



Food Antioxidants and Their Interaction with Human Proteins
 , , and
, , and
Abstract
:1. Food Antioxidants
2. Food Antioxidants, Their Stability, and Availability under Physiological Conditions
3. Binding Characteristics of Proteins and Their Interaction with Food Antioxidants
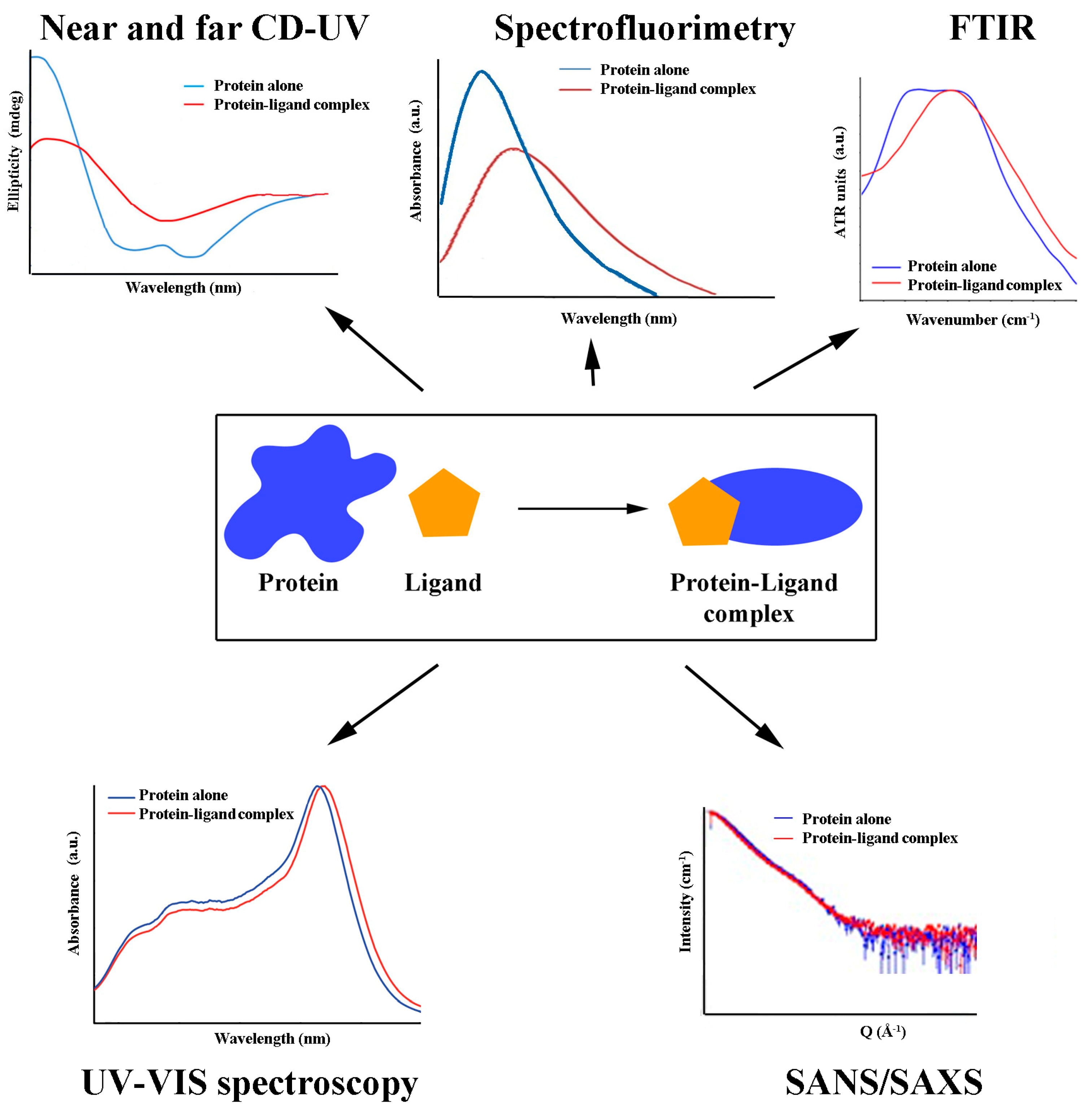
4. Methods for Studying Antioxidant/Protein Interactions
4.1. Binding Studies
4.2. Structural Aspects
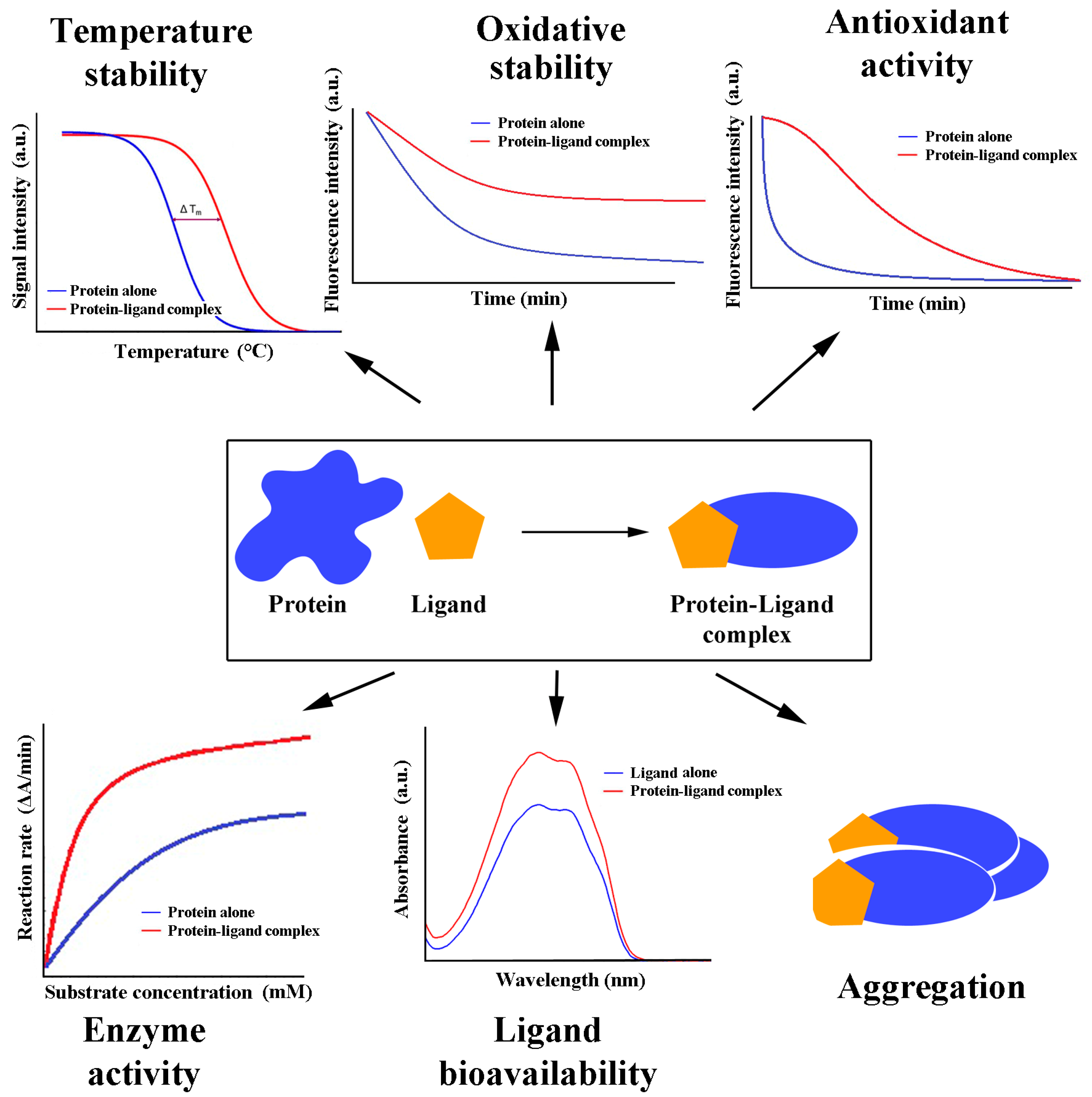
4.3. Functional Aspects

{kind=link}
{kind=link}
| Antioxidant Class (Subclass) | Chemical Compound | Effects (the Binding Constant in M−1, if Available) [Reference] |
|---|---|---|
| Flavonoids (Flavonols) | Quercetin | No significant influence on the structure [87]; no effects were studied [88]; inhibition of the oxidation of HSA-bound linoleic acid (1.2 × 105 at 25 °C) [89] |
| Fisetin | No effects were studied (1.2 × 105 at 25 °C) [88] | |
| Galangin | No effects were studied (2.3 × 105 at 25 °C) [88] | |
| Rhamnetin | No effects were studied (1.3 × 105 at 25 °C) [88] | |
| Myricetin | No effects were studied [90] | |
| Kaempferol | HSA unfolding to some degree (3.5 × 105 at 25 °C) [91]; negligible structural alteration (3.5 × 105 at 25 °C) [92] | |
| Morin | Reduction of α-helix and β-sheet structures (1.1 × 105 at 37 °C) [93] | |
| Astilbin | Reduction of α-helix content of HSA and antioxidant capacity of astilbin (4.5 × 105 at 37 °C) [94] | |
| Flavonoids (Flavones) | Luteolin | Reduction of α-helix and increase in β-turn structures; altered configuration of two disulfide bridges (1.6 × 105 at 25 °C) [95]; HSA stabilization by inhibition of fibrillation and glycation [96] |
| Apigenin | No effects were studied (1.3 × 105 at 25 °C [88]; 4.6 × 106 at 20 °C [97]) | |
| Rutin | No effects were studied (0.7 × 105 at 25 °C) [98]; reduction in α-helix content (2.4 × 106 at 25 °C) [99] | |
| Chrysin | No effects were studied (2.0 × 105 at 25 °C [88]; 2.5 × 105 at 25 °C [100]); stabilization of HSA by inhibition of fibrillation and glycation [96] | |
| Diosmetin | No effects were studied [88]; slight alteration of HSA structure (1.2 × 105 at 25 °C) [101] | |
| Flavone | No effects were studied (0.6 × 105 at 25 °C) [88] | |
| Trimethoxy flavone | Partial unfolding of protein secondary structure (1.0 × 103 at 25 °C) [102] | |
| Flavonoids (Isoflavones) | Genistein | Binding of oleic acid decreases affinity to HSA (7.8 × 106 at 20 °C) [103]; reduction in α-helix content (0.2 × 105 at 25 °C) [104]; no effects were studied (0.5 × 105 at 25 °C [88]; 1.5 × 105 at 27 °C [105]); |
| Formononetin | No effects were studied (0.2 × 105 at 25 °C) [88]; alteration of HSA structure (0.6 × 105 at 25 °C) [106] | |
| Daidzein | No effects were studied [105]; slight alteration of HSA structure (7.8 × 106 at 20 °C) [107] | |
| Prunetin | No effects were studied (0.4 × 105 at 25 °C) [88] | |
| Biochanin | Reduction of α-helix content (0.2 × 105 at 20 °C) [108] | |
| Flavonoids (Flavanols) | Catechin | Reduction of α-helix content (2.9 × 105 at 25 °C [80]; 0.2 × 105 at 20 °C [109]) |
| Epicatechin | Prolonged stabilization of ligand [110] | |
| Epigallocatechin | Prolonged stabilization of ligand [110] | |
| Epicatechin gallate | Prolonged stabilization of ligand [110]; reduction in α-helix content (3.1 × 105 at 25 °C) [80] | |
| Epigallocatechin galate | Protection and stabilization of ligand from oxidation [111]; prolonged stabilization of ligand [110]; reduction in α-helix content (3.3 × 105 at 25 °C [80]; 3.2 × 105 at 25 °C [112]); increase in protein aggregation and promotion of heterogeneous aggregate formation [113] | |
| Flavonoids (Flavanonols) | Taxifolin | Alteration of HSA conformation (1.8 × 105 at 37 °C) [114]; reduction in α-helix content (1.1 × 105 at 25 °C) [115] |
| Flavonoids (Flavanones) | Hesperetin | Reduction of α-helix content (0.2 × 105 at 25 °C [116]; 0.8 × 105 at 25 °C [117]) |
| Narirutin | No effects were studied (0.7 × 105 at 27 °C) [118] | |
| Naringin | No effects were studied (0.3 × 105 at 27 °C) [118] | |
| Nobiletin | No effects were studied (1.3 × 105 at 27 °C) [118] | |
| Tangeretin | No effects were studied (1.0 × 105 at 27 °C) [118] | |
| Naringenin | The binding of oleic acid decreases its affinity to HSA (5.3 × 106 at 20 °C) [103]; no effects were studied (0.7 × 105 at 27 °C) [118] | |
| Flavanone | No effects were studied (0.5 × 105 at 25 °C) [88] | |
| Sakuranetin | No effects were studied (0.2 × 105 at 25 °C) [88] | |
| Flavonoids (Anthocyanins) | Pelargonidin | Lower pH induces stronger binding (2.1 × 105 at 37 °C) [119] |
| Cyanidin | Lower pH induces stronger binding (3.1 × 105 at 37 °C) [119] | |
| Delphinidin | Lower pH induces stronger binding (3.4 × 105 at 37 °C) [119] | |
| Malvidin | Lower pH induces stronger binding (1.7 × 105 at 37 °C) [119] | |
| Pelargonidin-3-O-glucoside | Lower pH induces stronger binding (3.7 × 105 at 37 °C) [119] | |
| Flavonoids (Dihydrochalcones) | Phloretin | Reduction of α-helix content; increased resistance to aggregation, fibrillation, and oxidative modification (5.4 × 105 at 37 °C) [120] |
| Phenolic acids (Benzoic acid derivatives) | p-hydroxybenzoic acid | No effects were studied (1.0 × 103 at 25 °C) [121] |
| Gallic acid | Increase its antioxidant activity (2.0 × 103 at 25 °C) [122]; increase in α-helix content (9.0 × 103 at 25 °C) [79]; transition of α-helix to β-turn structures (1.0 × 104 at 25 °C) [123] | |
| Ellagic acid | Reduction of α-helix content (1.6 × 105 at 25 °C) [124] | |
| Vanillic acid | No effects were studied (1.0 × 103 at 25 °C) [121] | |
| Isovanillic acid | No effects were studied (2.0 × 103 at 25 °C) [122] | |
| Syringic acid | Increase in antioxidant activity of ligand [122] | |
| Protocatechuic acid | Increase in antioxidant activity of protocatechuic acid (2.0 × 103 at 25 °C) [122] | |
| Gentisic acid | Increase in antioxidant activity of gentisic acid (5.0 × 103 at 25 °C) [122] | |
| Phenolic acids (Cinnamic acid derivatives) | Cinnamic acid | Reduction of α-helix content (0.4 × 105 at 25 °C) [125] |
| Caffeic acid | Reduction of α-helix content (1.6 × 105 at 25 °C) [125]; stabilization of protein structure (0.3 × 105 at 37 °C) [126]; inhibition of the oxidation of HSA-bound linoleic acid [89]; no effects were studied (0.3 × 105 at 37 °C) [127]; alteration of protein structure (0.2 × 105 at 25 °C) [128] | |
| Ferulic acid | The partial unfolding of HSA (0.3 × 105 at 25 °C) [129]; binding of ferulic acid inhibits the oxidation of HSA-bound linoleic acid [89]; no effects were studied (2.3 × 106 at 25 °C) [130] | |
| Sinapic acid | Transition of α-helix to β-turn structures (6.9 × 107 at 25 °C) [123] | |
| Rosmarinic acid | Changes in tertiary structure with the reduction in α-helix content (0.6 × 105 at 37 °C) [131]; Inhibition of protein glycation and aggregation [132] | |
| p-Coumaric acid | Reduction of α-helix content (1.1 × 105 at 25 °C) [125] | |
| Chlorogenic acid | The partial unfolding of HSA (0.4 × 105 at 25 °C) [129]; binding of chlorogenic acid inhibits oxidation of HSA-bound linoleic acid [89]; no effects were studied (9.2 × 106 at 25 °C) [130] | |
| Phenolic aldehydes | Vanillin | Reduction of α-helix content (0.6 × 105 at 37 °C) [133] |
| Protocatechuic aldehyde | Disordered structure of HSA (9.8 × 107 at 37 °C) [134] | |
| Terpenes (Monoterpenes) | Menthol | Reduction of α-helix content, an increase in β-sheet and random coils [135] |
| Cuminaldehyde | Reduction of α-helix content (8.0 × 103 at 25 °C) [136] | |
| Cuminol | Reduction of α-helix content (1.0 × 103 at 25 °C) [136] | |
| Saphranal | Reduction of α-helix content, an increase in β-sheet and random coils (3.0 × 103 at 25 °C) [137] | |
| Terpenes (Diterpenes) | Leoheterin | Increase of α-helix content (1.2 × 105 at 25 °C) [138] |
| Cafestol | Reduction of α-helix content, increased affinity for warfarin (5.0 × 103 at 25 °C) [139] | |
| 16-O-methylcafestol | Reduction of α-helix content, increased affinity for warfarin (8.0 × 103 at 25 °C) [139] | |
| Terpenes (Triterpenes) | Betulinic acid | Reduction of α-helix content, an increase in β-sheet and random coils (1.7 × 106 at 25 °C) [140] |
| Asiatic acid | Reduction of α-helix content, an increase in β-sheet and random coils (0.4 × 105 at 25 °C) [141] | |
| β-Carotene | Reduction of α-helix content, an increase in random coils and β-turns (2.7 × 105 at 37 °C) [142]; reduction in α-helix content, an increase in β-turns (3.0 × 105 at 37 °C) [143] | |
| Isorenieratene | Reduction of α-helix content, increase in β-turns (3.5 × 105 at 37 °C) [143] | |
| Terpenes (Xanthophylls) | Lutein | Reduction of α-helix content, increase in β-turns (3.5 × 105 at 37 °C) [143] |
| Astaxanthin | Reduction of α-helix content, an increase in random coils and β-turns (2.6 × 106 at 37 °C) [142] | |
| Apocarotenoids | Crocetin | Reduction of α-helix content, an increase in β-sheet and random coils (2.0 × 103 at 25 °C) [137] |
| Stilbenes | Resveratrol | Increase of α-helix content, displacement of aflatoxin B1 (6.4 × 106 at 25 °C) [144,145]; protein thermal stabilization [146] |
| Rhaponticin | Reduction of α-helix content (1.4 × 105 at 25 °C) [147] | |
| Esculin | Reduction of α-helix content (4.6 × 105 at 25 °C) [148] | |
| Esculetin | Reduction of α-helix and increase in β-sheet content (0.3 × 105 at 25 °C) [149]; reduction in α-helix content (0.7 × 105 at 37 °C) [148] | |
| Scopoletin | Reduction of α-helix content (2.6 × 105 at 25 °C) [86] | |
| Fraxin | Reduction of α-helix content (3.1 × 105 at 25 °C) [148] | |
| Fraxetin | Reduction of α-helix content (0.9 × 105 at 25 °C) [148] | |
| Daphnetin | Reduction of α-helix content (1.7 × 106 at 37 °C) [150] | |
| Osthole | Conformational change of HSA (1.0 × 105 at 25 °C) [151]; slight reduction in α-helix content, inhibition of HSA esterase activity (8.9 × 105 at 25 °C) [152] | |
| Vitamins | Ascorbic acid | Reduction of α-helix content and increase in β-sheet and random coils (0.2 × 105 at 25 °C) [153]; reduction in α-helix content (0.2 × 105 at 25 °C) [154]; slight changes in secondary structure (3.0 × 103 at 25 °C) [155] |
| α-Tocopherol | Reduction of α-helix content and random coils, an increase in β-sheet and β-turn (4.0 × 103 at 25 °C) [153]; impaired binding of diazepam (7.0 × 106 at 25 °C) [156] | |
| Retinol | Protein stabilization: Increase in α-helix content and reduction in β-sheet content (1.3 × 105 at 25 °C) [157] | |
| Sulfur-containing compounds | Lipoic/Dihydro- lipoic acid | Restoration of zinc ion binding to protein [63]; thermal stabilization of HSA, no effect on trypsin digestion (0.1 × 105 at 37 °C) [56] |
| Phycobilins | Phycocyanobilin | Displacement of bound bilirubin (2.2 × 106 at 25 °C) [37]; increase in α-helix content and reduction in random coils; increased thermal and proteolytic stability; and conformational change of ligand [49] |
| Antioxidant Class | Antioxidant Subclass | Chemical Compound | Effects (the Binding Constant in M−1, if Available) [Reference] |
|---|---|---|---|
| Flavonoids | Flavonols | Quercetin | Reduction of hydrophobicity in the microenvironment of Trp residue; the unfolding of protein backbone; increase in β-sheet followed by reduction in α-helix and β-turn structures [158] |
| Fisetin | Moderate interaction; thermal denaturation of protein (1.4 × 106 at 25 °C) [159] | ||
| Galangin | Reduction of hydrophobicity in the microenvironment of Trp; unfolding of transferrin backbone; increase in β-sheet followed by reduction in α-helix and β-turn structures [158] | ||
| Myricetin | Reduction of hydrophobicity in the microenvironment of Trp; unfolding of transferrin backbone; increase in β-sheet followed by reduction in α-helix and β-turn structures [158] | ||
| Kaempferol | Reduction of hydrophobicity in the microenvironment of Trp; unfolding of transferrin backbone; increase in β-sheet followed by reduction in α-helix and β-turn structures [158] | ||
| Flavones | Luteolin | Increase in hydrophobicity of the Trp microenvironment and thermal stabilization (1.0 × 105 at 35 °C) [160] | |
| Apigenin | Increase in α-helix content (6.7 × 104 at 25 °C) [161] Increase of hydrophobicity in the Trp microenvironment; thermal stabilization (1.0 × 105 at 35 °C) [160] | ||
| Rutin | Increase of hydrophobicity in the Trp microenvironment; thermal stabilization (2.1 × 105 at 35 °C) [160] | ||
| Isoflavones | Genistein | Slight increase in hydrophobicity in the Trp microenvironment; slight increase in α-helix content (1.3 × 104 at 25 °C) [162] | |
| Daidzein | Increase of hydrophobicity in the Trp microenvironment; increase in α-helix content (2.9 × 105 at 25 °C) [162] | ||
| Flavanones | Naringenin | Stabilization of transferrin structure; increase in α-helix content; high binding affinity (6.3 × 106 at 25 °C) [161]; high binding affinity; no structural effects were observed [163] | |
| Anthocyanins | Cyanidin | Changes of hydrophobicity in the microenvironment of Trp and Tyr [164] | |
| Phenolic acids | Cinnamic acid derivatives | Rosmarinic acid | Alteration of protein structure and conformation (4.7 × 107 at 18 °C) [165] |
| Carotenoids | Xanthophylls | β-Cryptoxanthin | No effects were studied [166] |
| Vitamins | Ascorbic acid | Reduction in α-helix and increase in β-sheet content; inhibition of ligand acid-free radical scavenging activity upon binding (1.1 × 104 at 25 °C) [167] |
| Antioxidant Class | Antioxidant Subclass | Chemical Compound | Effects (the Binding Constant in M−1, if Available) [Reference] |
|---|---|---|---|
| Flavonoids | Flavonols | Quercetin | Reduction of hydrophobicity in the Trp microenvironment and α-helix content; reduction in inhibitory activity (4.4 × 103 at 25 °C) [168] |
| Myricetin | Increase of hydrophobicity in the Trp and Tyr microenvironment; slight decrease in α-helix content (2.4 × 103 at 45 °C) [169] | ||
| Phenolic acids | Benzoic acid derivatives | Gallic acid | Reduction of hydrophobicity in the Trp microenvironment; increase in the α-helix content; reduction in inhibitory activity (2.9 × 104 at 25 °C) [170] |
| Cinnamic acid derivatives | Ferulic acid | No significant structural changes were observed (6.7 × 104 at 25 °C) [171] | |
| Vitamins | Ascorbic acid | Induction of slight conformational changes [172] |
| Antioxidant Class | Antioxidant Subclass | Chemical Compound | Effects (the Binding Constant in M−1, if Available) [Reference] |
|---|---|---|---|
| Flavonoids | Flavonols | Myricetin | Reduction of α-helix content (2.0 × 104 at 25 °C) [173] |
| Flavones | Rutin | No effects were studied (2.1 × 104 at 25 °C) [173] | |
| Isoflavones | Genistein | Reduction of α-helix content (1.7 × 104 at 25 °C) [173] | |
| Puerarin | Reduction of α-helix content (8.8 × 103 at 25 °C) [173] | ||
| Flavanols | (-)-Epigallocatechin | No effects were studied [174] | |
| Flavanones | Hesperidin | Reduction of α-helix content (2.3 × 104 at 25 °C) [173] | |
| Naringin | Reduction of α-helix content (1.5 × 104 at 25 °C) [173] | ||
| Phenolic acids | Cinnamic acid derivatives | Caffeic acid | No effects were studied [175] |
| Ferulic acid | No effects were studied [175] | ||
| Stilbenes | Resveratrol | No effects were studied [176]; no fibrinogen unfolding/destabilization; mutually protective effect against free radical-induced oxidation (2.6 × 103 at 25 °C) [36] | |
| Tannins | Hydrolyzable tannins | Tannic acid | No effects were studied [176] |
| Sulfur- containing compounds | Fatty acids | Dehydrolypoic acid | Slight secondary structural alteration, more ordered protein molecular organization; formation of fibrin with thicker fibers; protection from oxidation (1.0 × 104 at 25 °C) [59] |
| Antioxidant Class | Antioxidant Subclass | Chemical Compound | Effects (the Binding Constant in M−1, if Available) [Reference] |
|---|---|---|---|
| Flavonoids | Flavonols | Quercetin | Reduction of α-helix content (1.2 × 104 at 25 °C) [177] |
| Dihydromyricetin | No alteration of secondary structure (2.8 × 104 at 23 °C) [178] | ||
| Isoflavones | Genistein | No alteration of secondary structure (3.5 × 104 at 25 °C) [179] | |
| Diadzein | No alteration of secondary structure (8.7 × 103 at 25 °C) [180] | ||
| Flavanols | Catechin | No alteration of α-helix content; pro-oxidative effect towards metHb formation at higher catechin concentration (7 × 107 at 25 °C) [181] | |
| Flavanones | Hesperidin | Stabilization of secondary structure (1.4 × 104 at 25 °C) [182] Slight alteration of secondary structure (2.2 × 104 at 25 °C) [183] | |
| Naringenin | No alteration of secondary structure (1.5 × 104 at 25 °C) [184] | ||
| Anthocyanidins | Procyanidin B3 | No effects were studied (0.9 × 103 at 37 °C) [185] | |
| Carotenoids | Terpenes | Astaxanthin | No effects were studied (2.2 × 109 at 37 °C) [185] |
| Carotenes | β-Carotene | No effects were studied (2.3 × 105 at 37 °C) [185] | |
| Tannins | Hydrolyzable tannins | Tannic acid | Structural changes and protein denaturation [186]; no effects were studied (1.5 × 104 at 25 °C) [187] |
| Vitamins | L-Ascorbic acid | No effects were studied (4.6 × 106 at 37 °C) [185] | |
| Tocopherols | α-Tocopherol | No effects were studied (3.2 × 103 at 37 °C) [185] | |
| Other | Turmeric | Curcumin | Unfolding of protein; reduction in thermal stability (4.9 × 105 at 20 °C) [188] |
5. Redox Status, Antioxidants, and Diseases
6. Future Perspectives in the Analysis of Antioxidant/Protein Interactions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gulcin, İ. Antioxidants and Antioxidant Methods: An Updated Overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunyadi, A. The mechanism(s) of action of antioxidants: From scavenging reactive oxygen/nitrogen species to redox signaling and the generation of bioactive secondary metabolites. Med. Res. Rev. 2019, 39, 2505–2533. [Google Scholar] [CrossRef] [Green Version]
- Rochette, L.; Ghibu, S.; Muresan, A.; Vergely, C. Alpha-Lipoic Acid: Molecular Mechanisms and Therapeutic Potential in Diabetes. Can. J. Physiol. Pharmacol. 2015, 93, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, M.C.; Orellana Palacios, J.C.; Hesami, G.; Jafarzadeh, S.; Lorenzo, J.M.; Domínguez, R.; Moreno, A.; Hadidi, M. Spectrophotometric Methods for Measurement of Antioxidant Activity in Food and Pharmaceuticals. Antioxidants 2022, 11, 2213. [Google Scholar] [CrossRef] [PubMed]
- Thorat, I.D.; Jagtap, D.D.; Mohapatra, D.; Joshi, D.C.; Sutar, R.F.; Kapdi, S.S. Antioxidants, Their Properties, Uses in Food Products and Their Legal Implications. Int. J. Food Stud. 2013, 2, 81–104. [Google Scholar] [CrossRef]
- Carlsen, M.H.; Halvorsen, B.L.; Holte, K.; Bøhn, S.K.; Dragland, S.; Sampson, L.; Willey, C.; Senoo, H.; Umezono, Y.; Sanada, C.; et al. The Total Antioxidant Content of More than 3100 Foods, Beverages, Spices, Herbs and Supplements Used Worldwide. Nutr. J. 2010, 9, 3. [Google Scholar] [CrossRef]
- MacDonald-Wicks, L.K.; Wood, L.G.; Garg, M.L. Methodology for the Determination of Biological Antioxidant Capacity in Vitro: A Review. J. Sci. Food Agric. 2006, 86, 2046–2056. [Google Scholar] [CrossRef]
- Cömert, E.D.; Gökmen, V. Physiological Relevance of Food Antioxidants. In Advances in Food and Nutrition Research; Toldrá, F., Ed.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2020; pp. 205–250. [Google Scholar]
- Neilson, A.P.; Goodrich, K.M.; Ferruzzi, M.G. Bioavailability and Metabolism of Bioactive Compounds From Foods. In Nutrition in the Prevention and Treatment of Disease; Coulston, A.M., Boushey, C.J., Ferruzzi, M.G., Delahanty, L.M., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2017; pp. 301–319. [Google Scholar]
- Rodriguez-Mateos, A.; Vauzour, D.; Krueger, C.G.; Shanmuganayagam, D.; Reed, J.; Calani, L.; Mena, P.; Del Rio, D.; Crozier, A. Bioavailability, Bioactivity and Impact on Health of Dietary Flavonoids and Related Compounds: An Update. Arch. Toxicol. 2014, 88, 1803–1853. [Google Scholar] [CrossRef]
- Selma, M.V.; Espín, J.C.; Tomás-Barberán, F.A. Interaction between Phenolics and Gut Microbiota: Role in Human Health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef]
- Cömert, E.D.; Gökmen, V. Antioxidants Bound to an Insoluble Food Matrix: Their Analysis, Regeneration Behavior, and Physiological Importance. Compr. Rev. Food Sci. Food Saf. 2017, 16, 382–399. [Google Scholar] [CrossRef] [Green Version]
- Dupas, C.J.; Marsset-Baglieri, A.C.; Ordonaud, C.S.; Ducept, F.M.G.; Maillard, M.-N. Coffee Antioxidant Properties: Effects of Milk Addition and Processing Conditions. J. Food Sci. 2006, 71, S253–S258. [Google Scholar] [CrossRef]
- Duarte, G.S.; Farah, A. Effect of Simultaneous Consumption of Milk and Coffee on Chlorogenic Acids’ Bioavailability in Humans. J. Agric. Food Chem. 2011, 59, 7925–7931. [Google Scholar] [CrossRef]
- Chima, B.; Mathews, P.; Morgan, S.; Johnson, S.A.; Van Buiten, C.B. Physicochemical Characterization of Interactions between Blueberry Polyphenols and Food Proteins from Dairy and Plant Sources. Foods 2022, 11, 2846. [Google Scholar] [CrossRef] [PubMed]
- Reboul, E. Mechanisms of Carotenoid Intestinal Absorption: Where Do We Stand? Nutrients 2019, 11, 838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peinado, J.; López de Lerma, N.; Peinado, R.A. Synergistic Antioxidant Interaction between Sugars and Phenolics from a Sweet Wine. Eur. Food Res. Technol. 2010, 231, 363–370. [Google Scholar] [CrossRef]
- Agulló, V.; Villaño, D.; García-Viguera, C.; Domínguez-Perles, R. Anthocyanin Metabolites in Human Urine after the Intake of New Functional Beverages. Molecules 2020, 25, 371. [Google Scholar] [CrossRef] [Green Version]
- Cömert, E.D.; Gökmen, V. Effect of Food Combinations and Their Co-Digestion on Total Antioxidant Capacity under Simulated Gastrointestinal Conditions. Curr. Res. Food Sci. 2022, 5, 414–422. [Google Scholar] [CrossRef]
- Kumar, S.; Ma, B.; Tsai, C.-J.; Sinha, N.; Nussinov, R. Folding and Binding Cascades: Dynamic Landscapes and Population Shifts. Protein Sci. 2000, 9, 10–19. [Google Scholar] [CrossRef]
- Weikl, T.R.; Paul, F. Conformational Selection in Protein Binding and Function. Protein Sci. 2014, 23, 1508–1518. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, N.; Abyzov, A.; Clarke, D.; Shou, C.; Gerstein, M.B. Integration of Protein Motions with Molecular Networks Reveals Different Mechanisms for Permanent and Transient Interactions. Protein Sci. 2011, 20, 1745–1754. [Google Scholar] [CrossRef] [Green Version]
- Hatasa, Y.; Chikazawa, M.; Furuhashi, N.; Nakashima, F.; Shibata, T.; Kondo, T.; Akagawa, M.; Hamagami, H.; Tanaka, H.; Tachibana, H.; et al. Oxidative Deamination of Serum Albumins by (-)-Epigallocatechin-3-O-Gallate: A Potential Mechanism for the Formation of Innate Antigens by Antioxidants. PLoS ONE 2016, 11, e0153002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, T.; Mori, T.; Tanaka, T.; Mizuno, D.; Yamaji, R.; Kumazawa, R.; Nakayama, T.; Akagawa, M. Covalent modification of proteins by green tea polyphenol (-)-epigallocatechin-3-gallate through autoxidation. Free Radic. Biol. Med. 2008, 45, 1384–1394. [Google Scholar] [CrossRef]
- Walle, T.; Vincent, T.S.; Walle, U.K. Evidence of covalent binding of the dietary flavonoid quercetin to DNA and protein in human intestinal and hepatic cells. Biochem. Pharmacol. 2003, 65, 1603–1610. [Google Scholar] [CrossRef]
- Kaldas, M.I.; Walle, U.K.; van der Woude, H.; McMillan, J.E.M.; Walle, T. Covalent binding of the flavonoid quercetin to human serum albumin. J. Agric. Food Chem. 2005, 53, 4194–4197. [Google Scholar] [CrossRef] [PubMed]
- Saqib, U.; Kelley, T.T.; Panguluri, S.K.; Liu, D.; Savai, R.; Baig, M.S.; Schürer, S.C. Polypharmacology or Promiscuity? Structural Interactions of Resveratrol With Its Bandwagon of Targets. Front. Pharmacol. 2018, 9, 1201. [Google Scholar] [CrossRef] [Green Version]
- Nwachukwu, J.C.; Srinivasan, S.; Bruno, N.E.; Parent, A.A.; Hughes, T.S.; Pollock, J.A.; Gjyshi, O.; Cavett, V.; Nowak, J.; Garcia-Ordonez, R.D.; et al. Resveratrol Modulates the Inflammatory Response via an Estrogen Receptor-Signal Integration Network. eLife 2014, 3, e02057. [Google Scholar] [CrossRef] [PubMed]
- Baruah, I.; Kashyap, C.; Guha, A.K.; Borgohain, G. Insights into the Interaction between Polyphenols and β-Lactoglobulin through Molecular Docking, MD Simulation, and QM/MM Approaches. ACS Omega 2022, 7, 23083–23095. [Google Scholar] [CrossRef]
- Shafqat, N.; Muniz, J.R.C.; Pilka, E.S.; Papagrigoriou, E.; von Delft, F.; Oppermann, U.; Yue, W.W. Insight into S-Adenosylmethionine Biosynthesis from the Crystal Structures of the Human Methionine Adenosyltransferase Catalytic and Regulatory Subunits. Biochem. J. 2013, 452, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Latruffe, N.; Menzel, M.; Delmas, D.; Buchet, R.; Lançon, A. Compared Binding Properties between Resveratrol and Other Polyphenols to Plasmatic Albumin: Consequences for the Health Protecting Effect of Dietary Plant Microcomponents. Molecules 2014, 19, 17066–17077. [Google Scholar] [CrossRef] [Green Version]
- Gomes, M.B.; Negrato, C.A. Alpha-Lipoic Acid as a Pleiotropic Compound with Potential Therapeutic Use in Diabetes and Other Chronic Diseases. Diabetol. Metab. Syndr. 2014, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Atukeren, P.; Aydin, S.; Uslu, E.; Gumustas, M.K.; Cakatay, U. Redox Homeostasis of Albumin in Relation to Alpha-Lipoic Acid and Dihydrolipoic Acid. Oxid. Med. Cell. Longev. 2010, 3, 206–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Weert, M.; Stella, L. Fluorescence Quenching and Ligand Binding: A Critical Discussion of a Popular Methodology. J. Mol. Struct. 2011, 998, 144–150. [Google Scholar] [CrossRef]
- Zhang, J.; Mi, Q.; Shen, M. Resveratrol Binding to Collagen and Its Biological Implication. Food Chem. 2012, 131, 879–884. [Google Scholar] [CrossRef]
- Gligorijević, N.; Radomirović, M.; Rajković, A.; Nedić, O.; Ćirković Veličković, T. Fibrinogen Increases Resveratrol Solubility and Prevents It from Oxidation. Foods 2020, 9, 780. [Google Scholar] [CrossRef]
- Minic, S.L.; Milcic, M.; Stanic-Vucinic, D.; Radibratovic, M.; Sotiroudis, T.G.; Nikolic, M.R.; Velickovic, T.Ć. Phycocyanobilin, a Bioactive Tetrapyrrolic Compound of Blue-Green Alga Spirulina, Binds with High Affinity and Competes with Bilirubin for Binding on Human Serum Albumin. RSC Adv. 2015, 5, 61787–61798. [Google Scholar] [CrossRef] [Green Version]
- Frazier, R.A.; Papadopoulou, A.; Green, R.J. Isothermal Titration Calorimetry Study of Epicatechin Binding to Serum Albumin. J. Pharm. Biomed. Anal. 2006, 41, 1602–1605. [Google Scholar] [CrossRef]
- Eaton, J.D.; Williamson, M.P. Multi-Site Binding of Epigallocatechin Gallate to Human Serum Albumin Measured by NMR and Isothermal Titration Calorimetry. Biosci. Rep. 2017, 37, BSR20170209. [Google Scholar] [CrossRef] [Green Version]
- Duff, M.R., Jr.; Grubbs, J.; Howell, E.E. Isothermal Titration Calorimetry for Measuring Macromolecule-Ligand Affinity. J. Vis. Exp. 2011, 55, 2796. [Google Scholar] [CrossRef] [Green Version]
- Rich, R.L.; Day, Y.S.N.; Morton, T.A.; Myszka, D.G. High-Resolution and High-Throughput Protocols for Measuring Drug/Human Serum Albumin Interactions Using BIACORE. Anal. Biochem. 2001, 15, 197–207. [Google Scholar] [CrossRef]
- Ishii, K.; Noda, M.; Uchiyama, S. Mass spectrometric analysis of protein-ligand interactions. Biophys. Physicobiol. 2016, 13, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Minic, S.; Radomirovic, M.; Savkovic, N.; Radibratovic, M.; Mihailovic, J.; Vasovic, T.; Nikolic, M.; Milcic, M.; Stanic-Vucinic, D.; Cirkovic Velickovic, T. Covalent binding of food-derived blue pigment phycocyanobilin to bovine β-lactoglobulin under physiological conditions. Food Chem. 2018, 269, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Stanic-Vucinic, D.; Prodic, I.; Apostolovic, D.; Nikolic, M.; Cirkovic Velickovic, T. Structure and antioxidant activity of β-lactoglobulin-glycoconjugates obtained by high-intensity-ultrasound-induced Maillard reaction in aqueous model systems under neutral conditions. Food Chem. 2013, 138, 590–599. [Google Scholar] [CrossRef]
- Woody, R.W.; Dunker, A.K. Aromatic and Cystine Side-Chain Circular Dichroism in Proteins. In Circular Dichroism and the Conformational Analysis of Biomolecules; Fasman, G., Ed.; Plenum Press: New York, NY, USA, 1996; pp. 110–144. [Google Scholar]
- Micsonai, A.; Moussong, É.; Wien, F.; Boros, E.; Vadászi, H.; Murvai, N.; Lee, Y.-H.; Molnár, T.; Réfrégiers, M.; Goto, Y.; et al. BeStSel: Webserver for Secondary Structure and Fold Prediction for Protein CD Spectroscopy. Nucleic Acids Res. 2022, 50, W90–W98. [Google Scholar] [CrossRef]
- Pelton, J.T.; McLean, L.R. Spectroscopic Methods for Analysis of Protein Secondary Structure. Anal. Biochem. 2000, 277, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Yu, S. Fourier Transform Infrared Spectroscopic Analysis of Protein Secondary Structures. Acta Biochim. Biophys. Sin. 2007, 39, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radibratovic, M.; Minic, S.; Stanic-Vucinic, D.; Nikolic, M.; Milcic, M.; Cirkovic Velickovic, T. Stabilization of Human Serum Albumin by the Binding of Phycocyanobilin, a Bioactive Chromophore of Blue-Green Alga Spirulina: Molecular Dynamics and Experimental Study. PLoS ONE 2016, 11, e0167973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doniach, S. Changes in Biomolecular Conformation Seen by Small Angle X-Ray Scattering. Chem. Rev. 2001, 101, 1763–1778. [Google Scholar] [CrossRef]
- Rambo, R.P.; Tainer, J.A. Characterizing Flexible and Intrinsically Unstructured Biological Macromolecules by SAS Using the Porod-Debye Law. Biopolymers 2011, 95, 559–571. [Google Scholar] [CrossRef] [Green Version]
- Minić, S.; Annighöfer, B.; Hélary, A.; Hamdane, D.; Hui Bon Hoa, G.; Loupiac, C.; Brûlet, A.; Combet, S. Effect of Ligands on HP-Induced Unfolding and Oligomerization of β-Lactoglobulin. Biophys. J. 2020, 119, 2262–2274. [Google Scholar] [CrossRef]
- Gligorijević, N.; Minić, S.; Radibratović, M.; Papadimitriou, V.; Nedić, O.; Sotiroudis, T.G.; Nikolić, M.R. Nutraceutical Phycocyanobilin Binding to Catalase Protects the Pigment from Oxidation without Affecting Catalytic Activity. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 251, 119483. [Google Scholar] [CrossRef]
- Sjödin, T.; Hansson, R.; Sjöholm, I. Isolation and Identification of a Trypsin-Resistant Fragment of Human Serum Albumin with Bilirubin- and Drug-Binding Properties. Biochim. Biophys. Acta-Protein Struct. 1977, 494, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ghanizadeh, H.; Li, X.; Han, Z.; Qiu, Y.; Zhang, Y.; Chen, X.; Wang, A. A Study of the Interaction, Morphology, and Structure in Trypsin-Epigallocatechin-3-Gallate Complexes. Molecules 2021, 26, 4567. [Google Scholar] [CrossRef] [PubMed]
- Gligorijevic, N.; Sukalovic, V.; Minic, S.; Miljus, G.; Nedic, O.; Penezic, A. Physicochemical Characterisation of Dihydro-Alpha-Lipoic Acid Interaction with Human Serum Albumin by Multi-Spectroscopic and Molecular Modelling Approaches. J. Serb. Chem. Soc. 2021, 86, 795–807. [Google Scholar] [CrossRef]
- Kim, H.G.; Lee, J.H.; Lee, S.J.; Oh, J.-H.; Shin, E.; Jang, Y.P.; Lee, Y.-J. The Increased Cellular Uptake and Biliary Excretion of Curcumin by Quercetin: A Possible Role of Albumin Binding Interaction. Drug Metab. Dispos. 2012, 40, 1452–1455. [Google Scholar] [CrossRef] [Green Version]
- Malinowska, J.; Olas, B. Effect of Resveratrol on Hemostatic Properties of Human Fibrinogen and Plasma during Model of Hyperhomocysteinemia. Thromb. Res. 2010, 126, e379–e382. [Google Scholar] [CrossRef]
- Gligorijević, N.; Šukalović, V.; Penezić, A.; Nedić, O. Characterisation of the Binding of Dihydro-Alpha-Lipoic Acid to Fibrinogen and the Effects on Fibrinogen Oxidation and Fibrin Formation. Int. J. Biol. Macromol. 2020, 147, 319–325. [Google Scholar] [CrossRef]
- Rashtbari, S.; Khataee, S.; Iranshahi, M.; Moosavi-Movahedi, A.A.; Hosseinzadeh, G.; Dehghan, G. Experimental Investigation and Molecular Dynamics Simulation of the Binding of Ellagic Acid to Bovine Liver Catalase: Activation Study and Interaction Mechanism. Int. J. Biol. Macromol. 2020, 143, 850–861. [Google Scholar] [CrossRef]
- Collazos, N.; García, G.; Malagón, A.; Caicedo, O.; Vargas, E.F. Binding Interactions of a Series of Sulfonated Water-Soluble Resorcinarenes with Bovine Liver Catalase. Int. J. Biol. Macromol. 2019, 139, 75–84. [Google Scholar] [CrossRef]
- Rashtbari, S.; Dehghan, G.; Yekta, R.; Jouyban, A. Investigation of the Binding Mechanism and Inhibition of Bovine Liver Catalase by Quercetin: Multi-Spectroscopic and Computational Study. BioImpacts 2017, 7, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Al-Harthi, S.; Chandra, K.; Jaremko, Ł. Lipoic Acid Restores Binding of Zinc Ions to Human Serum Albumin. Front. Chem. 2022, 10, 942585. [Google Scholar] [CrossRef]
- Ding, S.; Jiang, H.; Fang, J. Regulation of Immune Function by Polyphenols. J. Immunol. Res. 2018, 2018, 1264074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negri, A.; Naponelli, V.; Rizzi, F.; Bettuzzi, S. Molecular Targets of Epigallocatechin-Gallate (EGCG): A Special Focus on Signal Transduction and Cancer. Nutrients 2018, 10, 1936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oettl, K.; Stauber, R.E. Physiological and Pathological Changes in the Redox State of Human Serum Albumin Critically Influence Its Binding Properties. Br. J. Pharmacol. 2007, 151, 580–590. [Google Scholar] [CrossRef]
- Minic, S.; Stanic-Vucinic, D.; Radomirovic, M.; Radibratovic, M.; Milcic, M.; Nikolic, M.; Cirkovic Velickovic, T. Characterization and Effects of Binding of Food-Derived Bioactive Phycocyanobilin to Bovine Serum Albumin. Food Chem. 2018, 239, 1090–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.-L.; Wang, G.-H.; Xiang, W.-Z.; Li, T.; He, H. Stability and Antioxidant Activity of Food-Grade Phycocyanin Isolated from Spirulina Platensis. Int. J. Food Prop. 2016, 19, 2349–2362. [Google Scholar] [CrossRef] [Green Version]
- Zou, D.; Xie, A. Influence of polyphenol-plasma protein interaction on the antioxidant properties of polyphenols. Curr. Drug Metab. 2013, 14, 451–455. [Google Scholar] [CrossRef]
- Romay, C.C.; Pascual, C.; Lissi, E.A. The reaction between ABTS radical cation and antioxidants and its use to evaluate the antioxidant status of serum samples. Braz. J. Med. Biol. Res. 1996, 29, 175–183. [Google Scholar]
- Rohn, S.; Rawel, H.M.; Kroll, J. Antioxidant activity of protein-bound quercetin. J. Agric. Food Chem. 2004, 52, 4725–4729. [Google Scholar] [CrossRef]
- Li, Q.; Wei, Q.; Yuan, E.; Yang, J.; Ning, Z. Interaction between four flavonoids and trypsin: Effect on the characteristics of trypsin and antioxidant activity of flavonoids. Int. J. Food Sci. 2014, 49, 1063–1069. [Google Scholar] [CrossRef]
- Mihajlovic, L.; Radosavljevic, J.; Nordlund, E.; Krstic, M.; Bohn, T.; Smit, J.; Buchert, J.; Cirkovic Velickovic, T. Peanut protein structure, polyphenol content and immune response to peanut proteins in vivo are modulated by laccase. Food Funct. 2016, 7, 2357. [Google Scholar] [CrossRef]
- Heck, T.; Faccio, G.; Richter, M.; Thöny-Meyer, L. Enzyme-catalyzed protein crosslinking. Appl. Microbiol. Biotechnol. 2013, 97, 461–475. [Google Scholar] [CrossRef] [Green Version]
- Boutureira, O.; Bernardes, G.J.L. Advances in chemical protein modification. Chem. Rev. 2015, 115, 2174–2195. [Google Scholar] [CrossRef]
- Radomirovic, M.; Minic, S.; Stanic-Vucinic, D.; Nikolic, M.; Van Haute, S.; Rajkovic, A.; Cirkovic Velickovic, T. Phycocyanobilin-modified β-lactoglobulin exhibits increased antioxidant properties and stability to digestion and heating. Food Hydrocoll. 2022, 123, 107169. [Google Scholar] [CrossRef]
- Stojadinovic, M.; Pieters, R.; Smit, J.; Velickovic, C. Cross-Linking of β-Lactoglobulin Enhances Allergic Sensitization Through Changes in Cellular Uptake and Processing. Toxicol. Sci. 2014, 140, 224–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tantoush, Z.; Stanic, D.; Stojadinovic, M.; Ognjenovic, J.; Mihajlovic, L.; Atanaskovic-Markovic, M.; Cirkovic-Velickovic, T. Digestibility and allergenicity of beta-lactoglobulin following laccase-mediated cross-linking in the presence of sour cherry phenolics. Food Chem. 2011, 125, 84–91. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, L.; LI, J.; Chen, X. Studies on the Interaction of Gallic Acid with Human Serum Albumin in Membrane Mimetic Environments. Talanta 2008, 76, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Chanphai, P.; Tajmir-Riahi, H.A. Tea Polyphenols Bind Serum Albumins: A Potential Application for Polyphenol Delivery. Food Hydrocoll. 2019, 89, 461–467. [Google Scholar] [CrossRef]
- Beema Shafreen, R.; Dymerski, T.; Namieśnik, J.; Jastrzębski, Z.; Vearasilp, S.; Gorinstein, S. Interaction of Human Serum Albumin with Volatiles and Polyphenols from Some Berries. Food Hydrocoll. 2017, 72, 297–303. [Google Scholar] [CrossRef]
- Beg, M.; Maji, A.; Nayim, S.; Aktara, M.N.; Sahoo, N.K.; Jana, G.C.; Islam, M.M.; Hossain, M. Biophysical Insights into the Interaction of Human Serum Albumin with Cassia Fistula Leaf Extracts Inspired Biogenic Potent Antibacterial and Anticancerous Gold Nanoparticles. J. Biomol. Struct. Dyn. 2021, 39, 4567–4581. [Google Scholar] [CrossRef]
- Roufegarinejad, L.; Jahanban-Esfahlan, A.; Sajed-Amin, S.; Panahi-Azar, V.; Tabibiazar, M. Molecular Interactions of Thymol with Bovine Serum Albumin: Spectroscopic and Molecular Docking Studies. J. Mol. Recognit. 2018, 31, e2704. [Google Scholar] [CrossRef]
- Chaturvedi, S.K.; Ahmad, E.; Khan, J.M.; Alam, P.; Ishtikhar, M.; Khan, R.H. Elucidating the Interaction of Limonene with Bovine Serum Albumin: A Multi-Technique Approach. Mol. Biosyst. 2015, 11, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Galdón, B.; Pinto Corraliza, C.; Cestero Carrillo, J.J.; Macías Laso, P. Spectroscopic Study of the Interaction between Lycopene and Bovine Serum Albumin. Luminescence 2013, 28, 765–770. [Google Scholar] [CrossRef]
- Cheng, Z. Studies on the Interaction between Scopoletin and Two Serum Albumins by Spectroscopic Methods. J. Lumin. 2012, 132, 2719–2729. [Google Scholar] [CrossRef]
- Sengupta, B.; Sengupta, P.K. Binding of Quercetin with Human Serum Albumin: A Critical Spectroscopic Study. Biopolymers 2003, 72, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Rimac, H.; Debeljak, Ž.; Šakić, D.; Weitner, T.; Gabričević, M.; Vrček, V.; Zorc, B.; Bojić, M. Structural and Electronic Determinants of Flavonoid Binding to Human Serum Albumin: An Extensive Ligand-Based Study. RSC Adv. 2016, 6, 75014–75022. [Google Scholar] [CrossRef] [Green Version]
- Roche, M.; Dufour, C.; Loonis, M.; Reist, M.; Carrupt, P.-A.; Dangles, O. Olive Phenols Efficiently Inhibit the Oxidation of Serum Albumin-Bound Linoleic Acid and Butyrylcholine Esterase. Biochim. Biophys. Acta-Gen. Subj. 2009, 1790, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Xie, M.-X.; Liu, Y. Characterization of the Myricetin−Human Serum Albumin Complex by Spectroscopic and Molecular Modeling Approaches. Biomacromolecules 2007, 8, 2182–2189. [Google Scholar] [CrossRef]
- Matei, I.; Hillebrand, M. Interaction of Kaempferol with Human Serum Albumin: A Fluorescence and Circular Dichroism Study. J. Pharm. Biomed. Anal. 2010, 51, 768–773. [Google Scholar] [CrossRef]
- Das, S.; Hazarika, Z.; Sarmah, S.; Baruah, K.; Rohman, M.A.; Paul, D.; Jha, A.N.; Singha Roy, A. Exploring the Interaction of Bioactive Kaempferol with Serum Albumin, Lysozyme and Hemoglobin: A Biophysical Investigation Using Multi-Spectroscopic, Docking and Molecular Dynamics Simulation Studies. J. Photochem. Photobiol. B Biol. 2020, 205, 111825. [Google Scholar] [CrossRef]
- Xie, M.-X.; Long, M.; Liu, Y.; Qin, C.; Wang, Y.-D. Characterization of the Interaction between Human Serum Albumin and Morin. Biochim. Biophys. Acta-Gen. Subj. 2006, 1760, 1184–1191. [Google Scholar] [CrossRef]
- Han, X.; Sun, J.; Niu, T.; Mao, B.; Gao, S.; Zhao, P.; Sun, L. Molecular Insight into the Binding of Astilbin with Human Serum Albumin and Its Effect on Antioxidant Characteristics of Astilbin. Molecules 2022, 27, 4487. [Google Scholar] [CrossRef] [PubMed]
- Jurasekova, Z.; Marconi, G.; Sanchez-Cortes, S.; Torreggiani, A. Spectroscopic and Molecular Modeling Studies on the Binding of the Flavonoid Luteolin and Human Serum Albumin. Biopolymers 2009, 91, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Sarmah, S.; Das, S.; Roy, A.S. Protective Actions of Bioactive Flavonoids Chrysin and Luteolin on the Glyoxal Induced Formation of Advanced Glycation End Products and Aggregation of Human Serum Albumin: In Vitro and Molecular Docking Analysis. Int. J. Biol. Macromol. 2020, 165, 2275–2285. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.-L.; Lv, Z.; Liu, Z.-G.; Hu, Z.; Zou, G.-L. Study on Interaction between Apigenin and Human Serum Albumin by Spectroscopy and Molecular Modeling. J. Photochem. Photobiol. A Chem. 2007, 191, 104–113. [Google Scholar] [CrossRef]
- Pastukhov, A.V.; Levchenko, L.A.; Sadkov, A.P. Spectroscopic Study on Binding of Rutin to Human Serum Albumin. J. Mol. Struct. 2007, 842, 60–66. [Google Scholar] [CrossRef]
- Sengupta, P.; Sardar, P.S.; Roy, P.; Dasgupta, S.; Bose, A. Investigation on the Interaction of Rutin with Serum Albumins: Insights from Spectroscopic and Molecular Docking Techniques. J. Photochem. Photobiol. B Biol. 2018, 183, 101–110. [Google Scholar] [CrossRef]
- Mohos, V.; Fliszár-Nyúl, E.; Schilli, G.; Hetényi, C.; Lemli, B.; Kunsági-Máté, S.; Bognár, B.; Poór, M. Interaction of Chrysin and Its Main Conjugated Metabolites Chrysin-7-Sulfate and Chrysin-7-Glucuronide with Serum Albumin. Int. J. Mol. Sci. 2018, 19, 4073. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Wang, L.; Pan, J. Probing the Binding of the Flavonoid Diosmetin to Human Serum Albumin by Multispectroscopic Techniques. J. Agric. Food Chem. 2012, 60, 2721–2729. [Google Scholar] [CrossRef]
- Gokara, M.; Sudhamalla, B.; Amooru, D.G.; Subramanyam, R. Molecular Interaction Studies of Trimethoxy Flavone with Human Serum Albumin. PLoS ONE 2010, 5, e8834. [Google Scholar] [CrossRef] [Green Version]
- Bolli, A.; Marino, M.; Rimbach, G.; Fanali, G.; Fasano, M.; Ascenzi, P. Flavonoid Binding to Human Serum Albumin. Biochem. Biophys. Res. Commun. 2010, 398, 444–449. [Google Scholar] [CrossRef]
- Bian, Q.; Liu, J.; Tian, J.; Hu, Z. Binding of Genistein to Human Serum Albumin Demonstrated Using Tryptophan Fluorescence Quenching. Int. J. Biol. Macromol. 2004, 34, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Mahesha, H.G.; Singh, S.A.; Srinivasan, N.; Rao, A.G.A. A Spectroscopic Study of the Interaction of Isoflavones with Human Serum Albumin. FEBS J. 2006, 273, 451–467. [Google Scholar] [CrossRef]
- Li, Y.; He, W.; Dong, Y.; Sheng, F.; Hu, Z. Human Serum Albumin Interaction with Formononetin Studied Using Fluorescence Anisotropy, FT-IR Spectroscopy, and Molecular Modeling Methods. Bioorg. Med. Chem. 2006, 14, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, W.; Liu, H.; Yao, X.; Hu, Z. Daidzein Interaction with Human Serum Albumin Studied Using Optical Spectroscopy and Molecular Modeling Methods. J. Mol. Struct. 2007, 831, 144–150. [Google Scholar] [CrossRef]
- Xue, Z.; Cheng, A.; Li, Y.; Yu, W.; Kou, X. Investigating Interaction Between Biochanin A and Human Serum Albumin by Multi-Spectroscopic and Molecular Simulation Methods. Trans. Tianjin Univ. 2017, 23, 325–333. [Google Scholar] [CrossRef]
- Li, X.; Wang, S. Study on the Interaction of (+)-Catechin with Human Serum Albumin Using Isothermal Titration Calorimetry and Spectroscopic Techniques. New J. Chem. 2015, 39, 386–395. [Google Scholar] [CrossRef]
- Zinellu, A.; Sotgia, S.; Scanu, B.; Forteschi, M.; Giordo, R.; Cossu, A.; Posadino, A.M.; Carru, C.; Pintus, G. Human Serum Albumin Increases the Stability of Green Tea Catechins in Aqueous Physiological Conditions. PLoS ONE 2015, 10, e0134690. [Google Scholar] [CrossRef]
- Bae, M.-J.; Ishii, T.; Minoda, K.; Kawada, Y.; Ichikawa, T.; Mori, T.; Kamihira, M.; Nakayama, T. Albumin Stabilizes (-)-Epigallocatechin Gallate in Human Serum: Binding Capacity and Antioxidant Property. Mol. Nutr. Food Res. 2009, 53, 709–715. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, M.; Sun, B.; Liu, J.; Wei, X.; Wang, Z.; Wang, B.; Han, J. Calorimetric and Spectroscopic Studies on the Competitive Behavior between (−)-Epigallocatechin-3-Gallate and 5-Fluorouracil with Human Serum Albumin. J. Mol. Liq. 2017, 248, 330–339. [Google Scholar] [CrossRef]
- Sun, X.; Ferguson, H.N.; Hagerman, A.E. Conformation and Aggregation of Human Serum Albumin in the Presence of Green Tea Polyphenol (EGCG) and/or Palmitic Acid. Biomolecules 2019, 9, 705. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; He, H.; Zhang, M.; Lv, X.; Li, W.; Wang, R.; Chang, J. Investigation of the Interactions between Three Flavonoids and Human Serum Albumin by Isothermal Titration Calorimetry, Spectroscopy, and Molecular Docking. New J. Chem. 2022, 46, 12814–12824. [Google Scholar] [CrossRef]
- Mohseni-Shahri, F.S.; Housaindokht, M.R.; Bozorgmehr, M.R.; Moosavi-Movahedi, A.A. Influence of Taxifolin on the Human Serum Albumin–Propranolol Interaction: Multiple Spectroscopic and Chemometrics Investigations and Molecular Dynamics Simulation. J. Solution Chem. 2016, 45, 265–285. [Google Scholar] [CrossRef]
- Ding, F.; Diao, J.-X.; Sun, Y.; Sun, Y. Bioevaluation of Human Serum Albumin–Hesperidin Bioconjugate: Insight into Protein Vector Function and Conformation. J. Agric. Food Chem. 2012, 60, 7218–7228. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.-X.; Xu, X.-Y.; Wang, Y.-D. Interaction between Hesperetin and Human Serum Albumin Revealed by Spectroscopic Methods. Biochim. Biophys. Acta-Gen. Subj. 2005, 1724, 215–224. [Google Scholar] [CrossRef]
- Cao, H.; Chen, L.; Xiao, J. Binding Citrus Flavanones to Human Serum Albumin: Effect of Structure on Affinity. Mol. Biol. Rep. 2011, 38, 2257–2262. [Google Scholar] [CrossRef]
- Cahyana, Y.; Gordon, M.H. Interaction of Anthocyanins with Human Serum Albumin: Influence of PH and Chemical Structure on Binding. Food Chem. 2013, 141, 2278–2285. [Google Scholar] [CrossRef]
- Barreca, D.; Laganà, G.; Toscano, G.; Calandra, P.; Kiselev, M.A.; Lombardo, D.; Bellocco, E. The Interaction and Binding of Flavonoids to Human Serum Albumin Modify Its Conformation, Stability and Resistance against Aggregation and Oxidative Injuries. Biochim. Biophys. Acta-Gen. Subj. 2017, 1861, 3531–3539. [Google Scholar] [CrossRef]
- Myint, O.; Wattanapongpitak, S.; Supawat, B.; Kothan, S.; Udomtanakunchai, C.; Tima, S.; Tungjai, M. Protein Binding of 4-Hydroxybenzoic Acid and 4-Hydroxy-3-Methoxybenzoic Acid to Human Serum Albumin and Their Anti-Proliferation on Doxorubicin-Sensitive and Doxorubicin-Resistant Leukemia Cells. Toxicol. Rep. 2021, 8, 1381–1388. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, S.; Qin, Y.; Liu, J.; Liu, J.; Wang, Q.; Ren, F.; Zhang, H. Interaction of Phenolic Acids and Their Derivatives with Human Serum Albumin: Structure–Affinity Relationships and Effects on Antioxidant Activity. Food Chem. 2018, 240, 1072–1080. [Google Scholar] [CrossRef]
- Liu, Y.; Xie, M.-X.; Jiang, M.; Wang, Y.-D. Spectroscopic Investigation of the Interaction between Human Serum Albumin and Three Organic Acids. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2005, 61, 2245–2251. [Google Scholar] [CrossRef]
- Pattanayak, R.; Basak, P.; Sen, S.; Bhattacharyya, M. An Insight to the Binding of Ellagic Acid with Human Serum Albumin Using Spectroscopic and Isothermal Calorimetry Studies. Biochem. Biophys. Rep. 2017, 10, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Meng-Xia, X.; Dong, Z.; Yuan, L.; Xiao-Yu, L.; Xing, C. Spectroscopic Studies on the Interaction of Cinnamic Acid and Its Hydroxyl Derivatives with Human Serum Albumin. J. Mol. Struct. 2004, 692, 71–80. [Google Scholar] [CrossRef]
- Zhang, Y.; Yue, Y.; Li, J.; Chen, X. Studies on the Interaction of Caffeic Acid with Human Serum Albumin in Membrane Mimetic Environments. J. Photochem. Photobiol. B Biol. 2008, 90, 141–151. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Li, Q.; Chen, J.; Gao, X.; Chen, H.; Xiao, C.; Bian, L.; Zheng, J.; Zhao, X.; Zheng, X. Binding of Caffeic Acid to Human Serum Albumin by the Retention Data and Frontal Analysis. Biomed. Chromatogr. 2014, 28, 1881–1886. [Google Scholar] [CrossRef] [PubMed]
- Jahanban-Esfahlan, A.; Roufegarinejad, L.; Tabibiazar, M.; Lorenzo, J.; Amarowicz, R. Exploring the Interactions Between Caffeic Acid and Human Serum Albumin Using Spectroscopic and Molecular Docking Techniques. Pol. J. Food Nutr. Sci. 2021, 71, 69–77. [Google Scholar] [CrossRef]
- Kang, J.; Liu, Y.; Xie, M.; Li, S.; Jiang, M.; Wang, Y. Interactions of Human Serum Albumin with Chlorogenic Acid and Ferulic Acid. Biochim. Biophys. Acta-Gen. Subj. 2004, 1674, 205–214. [Google Scholar] [CrossRef]
- Sinisi, V.; Forzato, C.; Cefarin, N.; Navarini, L.; Berti, F. Interaction of Chlorogenic Acids and Quinides from Coffee with Human Serum Albumin. Food Chem. 2015, 168, 332–340. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Wang, X.; Qi, W.; Su, R.; He, Z. Affinity of Rosmarinic Acid to Human Serum Albumin and Its Effect on Protein Conformation Stability. Food Chem. 2016, 192, 178–187. [Google Scholar] [CrossRef]
- Shamsi, A.; Ahmed, A.; Khan, M.S.; Husain, F.M.; Bano, B. Rosmarinic Acid Restrains Protein Glycation and Aggregation in Human Serum Albumin: Multi Spectroscopic and Microscopic Insight—Possible Therapeutics Targeting Diseases. Int. J. Biol. Macromol. 2020, 161, 187–193. [Google Scholar] [CrossRef]
- Wang, X.; Xie, X.; Ren, C.; Yang, Y.; Xu, X.; Chen, X. Application of Molecular Modelling and Spectroscopic Approaches for Investigating Binding of Vanillin to Human Serum Albumin. Food Chem. 2011, 127, 705–710. [Google Scholar] [CrossRef]
- Tian, J.; Chen, C.; Xue, M. The Interaction between Protocatechuic Aldehyde and Human Serum Albumin Using Three-Dimensional Fluorescence Techniques. Spectroscopy 2011, 26, 195–201. [Google Scholar] [CrossRef]
- Yeggoni, D.P.; Rachamallu, A.; Dubey, S.; Mitra, A.; Subramanyam, R. Probing the Interaction Mechanism of Menthol with Blood Plasma Proteins and Its Cytotoxicity Activities. J. Biomol. Struct. Dyn. 2018, 36, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.S.; Rehman, M.T.; Al-Lohedan, H.; AlAjmi, M.F. Spectroscopic and Molecular Docking Investigation on the Interaction of Cumin Components with Plasma Protein: Assessment of the Comparative Interactions of Aldehyde and Alcohol with Human Serum Albumin. Int. J. Mol. Sci. 2022, 23, 4078. [Google Scholar] [CrossRef] [PubMed]
- Kanakis, C.D.; Tarantilis, P.A.; Tajmir-Riahi, H.A.; Polissiou, M.G. Crocetin, Dimethylcrocetin, and Safranal Bind Human Serum Albumin: Stability and Antioxidative Properties. J. Agric. Food Chem. 2007, 55, 970–977. [Google Scholar] [CrossRef]
- Ali, M.S.; Amina, M.; Al-Lohedan, H.A.; Al Musayeib, N.M. Human Serum Albumin Binding to the Biologically Active Labdane Diterpene “Leoheterin”: Spectroscopic and in Silico Analysis. J. Photochem. Photobiol. B Biol. 2018, 182, 9–17. [Google Scholar] [CrossRef]
- Guercia, E.; Forzato, C.; Navarini, L.; Berti, F. Interaction of Coffee Compounds with Serum Albumins. Part II: Diterpenes. Food Chem. 2016, 199, 502–508. [Google Scholar] [CrossRef] [Green Version]
- Subramanyam, R.; Gollapudi, A.; Bonigala, P.; Chinnaboina, M.; Amooru, D.G. Betulinic Acid Binding to Human Serum Albumin: A Study of Protein Conformation and Binding Affinity. J. Photochem. Photobiol. B Biol. 2009, 94, 8–12. [Google Scholar] [CrossRef]
- Gokara, M.; Malavath, T.; Kalangi, S.K.; Reddana, P.; Subramanyam, R. Unraveling the Binding Mechanism of Asiatic Acid with Human Serum Albumin and Its Biological Implications. J. Biomol. Struct. Dyn. 2014, 32, 1290–1302. [Google Scholar] [CrossRef]
- Li, X.; Wang, G.; Chen, D.; Lu, Y. β-Carotene and Astaxanthin with Human and Bovine Serum Albumins. Food Chem. 2015, 179, 213–221. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, Y.; Chen, M.; Xie, B.; Yang, J.; Chen, J.; Sun, Z. Isorenieratene Interaction with Human Serum Albumin: Multi-Spectroscopic Analyses and Docking Simulation. Food Chem. 2018, 258, 393–399. [Google Scholar] [CrossRef]
- N’ soukpoé-Kossi, C.N.; St-Louis, C.; Beauregard, M.; Subirade, M.; Carpentier, R.; Hotchandani, S.; Tajmir-Riahi, H.A. Resveratrol Binding to Human Serum Albumin. J. Biomol. Struct. Dyn. 2006, 24, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, M.A.; Javed, S. Investigating Binding Dynamics of Trans Resveratrol to HSA for an Efficient Displacement of Aflatoxin B1 Using Spectroscopy and Molecular Simulation. Sci. Rep. 2022, 12, 2400. [Google Scholar] [CrossRef]
- Stirpe, A.; Pantusa, M.; Rizzuti, B.; De Santo, M.P.; Sportelli, L.; Bartucci, R.; Guzzi, R. Resveratrol Induces Thermal Stabilization of Human Serum Albumin and Modulates the Early Aggregation Stage. Int. J. Biol. Macromol. 2016, 92, 1049–1056. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ji, Z.; Liang, X.; Li, G.; Yang, S.; Wei, S.; Zhao, Y.; Hu, X.; Fan, J. Studies on the Binding of Rhaponticin with Human Serum Albumin by Molecular Spectroscopy, Modeling and Equilibrium Dialysis. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2012, 87, 171–178. [Google Scholar] [CrossRef]
- Xu, L.; Yang, H.; Hu, R.; Liang, Y.; Li, Y.; Xu, W.; Fan, X.; Liu, Y. Comparing the Interaction of Four Structurally Similar Coumarins from Fraxinus Chinensis Roxb. with HSA through Multi-Spectroscopic and Docking Studies. J. Mol. Liq. 2021, 340, 117234. [Google Scholar] [CrossRef]
- Yue, Y.; Zhang, Y.; Qin, J.; Chen, X. Study of the Interaction between Esculetin and Human Serum Albumin by Multi-Spectroscopic Method and Molecular Modeling. J. Mol. Struct. 2008, 888, 25–32. [Google Scholar] [CrossRef]
- Liu, J.; Tian, J.; Li, Y.; Yao, X.; Hu, Z.; Chen, X. Binding of the Bioactive Component Daphnetin to Human Serum Albumin Demonstrated Using Tryptophan Fluorescence Quenching. Macromol. Biosci. 2004, 4, 520–525. [Google Scholar] [CrossRef]
- Yang, G.-D.; Li, C.; Zeng, A.-G.; Zhao, Y.; Yang, R.; Bian, X.-L. Fluorescence Spectroscopy of Osthole Binding to Human Serum Albumin. J. Pharm. Anal. 2013, 3, 200–204. [Google Scholar] [CrossRef] [Green Version]
- Bijari, N.; Shokoohinia, Y.; Ashrafi-Kooshk, M.R.; Ranjbar, S.; Parvaneh, S.; Moieni-Arya, M.; Khodarahmi, R. Spectroscopic Study of Interaction between Osthole and Human Serum Albumin: Identification of Possible Binding Site of the Compound. J. Lumin. 2013, 143, 328–336. [Google Scholar] [CrossRef]
- Li, X.; Chen, D.; Wang, G.; Lu, Y. Study of Interaction between Human Serum Albumin and Three Antioxidants: Ascorbic Acid, α-Tocopherol, and Proanthocyanidins. Eur. J. Med. Chem. 2013, 70, 22–36. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhu, Z.; Ni, Y. Interaction between Aspirin and Vitamin C with Human Serum Albumin as Binary and Ternary Systems. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 236, 118356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, Z.-Q.; Liu, X.-R.; Qi, Z.-D.; Jiang, F.-L.; Liu, Y. Conformation and Thermodynamic Properties of the Binding of Vitamin C to Human Serum Albumin. J. Solut. Chem. 2012, 41, 351–366. [Google Scholar] [CrossRef]
- Fanali, G.; Fasano, M.; Ascenzi, P.; Zingg, J.-M.; Azzi, A. α-Tocopherol Binding to Human Serum Albumin. BioFactors 2013, 39, 294–303. [Google Scholar] [CrossRef] [PubMed]
- N’soukpoé-Kossi, C.N.; Sedaghat-Herati, R.; Ragi, C.; Hotchandani, S.; Tajmir-Riahi, H.A. Retinol and Retinoic Acid Bind Human Serum Albumin: Stability and Structural Features. Int. J. Biol. Macromol. 2007, 40, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Han, L.; Song, Z.; Xu, R.; Wang, L. Comparative Study on the Interaction between Transferrin and Flavonols: Experimental and Computational Modeling Approaches. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2023, 288, 122128. [Google Scholar] [CrossRef]
- Sandu, N.; Popescu, A.I.; Chilom, C.G. Probing the Interaction of Fisetin with Human Serum Transferrin via Spectroscopic and Molecular Docking Approaches. J. Biomol. Struct. Dyn. 2022, 40, 9613–9619. [Google Scholar] [CrossRef] [PubMed]
- Sandu, N.; Chilom, C.G.; Popescu, A.I. Structural and molecular aspects of flavonoids as ligands for serum transferrin. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2021, 254, 119600. [Google Scholar] [CrossRef]
- Zhang, X.; Han, R.; Sun, X.; Li, G.; Yang, Q.; Li, Q.; Gai, W.; Zhang, M.; Chen, L.; Yang, G.; et al. The Effect of the Skeleton Structure of Flavanone and Flavonoid on Interaction with Transferrin. Bioorg. Med. Chem. Lett. 2013, 23, 6677–6681. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, X.; Liu, Y.; Han, R.; Lan, L.; Chen, H.; Sun, X.; Li, Q.; Tang, Y.-L. Interaction of Isoflavones with Different Structures and Transferrin. Spectrosc. Lett. 2016, 49, 596–601. [Google Scholar] [CrossRef]
- Khan, S.; Alhumaydhi, F.A.; Khan, M.S.; Sharaf, S.E.; Al Abdulmonem, W.; Hassan, M.I.; Shamsi, A.; Kumar Yadav, D. Exploring Binding Mechanism of Naringenin to Human Transferrin Using Combined Spectroscopic and Computational Methods: Towards Therapeutic Targeting of Neurodegenerative Diseases. J. Mol. Liq. 2022, 356, 119001. [Google Scholar] [CrossRef]
- Khashkhashi-Moghadam, S.; Ezazi-Toroghi, S.; Kamkar-Vatanparast, M.; Jouyaeian, P.; Mokaberi, P.; Yazdyani, H.; Amiri-Tehranizadeh, Z.; Reza Saberi, M.; Chamani, J. Novel Perspective into the Interaction Behavior Study of the Cyanidin with Human Serum Albumin-Holo Transferrin Complex: Spectroscopic, Calorimetric and Molecular Modeling Approaches. J. Mol. Liq. 2022, 356, 119042. [Google Scholar] [CrossRef]
- Shamsi, A.; Anwar, S.; Shahbaaz, M.; Mohammad, T.; Alajmi, M.F.; Hussain, A.; Hassan, I.; Ahmad, F.; Islam, A. Evaluation of Binding of Rosmarinic Acid with Human Transferrin and Its Impact on the Protein Structure: Targeting Polyphenolic Acid-Induced Protection of Neurodegenerative Disorders. Oxid. Med. Cell. Longev. 2020, 2020, 1245875. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Hazra, U.; Dutta, D. Role of β-Cryptoxanthin as an Antioxidant and Its Ability to Bind with Transferrin. Int. J. Biosci. Biochem. Bioinforma. 2019, 9, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Zhou, L.; Zhou, B.; Zhang, S.; Ma, X.; Zhou, H.; Tuo, X. Elucidation on the Interaction between Transferrin and Ascorbic Acid: A Study Based on Spectroscopic Analysis, Molecular Docking Technology, and Antioxidant Evaluation. J. Mol. Liq. 2022, 360, 119413. [Google Scholar] [CrossRef]
- Siddiqui, T.; Zia, M.K.; Ahsan, H.; Khan, F.H. Quercetin-Induced Inactivation and Conformational Alterations of Alpha-2-Macroglobulin: Multi-Spectroscopic and Calorimetric Study. J. Biomol. Struct. Dyn. 2020, 38, 4107–4118. [Google Scholar] [CrossRef]
- Ansari, S.; Ahsan, H.; Zia, M.K.; Gatasheh, M.K.; Khan, F.H. Exploring the Interaction of Myricetin with Human Alpha-2-Macroglobulin: Biophysical and in-Silico Analysis. J. Biol. Phys. 2023, 20, 29–48. [Google Scholar] [CrossRef]
- Siddiqui, T.; Zia, M.K.; Ali, S.S.; Ahsan, H.; Khan, F.H. Inactivation of Alpha-2-Macroglobulin by Photo-Illuminated Gallic Acid. J. Fluoresc. 2019, 29, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.A.; Sarwar, T.; Arif, H.; Ali, S.S.; Ahsan, H.; Tabish, M.; Khan, F.H. Spectroscopic and Thermodynamic Studies on Ferulic Acid—Alpha-2-Macroglobulin Interaction. J. Mol. Struct. 2017, 1144, 254–259. [Google Scholar] [CrossRef]
- Ali, S.S.; Zia, M.K.; Siddiqui, T.; Ahsan, H.; Khan, F.H. Influence of Ascorbic Acid on the Structure and Function of Alpha-2- Macroglobulin: Investigations Using Spectroscopic and Thermodynamic Techniques. Protein Pept. Lett. 2020, 27, 201–209. [Google Scholar] [CrossRef]
- Li, X.; Duan, H.; Song, Z.; Xu, R. Comparative Study on the Interaction between Fibrinogen and Flavonoids. J. Mol. Struct. 2022, 1262, 132963. [Google Scholar] [CrossRef]
- Suzuki, Y.; Isemura, M. Binding Interaction between (−)-Epigallocatechin Gallate Causes Impaired Spreading of Cancer Cells on Fibrinogen. Biomed. Res. 2013, 34, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Shafreen, R.M.B.; Lakshmi, S.A.; Pandian, S.K.; Park, Y.S.; Kim, Y.M.; Paśko, P.; Deutsch, J.; Katrich, E.; Gorinstein, S. Unraveling the Antioxidant, Binding and Health-Protecting Properties of Phenolic Compounds of Beers with Main Human Serum Proteins: In Vitro and In Silico Approaches. Molecules 2020, 25, 4962. [Google Scholar] [CrossRef] [PubMed]
- Shafreen, R.M.B.; Lakshmi, S.A.; Pandian, S.K.; Kim, Y.-M.; Deutsch, J.; Katrich, E.; Gorinstein, S. In Vitro and In Silico Interaction Studies with Red Wine Polyphenols against Different Proteins from Human Serum. Molecules 2021, 26, 6686. [Google Scholar] [CrossRef] [PubMed]
- Sett, R.; Paul, B.K.; Guchhait, N. Deciphering the Fluorescence Quenching Mechanism of a Flavonoid Drug Following Interaction with Human Hemoglobin. J. Phys. Org. Chem. 2022, 35, e4307. [Google Scholar] [CrossRef]
- Chen, T.; Zhu, S.; Shang, Y.; Ge, C.; Jiang, G. Binding of Dihydromyricetin to Human Hemoglobin: Fluorescence and Circular Dichroism Studies. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2012, 93, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Pahari, B.; Chakraborty, S.; Sengupta, B.; Chaudhuri, S.; Martin, W.; Taylor, J.; Henley, J.; Davis, D.; Biswas, P.K.; Sharma, A.K.; et al. Biophysical Characterization of Genistein in Its Natural Carrier Human Hemoglobin Using Spectroscopic and Computational Approaches. Food Nutr. Sci. 2013, 4, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, B.; Chakraborty, S.; Crawford, M.; Taylor, J.M.; Blackmon, L.E.; Biswas, P.K.; Kramer, W.H. Characterization of Diadzein–Hemoglobin Binding Using Optical Spectroscopy and Molecular Dynamics Simulations. Int. J. Biol. Macromol. 2012, 51, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Das, D.; Sen, S.; Sen, K. Caffeine and Catechin towards Prevention of Drug Induced Oxidation of Hemoglobin: A Spectroscopic Study. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 232, 118167. [Google Scholar] [CrossRef]
- Ding, F.; Sun, Y.; Diao, J.-X.; Li, X.-N.; Yang, X.-L.; Sun, Y.; Zhang, L. Features of the Complex of Food Additive Hesperidin to Hemoglobin. J. Photochem. Photobiol. B Biol. 2012, 106, 53–60. [Google Scholar] [CrossRef]
- Chakraborty, S.; Chaudhuri, S.; Pahari, B.; Taylor, J.; Sengupta, P.K.; Sengupta, B. A Critical Study on the Interactions of Hesperitin with Human Hemoglobin: Fluorescence Spectroscopic and Molecular Modeling Approach. J. Lumin. 2012, 132, 1522–1528. [Google Scholar] [CrossRef] [Green Version]
- Maity, S.; Chakraborty, S.; Chakraborti, A.S. Critical Insight into the Interaction of Naringenin with Human Haemoglobin: A Combined Spectroscopic and Computational Modeling Approaches. J. Mol. Struct. 2017, 1129, 256–262. [Google Scholar] [CrossRef]
- Yang, Z.; Cheng, X.; Li, X. Investigation of the Interaction between Human Hemoglobin and Five Antioxidants by Fluorescence Spectroscopy and Molecular Modeling. J. Iran. Chem. Soc. 2018, 15, 245–257. [Google Scholar] [CrossRef]
- Grigoryan, K.R.; Sargsyan, L.S. Denaturation of Hemoglobin in the Presence of Tannic Acid. Proc. Yerevan State Univ. 2014, 48, 23–27. [Google Scholar] [CrossRef]
- Sargsyan, L.C.; Grigoryan, K.R.; Markarian, S.A.; Burkholz, T.; Jacob, C. Fluorescence Characterization of Human Hemoglobin Binding with Tannic Acid. Chem. J. Armen. 2014, 67, 181–187. [Google Scholar]
- Basu, A.; Kumar, G.S. Interaction of the Dietary Pigment Curcumin with Hemoglobin: Energetics of the Complexation. Food Funct. 2014, 5, 1949–1955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thyagarajan, A.; Sahu, R.P. Potential Contributions of Antioxidants to Cancer Therapy: Immunomodulation and Radiosensitization. Integr. Cancer Ther. 2018, 17, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Wieland, L.S.; Moffet, I.; Shade, S.; Emadi, A.; Knott, C.; Gorman, E.F.; D’Adamo, C. Risks and Benefits of Antioxidant Dietary Supplement Use during Cancer Treatment: Protocol for a Scoping Review. BMJ Open 2021, 11, e047200. [Google Scholar] [CrossRef]
- Ambrosone, C.B.; Zirpoli, G.R.; Hutson, A.D.; McCann, W.E.; McCann, S.E.; Barlow, W.E.; Kelly, K.M.; Cannioto, R.; Sucheston-Campbell, L.E.; Hershman, D.L.; et al. Dietary Supplement Use During Chemotherapy and Survival Outcomes of Patients With Breast Cancer Enrolled in a Cooperative Group Clinical Trial (SWOG S0221). J. Clin. Oncol. 2020, 38, 804–814. [Google Scholar] [CrossRef]
- Reddy, P.H. Amyloid Precursor Protein-Mediated Free Radicals and Oxidative Damage: Implications for the Development and Progression of Alzheimer’s Disease. J. Neurochem. 2006, 96, 1–13. [Google Scholar] [CrossRef]
- Bajaj, S.; Khan, A. Antioxidants and Diabetes. Indian J. Endocrinol. Metab. 2012, 16, 267. [Google Scholar] [CrossRef]
- Wang, X.; Qi, Y.; Zheng, H. Dietary Polyphenol, Gut Microbiota, and Health Benefits. Antioxidants 2022, 11, 1212. [Google Scholar] [CrossRef] [PubMed]
- Riaz Rajoka, M.S.; Thirumdas, R.; Mehwish, H.M.; Umair, M.; Khurshid, M.; Hayat, H.F.; Phimolsiripol, Y.; Pallarés, N.; Martí-Quijal, F.J.; Barba, F.J. Role of Food Antioxidants in Modulating Gut Microbial Communities: Novel Understandings in Intestinal Oxidative Stress Damage and Their Impact on Host Health. Antioxidants 2021, 10, 1563. [Google Scholar] [CrossRef]
- Gammoh, S.; Alhamad, M.N.; Rababah, T.; Ereifej, K.; Almajwal, A. Characterization of phenolic compounds extracted from wheat protein fractions using high-performance liquid chromatography/liquid chromatography mass spectrometry in relation to an-ti-allergenic, anti-oxidant, anti-hypertension, and anti-diabetic properties. Int. J. Food Prop. 2017, 20, 2383–2395. [Google Scholar] [CrossRef] [Green Version]
- Nagy, K.; Courtet-Compondu, M.-C.; Williamson, G.; Rezzi, S.; Kussmann, M.; Rytz, A. Non-covalent binding of proteins to polyphenols correlates with their amino acid sequence. Food Chem. 2012, 132, 1333–1339. [Google Scholar] [CrossRef] [PubMed]
- Mirzaee, F.; Hosseinzadeh, L.; Ashrafi-Kooshk, M.R.; Esmaeili, S.; Ghobad, S.; Farzaei, M.H.; Zad-Bari, M.R.; Khodarahmi, R. Diverse Effects of Different “Protein-Based” Vehicles on the Stability and Bioavailability of Curcumin: Spectroscopic Evaluation of the Antioxidant Activity and Cytotoxicity In Vitro. Protein Pept. Lett. 2019, 26, 132–147. [Google Scholar] [CrossRef]
- Ponomarenko, E.A.; Poverennaya, E.V.; Ilgisonis, E.V.; Pyatnitskiy, M.A.; Kopylov, A.T.; Zgoda, V.G.; Lisitsa, A.V.; Archakov, A.I. The Size of the Human Proteome: The Width and Depth. Int. J. Anal. Chem. 2016, 2016, 7436849. [Google Scholar] [CrossRef] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nedić, O.; Penezić, A.; Minić, S.; Radomirović, M.; Nikolić, M.; Ćirković Veličković, T.; Gligorijević, N. Food Antioxidants and Their Interaction with Human Proteins. Antioxidants 2023, 12, 815. https://doi.org/10.3390/antiox12040815
Nedić O, Penezić A, Minić S, Radomirović M, Nikolić M, Ćirković Veličković T, Gligorijević N. Food Antioxidants and Their Interaction with Human Proteins. Antioxidants. 2023; 12(4):815. https://doi.org/10.3390/antiox12040815
Chicago/Turabian StyleNedić, Olgica, Ana Penezić, Simeon Minić, Mirjana Radomirović, Milan Nikolić, Tanja Ćirković Veličković, and Nikola Gligorijević. 2023. "Food Antioxidants and Their Interaction with Human Proteins" Antioxidants 12, no. 4: 815. https://doi.org/10.3390/antiox12040815