
CoQ10Phytosomes Improve Cellular Ubiquinone Uptake in Skeletal Muscle Cells: An Ex Vivo Study Using CoQ10-Enriched Low-Density Lipoproteins Obtained in a Randomized Crossover Study
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
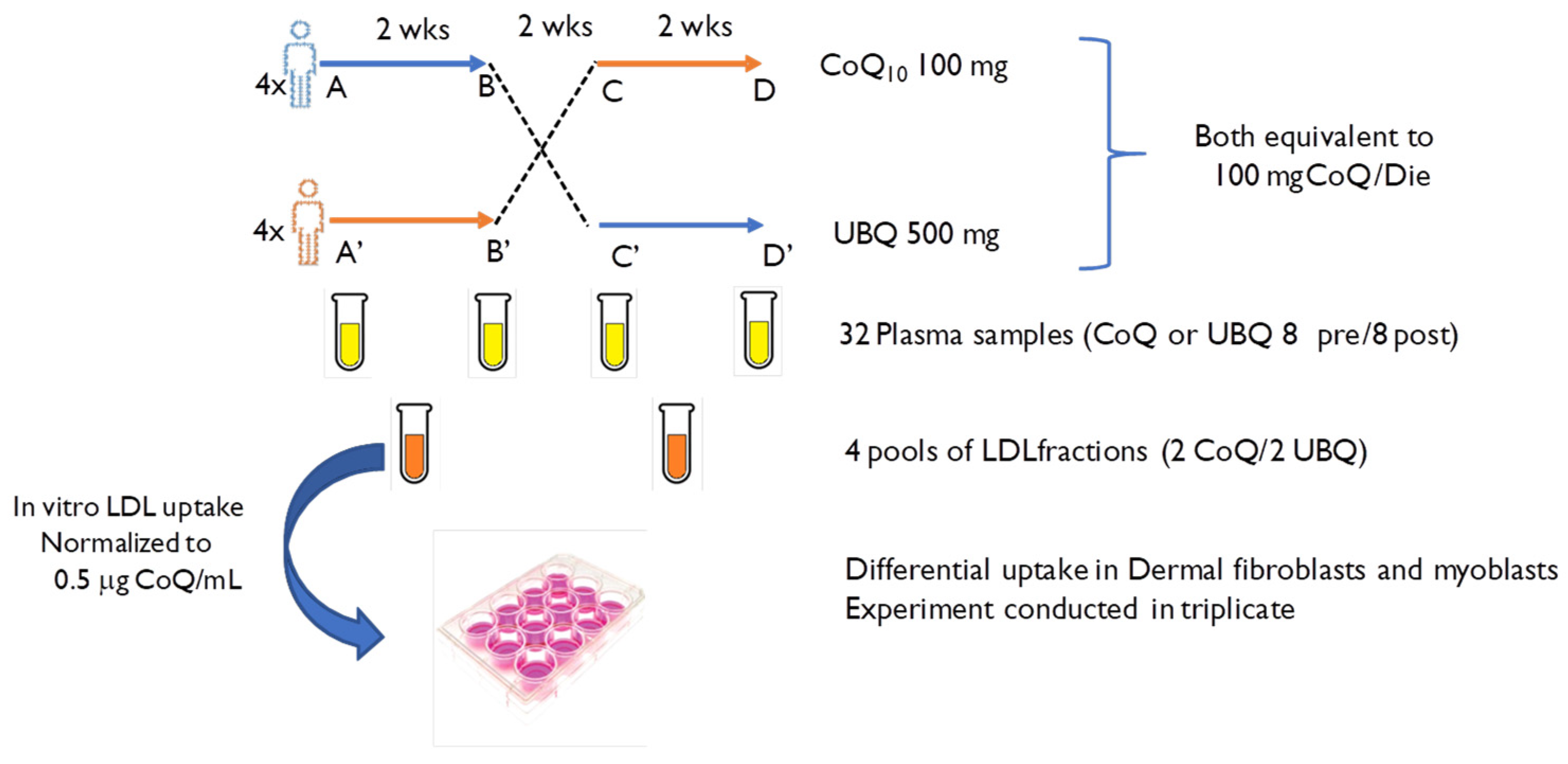
2.1. Experimental Design
2.2. Blood Samples and Extraction of Enriched LDL from Plasma
2.3. Cell Culture and LDL Treatment
2.4. Coenzyme Q10Level and Its Oxidative Status Determination in Plasma and Cell Samples
2.5. Sample Size and Statistical Analysis
3. Results
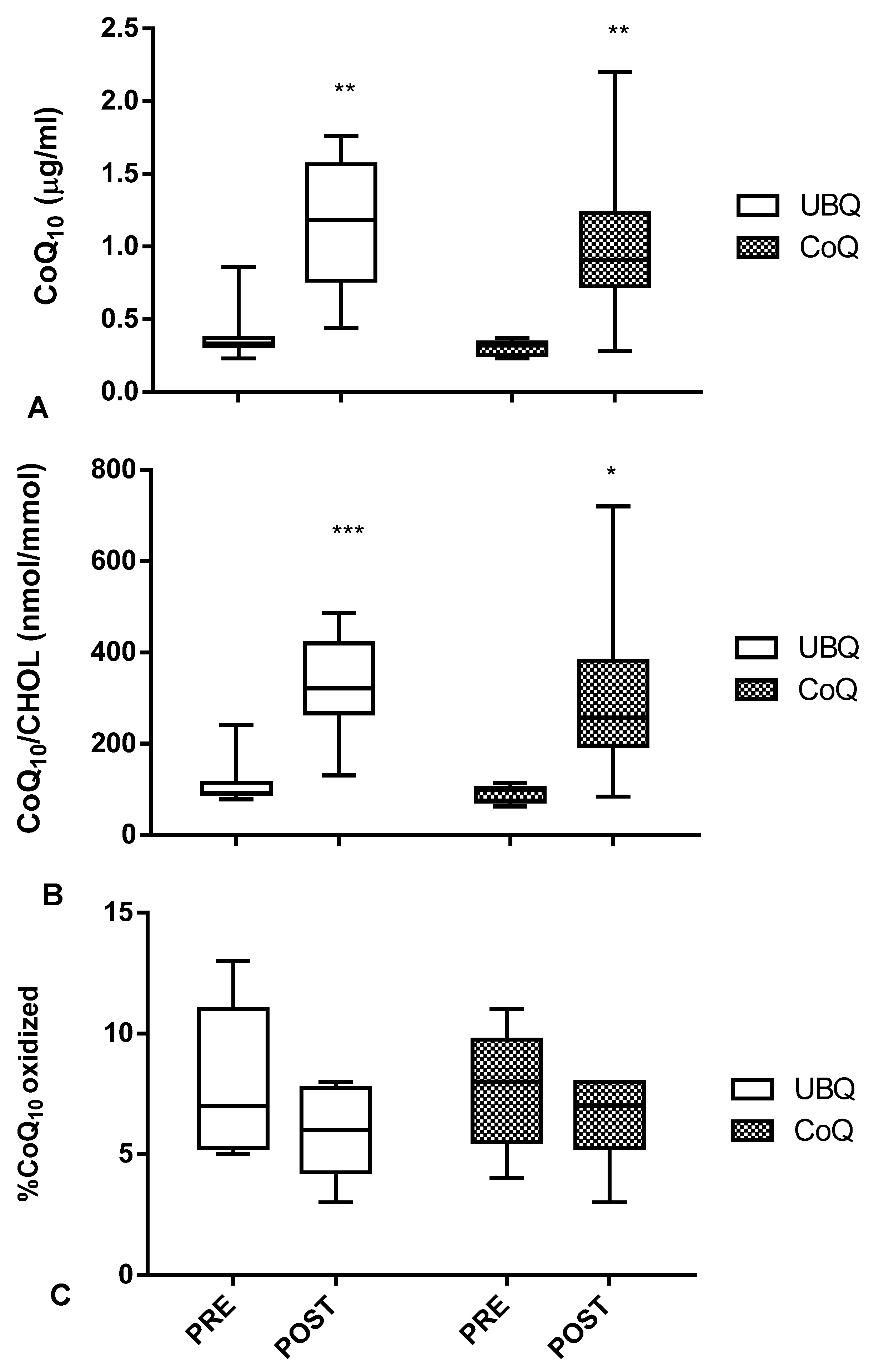
3.1. Both Crystalline CoQ10 and Ubiqsome Supplementation Produced a Significant Increase in CoQ10Plasma Levelsand Improved Its Oxidative Status
3.2. CoQ10-Enriched LDL from Ubiqsome Supplemented Subjects Are More Efficient Vectors of CoQ10 to Cultured Cells
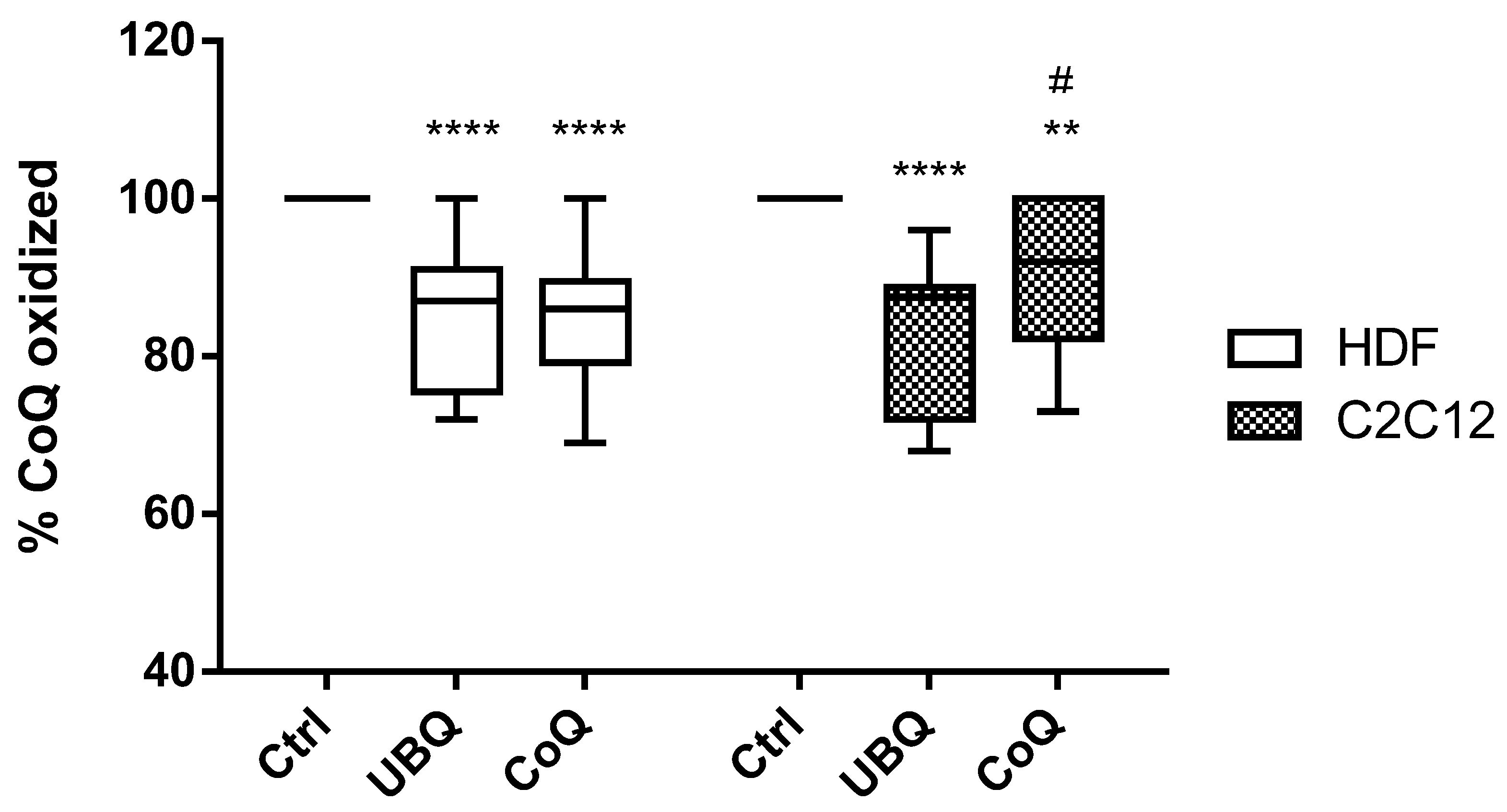
3.3. Ubiqsome Enriched LDL Were More Effective in Improving Cellular CoQ10Oxidative Status in Murine Myoblasts
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lenaz, G.; Genova, M.L. Structure and organization of mitochondrial respiratory complexes: A new understanding of an old subject. Antioxid. Redox Signal. 2010, 12, 961–1008. [Google Scholar] [CrossRef] [PubMed]
- Bentinger, M.; Brismar, K.; Dallner, G. The antioxidant role of coenzyme Q. Mitochondrion 2007, 7, 41–50. [Google Scholar] [CrossRef]
- Navas, P.; Villalba, J.M.; Cabo, R.D. The importance of plasma membrane coenzyme Q in aging and stress responses. Mitochondrion 2007, 7, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Cabo, R.D.; Cabello, R.; Rios, M.; Lopez-Lluch, G.; Ingram, D.K.; Lane, M.A.; Navas, P. Calorie restriction attenuates age-related alterations in the plasma membrane antioxidant system in rat liver. Exp. Gerontol. 2004, 39, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Echtay, K.S.; Winkler, E.; Klingenberg, M. Coenzyme Q is an obligatory cofactor for uncoupling protein function. Nature 2000, 408, 609–613. [Google Scholar] [CrossRef]
- Sluse, F.E.; Jarmuszkiewicz, W.; Navet, R.; Douette, P.; Mathy, G.; Sluse-Goffart, C.M. Mitochondrial UCPs: New insights into regulation and impact. Biochim. Biophys. Acta 2006, 1757, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Belliere, J.; Devun, F.; Cottet-Rousselle, C.; Batandier, C.; Leverve, X.; Fontaine, E. Prerequisites for ubiquinone analogs to prevent mitochondrial permeability transition-induced cell death. J. Bioenerg. Biomembr. 2012, 44, 207–212. [Google Scholar] [CrossRef]
- Molyneux, S.L.; Florkowski, C.M.; George, P.M.; Pilbrow, A.P.; Frampton, C.M.; Lever, M.; Richards, A.M. Coenzyme Q10: An independent predictor of mortality in chronic heart failure. J. Am. Coll. Cardiol. 2008, 52, 1435–1441. [Google Scholar] [CrossRef]
- Cirilli, I.; Damiani, E.; Dludla, P.; Hargreaves, I.; Marcheggiani, F.; Millichap, L.E.; Orlando, P.; Silvestri, S.; Tiano, L. Role of Coenzyme Q10 in Health and Disease: An Update on the Last 10 Years (2010–2020). Antioxidants 2021, 10, 1325. [Google Scholar] [CrossRef]
- Kon, M.; Tanabe, K.; Akimoto, T.; Fuminori, K.; Yuko, T.; Kazuhiro, S.; Tadashi, O.; Ichiro, K. Reducing exercise-induced muscular injury in kendo athletes with supplementation of coenzyme Q10. Br. J. Nutr. 2008, 100, 903–909. [Google Scholar] [CrossRef]
- Cooke, M.; Iosia, M.; Buford, T.; Shelmadine, B.; Hudson, G.; Kerksick, C.; Rasmussen, C.; Greenwood, M.; Leutholtz, B.; Willoughby, D.; et al. Effects of acute and 14-day coenzyme Q10 supplementation on exercise performance in both trained and untrained individuals. J. Int. Soc. Sport. Nutr. 2008, 4, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Orlando, P.; Silvestri, S.; Galeazzi, R.; Antonicelli, R.; Marcheggiani, F.; Cirilli, I.; Bacchetti, T.; Tiano, L. Effect of ubiquinol supplementation on biochemical and oxidative stress indexes after intense exercise in youngathletes. Redox Rep. 2018, 23, 136–145. [Google Scholar] [CrossRef]
- Marcheggiani, F.; Kordes, S.; Cirilli, I.; Orlando, P.; Silvestri, S.; Vogelsang, A.; Möller, N.; Blatt, T.; Weise, J.M.; Damiani, E.; et al. Anti-ageing effects of ubiquinone and ubiquinol in a senescence model of human dermal fibroblasts. Free Radic. Biol. Med. 2021, 165, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Fazakerley, D.J.; Chaudhuri, R.; Yang, P.; Maghzal, G.J.; Thomas, K.C.; Krycer, J.R.; Humphrey, S.J.; Parker, B.L.; Fisher-Wellman, K.H.; Meoli, C.C.; et al. Mitochondrial CoQ deficiency is a common driver of mitochondrial oxidants and insulin resistance. Elife 2018, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Miyazaki, T.; Takagi, A.; Sugita, Y.; Yatsu, S.; Murata, A.; Kato, T.; Suda, S.; Ouchi, S.; Aikawa, T.; et al. Low circulating coenzyme Q10 during acutephase is associated with inflammation, malnutrition, and in-hospital mortality in patients admitted to the coronary careunit. Heart Vessel. 2017, 32, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Pravst, I.; Žmitek, K.; Žmitek, J. Coenzyme Q10 Contents in Foods and Fortification Strategies. Crit. Rev. Food Sci. Nutr. 2010, 50, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Bhagavan, H.N.; Chopra, R.K. Coenzyme Q10: Absorption, tissue uptake, metabolism and pharmacokinetics. Free Radic. Res. 2006, 40, 445–453. [Google Scholar] [CrossRef]
- Bentinger, M.; Dallner, G.; Chojnacki, T.; Swiezewska, E. Distribution and breakdown of labeled coenzyme Q10 in rat. Free Radic. Biol. Med. 2003, 34, 563–575. [Google Scholar] [CrossRef]
- Zhang, Y.; Åberg, F.; Appelkvist, E.L.; Dallner, G.; Ernster, L. Uptake of dietary coenzyme Q supplement is limited in rats. J. Nutr. 1995, 125, 446–453. [Google Scholar]
- Zhang, Y.; Turunen, M.; Appelkvist, E.L. Restricted uptake of dietary coenzyme Q is in contrast to the unrestricted uptake of alpha-tocopherol into rat organs and cells. J. Nutr. 1996, 26, 2089–2097. [Google Scholar] [CrossRef]
- Drobnic, F.; Riera, J.; Artuch, V.; Jou, C.; Codina, A.; Montero, R.; Paredes-Fuentes, A.J.; Domingo, J.C.; Banquells, M.; Riva, A.; et al. Efficient Muscle Distribution Reflects the Positive Influence of Coenzyme Q10 Phytosome in Healthy Aging Athletes after Stressing Exercise. J. Food Sci. Nutr. Res. 2020, 3, 262–275. [Google Scholar] [CrossRef]
- Weiland, H.; Seidel, D. A simple specific method for precipitation of low density lipoproteins. J. Lipid Res. 1983, 24, 904–909. [Google Scholar] [CrossRef]
- Andreani, C.; Bartolacci, C.; Guescini, M.; Battistelli, M.; Stocchi, V.; Orlando, F.; Provinciali, M.; Amici, A.; Marchini, C.; Tiano, L.; et al. Combination of Coenzyme Q(10) Intake and Moderate Physical Activity Counteracts Mitochondrial Dysfunctions in a SAMP8 Mouse Model. Oxidative Med. Cell. Longev. 2018, 2018, 8936251. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Onur, S.; Niklowitz, P.; Menke, T.; Laudes, M.; Rimbach, G.; Döring, F. Coenzyme Q10 status as a determinant of muscular strength in two independent cohorts. PLoS ONE 2016, 11, e0167124. [Google Scholar] [CrossRef]
- Martinefski, M.; Samassa, P.; Buontempo, F.; Hocht, C.; Lucangioli, S.; Tripodi, V. Relative bioavailability of coenzyme Q10 formulation for paediatric individualized therapy. J. Pharm. Pharmacol. 2016, 69, 567–573. [Google Scholar] [CrossRef]
- Langsjoen, P.H.; Langsjoen, A.M. Overview of the use of CoQ10 in cardiovascular disease. Biofactors 1999, 2, 273–284. [Google Scholar] [CrossRef]
- La Guardia, P.G.; Alberici, L.C.; Ravagnani, F.G.; Catharino, R.R.; Vercesi, A.E. Protection of rat skeletal muscle fibers by either L-carnitine or coenzyme Q10 against statins toxicity mediated by mitochondrial reactive oxygen generation. Front. Physiol. 2013, 4, 103. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.W.; Jabbour, A.; Hayward, C.S.; Furlong, T.J.; Girgis, L.; Macdonald, P.S.; Keogh, A.M. Potential role of coenzyme Q10 in facilitating recovery from statin-induced rhabdomyolysis. Intern. Med. J. 2015, 45, 451–453. [Google Scholar] [CrossRef]
- Lopez-Lluch, G.; Rodriguez-Aguilera, J.C.; Santos-Ocana, C.; Navas, P. Is coenzyme Q a key factor in aging? Mech. Ageing Dev. 2010, 131, 225–235. [Google Scholar] [CrossRef]
- Olivieri, F.; Lazzarini, R.; Babini, L.; Prattichizzo, F.; Rippo, M.R.; Tiano, L.; Di Nuzzo, S.; Graciotti, L.; Festa, R.; Bruge, F.; et al. Anti-inflammatory effect of ubiquinol-10 on young and senescent endothelial cells via miR-146a modulation. Free Radic. Biol. Med. 2013, 63, 410–420. [Google Scholar] [CrossRef]
- Lee, B.J.; Tseng, Y.F.; Yen, C.H.; Lin, P.T. Effects of coenzyme Q10 supplementation (300 mg/day) on antioxidation and antiinflammation in coronary artery disease patients during statins therapy: A randomized, placebo-controlled trial. Nutr. J. 2013, 12, 142. [Google Scholar] [CrossRef] [PubMed]
- Stone, L.H.; Chappuis, E.; Wright, C.; Kelly, R.F.; McFalls, O.E. CoQ10 enhances PGC1α and increases expression of mitochondrial antioxidant proteins in chronically ischemic swine myocardium. Nutr. Metab. 2019, 16, 92. [Google Scholar] [CrossRef]
- Bergamini, C.; Moruzzi, N.; Sblendido, A.; Lenaz, G.; Fato, R. A Water Soluble CoQ10 Formulation Improves Intracellular Distribution and Promotes Mitochondrial Respiration in Cultured Cells. PLoS ONE 2012, 7, e33712. [Google Scholar] [CrossRef]
- Orlando, P.; Sabbatinelli, J.; Silvestri, S.; Marcheggiani, F.; Cirilli, I.; Dludla, P.V.; Molardi, A.; Nicolini, F.; Tiano, L. Ubiquinol supplementation in elderly patients undergoing aortic valve replacement: Biochemical and clinical aspects. Aging 2020, 12, 15514–15531. [Google Scholar] [CrossRef]
- Rosenfeldt, F.L.; Pepe, S.; Linnane, A.; Nagley, P.; Rowland, M.; Ou, R.; Marasco, S.; Lyon, W.; Esmore, D. Coenzyme Q10 protects the aging heart against stress: Studies in rats, human tissues, and patients. Ann. N. Y. Acad. Sci. 2002, 959, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Kamzalov, S.; Sumien, N.; Forster, M.J.; Sohal, R.S. Coenzyme Q intake elevates the mitochondrial and tissue levels of Coenzyme Q and alpha-tocopherol in young mice. J. Nutr. 2003, 133, 3175–3180. [Google Scholar] [CrossRef]
- Zaki, N.M. Strategies for oral delivery and mitochondrial targeting of CoQ10. Drug Deliv. 2016, 6, 1868–1881. [Google Scholar] [CrossRef]
- Zmitek, J.; Zmitek, K.; Pravst, I. Improving the bioavailability of coenzyme Q10: From theory to practice. Agro-Food Ind. 2008, 19, 8–10. [Google Scholar]
- Xia, S.; Xu, S.; Zhang, X.; Zhong, F.; Wang, Z. Nanoliposomes mediate coenzyme Q10 transport and accumulation across human intestinal Caco-2 cell monolayer. J. Agric. Food Chem. 2009, 57, 7989–7996. [Google Scholar] [CrossRef]
- Petrangolini, G.; Ronchi, M.; Frattini, E.; De Combarieu, E.; Allegrini, P.; Riva, A. A New Food-grade Coenzyme Q10 Formulation Improves Bioavailability: Single and Repeated Pharmacokinetic Studies in Healthy Volunteers. Curr. Drug Deliv. 2019, 16, 759–767. [Google Scholar] [CrossRef]
- Rizzardi, N.; Liparulo, I.; Antonelli, G.; Orsini, F.; Riva, A.; Bergamini, C.; Fato, R. Coenzyme Q10 Phytosome Formulation Improves CoQ10 Bioavailability and Mitochondrial Functionality in Cultured Cells. Antioxidants 2021, 10, 927. [Google Scholar] [CrossRef] [PubMed]
- Lenaz, G.; Fato, R.; Castelluccio, C.; Genova, M.L.; Bovina, C.; Estornell, E.; Valls, V.; Pallotti, F.; Castelli, G.P. The function of coenzyme Q in mitochondria. Clin. Investig. 1993, 71, S66–S70. [Google Scholar] [CrossRef] [PubMed]
- Kemmerer, Z.A.; Robinson, K.P.; Schmitz, J.M.; Manicki, M.; Paulson, B.R.; Jochem, A.; Hutchins, P.D.; Coon, J.J.; Pagliarini, D.J. UbiB proteins regulate cellular CoQ distribution in Saccharomyces cerevisiae. Nat. Commun. 2021, 6, 4769. [Google Scholar] [CrossRef] [PubMed]
- Lohman, D.C.; Aydin, D.; Von Bank, H.C.; Smith, R.W.; Linke, V.; Weisenhorn, E.; McDevitt, M.T.; Hutchins, P.; Wilkerson, E.M.; Wancewicz, B.; et al. An Isoprene Lipid-Binding Protein Promotes Eukaryotic Coenzyme Q Biosynthesis. Mol. Cell 2019, 21, 763–774. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | CoQ9 (ng/mg Protein) | CoQ10 (ng/mg Protein) |
|---|---|---|
| Ctrl | 63.5 ± 42.9 | 2.6 ± 0.5 |
| UBQ | 83.3 ± 38.3 | 30.2 ± 12.6 (a;b) |
| CoQ | 68.9 ± 30.1 | 7.9 ± 2.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcheggiani, F.; Orlando, P.; Silvestri, S.; Cirilli, I.; Riva, A.; Petrangolini, G.; Orsini, F.; Tiano, L. CoQ10Phytosomes Improve Cellular Ubiquinone Uptake in Skeletal Muscle Cells: An Ex Vivo Study Using CoQ10-Enriched Low-Density Lipoproteins Obtained in a Randomized Crossover Study. Antioxidants 2023, 12, 964. https://doi.org/10.3390/antiox12040964
Marcheggiani F, Orlando P, Silvestri S, Cirilli I, Riva A, Petrangolini G, Orsini F, Tiano L. CoQ10Phytosomes Improve Cellular Ubiquinone Uptake in Skeletal Muscle Cells: An Ex Vivo Study Using CoQ10-Enriched Low-Density Lipoproteins Obtained in a Randomized Crossover Study. Antioxidants. 2023; 12(4):964. https://doi.org/10.3390/antiox12040964
Chicago/Turabian StyleMarcheggiani, Fabio, Patrick Orlando, Sonia Silvestri, Ilenia Cirilli, Antonella Riva, Giovanna Petrangolini, Francesca Orsini, and Luca Tiano. 2023. "CoQ10Phytosomes Improve Cellular Ubiquinone Uptake in Skeletal Muscle Cells: An Ex Vivo Study Using CoQ10-Enriched Low-Density Lipoproteins Obtained in a Randomized Crossover Study" Antioxidants 12, no. 4: 964. https://doi.org/10.3390/antiox12040964