
Human Microglia Synthesize Neurosteroids to Cope with Rotenone-Induced Oxidative Stress

 , , and
, , and 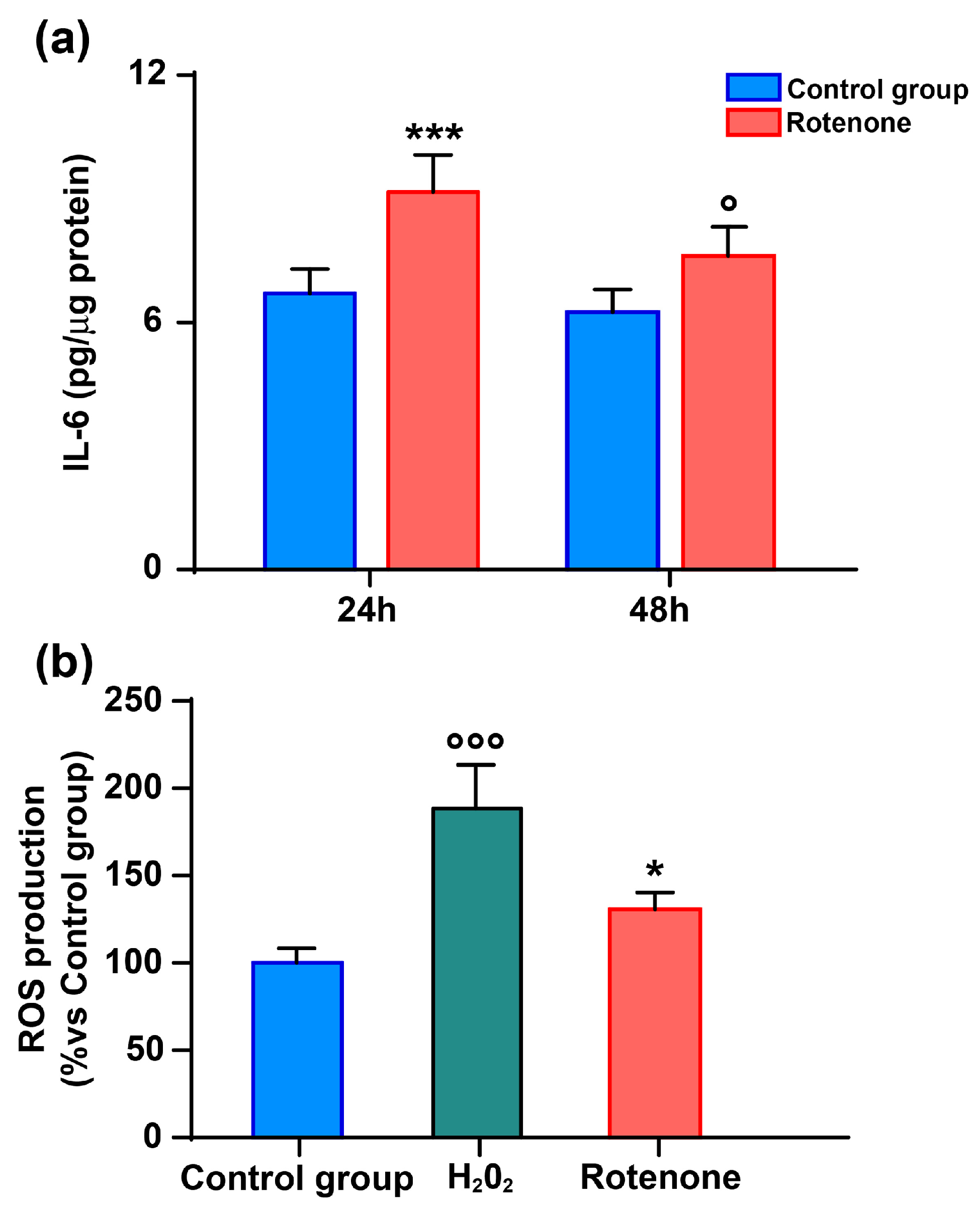{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture
2.3. IL-6 Quantification
2.4. Determination of ROS
2.5. Sample Processing for Neurosteroid Quantification
2.6. Working Solutions and Calibrators
2.7. LC-MS/MS Analysis
2.8. Viability of HMC3 Microglial Cells Treated with Rotenone
2.9. Statistical Analysis
3. Results
3.1. Oxidative Stress Activates HMC3 Microglia to Release IL-6
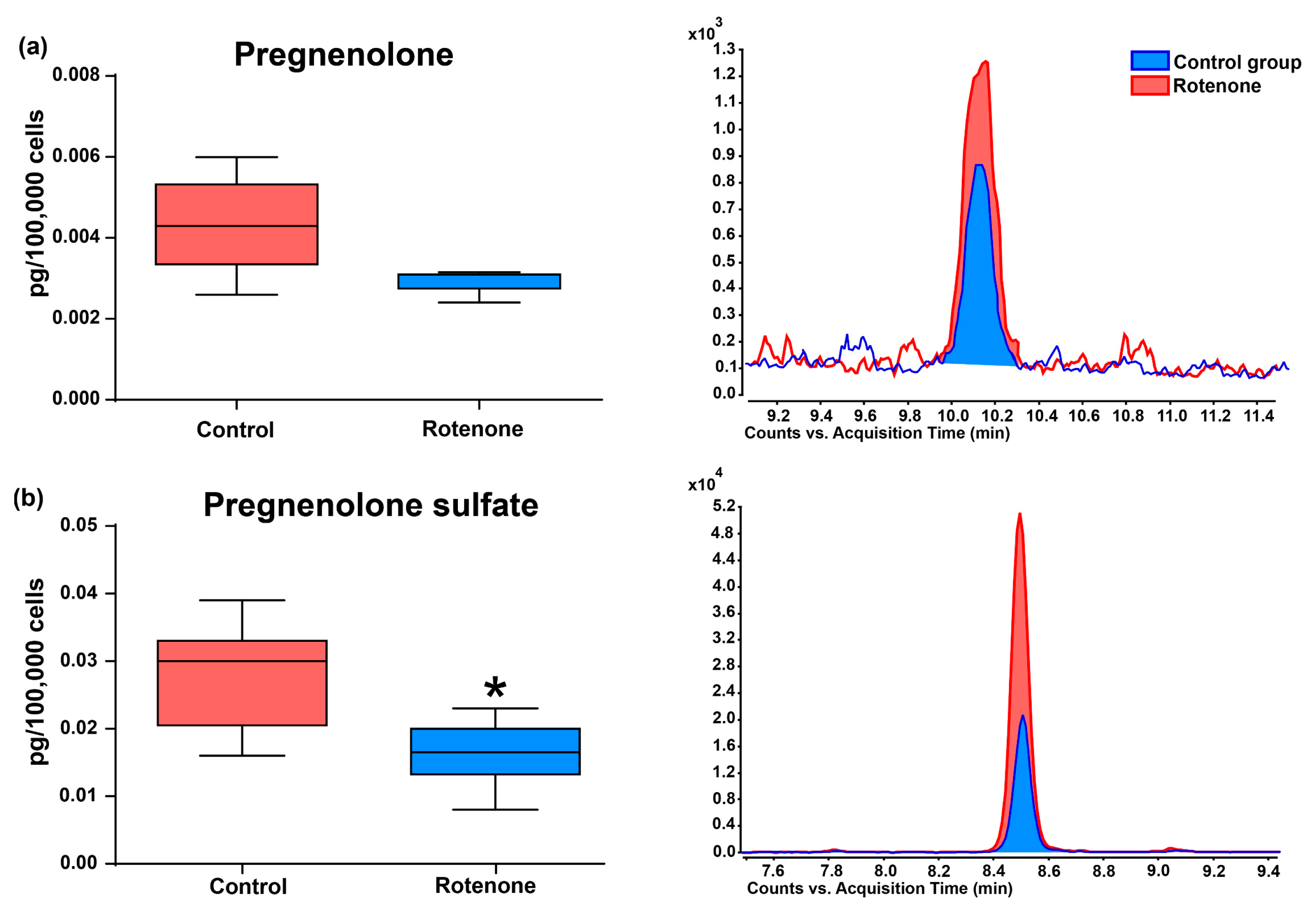
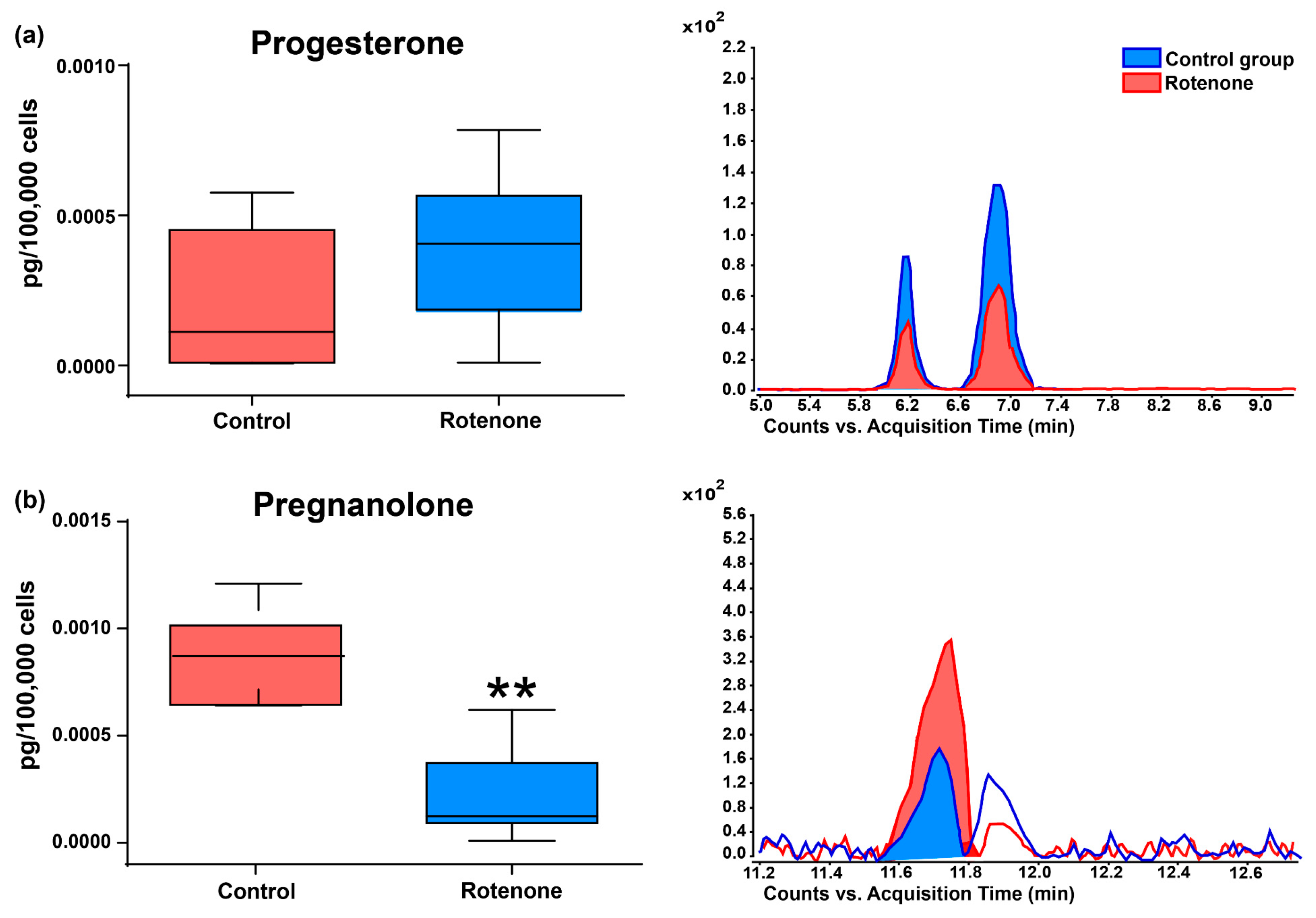
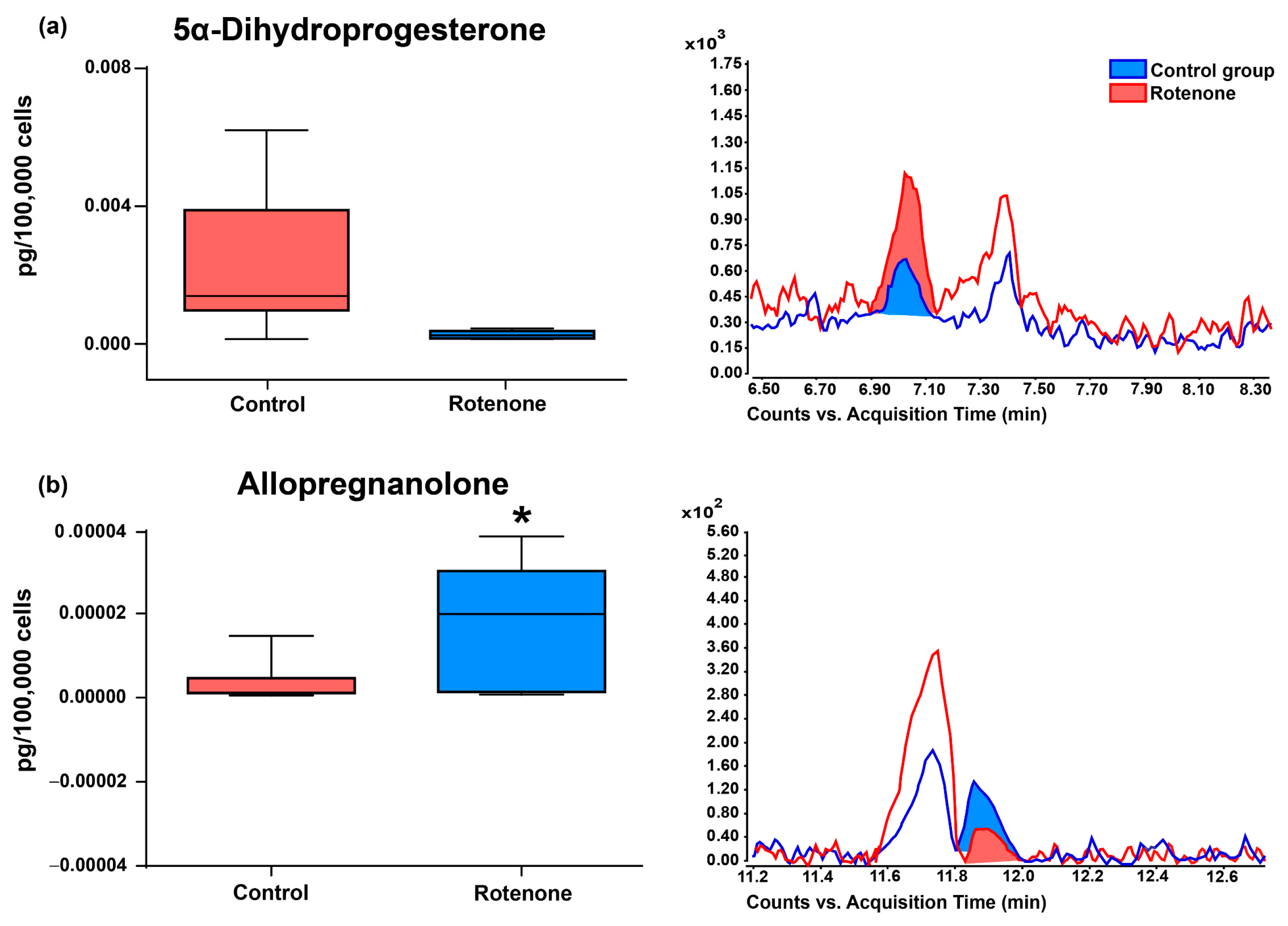
3.2. Effect of Rotenone on Neurosteroid Levels in Culture Medium of HMC3 Cells
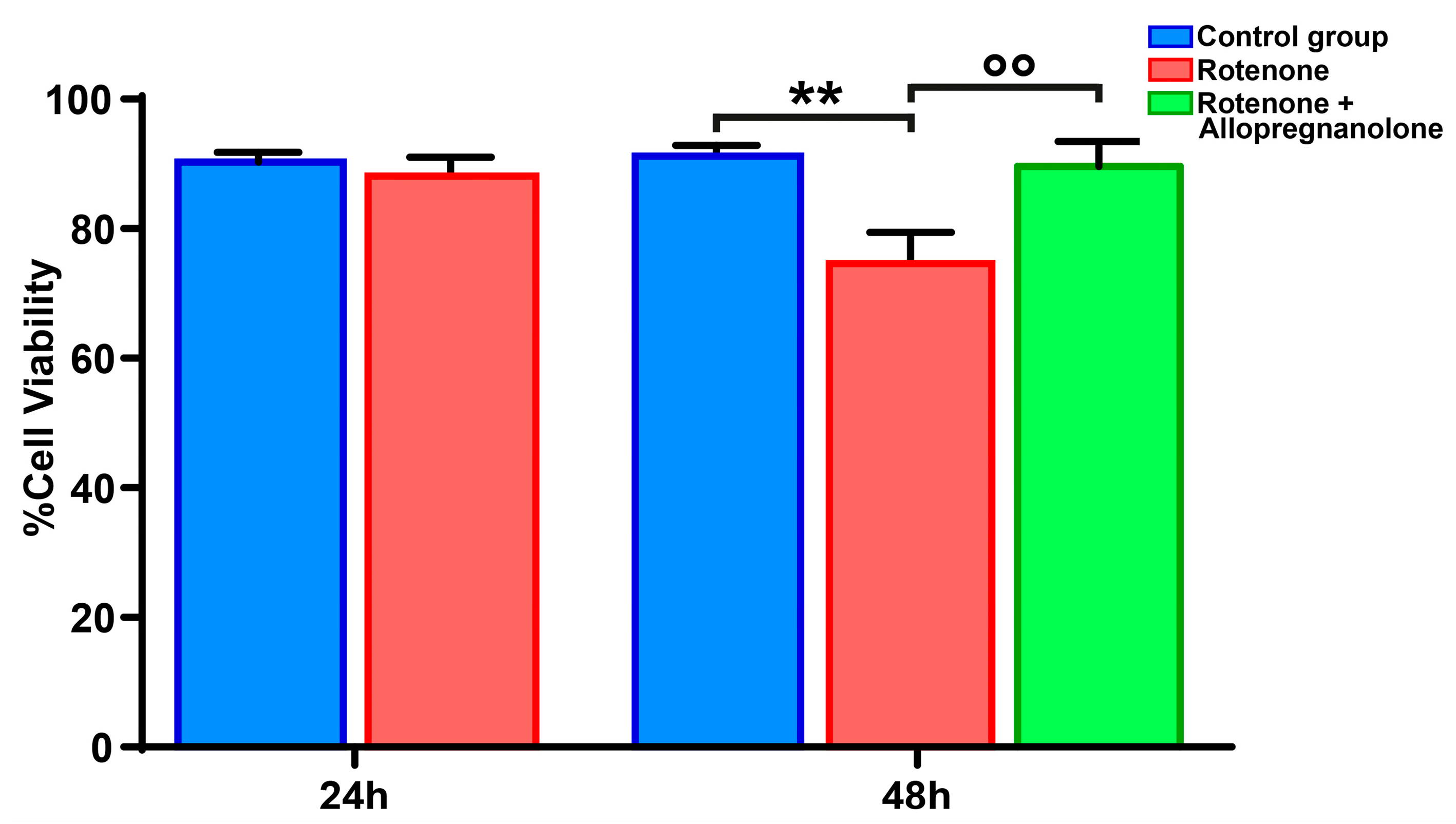
3.3. Exogenous Allopregnanolone Preserved HMC3 Cell Viability from Rotenone-Induced Damage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giatti, S.; Garcia-Segura, L.M.; Barreto, G.E.; Melcangi, R.C. Neuroactive Steroids, Neurosteroidogenesis and Sex. Prog. Neurobiol. 2019, 176, 1–17. [Google Scholar] [CrossRef]
- Gunn, B.G.; Cunningham, L.; Mitchell, S.G.; Swinny, J.D.; Lambert, J.J.; Belelli, D. GABAA Receptor-Acting Neurosteroids: A Role in the Development and Regulation of the Stress Response. Front. Neuroendocrinol. 2015, 36, 28–48. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, C.; Karali, K.; Fodelianaki, G.; Gravanis, A.; Chavakis, T.; Charalampopoulos, I.; Alexaki, V.I. Neurosteroids as Regulators of Neuroinflammation. Front. Neuroendocrinol. 2019, 55, 100788. [Google Scholar] [CrossRef]
- Balan, I.; Beattie, M.C.; O’Buckley, T.K.; Aurelian, L.; Morrow, A.L. Endogenous Neurosteroid (3α, 5α)3-Hydroxypregnan-20-One Inhibits Toll-like-4 Receptor Activation and Pro-Inflammatory Signaling in Macrophages and Brain. Sci. Rep. 2019, 9, 1220. [Google Scholar] [CrossRef]
- Diviccaro, S.; Cioffi, L.; Falvo, E.; Giatti, S.; Melcangi, R.C. Allopregnanolone: An Overview on Its Synthesis and Effects. J. Neuroendocrinol. 2022, 34, e12996. [Google Scholar] [CrossRef] [PubMed]
- Noorbakhsh, F.; Baker, G.B.; Power, C. Allopregnanolone and Neuroinflammation: A Focus on Multiple Sclerosis. Front. Cell. Neurosci. 2014, 8, 134. [Google Scholar] [CrossRef]
- di Michele, F.; Luchetti, S.; Bernardi, G.; Romeo, E.; Longone, P. Neurosteroid and Neurotransmitter Alterations in Parkinson’s Disease. Front. Neuroendocrinol. 2013, 34, 132–142. [Google Scholar] [CrossRef]
- Irwin, R.W.; Solinsky, C.M.; Brinton, R.D. Frontiers in Therapeutic Development of Allopregnanolone for Alzheimer’s Disease and Other Neurological Disorders. Front. Cell. Neurosci. 2014, 8, 203. [Google Scholar] [CrossRef]
- Lévesque, M.; Biagini, G.; Avoli, M. Neurosteroids and Focal Epileptic Disorders. Int. J. Mol. Sci. 2020, 21, 9391. [Google Scholar] [CrossRef]
- Lanthier, A.; Patwardhan, V.V. Sex Steroids and 5-En-3 Beta-Hydroxysteroids in Specific Regions of the Human Brain and Cranial Nerves. J. Steroid Biochem. 1986, 25, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, C.; Fiet, J.; Benais, J.P.; Gueux, B.; Bonete, R.; Villette, J.M.; Gourmel, B.; Dreux, C. Simultaneous Radioimmunoassay of Progesterone, Androst-4-Enedione, Pregnenolone, Dehydroepiandrosterone and 17-Hydroxyprogesterone in Specific Regions of Human Brain. J. Steroid Biochem. 1987, 28, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Bixo, M.; Andersson, A.; Winblad, B.; Purdy, R.H.; Bäckström, T. Progesterone, 5alpha-Pregnane-3,20-Dione and 3alpha-Hydroxy-5alpha-Pregnane-20-One in Specific Regions of the Human Female Brain in Different Endocrine States. Brain Res. 1997, 764, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.C.; Cascio, C.; Papadopoulos, V. Pathways of Neurosteroid Biosynthesis in Cell Lines from Human Brain: Regulation of Dehydroepiandrosterone Formation by Oxidative Stress and Beta-Amyloid Peptide. J. Neurochem. 2000, 74, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Cheung, G.; Porter, E.; Papadopoulos, V. The Neurosteroid Pregnenolone Is Synthesized by a Mitochondrial P450 Enzyme Other than CYP11A1 in Human Glial Cells. J. Biol. Chem. 2022, 298, 102110. [Google Scholar] [CrossRef]
- Stoffel-Wagner, B.; Beyenburg, S.; Watzka, M.; Blümcke, I.; Bauer, J.; Schramm, J.; Bidlingmaier, F.; Elger, C.E. Expression of 5alpha-Reductase and 3alpha-Hydroxisteroid Oxidoreductase in the Hippocampus of Patients with Chronic Temporal Lobe Epilepsy. Epilepsia 2000, 41, 140–147. [Google Scholar] [CrossRef]
- Yu, L.; Romero, D.G.; Gomez-Sanchez, C.E.; Gomez-Sanchez, E.P. Steroidogenic Enzyme Gene Expression in the Human Brain. Mol. Cell. Endocrinol. 2002, 190, 9–17. [Google Scholar] [CrossRef]
- Giatti, S.; Diviccaro, S.; Serafini, M.M.; Caruso, D.; Garcia-Segura, L.M.; Viviani, B.; Melcangi, R.C. Sex Differences in Steroid Levels and Steroidogenesis in the Nervous System: Physiopathological Role. Front. Neuroendocrinol. 2020, 56, 100804. [Google Scholar] [CrossRef]
- Schumacher, M.; Guennoun, R.; Robert, F.; Carelli, C.; Gago, N.; Ghoumari, A.; Gonzalez Deniselle, M.C.; Gonzalez, S.L.; Ibanez, C.; Labombarda, F.; et al. Local Synthesis and Dual Actions of Progesterone in the Nervous System: Neuroprotection and Myelination. Growth Horm. IGF Res. Off. J. Growth Horm. Res. Soc. Int. IGF Res. Soc. 2004, 14, 18–33. [Google Scholar] [CrossRef]
- Lin, Y.C.; Papadopoulos, V. Neurosteroidogenic Enzymes: CYP11A1 in the Central Nervous System. Front. Neuroendocrinol. 2021, 62, 100925. [Google Scholar] [CrossRef]
- Steckelbroeck, S.; Watzka, M.; Reichelt, R.; Hans, V.H.; Stoffel-Wagner, B.; Heidrich, D.D.; Schramm, J.; Bidlingmaier, F.; Klingmüller, D. Characterization of the 5alpha-Reductase-3alpha-Hydroxysteroid Dehydrogenase Complex in the Human Brain. J. Clin. Endocrinol. Metab. 2001, 86, 1324–1331. [Google Scholar] [CrossRef]
- Agís-Balboa, R.C.; Pinna, G.; Zhubi, A.; Maloku, E.; Veldic, M.; Costa, E.; Guidotti, A. Characterization of Brain Neurons That Express Enzymes Mediating Neurosteroid Biosynthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 14602–14607. [Google Scholar] [CrossRef] [PubMed]
- Gottfried-Blackmore, A.; Sierra, A.; Jellinck, P.H.; McEwen, B.S.; Bulloch, K. Brain Microglia Express Steroid-Converting Enzymes in the Mouse. J. Steroid Biochem. Mol. Biol. 2008, 109, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Germelli, L.; Da Pozzo, E.; Giacomelli, C.; Tremolanti, C.; Marchetti, L.; Wetzel, C.H.; Barresi, E.; Taliani, S.; Da Settimo, F.; Martini, C.; et al. De Novo Neurosteroidogenesis in Human Microglia: Involvement of the 18 kDa Translocator Protein. Int. J. Mol. Sci. 2021, 22, 3115. [Google Scholar] [CrossRef] [PubMed]
- Jolivel, V.; Brun, S.; Binamé, F.; Benyounes, J.; Taleb, O.; Bagnard, D.; De Sèze, J.; Patte-Mensah, C.; Mensah-Nyagan, A.-G. Microglial Cell Morphology and Phagocytic Activity Are Critically Regulated by the Neurosteroid Allopregnanolone: A Possible Role in Neuroprotection. Cells 2021, 10, 698. [Google Scholar] [CrossRef]
- Vidal-Itriago, A.; Radford, R.A.W.; Aramideh, J.A.; Maurel, C.; Scherer, N.M.; Don, E.K.; Lee, A.; Chung, R.S.; Graeber, M.B.; Morsch, M. Microglia Morphophysiological Diversity and Its Implications for the CNS. Front. Immunol. 2022, 13, 997786. [Google Scholar] [CrossRef]
- Salter, M.W.; Stevens, B. Microglia Emerge as Central Players in Brain Disease. Nat. Med. 2017, 23, 1018–1027. [Google Scholar] [CrossRef]
- Vainchtein, I.D.; Alsema, A.M.; Dubbelaar, M.L.; Grit, C.; Vinet, J.; van Weering, H.R.J.; Al-Izki, S.; Biagini, G.; Brouwer, N.; Amor, S.; et al. Characterizing Microglial Gene Expression in a Model of Secondary Progressive Multiple Sclerosis. Glia 2023, 71, 588–601. [Google Scholar] [CrossRef]
- Bennett, M.L.; Viaene, A.N. What Are Activated and Reactive Glia and What Is Their Role in Neurodegeneration? Neurobiol. Dis. 2021, 148, 105172. [Google Scholar] [CrossRef]
- Drago, F.; Sautière, P.-E.; Le Marrec-Croq, F.; Accorsi, A.; Van Camp, C.; Salzet, M.; Lefebvre, C.; Vizioli, J. Microglia of Medicinal Leech (Hirudo Medicinalis) Express a Specific Activation Marker Homologous to Vertebrate Ionized Calcium-Binding Adapter Molecule 1 (Iba1/alias Aif-1). Dev. Neurobiol. 2014, 74, 987–1001. [Google Scholar] [CrossRef]
- Jurga, A.M.; Paleczna, M.; Kuter, K.Z. Overview of General and Discriminating Markers of Differential Microglia Phenotypes. Front. Cell. Neurosci. 2020, 14, 198. [Google Scholar] [CrossRef] [PubMed]
- Sanders, L.H.; Timothy Greenamyre, J. Oxidative Damage to Macromolecules in Human Parkinson Disease and the Rotenone Model. Free Radic. Biol. Med. 2013, 62, 111–120. [Google Scholar] [CrossRef]
- Ibarra-Gutiérrez, M.T.; Serrano-García, N.; Orozco-Ibarra, M. Rotenone-Induced Model of Parkinson’s Disease: Beyond Mitochondrial Complex I Inhibition. Mol. Neurobiol. 2023, 60, 1929–1948. [Google Scholar] [CrossRef]
- Avallone, R.; Lucchi, C.; Puja, G.; Codeluppi, A.; Filaferro, M.; Vitale, G.; Rustichelli, C.; Biagini, G. BV-2 Microglial Cells Respond to Rotenone Toxic Insult by Modifying Pregnenolone, 5α-Dihydroprogesterone and Pregnanolone Levels. Cells 2020, 9, 2091. [Google Scholar] [CrossRef]
- Dello Russo, C.; Cappoli, N.; Coletta, I.; Mezzogori, D.; Paciello, F.; Pozzoli, G.; Navarra, P.; Battaglia, A. The Human Microglial HMC3 Cell Line: Where Do We Stand? A Systematic Literature Review. J. Neuroinflammation 2018, 15, 259. [Google Scholar] [CrossRef] [PubMed]
- Meletti, S.; Lucchi, C.; Monti, G.; Giovannini, G.; Bedin, R.; Trenti, T.; Rustichelli, C.; Biagini, G. Decreased Allopregnanolone Levels in Cerebrospinal Fluid Obtained during Status Epilepticus. Epilepsia 2017, 58, e16–e20. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.M.; Gol, M.; Lucchi, C.; Biagini, G. Antiepileptogenic Effects of Trilostane in the Kainic Acid Model of Temporal Lobe Epilepsy. Epilepsia 2023. [Google Scholar] [CrossRef]
- Šimončičová, E.; Gonçalves de Andrade, E.; Vecchiarelli, H.A.; Awogbindin, I.O.; Delage, C.I.; Tremblay, M.-È. Present and Future of Microglial Pharmacology. Trends Pharmacol. Sci. 2022, 43, 669–685. [Google Scholar] [CrossRef]
- Cho, I.; Kim, W.-J.; Kim, H.-W.; Heo, K.; Lee, B.I.; Cho, Y.-J. Increased Superoxide Dismutase 2 by Allopregnanolone Ameliorates ROS-Mediated Neuronal Death in Mice with Pilocarpine-Induced Status Epilepticus. Neurochem. Res. 2018, 43, 1464–1475. [Google Scholar] [CrossRef]
- Dziendzikowska, K.; Wilczak, J.; Grodzicki, W.; Gromadzka-Ostrowska, J.; Węsierska, M.; Kruszewski, M. Coating-Dependent Neurotoxicity of Silver Nanoparticles-An In Vivo Study on Hippocampal Oxidative Stress and Neurosteroids. Int. J. Mol. Sci. 2022, 23, 1365. [Google Scholar] [CrossRef] [PubMed]
- De Nicola, A.F.; Meyer, M.; Garay, L.; Kruse, M.S.; Schumacher, M.; Guennoun, R.; Gonzalez Deniselle, M.C. Progesterone and Allopregnanolone Neuroprotective Effects in the Wobbler Mouse Model of Amyotrophic Lateral Sclerosis. Cell. Mol. Neurobiol. 2022, 42, 23–40. [Google Scholar] [CrossRef]
- Mtchedlishvili, Z.; Bertram, E.H.; Kapur, J. Diminished allopregnanolone enhancement of GABAA receptor currents in a rat model of chronic temporal lobe epilepsy. J. Physiol. 2001, 537, 453–465. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucchi, C.; Codeluppi, A.; Filaferro, M.; Vitale, G.; Rustichelli, C.; Avallone, R.; Mandrioli, J.; Biagini, G. Human Microglia Synthesize Neurosteroids to Cope with Rotenone-Induced Oxidative Stress. Antioxidants 2023, 12, 963. https://doi.org/10.3390/antiox12040963
Lucchi C, Codeluppi A, Filaferro M, Vitale G, Rustichelli C, Avallone R, Mandrioli J, Biagini G. Human Microglia Synthesize Neurosteroids to Cope with Rotenone-Induced Oxidative Stress. Antioxidants. 2023; 12(4):963. https://doi.org/10.3390/antiox12040963
Chicago/Turabian StyleLucchi, Chiara, Alessandro Codeluppi, Monica Filaferro, Giovanni Vitale, Cecilia Rustichelli, Rossella Avallone, Jessica Mandrioli, and Giuseppe Biagini. 2023. "Human Microglia Synthesize Neurosteroids to Cope with Rotenone-Induced Oxidative Stress" Antioxidants 12, no. 4: 963. https://doi.org/10.3390/antiox12040963