
Spore Oil-Functionalized Selenium Nanoparticles Protect Pancreatic Beta Cells from Palmitic Acid-Induced Apoptosis via Inhibition of Oxidative Stress-Mediated Apoptotic Pathways
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
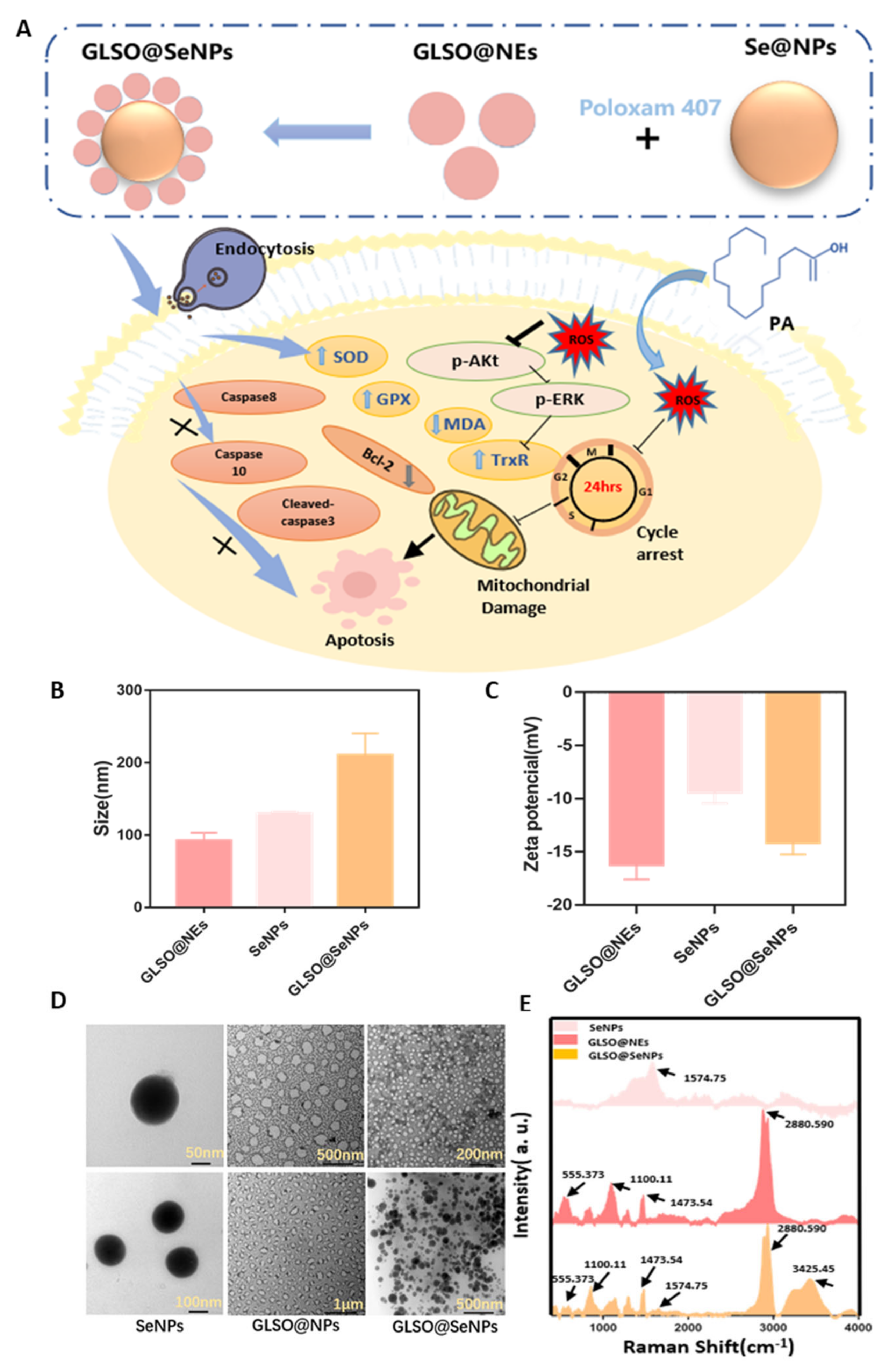
2.2. Synthesis and Characterization of GLSO Nanoemulsions (GLSO@NEs), SeNPs, and GLSO@SeNPs
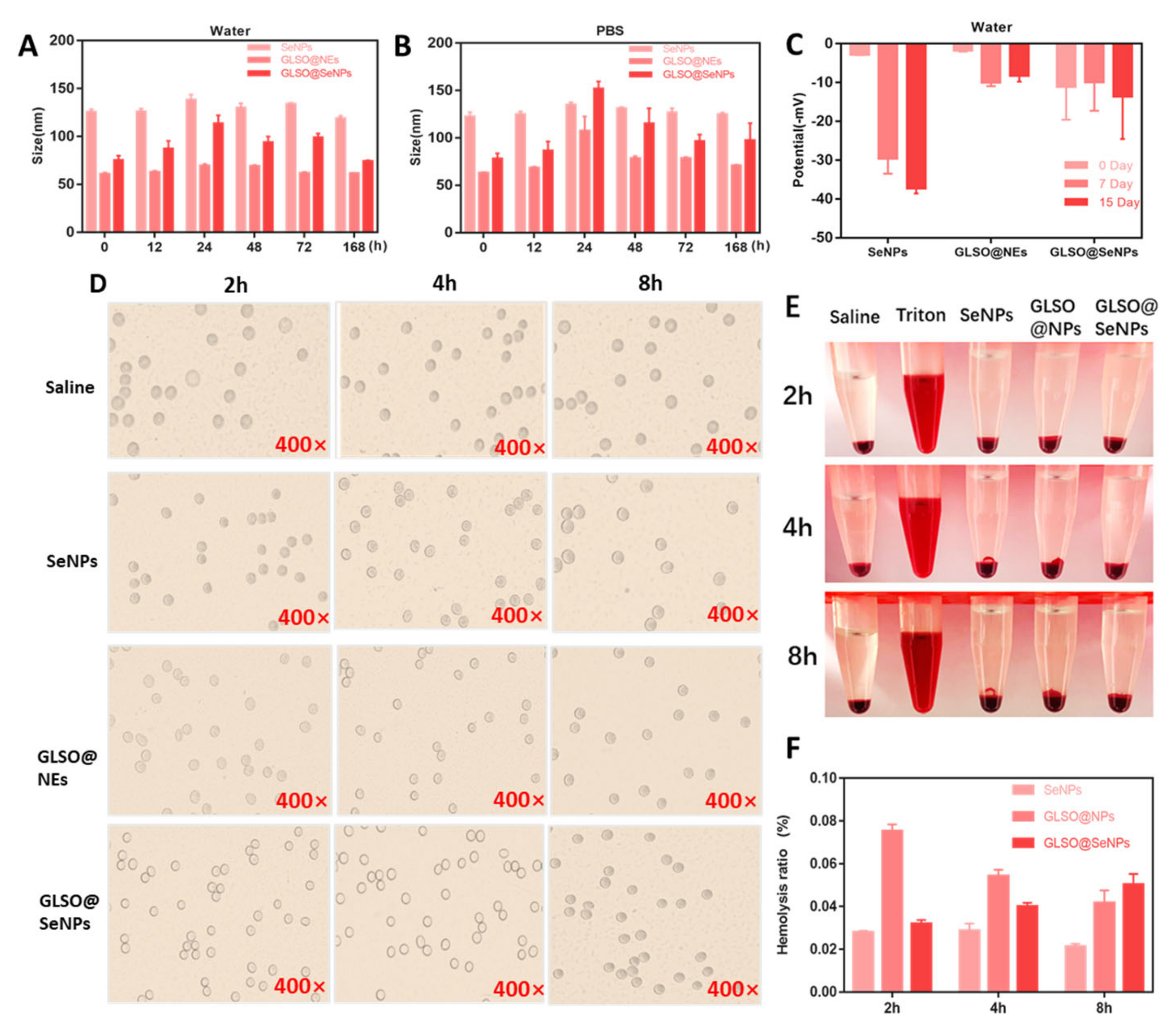
2.3. Stability Assessment of GLSO@NEs, SeNPs, and GLSO@SeNPs
2.4. Hemocompatibility of GLSO@NEs, SeNPs, and GLSO@SeNPs
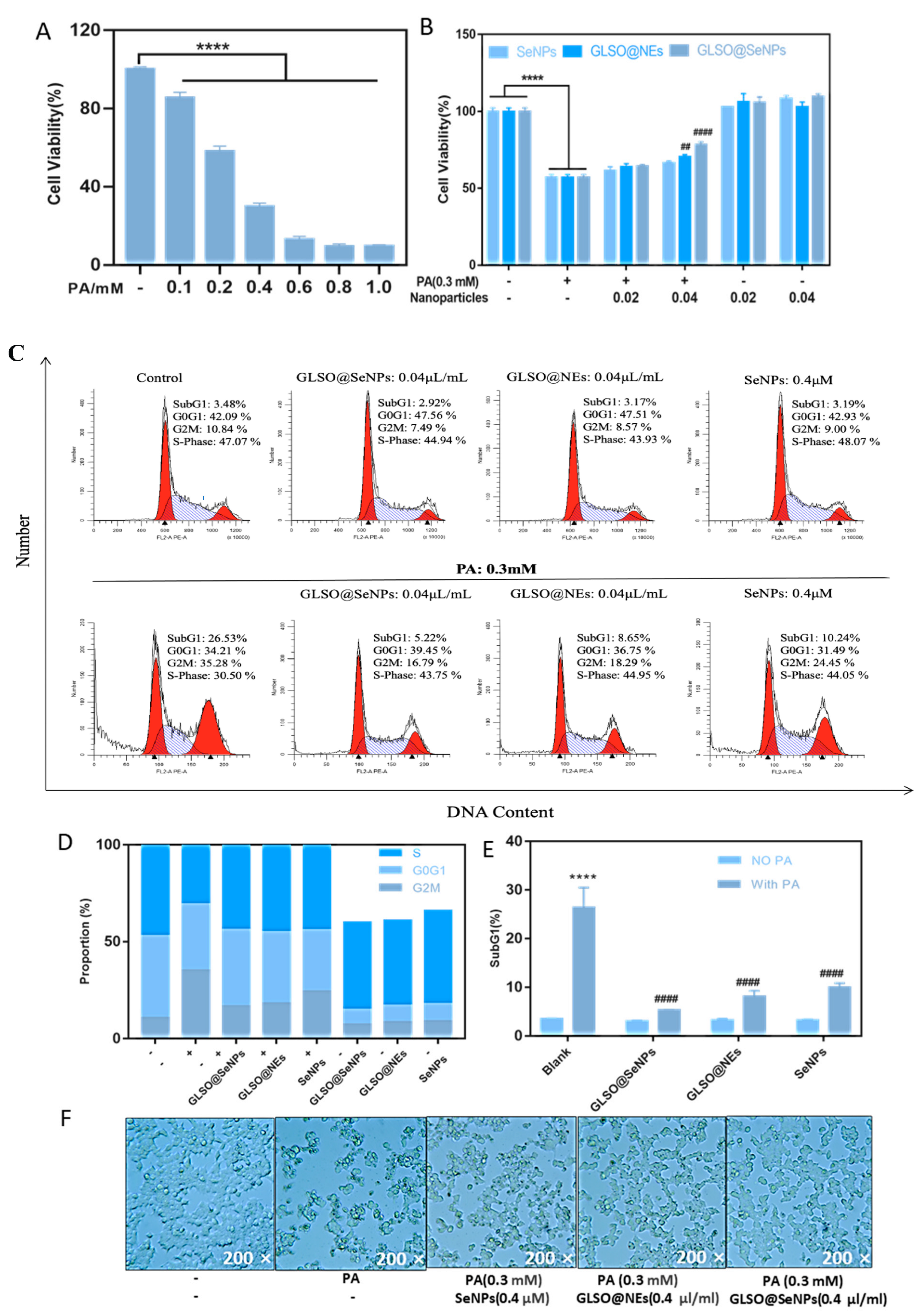
2.5. Cell Culture and Cell Viability Assay
2.6. Analysis of Cell Cycle Distribution
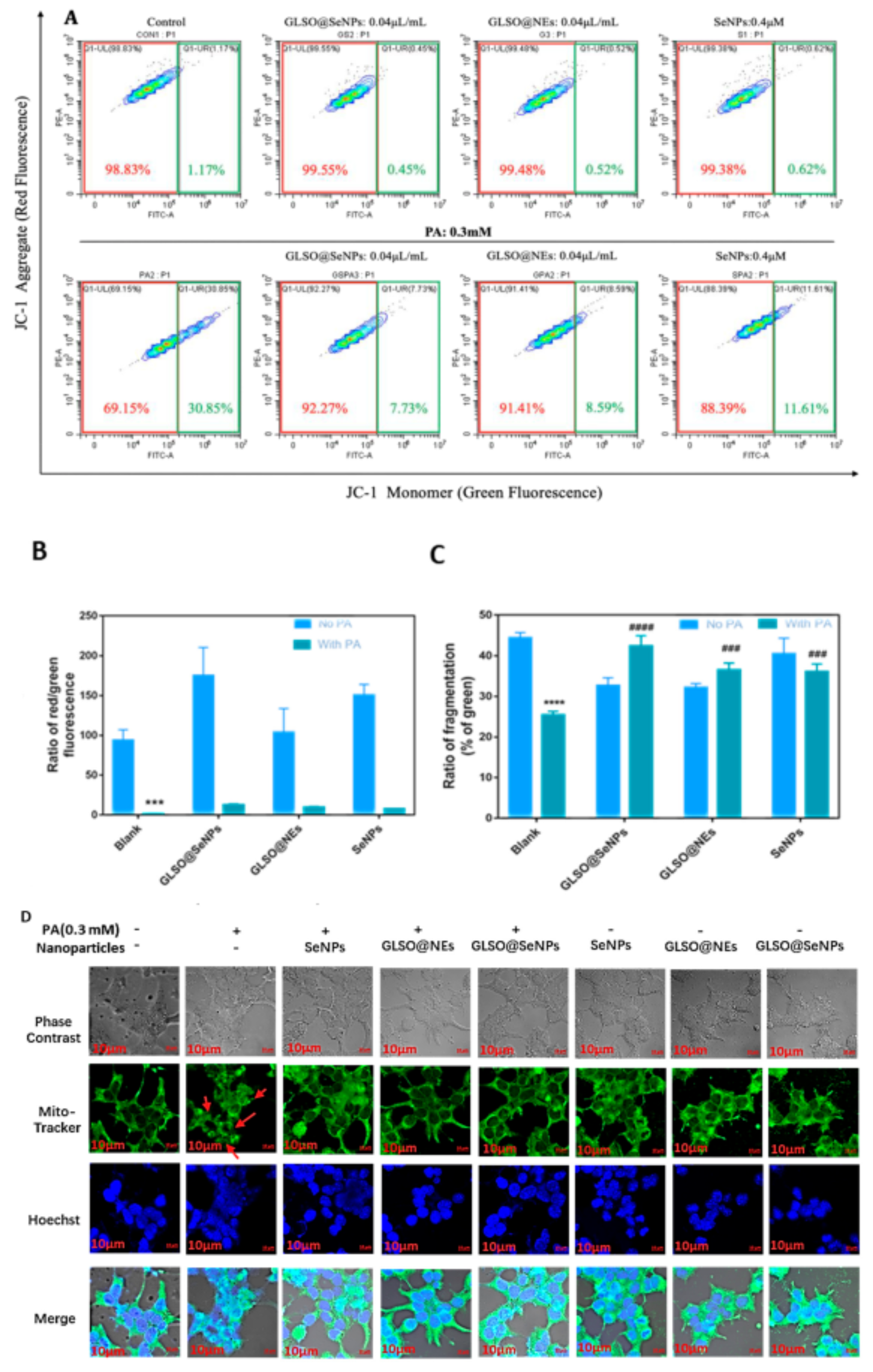
2.7. Mitochondrial Membrane Potential (ΔΨm) and Fragmentation Analysis
2.8. Assessment of ROS Generation
2.9. Measurement of SOD, MDA, and GSH
2.10. CAT, TrxR, and GPx Activity Assays
2.11. GLSO@SeNP Metabolism in INS-1E Cells
2.12. Western Blot Analysis
2.13. Statistical Analysis
3. Results
3.1. Preparation and Characterization of GLSO@NEs, SeNPs, and GLSO@SeNPs
3.2. Stability of GLSO@NEs, SeNPs, and GLSO@SeNPs in Water, PBS, Phenolic Red, and Human Blood
3.3. GLSO@SeNPs Protect against PA-Induced Cytotoxicity
3.4. GLSO@SeNPs Reverse PA–Induced Mitochondrial Dysfunction
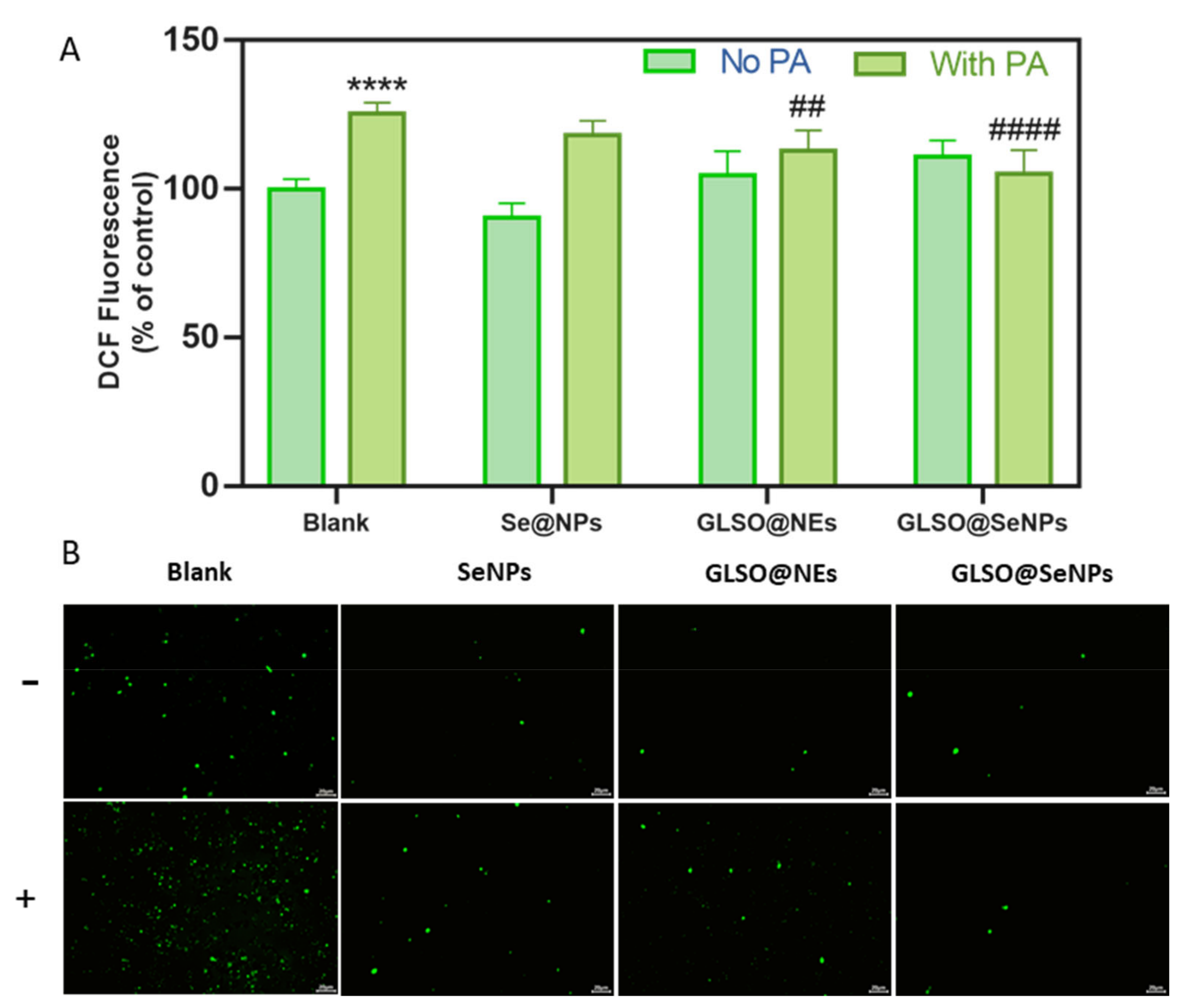
3.5. GLSO@SeNPs Reduce PA-Induced High Intracellular ROS Levels
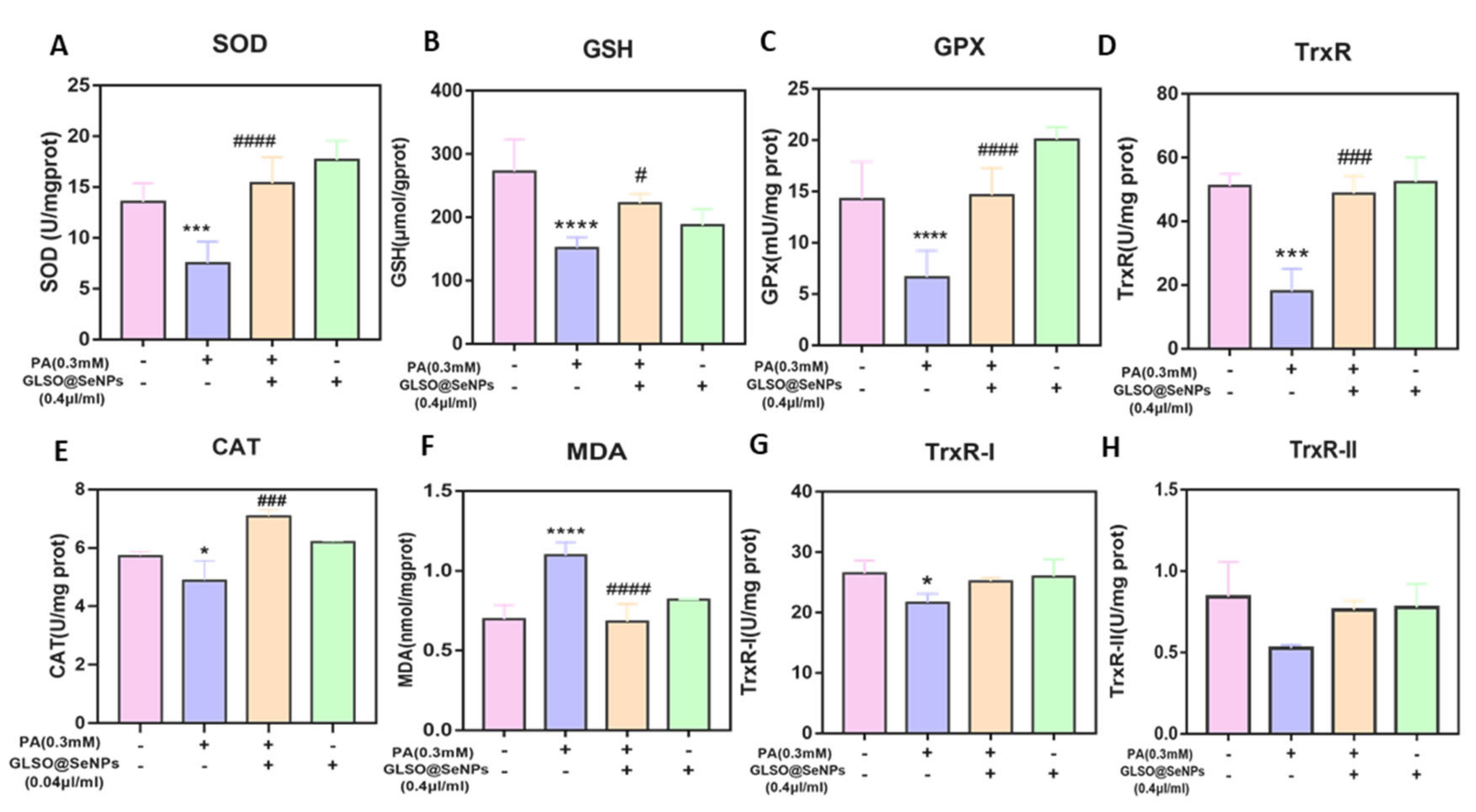
3.6. GLSO@SeNPs Regulate Antioxidant Enzyme Activity
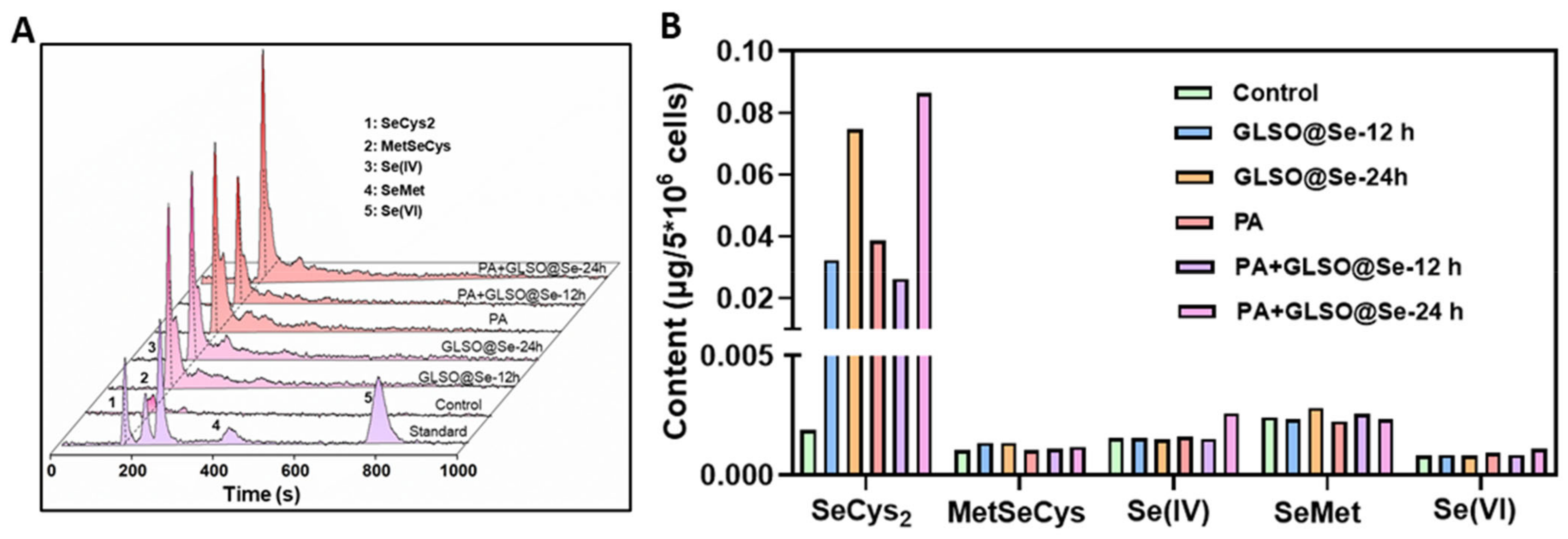
3.7. Metabolism of GLSO@SeNPs in INS-1E Cells
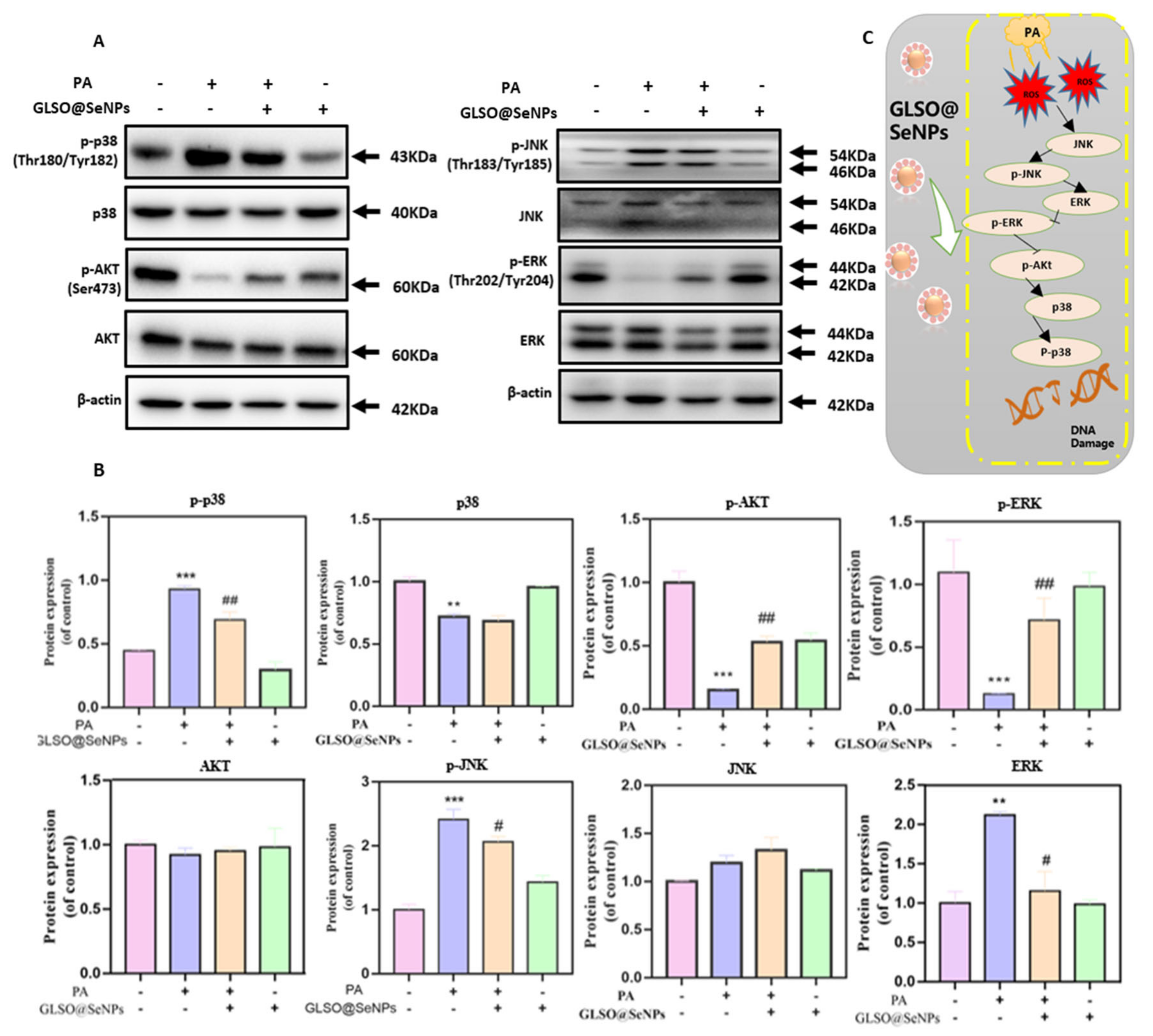
3.8. GLSO@SeNPs Regulate the ROS-Mediated Mitogen-Activated Protein Kinase (MAPK) Signaling Pathway
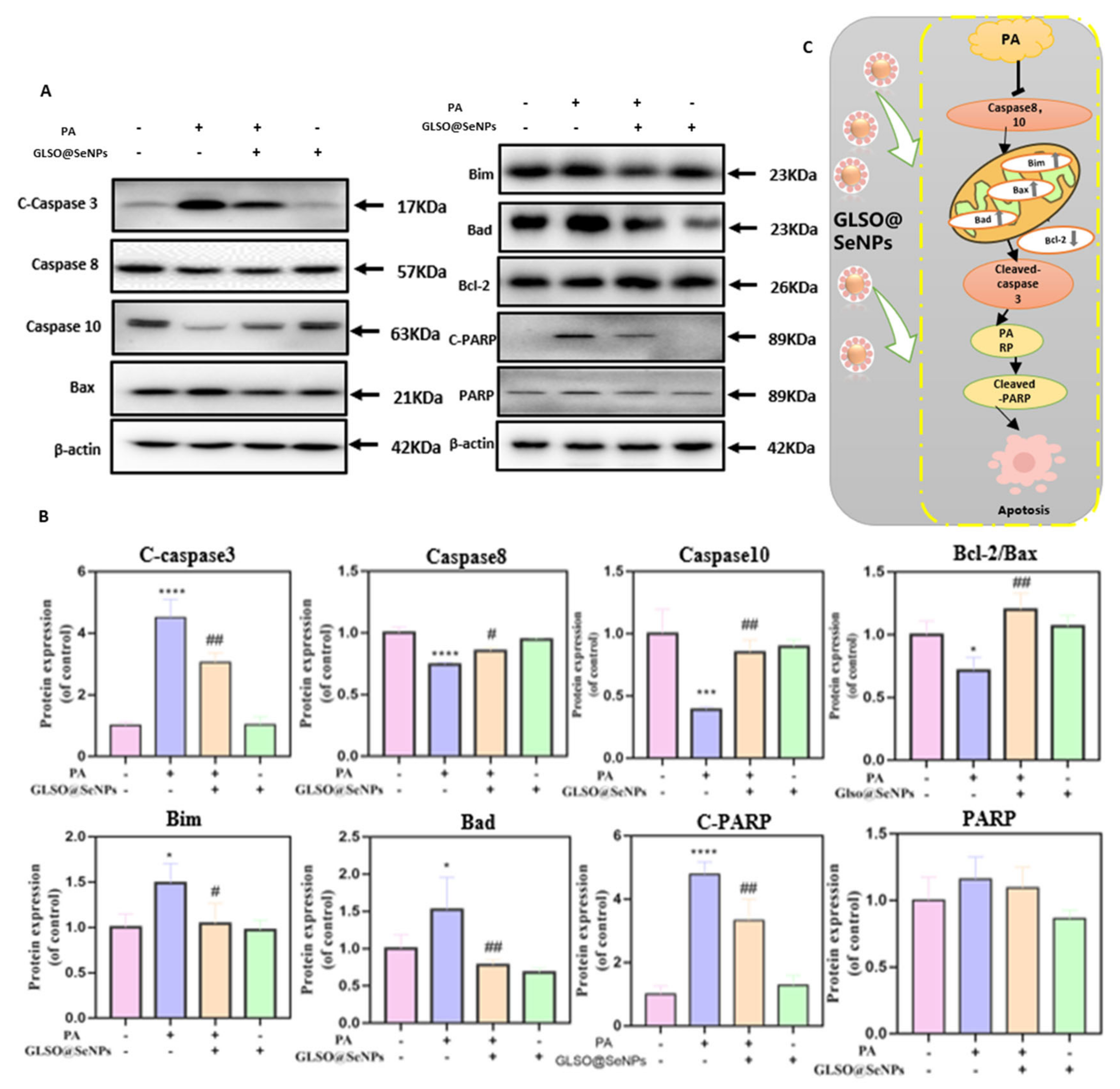
3.9. GLSO@SeNPs Suppress PA-Induced Caspase Activation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation Location: Brussels, Belgium, 2021. [Google Scholar]
- Jakubiak, G.K.; Osadnik, K.; Lejawa, M.; Osadnik, T.; Goławski, M.; Lewandowski, P.; Pawlas, N. “Obesity and Insulin Resistance” Is the Component of the Metabolic Syndrome Most Strongly Associated with Oxidative Stress. Antioxidants 2021, 11, 79. [Google Scholar] [CrossRef] [PubMed]
- Coral, D.E.; Fernandez-Tajes, J.; Tsereteli, N.; Pomares-Millan, H.; Fitipaldi, H.; Mutie, P.M.; Atabaki-Pasdar, N.; Kalamajski, S.; Poveda, A.; Miller-Fleming, T.W.; et al. A phenome-wide comparative analysis of genetic discordance between obesity and type 2 diabetes. Nat. Metab. 2023, 5, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Petry, S.F.; Römer, A.; Rawat, D.; Brunner, L.; Lerch, N.; Zhou, M.; Grewal, R.; Sharifpanah, F.; Sauer, H.; Eckert, G.P.; et al. Loss and Recovery of Glutaredoxin 5 Is Inducible by Diet in a Murine Model of Diabesity and Mediated by Free Fatty Acids In Vitro. Antioxidants 2022, 11, 788. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.S.; Guo, B.B.; Wang, G.H.; Zeng, L.M.; Hu, Y.H.; Wang, T.; Wang, H.Y. DGAT1 inhibitors protect pancreatic beta-cells from palmitic acid-induced apoptosis. Acta Pharmacol. Sin. 2021, 42, 264–271. [Google Scholar] [CrossRef]
- Yuan, J.; Li, F.; Cui, B.; Gao, J.; Yu, Z.; Lu, Z. Inhibition of GCN2 Alleviates Cardiomyopathy in Type 2 Diabetic Mice via Attenuating Lipotoxicity and Oxidative Stress. Antioxidants 2022, 11, 1379. [Google Scholar] [CrossRef]
- Zhu, Y.; Ren, C.; Zhang, M.; Zhong, Y. Perilipin 5 Reduces Oxidative Damage Associated With Lipotoxicity by Activating the PI3K/ERK-Mediated Nrf2-ARE Signaling Pathway in INS-1 Pancreatic beta-Cells. Front. Endocrinol. 2020, 11, 166. [Google Scholar] [CrossRef]
- Alnahdi, A.; John, A.; Raza, H. N-acetyl cysteine attenuates oxidative stress and glutathione-dependent redox imbalance caused by high glucose/high palmitic acid treatment in pancreatic Rin-5F cells. PLoS ONE 2019, 14, e0226696. [Google Scholar] [CrossRef]
- Qu, H.L.; Sun, L.J.; Li, X.; Liu, F.; Sun, H.H.; He, X.T.; Gan, D.; Yin, Y.; Tian, B.M.; Chen, F.M.; et al. Long non-coding RNA AC018926.2 regulates palmitic acid exposure-compromised osteogenic potential of periodontal ligament stem cells via the ITGA2/FAK/AKT pathway. Cell Prolif. 2023, e13411. [Google Scholar] [CrossRef]
- Cen, K.; Chen, M.; He, M.; Li, Z.; Song, Y.; Liu, P.; Jiang, Q.; Xu, S.; Jia, Y.; Shen, P. Sporoderm-Broken Spores of Ganoderma lucidum Sensitizes Ovarian Cancer to Cisplatin by ROS/ERK Signaling and Attenuates Chemotherapy-Related Toxicity. Front. Pharmacol. 2022, 13, 826716. [Google Scholar] [CrossRef]
- Sheng, Z.; Wen, L.; Yang, B. Structure identification of a polysaccharide in mushroom Lingzhi spore and its immunomodulatory activity. Carbohydr. Polym. 2021, 278, 118939. [Google Scholar] [CrossRef]
- Jiao, C.; Chen, W.; Tan, X.; Liang, H.; Li, J.; Yun, H.; He, C.; Chen, J.; Ma, X.; Xie, Y.; et al. Ganoderma lucidum spore oil induces apoptosis of breast cancer cells in vitro and in vivo by activating caspase-3 and caspase-9. J. Ethnopharmacol. 2020, 247, 112256. [Google Scholar] [CrossRef]
- Zhou, D.; Zhou, F.; Ma, J.; Ge, F. Microcapsulation of Ganoderma Lucidum spores oil: Evaluation of its fatty acids composition and enhancement of oxidative stability. Ind. Crops Prod. 2019, 131, 1–7. [Google Scholar] [CrossRef]
- Peng, H.; Zhong, L.; Cheng, L.; Chen, L.; Tong, R.; Shi, J.; Bai, L. Ganoderma lucidum: Current advancements of characteristic components and experimental progress in anti-liver fibrosis. Front. Pharmacol. 2022, 13, 1094405. [Google Scholar] [CrossRef]
- Viroel, F.J.M.; Laurino, L.F.; Caetano, E.L.A.; Jozala, A.F.; Spim, S.R.V.; Pickler, T.B.; Sercundes, M.K.; Gomes, M.C.; Hataka, A.; Grotto, D.; et al. Ganoderma lucidum Modulates Glucose, Lipid Peroxidation and Hepatic Metabolism in Streptozotocin-Induced Diabetic Pregnant Rats. Antioxidants 2022, 11, 1035. [Google Scholar] [CrossRef]
- Shokrani, H.; Shokrani, A.; Sajadi, S.M.; Yazdi, M.K.; Seidi, F.; Jouyandeh, M.; Zarrintaj, P.; Kar, S.; Kim, S.-J.; Kuang, T.; et al. Polysaccharide-based nanocomposites for biomedical applications: A critical review. Nanoscale Horiz. 2022, 7, 1136–1160. [Google Scholar] [CrossRef]
- Dai, C.; He, L.; Ma, B.; Chen, T. Facile Nanolization Strategy for Therapeutic Ganoderma Lucidum Spore Oil to Achieve Enhanced Protection against Radiation-Induced Heart Disease. Small 2019, 15, e1902642. [Google Scholar] [CrossRef]
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, B.; Peng, Q.; Baron, M.; Melcova, M.; Opatrilova, R.; Zidkova, J.; et al. Nano-selenium and its nanomedicine applications: A critical review. Int. J. Nanomed. 2018, 13, 2107–2128. [Google Scholar] [CrossRef]
- Deng, W.; Wang, H.; Wu, B.; Zhang, X. Selenium-layered nanoparticles serving for oral delivery of phytomedicines with hypoglycemic activity to synergistically potentiate the antidiabetic effect. Acta Pharm. Sin. B 2019, 9, 74–86. [Google Scholar] [CrossRef]
- Liu, C.; Fu, Y.; Li, C.E.; Chen, T.; Li, X. Phycocyanin-Functionalized Selenium Nanoparticles Reverse Palmitic Acid-Induced Pancreatic beta Cell Apoptosis by Enhancing Cellular Uptake and Blocking Reactive Oxygen Species (ROS)-Mediated Mitochondria Dysfunction. J. Agric. Food Chem. 2017, 65, 4405–4413. [Google Scholar] [CrossRef]
- Dai, C.; Tang, Z.; Li, X.; Chen, T. High-pressure homogenization and tailoring of size-tunable Ganoderma lucidum spore oil nanosystem for enhanced anticancer therapy. Chem. Eng. J. 2021, 406, 127125. [Google Scholar] [CrossRef]
- Yang, F.; Huang, J.; Liu, H.; Lin, W.; Li, X.; Zhu, X.; Chen, T. Lentinan-functionalized selenium nanosystems with high permeability infiltrate solid tumors by enhancing transcellular transport. Nanoscale 2020, 12, 14494–14503. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Han, Q.; Zhao, X.; Li, H.; Yuan, W.-E.; Fan, C. Asymmetrical 3D Nanoceria Channel for Severe Neurological Defect Regeneration. iScience 2019, 12, 216–231. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Velasco, S.; Delgado, J.; Peña, F.J.; Estévez, M. Ellagic Acid Triggers the Necrosis of Differentiated Human Enterocytes Exposed to 3-Nitro-Tyrosine: An MS-Based Proteomic Study. Antioxidants 2022, 11, 2485. [Google Scholar] [CrossRef] [PubMed]
- Cretin, E.; Lopes, P.; Vimont, E.; Tatsuta, T.; Langer, T.; Gazi, A.; Sachse, M.; Yu-Wai-Man, P.; Reynier, P.; Wai, T. High-throughput screening identifies suppressors of mitochondrial fragmentation in OPA1 fibroblasts. EMBO Mol. Med. 2021, 13, e13579. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Chen, X.; Yang, Y.; Wang, F.; Li, W.; Awuti, M.; Sun, Y.; Lian, C.; Li, Z.; et al. miR-365 promotes diabetic retinopathy through inhibiting Timp3 and increasing oxidative stress. Exp. Eye Res. 2018, 168, 89–99. [Google Scholar] [CrossRef]
- Ma, T.; Zhang, Y.; Zhang, C.; Luo, J.-G.; Kong, L.-Y. Downregulation of TIGAR sensitizes the antitumor effect of physapubenolide through increasing intracellular ROS levels to trigger apoptosis and autophagosome formation in human breast carcinoma cells. Biochem. Pharmacol. 2017, 143, 90–106. [Google Scholar] [CrossRef]
- Zhang, L.; Fan, J.; He, J.; Chen, W.; Jin, W.; Zhu, Y.; Sun, H.; Li, Y.; Shi, Y.; Jing, Y.; et al. Regulation of ROS-NF-kappaB axis by tuna backbone derived peptide ameliorates inflammation in necrotizing enterocolitis. J. Cell Physiol. 2019, 234, 14330–14338. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Y.; Wu, J.; Yang, J.; Li, M.; Chen, Q. The Potential Value of Targeting Ferroptosis in Early Brain Injury After Acute CNS Disease. Front. Mol. Neurosci. 2020, 13, 110. [Google Scholar] [CrossRef]
- Ali, A.O.; Abdalla, M.S.; Shahein, Y.E.; Shokeer, A.; Sharada, H.M.; Ali, K.A. Grafted carrageenan: Alginate gel beads for catalase enzyme covalent immobilization. 3 Biotech 2021, 11, 341. [Google Scholar] [CrossRef]
- Du, Y.; Zhang, Z.; Yang, Y.; Liu, T.; Chen, T.; Li, X. Highly active selenium nanotherapeutics combined with metformin to achieve synergistic sensitizing effect on NK cells for osteosarcoma therapy. Nanophotonics 2022, 11, 5101–5111. [Google Scholar] [CrossRef]
- Alnahdi, A.; John, A.; Raza, H. Augmentation of Glucotoxicity, Oxidative Stress, Apoptosis and Mitochondrial Dysfunction in HepG2 Cells by Palmitic Acid. Nutrients 2019, 11, 1979. [Google Scholar] [CrossRef]
- Schütz, C.A.; Juillerat-Jeanneret, L.; Mueller, H.; Lynch, I.; Riediker, M. Therapeutic nanoparticles in clinics and under clinical evaluation. Nanomedicine 2013, 8, 449–467. [Google Scholar] [CrossRef]
- Liang, X.; Liu, T.; Li, L.; Li, J.; Li, S.; Zeng, K.; Song, C.; Chen, T.; Zou, Y. Translational selenium nanotherapeutics counter-acts multiple risk factors to improve surgery-induced cognitive impairment. Chem. Eng. J. 2022, 441, 135984. [Google Scholar] [CrossRef]
- Oudih, S.B.; Tahtat, D.; Khodja, A.N.; Mahlous, M.; Hammache, Y.; Guittoum, A.; Gana, S.K. Chitosan nanoparticles with controlled size and zeta potential. Polym. Eng. Sci. 2023, 63, 1011–1021. [Google Scholar] [CrossRef]
- Karthikeyan, C.; Amawi, H.; Viana, A.G.; Sanglard, L.; Hussein, N.; Saddler, M.; Ashby, C.R., Jr.; Moorthy, N.H.N.; Trivedi, P.; Tiwari, A.K. lH-Pyrazolo[3,4-b]quinolin-3-amine derivatives inhibit growth of colon cancer cells via apoptosis and sub G1 cell cycle arrest. Bioorg. Med. Chem. Lett. 2018, 28, 2244–2249. [Google Scholar] [CrossRef]
- Youssef, N.S.; Elzaitony, A.S.; Baky, N.A.A. Diacerein attenuate LPS-induced acute lung injury via inhibiting ER stress and apoptosis: Impact on the crosstalk between SphK1/S1P, TLR4/NFkappaB/STAT3, and NLRP3/IL-1beta signaling pathways. Life Sci. 2022, 308, 120915. [Google Scholar] [CrossRef]
- Yu, X.; Peng, W.; Wang, Y.; Xu, W.; Chen, W.; Huang, L.; Xu, H.; He, X.; Wang, S.; Sun, Q.; et al. Palmitic Acid Inhibits the Growth and Metastasis of Gastric Cancer by Blocking the STAT3 Signaling Pathway. Cancers 2023, 15, 388. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, X.A.; Liu, C.; Shen, Q.; Wu, Y. Vitamin B6 Inhibits High Glucose-Induced Islet beta Cell Apoptosis by Upregulating Autophagy. Metabolites 2022, 12, 1048. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, Y.; Xu, C.; Liu, J.; Chen, J.; Li, G.; Huang, B.; Pan, Y.; Zhang, Y.; Wei, Q.; et al. Circular RNA circGlis3 protects against islet beta-cell dysfunction and apoptosis in obesity. Nat. Commun. 2023, 14, 351. [Google Scholar] [CrossRef]
- Wang, S.; Wu, H.; Zhang, X.; Luo, S.; Zhou, S.; Fan, H.; Lv, C. Preparation of nano-selenium from chestnut polysaccharide and characterization of its antioxidant activity. Front. Nutr. 2022, 9, 1054601. [Google Scholar] [CrossRef]
- Wang, X.; Liu, W.; Li, Y.; Ma, L.; Lin, Z.; Xu, J.; Guo, Y. Preparation and anti-tumor activity of selenium nanoparticles based on a polysaccharide from Paeonia lactiflora. Int. J. Biol. Macromol. 2023, 232, 123261. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.; Gonçalves, M.d.A.; de Macedo, L.F.; Torres, A.H.F.; Marena, G.D.; Chorilli, M.; Trovatti, E. Green nanotechnology for the development of nanoparticles based on alginate associated with essential and vegetable oils for application in fruits and seeds protection. Int. J. Biol. Macromol. 2023, 232, 123351. [Google Scholar] [CrossRef] [PubMed]
- Marmentini, C.; Guimarães, D.S.P.; de Lima, T.I.; Teófilo, F.B.S.; da Silva, N.S.; Soares, G.M.; Boschero, A.C.; Kurauti, M.A. Rosiglitazone protects INS-1E cells from human islet amyloid polypeptide toxicity. Eur. J. Pharmacol. 2022, 928, 175122. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, S.; Boger, M.; Lortz, S.; Mehmeti, I. Luminal H2O2 promotes ER Ca2+ dysregulation and toxicity of palmitate in insulin-secreting INS-1E cells. FASEB J. 2023, 37, e22685. [Google Scholar] [CrossRef] [PubMed]
- Ponomareva, S.; Joisten, H.; François, T.; Naud, C.; Morel, R.; Hou, Y.; Myers, T.O.; Joumard, I.; Dieny, B.; Carriere, M. Magnetic particles for triggering insulin release in INS-1E cells subjected to a rotating magnetic field. Nanoscale 2022, 14, 13274–13283. [Google Scholar] [CrossRef]
- Zhu, J.; Han, J.; Liu, L.; Liu, Y.; Xu, W.; Li, X.; Yang, L.; Gu, Y.; Tang, W.; Shi, Y.; et al. Pancreatic Islet beta-cell Expert Panel of the Chinese Diabetes, and A. Endocrinology Society of Jiangsu Medical, Clinical expert consensus on the assessment and protection of pancreatic islet beta-cell function in type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 2023, 197, 110568. [Google Scholar] [CrossRef]
- Huang, T.-S.; Wu, T.; Wu, Y.-D.; Li, X.-H.; Tan, J.; Shen, C.-H.; Xiong, S.-J.; Feng, Z.-Q.; Gao, S.-F.; Li, H.; et al. Long-term statins administration exacerbates diabetic nephropathy via ectopic fat deposition in diabetic mice. Nat. Commun. 2023, 14, 390. [Google Scholar] [CrossRef]
- Gong, W.; Zhang, S.; Chen, Y.; Shen, J.; Zheng, Y.; Liu, X.; Zhu, M.; Meng, G. Protective role of hydrogen sulfide against diabetic cardiomyopathy via alleviating necroptosis. Free. Radic. Biol. Med. 2022, 181, 29–42. [Google Scholar] [CrossRef]
- Yung, J.H.M.; Yeung, L.S.N.; Ivovic, A.; Tan, Y.F.; Jentz, E.M.; Batchuluun, B.; Gohil, H.; Wheeler, M.B.; Joseph, J.W.; Giacca, A.; et al. Prevention of Lipotoxicity in Pancreatic Islets with Gammahydroxybutyrate. Cells 2022, 11, 545. [Google Scholar] [CrossRef]
- Khoi, C.-S.; Xiao, C.-Q.; Hung, K.-Y.; Lin, T.-Y.; Chiang, C.-K. Oxidative Stress-Induced Growth Inhibitor (OSGIN1), a Target of X-Box-Binding Protein 1, Protects Palmitic Acid-Induced Vascular Lipotoxicity through Maintaining Autophagy. Biomedicines 2022, 10, 992. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, S.; Li, K.; Li, X.; Yin, H.; Li, S.; Gao, X.-J. TBBPA induced ROS overproduction promotes apoptosis and inflammation by inhibiting autophagy in mice lung. Ecotoxicol. Environ. Saf. 2023, 252, 114607. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, B.; Ye, B.; Sun, Z.; Qian, Z.; Yu, L.; Bi, Y.; Ma, L.; Ding, Y.; Du, Y.; et al. Mitochondrial-Targeted Delivery of Polyphenol-Mediated Antioxidases Complexes against Pyroptosis and Inflammatory Diseases. Adv. Mater. 2023, 35, e2208571. [Google Scholar] [CrossRef]
- Peterle, L.; Sanfilippo, S.; Borgia, F.; Cicero, N.; Gangemi, S. Alopecia Areata: A Review of the Role of Oxidative Stress, Possible Biomarkers, and Potential Novel Therapeutic Approaches. Antioxidants 2023, 12, 135. [Google Scholar] [CrossRef]
- Li, B.; Kwok, L.-Y.; Wang, D.; Li, L.; Guo, S.; Chen, Y. Integrating metabolomics, bionics, and culturomics to study probiotics-driven drug metabolism. Front. Pharmacol. 2023, 14, 1047863. [Google Scholar] [CrossRef]
- Farkhondeh, T.; Mehrpour, O.; Buhrmann, C.; Pourbagher-Shahri, A.; Shakibaei, M.; Samarghandian, S. Organophosphorus Compounds and MAPK Signaling Pathways. Int. J. Mol. Sci. 2020, 21, 4258. [Google Scholar] [CrossRef]
- Yang, W.; Zhu, L.; Lai, S.; Ding, Q.; Xu, T.; Guo, R.; Dou, X.; Chai, H.; Yu, Z.; Li, S. Cimifugin Ameliorates Lipotoxicity-Induced Hepatocyte Damage and Steatosis through TLR4/p38 MAPK- and SIRT1-Involved Pathways. Oxid. Med. Cell Longev. 2022, 2022, 4557532. [Google Scholar] [CrossRef]
- Vicinanza, M.; Rubinsztein, D.C. Deadly Encounter: Endosomes Meet Mitochondria to Initiate Apoptosis. Dev. Cell 2020, 53, 619–620. [Google Scholar] [CrossRef]
- Zhu, Q.; Yang, Q.; Lu, X.; Wang, H.; Tong, L.; Li, Z.; Liu, G.; Bao, Y.; Xu, X.; Gu, L.; et al. SETD2-mediated H3K14 trimethylation promotes ATR activation and stalled replication fork restart in response to DNA replication stress. Proc. Natl. Acad. Sci. USA 2021, 118, e2011278118. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, S.; Zhu, H.; Xiong, P.; Shi, L.; Bai, W.; Li, X. Spore Oil-Functionalized Selenium Nanoparticles Protect Pancreatic Beta Cells from Palmitic Acid-Induced Apoptosis via Inhibition of Oxidative Stress-Mediated Apoptotic Pathways. Antioxidants 2023, 12, 840. https://doi.org/10.3390/antiox12040840
Zhou S, Zhu H, Xiong P, Shi L, Bai W, Li X. Spore Oil-Functionalized Selenium Nanoparticles Protect Pancreatic Beta Cells from Palmitic Acid-Induced Apoptosis via Inhibition of Oxidative Stress-Mediated Apoptotic Pathways. Antioxidants. 2023; 12(4):840. https://doi.org/10.3390/antiox12040840
Chicago/Turabian StyleZhou, Sajin, Hongyan Zhu, Piaopiao Xiong, Lei Shi, Weibin Bai, and Xiaoling Li. 2023. "Spore Oil-Functionalized Selenium Nanoparticles Protect Pancreatic Beta Cells from Palmitic Acid-Induced Apoptosis via Inhibition of Oxidative Stress-Mediated Apoptotic Pathways" Antioxidants 12, no. 4: 840. https://doi.org/10.3390/antiox12040840