. J. Allergy Clin. Immunol. 2018, 121, 1184–1189. [Google Scholar] [CrossRef]
- Romano, M.; Cianci, E.; Simiele, F.; Recchiuti, A. Lipoxins and aspirin-triggered lipoxins in resolution of inflammation. Eur. J. Pharmacol. 2015, 760, 49–63. [Google Scholar] [CrossRef]
- Chandrasekharan, J.A.; Sharma-Walia, N. Lipoxins: Nature’s way to resolve inflammation. J. Inflamm. Res. 2015, 8, 181–192. [Google Scholar] [PubMed]
- Nanji, A.A. Animal models of nonalcoholic fatty liver disease and steatohepatitis. Clin. Liver Dis. 2004, 8, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Zhuge, L.; Su, D.; Yang, M.; Tao, S.; Li, J. Comparison between the efficacies of curcumin and puerarin in C57BL/6 mice with steatohepatitis induced by a methionine- and choline-deficient diet. Exp. Ther. Med. 2014, 7, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Gallage, S.; Avila, J.E.B.; Ramadori, P.; Focaccia, E.; Rahbari, M.; Ali, A.; Malek, N.P.; Anstee, Q.M.; Heikenwalder, M. A researcher’s guide to preclinical mouse NASH models. Nat. Metab. 2022, 4, 1632–1649. [Google Scholar] [CrossRef]
- Matsumoto, M.; Hada, N.; Sakamaki, Y.; Uno, A.; Shiga, T.; Tanaka, C.; Ito, T.; Katsume, A.; Sudoh, M. An improved mouse model that rapidly develops fibrosis in non-alcoholic steatohepatitis. Int. J. Exp. Pathol. 2013, 94, 93–103. [Google Scholar] [CrossRef]
- Turkseven, S.; Bolognesi, M.; Brocca, A.; Pesce, P.; Angeli, P.; Di Pascoli, M. Mitochondria-targeted antioxidant mitoquinone attenuates liver inflammation and fibrosis in cirrhotic rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G298–G304. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef]
- Duseja, A.; Singh, S.P.; De, A.; Madan, K.; Rao, P.N.; Shukla, A.; Choudhuri, G.; Saigal, S.; Arora, A.; Anand, A.C.; et al. Indian National Association for Study of the Liver (INASL) Guidance Paper on Nomenclature, Diagnosis and Treatment of Nonalcoholic Fatty Liver Disease (NAFLD). J Clin. Exp. Hepatol. 2023, 13, 273–302. [Google Scholar] [CrossRef]
- Chacko, B.K.; Srivastava, A.; Johnson, M.S.; Benavides, G.A.; Chang, M.J.; Ye, Y.; Jhala, N.; Murphy, M.P.; Kalyanaraman, B.; Darley-Usmar, V.M. Mitochondria-targeted ubiquinone (MitoQ) decreases ethanol-dependent micro and macro hepatosteatosis. Hepatology 2011, 54, 153–163. [Google Scholar] [CrossRef]
- Xu, X.-F.; Fan, J.-W.; Xin, J.-Q.; Wu, N.; Gao, H.; Duan, L.-F.; Zou, W.-B.; Zhang, H.; Li, Z.-S. Aspirin Ameliorates Pancreatic Inflammation and Fibrosis by Inhibiting COX-2 Expression in Experimental Chronic Pancreatitis. J. Inflamm. Res. 2022, ume 15, 4737–4749. [Google Scholar] [CrossRef]
- Wang, P.; Shen, Y.; Zhao, L. Chitosan nanoparticles loaded with aspirin and 5-fluororacil enable synergistic antitumour activity through the modulation of NF-κB/COX-2 signalling pathway. IET Nanobiotechnol. 2020, 14, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Jorda, A.; Aldasoro, M.; Aldasoro, C.; Guerra-Ojeda, S.; Iradi, A.; Vila, J.M.; Campos-Campos, J.; Valles, S.L. Action of low doses of Aspirin in Inflammation and Oxidative Stress induced by aβ1-42 on Astrocytes in primary culture. Int. J. Med. Sci. 2020, 17, 834–843. [Google Scholar] [CrossRef] [PubMed]
- Nakao, S.; Ogtata, Y.; Shimizu, E.; Yamazaki, M.; Furuyama, S.; Sugiya, H. Tumor necrosis factor alpha (TNF-alpha)-induced prostaglandin E2 release is mediated by the activation of cyclooxygenase-2 (COX-2) transcription via NFkappaB in human gingival fibroblasts. Mol. Cell Biochem. 2002, 238, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M.; Yang, C.C.; Hsiao, L.D.; Yu, C.Y.; Tseng, H.C.; Hsu, C.K.; Situmorang, J.H. Upregulation of COX-2 and PGE2 Induced by TNF-α Mediated Through TNFR1/MitoROS/PKCα/P38 MAPK, JNK1/2/FoxO1 Cascade in Human Cardiac Fibroblasts. J. Inflamm. Res. 2021, 145, 2807–2824. [Google Scholar] [CrossRef]
- Yu, J.; Ip, E.; Dela Peña, A.; Hou, J.Y.; Sesha, J.; Pera, N.; Hall, P.; Kirsch, R.; Leclercq, I.; Farrell, G.C. COX-2 induction in mice with experimental nutritional steatohepatitis: Role as pro-inflammatory mediator. Hepatology 2006, 43, 826–836. [Google Scholar] [CrossRef]
- Hui, A.Y.; Dannenberg, A.J.; Sung, J.J.; Subbaramaiah, K.; Du, B.; Olinga, P.; Friedman, S.L. Prostaglandin E2 inhibits transforming growth factor beta 1-mediated induction of collagen alpha 1(I) in hepatic stellate cells. J. Hepatol. 2004, 41, 251–258. [Google Scholar] [CrossRef]
- Motiño, O.; Agra, N.; Contreras, R.B.; Domínguez-Moreno, M.; García-Monzón, C.; Vargas-Castrillón, J.; Carnovale, C.E.; Boscá, L.; Casado, M.; Mayoral, R.; et al. Cyclooxygenase-2 expression in hepatocytes attenuates non-alcoholic steatohepatitis and liver fibrosis in mice. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2016, 1862, 1710–1723. [Google Scholar] [CrossRef]
- Fujita, K.; Nozaki, Y.; Wada, K.; Yoneda, M.; Endo, H.; Takahashi, H.; Iwasaki, T.; Inamori, M.; Abe, Y.; Kobayashi, N.; et al. Effectiveness of antiplatelet drugs against experimental non-alcoholic fatty liver disease. Gut 2008, 57, 1583–1591. [Google Scholar] [CrossRef]
- Turato, C.; Kent, P.; Sebastiani, G.; Cannito, S.; Morello, E.; Terrin, L.; Biasiolo, A.; Simonato, D.; Parola, M.; Pantopoulos, K.; et al. Serpinb3 is overexpressed in the liver in presence of iron overload. J. Investig. Med. 2017, 66, 32–38. [Google Scholar] [CrossRef]
- Novo, E.; Villano, G.; Turato, C.; Cannito, S.; Paternostro, C.; Busletta, C.; Biasiolo, A.; Quarta, S.; Morello, E.; Bocca, C.; et al. SerpinB3 Promotes Pro-fibrogenic Responses in Activated Hepatic Stellate Cells. Sci. Rep. 2017, 7, 3420. [Google Scholar] [CrossRef]
- Terrin, L.; Agostini, M.; Ruvoletto, M.; Martini, A.; Pucciarelli, S.; Bedin, C.; Nitti, D.; Pontisso, P. SerpinB3 upregulates the Cyclooxygenase-2/β-Catenin positive loop in colorectal cancer. Oncotarget 2017, 8, 15732–15743. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TGFβ1 | Sense 5′-TTGCTTCAGCTCCACAGAGA-3′ Rev 5′-TGGTTGTAGAGGGCAAGGAC-3′ |
| Col1a1 | Sense 5′-AAA TCT GCA CAC TGC CAT GA-3′ Rv 5′-GCA TGT TCG AAA TCC AGT GA-3′ |
| SOD-1 | Sense 5′-AACCATCCACTTCGAGCAGA-3′ Rev 5′-TACTGATGGACGTGGAACCC-3′ |
| SOD-2 | Sense 5′-GCCTGCTCTAATCAGGACCC-3′ Rev 5′-GTAGTAAGCGTGCTCCCACA-3′ |
| Catalase | Sense 5′-CACTGACGAGATGGCACACT-3′ Rev 5′-TGTGGAGAATCGAACGGCAA-3′ |
| TNFα | Sense 5′-AGCCCCCAGTCTGTATCCTT-3′ Rev 5′-CTCCCTTTGCAG AACTCAGG-3′ |
| GPx-1 | Sense 5′-AGTCCACCGTGTATGCCTTC-3′ Rev 5′-CCTCAGAGAGACGCGACATT-3′ |
| IL-6 | Sense 5′-AGTTGCCTTCTTGGGACTGA-3′ Rev 5′-CAGAATTGCCATTGCACAAC-3′ |
| IL-1β | Sense 5′-GAAATGCCACCTTTTGACAGTGAT-3′ Rev 5′-TTGGAAGCAGCCCTTCATCTT-3′ |
| Serpinb3 | Sense 5′-TCCTAGTGGGAGCCTAAGCA-3′ Rev 5′-ATCCCCCAGAAAGCTGAAGT-3′ |
| COX-1 | Sense 5′-TCTGCCTCAACACCAAGACC-3′ Rev 5′-AGACAGACCCGTCATCTCCA-3′ |
| COX-2 | Sense 5′-TATGCCACCATCTGGCTTCG-3′ Rev 5′-GTTGCTCATCACCCCACTCA-3′ |
| IL-10 | Sense 5′-CCAAGCCTTATCGGAAATGA-3′ Rev 5′-TTTTCACAGGGGAGAAATCG-3′ |
| MMP-2 | Sense 5′-CCA ACT ACA ACT TCT TCC CCC-3′ Rev 5′-CGA GCA AAA GCA TCA TCC AC-3′ |
| MMP-9 | Sense 5′-CCCTGGAACTCACACGACAT-3′ Rev 5′-TGGTTCACCTCATGGTCCAC-3′ |
| TIMP-1 | Sense 5′-ATGCCCACAAGTCCCAGAAC-3′ Rev 5′-TACGCCAGGGAACCAAGAAG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turkseven, S.; Turato, C.; Villano, G.; Ruvoletto, M.; Guido, M.; Bolognesi, M.; Pontisso, P.; Di Pascoli, M. Low-Dose Acetylsalicylic Acid and Mitochondria-Targeted Antioxidant Mitoquinone Attenuate Non-Alcoholic Steatohepatitis in Mice. Antioxidants 2023, 12, 971. https://doi.org/10.3390/antiox12040971
Turkseven S, Turato C, Villano G, Ruvoletto M, Guido M, Bolognesi M, Pontisso P, Di Pascoli M. Low-Dose Acetylsalicylic Acid and Mitochondria-Targeted Antioxidant Mitoquinone Attenuate Non-Alcoholic Steatohepatitis in Mice. Antioxidants. 2023; 12(4):971. https://doi.org/10.3390/antiox12040971
Chicago/Turabian StyleTurkseven, Saadet, Cristian Turato, Gianmarco Villano, Mariagrazia Ruvoletto, Maria Guido, Massimo Bolognesi, Patrizia Pontisso, and Marco Di Pascoli. 2023. "Low-Dose Acetylsalicylic Acid and Mitochondria-Targeted Antioxidant Mitoquinone Attenuate Non-Alcoholic Steatohepatitis in Mice" Antioxidants 12, no. 4: 971. https://doi.org/10.3390/antiox12040971