

Creatine Supplementation Potentiates Exercise Protective Effects against Doxorubicin-Induced Hepatotoxicity in Mice

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Study Design
2.2. ST Protocol
2.3. Euthanasia and Tissue Preparation
2.4. Histological Analysis
2.5. Interleukins, Oxidative Stress, and Oxidative Damage Analysis
2.6. Statistical Analyses
3. Results
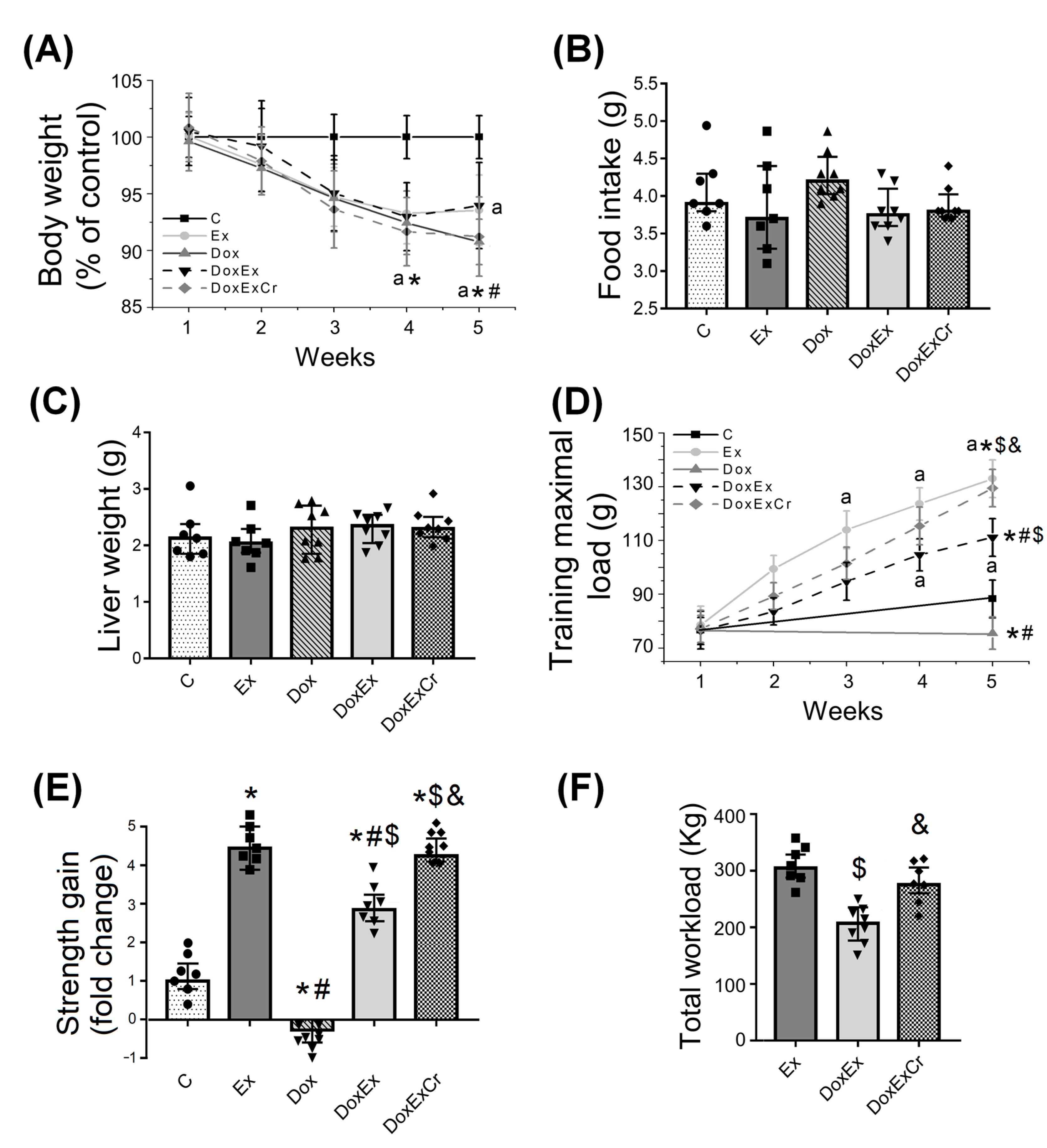
3.1. Effects of Creatine and ST on Doxorubicin-Induced Weight Loss and Muscle Strength
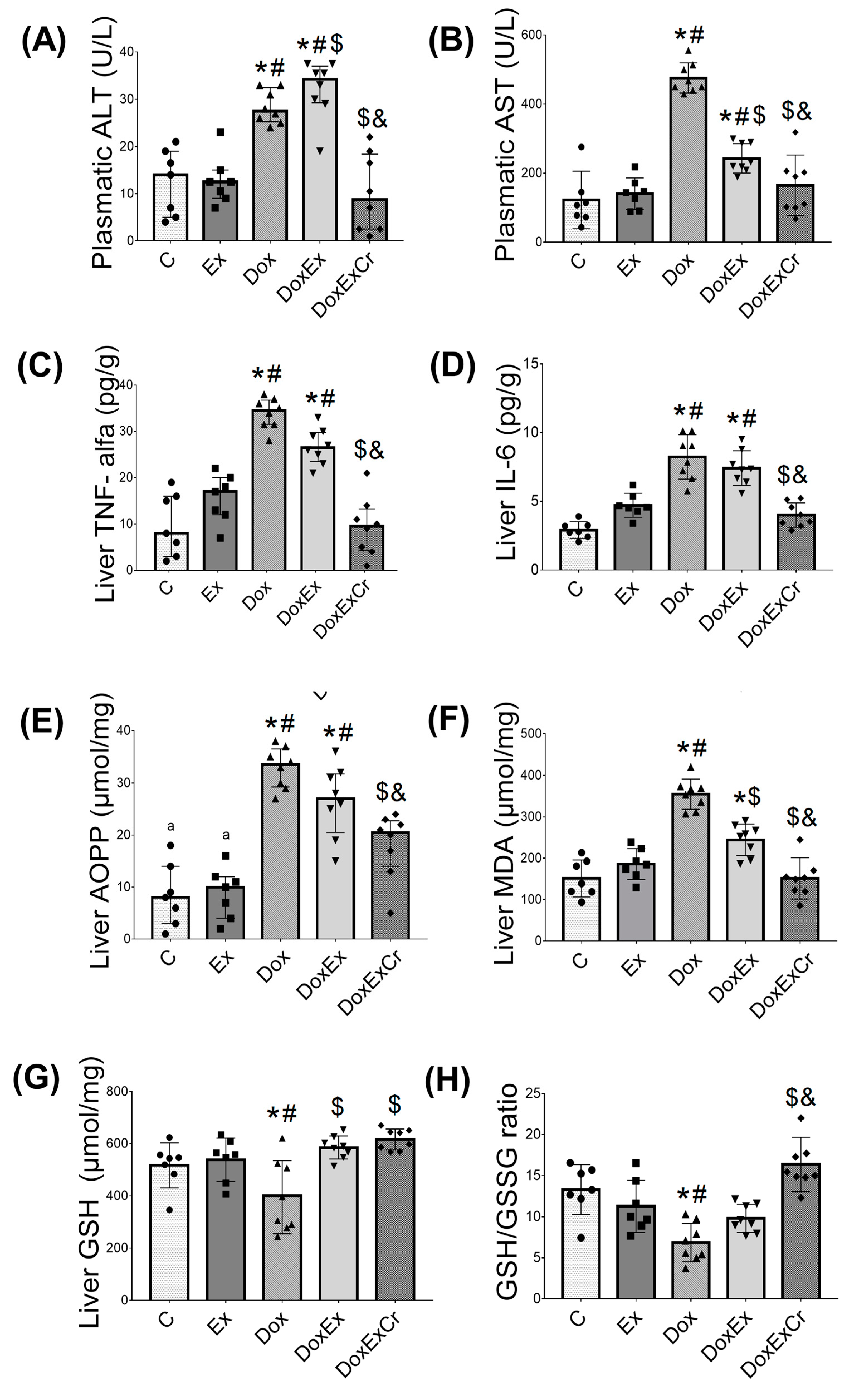
3.2. Effects of Creatine and ST on Doxorubicin-Induced Hepatotoxicity
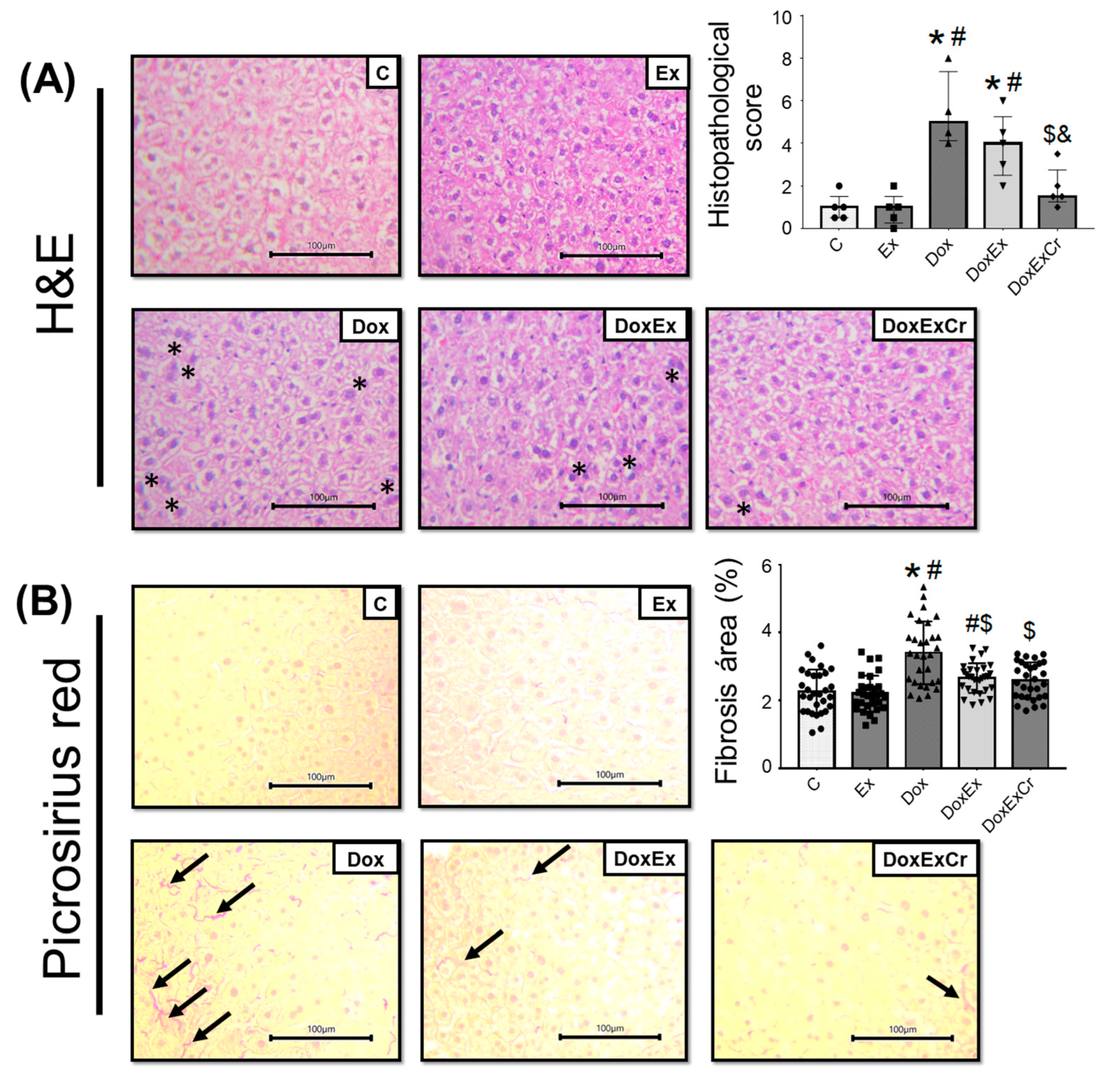
3.3. Histopathological Analyzes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ministério da Saúde Brasil; Secretaria de Atenção à Saúde. Clinical Practice Guidelines in Oncology; Ministério Da Saúde: Brasília, Brazil, 2014.
- Powers, S.K.; Duarte, J.A.; Le Nguyen, B.; Hyatt, H. Endurance exercise protects skeletal muscle against both doxorubicin-induced and inactivity-induced muscle wasting. Pflug. Arch. Eur. J. Physiol. 2019, 441, 53. [Google Scholar] [CrossRef] [PubMed]
- McGowan, J.V.; Chung, R.; Maulik, A.; Piotrowska, I.; Walker, J.M.; Yellon, D.M. Anthracycline Chemotherapy and Cardiotoxicity. Cardiovasc. Drugs 2017, 31, 63–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, V.S.; Lenihan, D.J.; Ky, B. Cancer therapy-induced cardiotoxicity: Basic mechanisms and potential cardioprotective therapies. J. Am. Heart Assoc. 2014, 3, e000665. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, Q.; Qi, H.; Wang, C.; Zhang, J.; Dong, L. Doxorubicin-induced systemic inflammation is driven by upregulation of toll-like receptor TLR4 and endotoxin leakage. Cancer Res. 2016, 76, 6631–6642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, S.; Ali, N.; Nafees, S.; Ahmad, S.T.; Arjumand, W.; Hasan, S.K. Alleviation of doxorubicin-induced nephrotoxicity and hepatotoxicity by chrysin in Wistar rats. Toxicol. Mech. Methods 2013, 23, 337–345. [Google Scholar] [CrossRef]
- Santos, D.L.; Moreno, A.J.M.; Leino, R.L.; Froberg, M.K.; Wallace, K.B. Carvedilol protects against doxorubicin-induced mitochondrial cardiomyopathy. Toxicol. Appl. Pharmacol. 2002, 185, 218–227. [Google Scholar] [CrossRef] [Green Version]
- Singla, S.; Kumar, N.; Kaur, J. In vivo studies on the protective effect of propolis on doxorubicin-induced toxicity in liver of male rats. Toxicol. Int. 2014, 21, 191–195. [Google Scholar] [CrossRef] [Green Version]
- AlAsmari, A.F.; Alharbi, M.; Alqahtani, F.; Alasmari, F.; AlSwayyed, M.; Alzarea, S.I. Diosmin alleviates doxorubicin-induced liver injury via modulation of oxidative stress-mediated hepatic inflammation and apoptosis via NfkB and MAPK pathway: A preclinical study. Antioxidants 2021, 10, 1998. [Google Scholar] [CrossRef]
- Alishahi, A.; Roshan, V.D.; Hedayyati, M. Pretreatment effects of regular aerobic training on the IGF system and hepatotoxicity induced by Doxorubicin in Rats. Asian Pac. J. Cancer Prev. 2013, 14, 7427–7431. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Nie, J.; Ferrari, G.; Rey-Lopez, J.P.; Rezende, L.F.M. Association of Physical Activity Intensity with Mortality: A National Cohort Study of 403681 US Adults. JAMA Intern. Med. 2021, 181, 203–211. [Google Scholar] [CrossRef]
- Zhao, M.; Veeranki, S.P.; Magnussen, C.G.; Xi, B. Recommended physical activity and all cause and cause specific mortality in US adults: Prospective cohort study. BMJ 2020, 370, m2031. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.L.; Winters-Stone, K.M.; Wiskemann, J.; May, A.M.; Schwartz, A.L.; Courneya, K.S. Exercise Guidelines for Cancer Survivors: Consensus Statement from International Multidisciplinary Roundtable. Med. Sci. Sports Exerc. 2019, 51, 2375–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, S.C.; Spence, R.R.; Galvão, D.A.; Newton, R.U. Australian Association for Exercise and Sport Science position stand: Optimising cancer outcomes through exercise. J. Sci. Med. Sport 2009, 12, 428–434. [Google Scholar] [CrossRef] [Green Version]
- Padilha, C.S.; Marinello, P.C.; Galvão, D.A.; Newton, R.U.; Borges, F.H.; Frajacomo, F.T.; Deminice, R. Evaluation of resistance training to improve muscular strength and body composition in cancer patients undergoing neoadjuvant and adjuvant therapy: A meta-analysis. J. Cancer Surviv. 2017, 11, 339–349. [Google Scholar] [CrossRef]
- Deminice, R.; Rezende, L.F.M.; Rosa, D.D.; Cangussu, R.; Garcia, L.M.T.; Riera, R.; Brum, P.C. Physical activity recommendations for cancer prevention and control: A Brazilian consortium. Braz. J. Oncol. 2022, 18, e-20220311. [Google Scholar] [CrossRef]
- Zolfagharzadeh, F.; Roshan, V.D. Pretreatment hepatoprotective effect of regular aerobic training against hepatic toxicity induced by doxorubicin in rats. Asian Pac. J. Cancer Prev. 2013, 14, 2931–2936. [Google Scholar] [CrossRef] [Green Version]
- Hinkley, J.M.; Morton, A.B.; Ichinoseki-Sekine, N.; Huertas, A.M.; Smuder, A.J. Exercise Training Prevents Doxorubicin-induced Mitochondrial Dysfunction of the Liver. Med. Sci. Sport Exerc. 2019, 6, 1106–1115. [Google Scholar] [CrossRef]
- Gualano, B.; Roschel, H.; Lancha Junior, A.H.; Brightbill, C.E.; Rawson, E.S. In sickness and in health: The widespread application of creatine supplementation. Amino Acids 2012, 43, 519–529. [Google Scholar] [CrossRef]
- Gualano, B.; Artioli, G.G.; Poortmans, J.R.; Lancha Junior, A.H. Exploring the therapeutic role of creatine supplementation. Amino Acids 2010, 38, 31–44. [Google Scholar] [CrossRef]
- Deminice, R.; Cella, P.S.; Padilha, C.S.; Borges, F.H.; Costa Mendes da Silva, L.E.; Campos-Ferraz, P.L. Creatine supplementation prevents hyperhomocysteinemia, oxidative stress and cancer-induced cachexia progression in Walker-256 tumor-bearing rats. Amino Acids 2016, 48, 2015–2024. [Google Scholar] [CrossRef]
- Deminice, R.; de Castro, G.S.; Brosnan, M.E.; Brosnan, J.T. Creatine supplementation as a possible new therapeutic approach for fatty liver disease: Early findings. Amino Acids 2016, 48, 1983–1991. [Google Scholar] [CrossRef]
- Deminice, R.; da Silva, R.P.; Lamarre, S.G.; Brown, C.; Furey, G.N.; McCarter, S.A. Creatine supplementation prevents the accumulation of fat in the livers of rats fed a high-fat diet. J. Nutr. 2011, 141, 1799–1804. [Google Scholar] [CrossRef] [Green Version]
- Aljobaily, N.; Viereckl, M.J.; Hydock, D.S.; Aljobaily, H.; Wu, T.Y.; Busekrus, R. Creatine alleviates doxorubicin-induced liver damage by inhibiting liver fibrosis, inflammation, oxidative stress, and cellular senescence. Nutrients 2020, 13, 41. [Google Scholar] [CrossRef] [PubMed]
- Padilha, C.S.; Cella, P.S.; Chimin, P.; Voltarelli, F.A.; Marinello, P.C.; Testa, M.T.J.; Guarnier, F.A.; Duarte, J.A.; Deminice, R. Resistance Training’s Ability to Prevent Cancer-induced Muscle Atrophy Extends Anabolic Stimulus. Med. Sci. Sports Exerc. 2021, 53, 1572–1582. [Google Scholar] [CrossRef] [PubMed]
- Padilha, C.S.; Borges, F.H.; Costa Mendes da Silva, L.E.; Frajacomo, F.T.T.; Jordao, A.A.; Duarte, J.A.; Deminice, R. Resistance exercise attenuates skeletal muscle oxidative stress, systemic pro-inflammatory state, and cachexia in Walker-256 tumor-bearing rats. Appl. Physiol. Nutr. Metab. 2017, 42, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Cella, P.S.; Marinello, P.C.; Borges, F.H.; Ribeiro, D.F.; Chimin, P.; Testa, M.T.J.; Duarte, J.A.; Deminice, R. Creatine supplementation in Walker-256 tumor-bearing rats prevents skeletal muscle atrophy by attenuating systemic inflammation and protein degradation signaling. Eur. J. Nutr. 2020, 59, 661–669. [Google Scholar] [CrossRef]
- Mori, N.; Maeda, K.; Fukami, Y.; Matsuyama, R.; Nonogaki, T.; Kato, R. High SARC-F score predicts poor survival of patients with cancer receiving palliative care. Support Care Cancer 2022, 30, 4065–4072. [Google Scholar] [CrossRef]
- Deminice, R.; Portari, G.V.; Vannucchi, H.; Jordao, A.A. Effects of creatine supplementation on homocysteine levels and lipid peroxidation in rats. Br. J. Nutr. 2009, 102, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Duarte, J.A.; Leão, A.; Magalhães, J.; Ascensão, A.; Bastos, M.L.; Amado, F.L. Strenuous exercise aggravates MDMA-induced skeletal muscle damage in mice. Toxicology 2005, 206, 349–358. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2007, 1, 3159–3165. [Google Scholar] [CrossRef]
- Spirlandeli, A.L.; Deminice, R.; Jordao, A.A. Plasma Malondialdehyde as Biomarker of Lipid Peroxidation: Effects of Acute Exercise. Int. J. Sports Med. 2014, 35, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Witko-Sarsat, V.; Friedlander, M.; Capeillère-Blandin, C.; Nguyen-Khoa, T.; Zingraff, J.; Jungers, P. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int. 1996, 49, 1304–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, A.D.; Marchant, E.D.; Munk, D.J.; Watt, R.K.; Hansen, J.M.; Thomson, D.M. Multitissue analysis of exercise and metformin on doxorubicin-induced iron dysregulation. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E922–E930. [Google Scholar] [CrossRef]
- Khorana, A.A.; Tullio, K.; Elson, P.; Pennell, N.A.; Grobmyer, S.R.; Kalady, M.F. Time to initial cancer treatment in the United States and association with survival over time: An observational study. PLoS ONE 2019, 14, e0213209. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.W.; Wilson, R.L.; Christopher, C.N.; Normann, A.J.; Barnes, O.; Lesansee, J.D. Exercise Cardio-Oncology: Exercise as a Potential Therapeutic Modality in the Management of Anthracycline-Induced Cardiotoxicity. Front. Cardiovasc. Med. 2022, 8, 805735. [Google Scholar] [CrossRef]
- Bredahl, E.C.; Sharif, S.; Siedlik, J.A.; Wagner, M.K.; Twaddell, M.D.; Tigner, A.T. Resistance Training during Chemotherapy with Doxorubicin. Med. Sci. Sports Exerc. 2020, 52, 2529–2537. [Google Scholar] [CrossRef]
- Smuder, A.J. Exercise stimulates beneficial adaptations to diminish doxorubicin-induced cellular toxicity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, 662–672. [Google Scholar] [CrossRef]
- Ahmadian, M.; Dabidi Roshan, V.; Leicht, A.S. Age-related effect of aerobic exercise training on antioxidant and oxidative markers in the liver challenged by doxorubicin in rats. Free Radic. Res. 2018, 52, 775–782. [Google Scholar] [CrossRef]
- Porter, C.; Reidy, P.T.; Bhattarai, N.; Sidossis, L.S.; Rasmussen, B.B. Resistance Exercise Training Alters Mitochondrial Function in Human Skeletal Muscle. Med. Sci. Sports Exerc. 2015, 47, 1922–1931. [Google Scholar] [CrossRef] [Green Version]
- Ko, T.H.; Marquez, J.C.; Kim, H.K.; Jeong, S.H.; Lee, S.R.; Youm, J.B. Resistance exercise improves cardiac function and mitochondrial efficiency in diabetic rat hearts. Pflug. Arch. Eur. J. Physiol. 2018, 470, 263–275. [Google Scholar] [CrossRef]
- Lawler, J.M.; Barnes, W.S.; Wu, G.; Song, W.; Demaree, S. Direct antioxidant properties of creatine. Biochem. Biophys. Res. Commun. 2002, 290, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Sestili, P.; Martinelli, C.; Colombo, E.; Barbieri, E.; Potenza, L.; Sartini, S. Creatine as an antioxidant. Amino Acids 2011, 40, 1385–1396. [Google Scholar] [CrossRef]
- Sestili, P.; Barbieri, E.; Stocchi, V. Effects of Creatine in Skeletal Muscle Cells and in Myoblasts Differentiating under Normal or Oxidatively Stressing Conditions. Mini Rev. Med. Chem. 2016, 16, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Deminice, R.; de Castro, G.S.F.; Francisco, L.V.; Costa Mendes da Silva, L.E.; Cardoso, J.F.R.; Frajocomo, F.T.T.; Jordão, A.A. Creatine supplementation prevents fatty liver in rats fed choline-deficient diet: A burden of one-carbon and fatty acid metabolism. J. Nutr. Biochem. 2015, 26, 391–397. [Google Scholar] [CrossRef]
- Barbieri, E.; Guescini, M.; Calcabrini, C.; Valloraini, L.; Diaz, A.R.; Fimognari, C. Creatine Prevents the Structural and Functional Damage to Mitochondria in Myogenic, Oxidatively Stressed C2C12 Cells and Restores Their Differentiation Capacity. Oxid. Med. Cell. Longev. 2016, 12, 5152029. [Google Scholar] [CrossRef]
- Alves, C.R.R.; Ferreira, J.C.; Siqueira-Filho, M.A.; Carvalho, C.R.; Lancha-Junior, A.H.; Gualano, B. Creatine-induced glucose uptake in type 2 diabetes: A role for AMPK-α? Amino Acids 2012, 43, 1803–1807. [Google Scholar] [CrossRef]
- Sestili, P.; Ambrogini, P.; Barbieri, E.; Sartini, S.; Fimognari, C.; Calcabrini, C. New insights into the trophic and cytoprotective effects of creatine in in vitro and in vivo models of cell maturation. Amino Acids 2015, 48, 1897–1911. [Google Scholar] [CrossRef] [PubMed]
- Wallimann, T.; Tokarska-Schlattner, M.; Schlattner, U. The creatine kinase system and pleiotropic effects of creatine. Amino Acids 2011, 40, 1271–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceddia, R.B.; Sweeney, G. Creatine supplementation increases glucose oxidation and AMPK phosphorylation and reduces lactate production in L6 rat skeletal muscle cells. J. Physiol. 2004, 555, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; Moura, L.P.; Junior, R.C.V.; Junior, M.C.; Dalia, R.A.; Sponton, A.C. Creatine supplementation and oxidative stress in rat. J. Int. Soc. Sports Nutr. 2013, 10, 54. [Google Scholar] [CrossRef] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa Godinho, L.R.L.; Cella, P.S.; Guimarães, T.A.S.; Palma, G.H.D.; Nunes, J.H.C.; Deminice, R. Creatine Supplementation Potentiates Exercise Protective Effects against Doxorubicin-Induced Hepatotoxicity in Mice. Antioxidants 2023, 12, 823. https://doi.org/10.3390/antiox12040823
Costa Godinho LRL, Cella PS, Guimarães TAS, Palma GHD, Nunes JHC, Deminice R. Creatine Supplementation Potentiates Exercise Protective Effects against Doxorubicin-Induced Hepatotoxicity in Mice. Antioxidants. 2023; 12(4):823. https://doi.org/10.3390/antiox12040823
Chicago/Turabian StyleCosta Godinho, Loriane R. L., Paola S. Cella, Tatiana A. S. Guimarães, Guilherme H. Dantas Palma, Jonathan H. C. Nunes, and Rafael Deminice. 2023. "Creatine Supplementation Potentiates Exercise Protective Effects against Doxorubicin-Induced Hepatotoxicity in Mice" Antioxidants 12, no. 4: 823. https://doi.org/10.3390/antiox12040823