

Loss of the Kidney Urate Transporter, Urat1, Leads to Disrupted Redox Homeostasis in Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Wildtype and RST Knockout Mice
2.2. Transcriptomic Profiling
2.3. Metabolomic Profiling
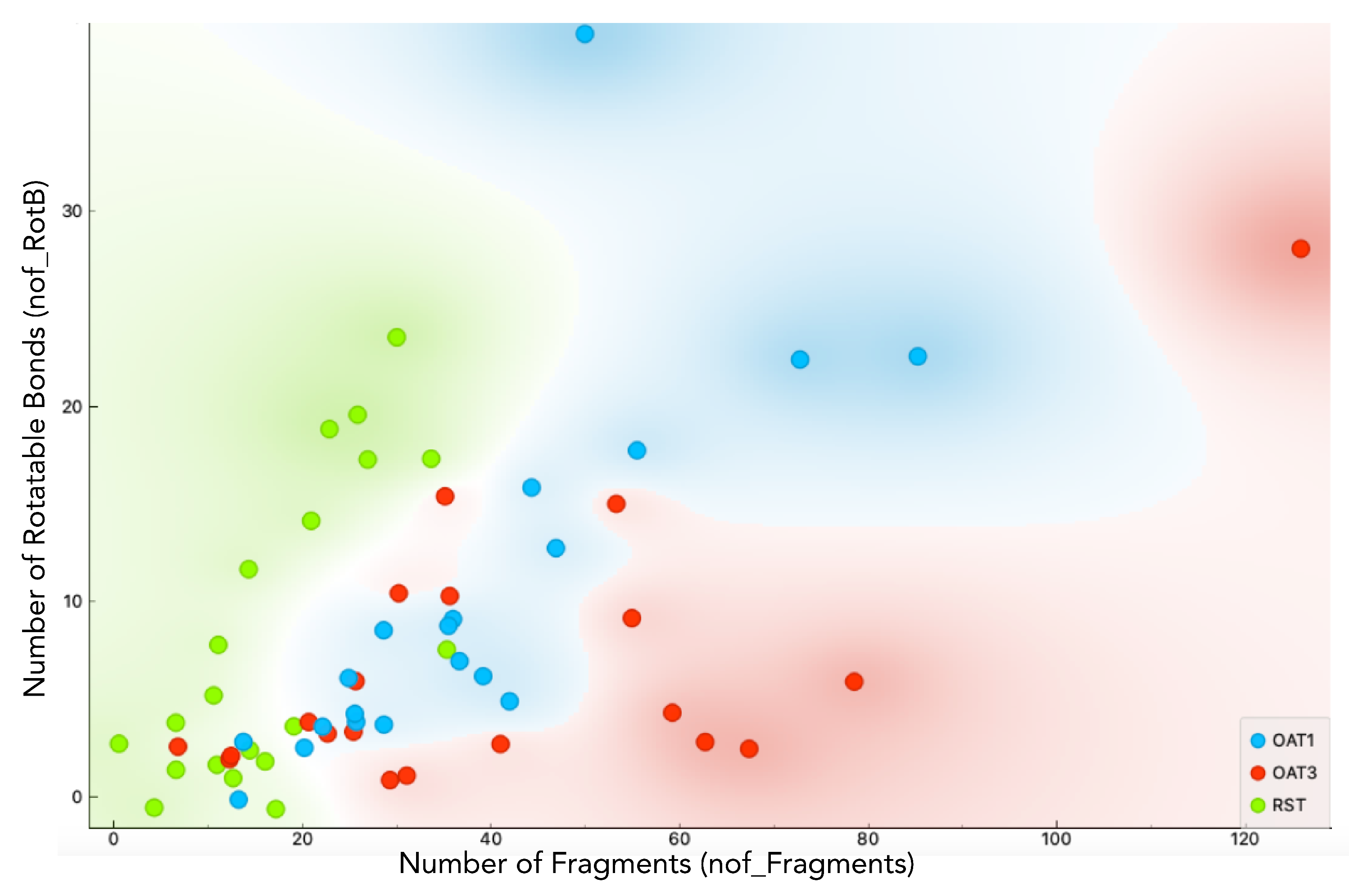
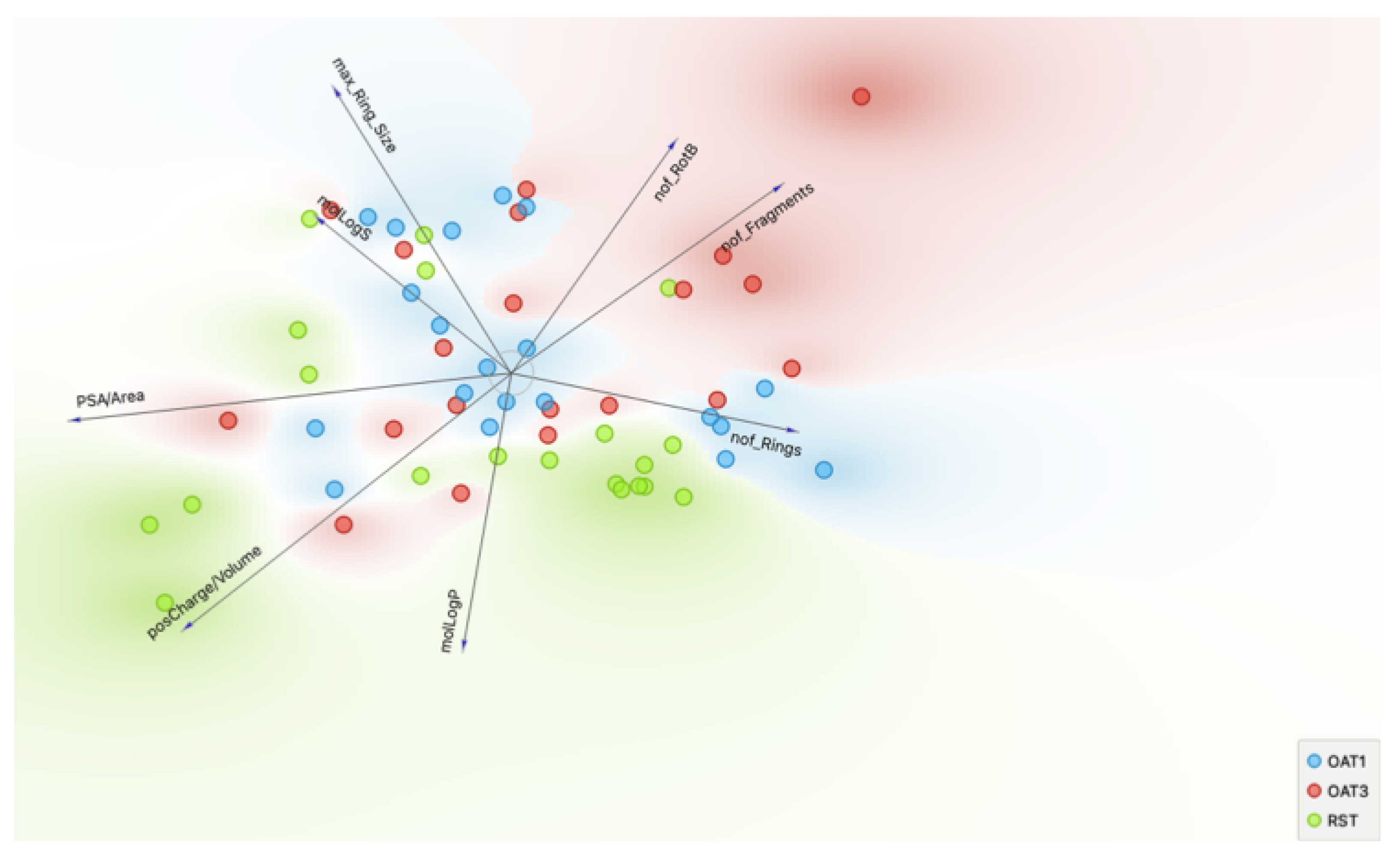
2.4. Chemoinformatics and Machine Learning
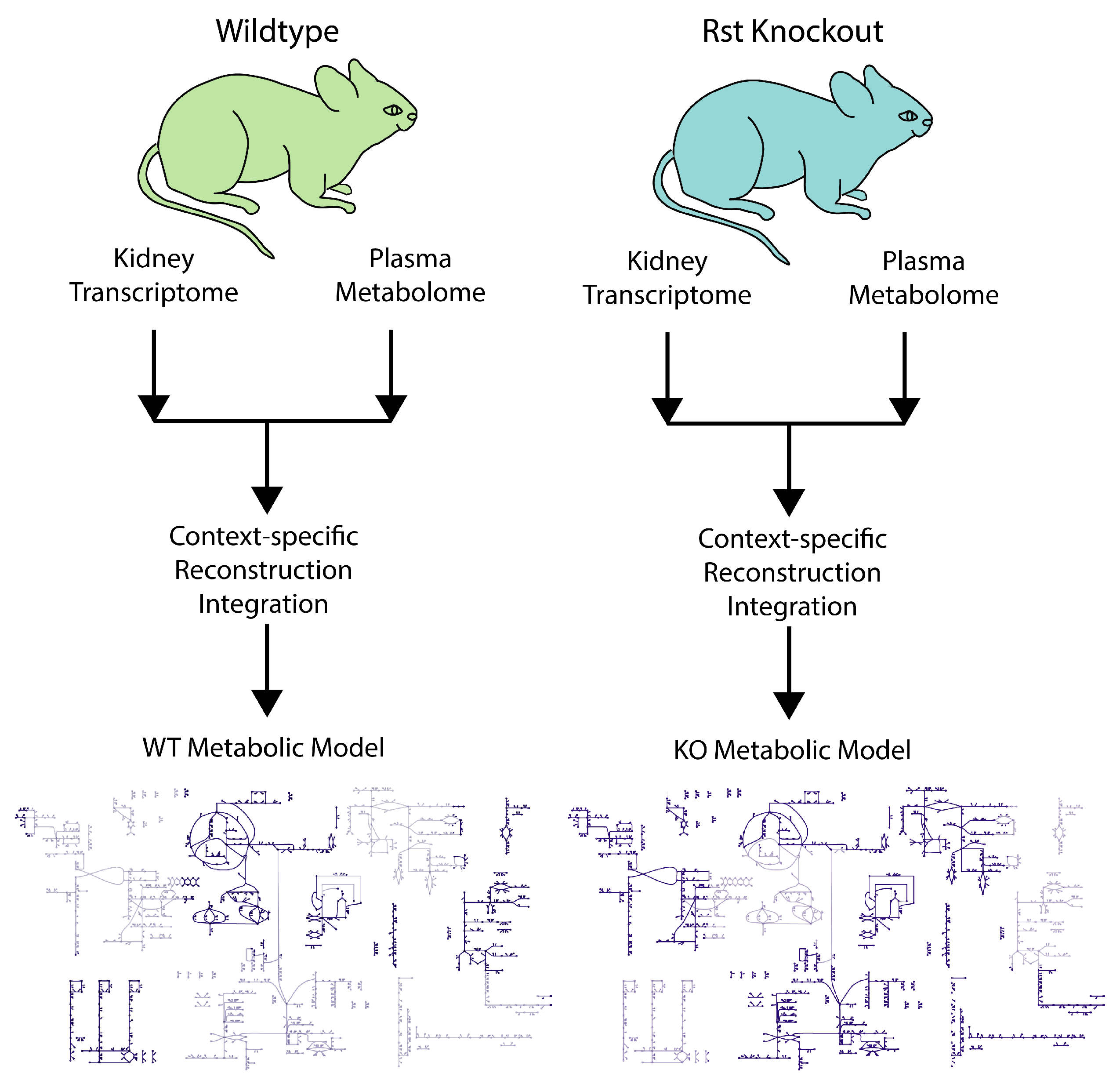
2.5. Modeling and Analysis
- ▸
- If KO/WT > 1 and the metabolite could only be secreted, then the metabolite exchange was constrained with a non-zero lower bound (10% of maximum secretion)
- ▸
- If KO/WT > 1 and the metabolite could only be taken up, then the metabolite exchange was constrained with a lower bound that was greater than the minimum
- ▸
- If KO/WT < 1 and the metabolite could only be secreted, then the metabolite exchange was constrained with an upper bound that was less than the maximum
- ▸
- If KO/WT < 1 and the metabolite could only be taken up, then the metabolite exchange was constrained with upper bound that was less than zero (90% of maximum uptake)
3. Results
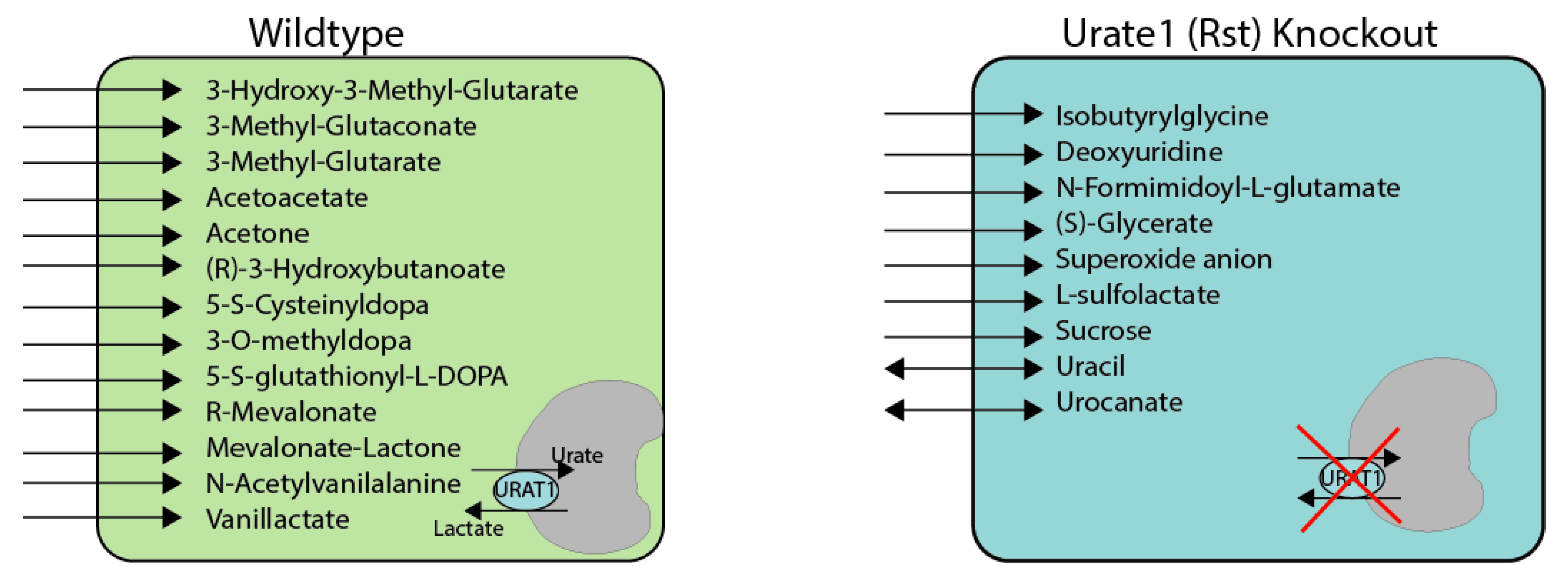
3.1. Metabolite Alterations Resulting from Urat1 Deletion
3.2. Chemoinformatics and Machine Learning Analysis of Metabolites Dependent Upon Urat1 Function In Vivo
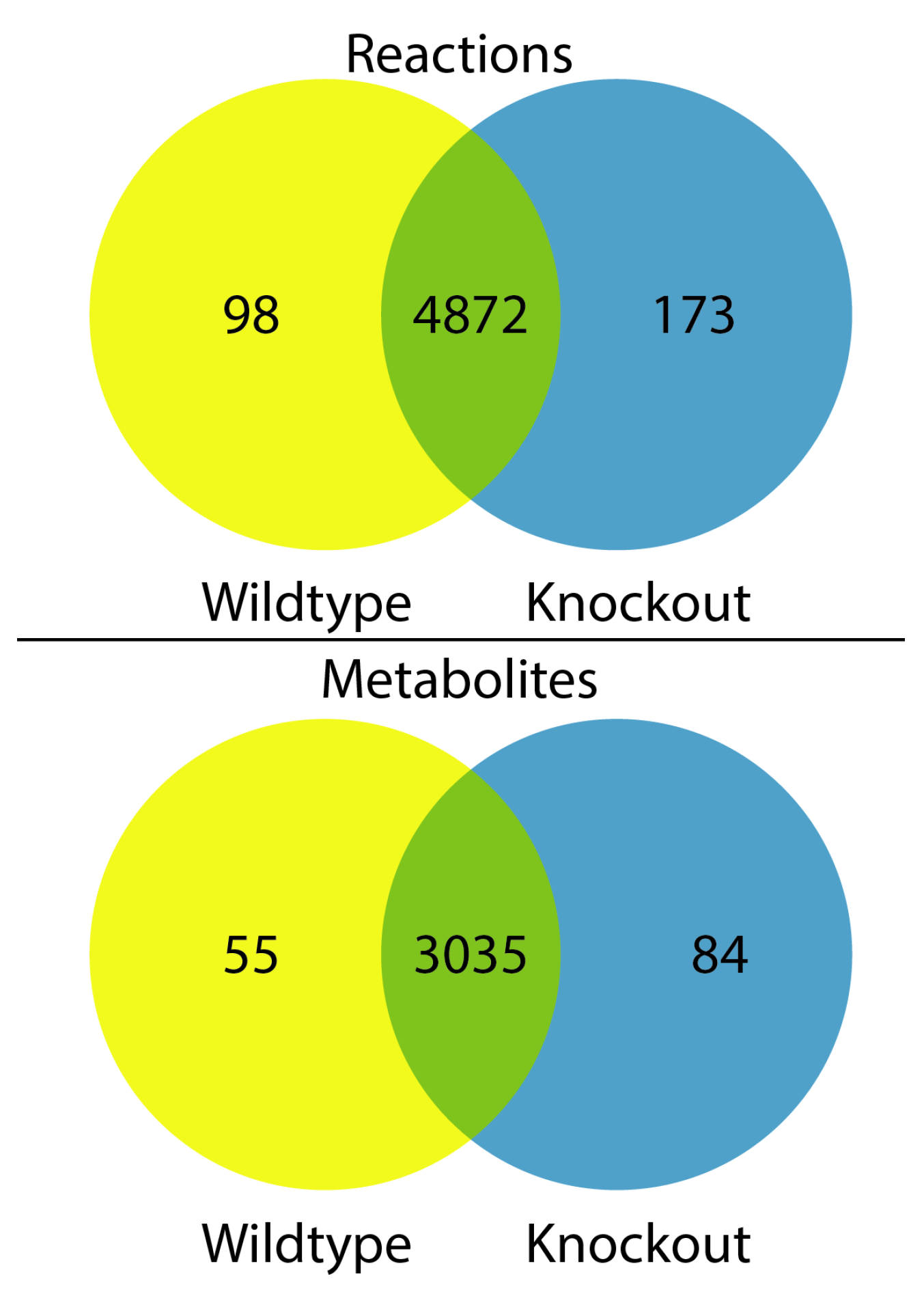
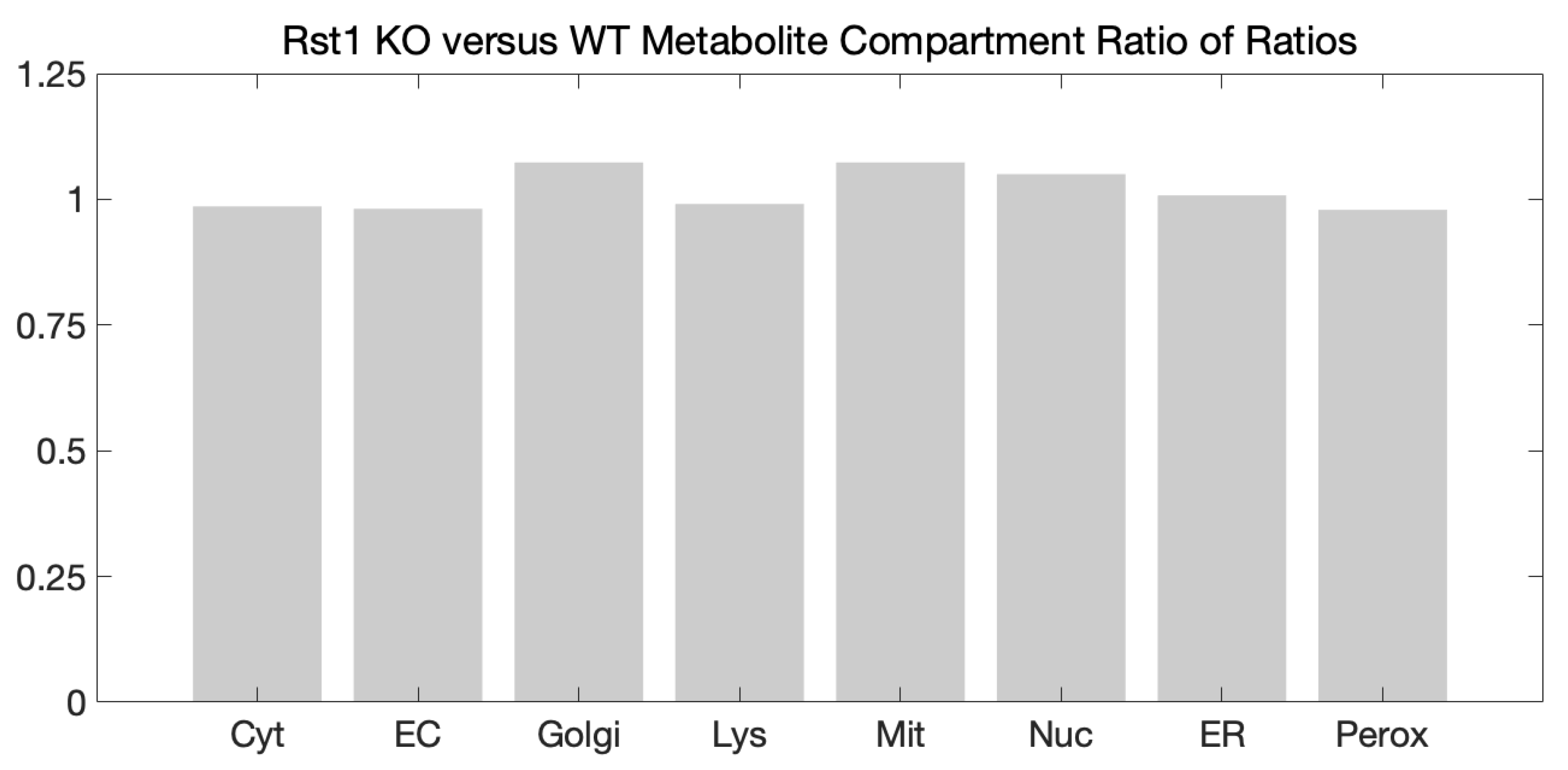
3.3. Metabolic Network Structural and Functional Differences of Wildtype and Urat1 Knockout Mice
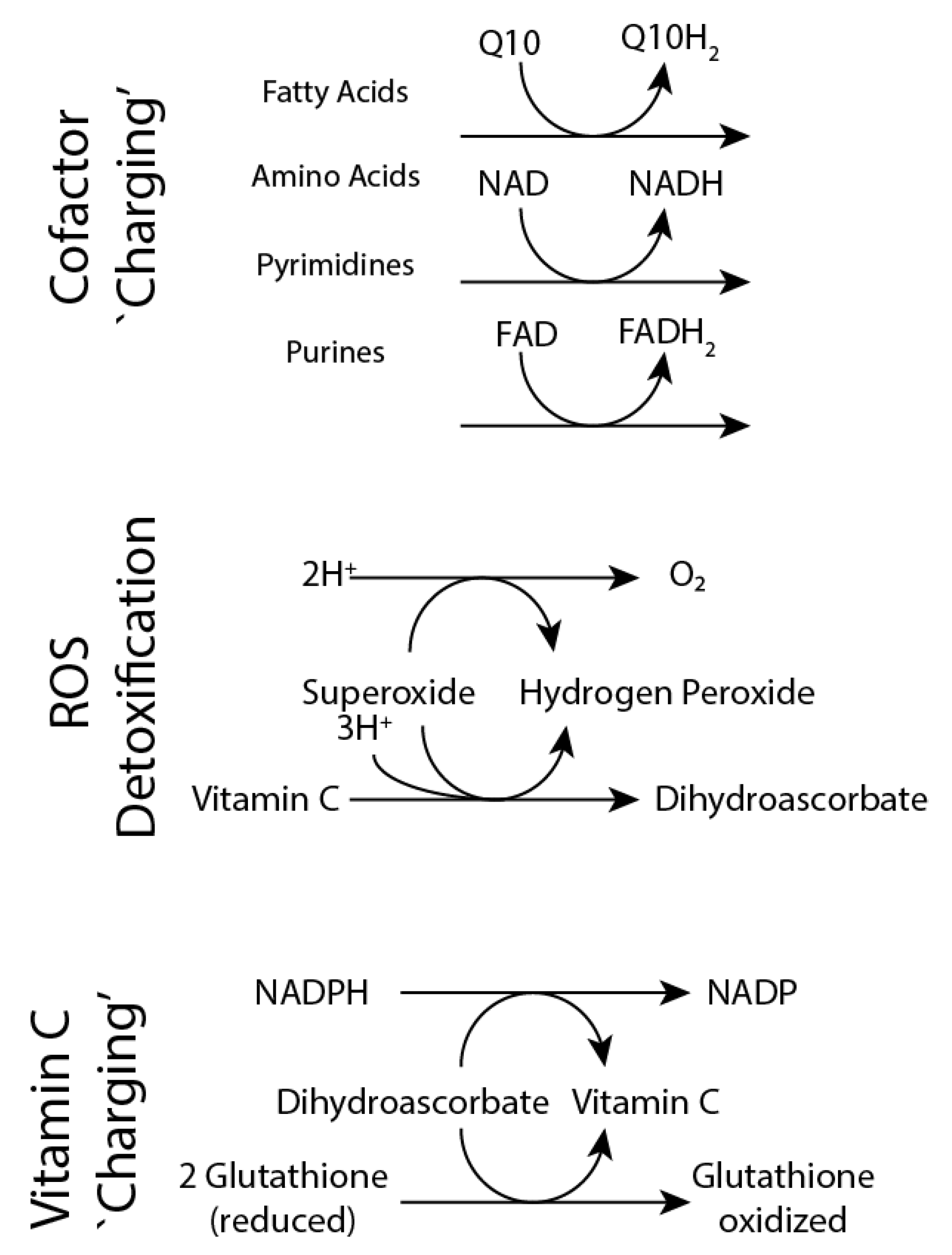
3.4. Altered Tissue and Systemic Redox State in the Urat1KO Mouse
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ANOVA | Two way analysis of variance |
| COBRA | Constraint-based Reconstruction Analysis |
| CKD | Chronic Kidney Disease |
| FBA | Flux Balance Analysis |
| FVA | Flux Variability Analysis |
| GEM | Genome-scale model |
| GIMME | Gene Inactivity Moderated by Metabolism and Expression |
| IACUC | Institutional Animal Care and Use Committee |
| KO | knockout |
| OAT1 | Organic Anion Transporter 1 |
| OAT3 | Organic Anion Transporter 3 |
| ROS | reactive oxygen species |
| RSST | Remote Sensing and Signaling Theory |
| WT | wildtype |
| URAT1 | Urate transporter 1 (known as RST in mice) |
References
- Nigam, S.K.; Bhatnagar, V. The systems biology of uric acid transporters: The role of remote sensing and signaling. Curr. Opin. Nephrol. Hypertens. 2018, 27, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, V.; Richard, E.L.; Wu, W.; Nievergelt, C.M.; Lipkowitz, M.S.; Jeff, J.; Maihofer, A.X.; Nigam, S.K. Analysis of ABCG2 and other urate transporters in uric acid homeostasis in chronic kidney disease: Potential role of remote sensing and signaling. Clin. Kidney J. 2016, 9, 444–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komori, H.; Yamada, K.; Tamai, I. Hyperuricemia enhances intracellular urate accumulation via down-regulation of cell-surface BCRP/ABCG2 expression in vascular endothelial cells. Biochim. Biophys. Acta Biomembr. 2018, 1860, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.Y.; Nigam, S.K. Toward a systems level understanding of organic anion and other multispecific drug transporters: A remote sensing and signaling hypothesis. Mol. Pharmacol. 2009, 76, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Nigam, S.K.; Granados, J.C. OAT, OATP, and MRP Drug Transporters and the Remote Sensing and Signaling Theory. Annu. Rev. Pharmacol. Toxicol. 2022, 63, 637–660. [Google Scholar] [CrossRef]
- Nigam, S.K.; Granados, J.C. A Biological Basis for Pharmacokinetics: The Remote Sensing and Signaling Theory. Clin. Pharmacol. Ther. 2022, 112, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Nigam, S.K. What do drug transporters really do? Nat. Rev. Drug Discov. 2015, 14, 29–44. [Google Scholar] [CrossRef] [Green Version]
- Pavelcova, K.; Bohata, J.; Pavlikova, M.; Bubenikova, E.; Pavelka, K.; Stiburkova, B. Evaluation of the Influence of Genetic Variants of SLC2A9 (GLUT9) and SLC22A12 (URAT1) on the Development of Hyperuricemia and Gout. J. Clin. Med. 2020, 9, 2510. [Google Scholar] [CrossRef]
- Cho, S.K.; Kim, S.; Chung, J.Y.; Jee, S.H. Discovery of URAT1 SNPs and association between serum uric acid levels and URAT1. BMJ Open 2015, 5, e009360. [Google Scholar] [CrossRef] [Green Version]
- Eraly, S.A.; Vallon, V.; Rieg, T.; Gangoiti, J.A.; Wikoff, W.R.; Siuzdak, G.; Barshop, B.A.; Nigam, S.K. Multiple organic anion transporters contribute to net renal excretion of uric acid. Physiol. Genom. 2008, 33, 180–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, S.Y.; Jamshidi, N.; Mo, M.L.; Wu, W.; Eraly, S.A.; Dnyanmote, A.; Bush, K.T.; Gallegos, T.F.; Sweet, D.H.; Palsson, B.O.; et al. Linkage of organic anion transporter-1 to metabolic pathways through integrated “omics”-driven network and functional analysis. J. Biol. Chem. 2011, 286, 31522–31531. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.C.; Jamshidi, N.; Chen, Y.; Eraly, S.A.; Cho, S.Y.; Bhatnagar, V.; Wu, W.; Bush, K.T.; Abagyan, R.; Palsson, B.O.; et al. An Organic Anion Transporter 1 (OAT1)-centered Metabolic Network. J. Biol. Chem. 2016, 291, 19474–19486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, S.Y.; Eraly, S.A.; Tsigelny, I.; Nigam, S.K. Interaction of organic cations with organic anion transporters. J. Biol. Chem. 2009, 284, 31422–31430. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Azad, P.; Engelhart, D.C.; Haddad, G.G.; Nigam, S.K. SLC22 Transporters in the Fly Renal System Regulate Response to Oxidative Stress In Vivo. Int. J. Mol. Sci. 2021, 22, 13407. [Google Scholar] [CrossRef]
- Whisenant, T.C.; Nigam, S.K. Organic Anion Transporters (OAT) and Other SLC22 Transporters in Progression of Renal Cell Carcinoma. Cancers 2022, 14, 4772. [Google Scholar] [CrossRef]
- Xu, J.; Liu, Y.; Liu, J.; Shou, Y.; Xiong, Z.; Xiong, H.; Xu, T.; Wang, Q.; Liu, D.; Liang, H.; et al. Low Expression Levels of SLC22A12 Indicates a Poor Prognosis and Progresses Clear Cell Renal Cell Carcinoma. Front. Oncol. 2021, 11, 659208. [Google Scholar] [CrossRef] [PubMed]
- Dufour, I.; Werion, A.; Belkhir, L.; Wisniewska, A.; Perrot, M.; De Greef, J.; Schmit, G.; Yombi, J.C.; Wittebole, X.; Laterre, P.F.; et al. , Serum uric acid, disease severity and outcomes in COVID-19. Crit. Care 2021, 25, 212–11. [Google Scholar] [CrossRef]
- Hosoyamada. M.; Ichida. K.; Enomoto, A.; Hosoya, T.; Endou, H. Function and localization of urate transporter 1 in mouse kidney. J. Am. Soc. Nephrol. 2004, 15, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigam, A.K.; Li, J.G.; Lall, K.; Shi, D.; Bush, K.T.; Bhatnagar, V.; Abagyan, R.; Nigam, S.K. Unique metabolite preferences of the drug transporters OAT1 and OAT3 analyzed by machine learning. J. Biol. Chem. 2020, 295, 1829–1842. [Google Scholar] [CrossRef]
- Bush, K.T.; Wu, W.; Lun, C.; Nigam, S.K. The drug transporter OAT3 (SLC22A8) and endogenous metabolite communication via the gut-liver-kidney axis. J. Biol. Chem. 2017, 292, 15789–15803. [Google Scholar] [CrossRef] [Green Version]
- Nigam, A.K.; Ojha, A.A.; Li, J.G.; Shi, D.; Bhatnagar, V.; Nigam, K.B.; Abagyan, R.; Nigam, S.K. Molecular Properties of Drugs Handled by Kidney OATs and Liver OATPs Revealed by Chemoinformatics and Machine Learning: Implications for Kidney and Liver Disease. Pharmaceutics 2021, 13, 1720. [Google Scholar] [CrossRef] [PubMed]
- Demsar, J.; C. T.; Erjavec, A.; Gorup, C.; Hocevar, T.; Milutinovic, M.; Mozina, M.; Polajnar, M.; Toplak, M.; Staric, A.; et al. Orange: Data mining toolbox in python. J. Mach. Learn. Res. 2013, 14, 2349–2353. [Google Scholar]
- Brunk, E.; Sahoo, S.; Zielinski, D.C.; Altunkaya, A.; Drager, A.; Mih, N.; Gatto, F.; Nilsson, A.; Preciat Gonzalez, G.A.; Aurich, M.K. ; Recon3D enables a three-dimensional view of gene variation in human metabolism. Nat. Biotechnol. 2018, 36, 272–281. [Google Scholar] [CrossRef]
- Duarte, N.C.; Becker, S.A.; Jamshidi, N.; Thiele, I.; Mo, M.L.; Vo, T.D.; Srivas, R.; Palsson, B.O. Global reconstruction of the human metabolic network based on genomic and bibliomic data. Proc. Natl. Acad. Sci. USA 2007, 104, 1777–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, S.A.; Palsson, B.O. Context-specific metabolic networks are consistent with experiments. PLoS Comput. Biol. 2008, 4, e1000082. [Google Scholar] [CrossRef] [Green Version]
- Schellenberger, J.; Que, R.; Fleming, R.M.; Thiele, I.; Orth, J.D.; Feist, A.M.; Zielinski, D.C.; Bordbar, A.; Lewis, N.E.; Rahmanian, S.; et al. Quantitative prediction of cellular metabolism with constraint-based models: The COBRA Toolbox v2.0. Nat. Protoc. 2011, 6, 1290–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahadevan, R.; Schilling, C.H. The effects of alternate optimal solutions in constraint-based genome-scale metabolic models. Metab. Eng. 2003, 5, 264–276. [Google Scholar] [CrossRef]
- Bordbar, A.; Lewis, N.E.; Schellenberger, J.; Palsson, B.O.; Jamshidi, N. Insight into human alveolar macrophage and M. tuberculosis interactions via metabolic reconstructions. Mol. Syst. Biol. 2010, 6, 422. [Google Scholar] [CrossRef] [PubMed]
- Jerby, L.; Shlomi, T.; Ruppin, E. Computational reconstruction of tissue-specific metabolic models: Application to human liver metabolism. Mol. Syst. Biol. 2010, 6, 401. [Google Scholar] [CrossRef]
- André, Schultz. ; Amina, A.Q. Reconstruction of Tissue-Specific Metabolic Networks Using CORDA. PLoS Comput. Biol. 2016, 12, 12. [Google Scholar]
- Ebrahim, A.; Lerman, J.A.; Palsson, B.O.; Hyduke, D.R. COBRApy: COnstraints-Based Reconstruction and Analysis for Python. BMC Syst. Biol. 2013, 7, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, Z.A.; Drager, A.; Ebrahim, A.; Sonnenschein, N.; Lewis, N.E.; Palsson, B.O. Escher: A Web Application for Building, Sharing, and Embedding Data-Rich Visualizations of Biological Pathways. PLoS Comput. Biol. 2015, 11, e1004321. [Google Scholar] [CrossRef] [Green Version]
- Eraly, S.A.; Liu, H.C.; Jamshidi, N.; Nigam, S.K. Transcriptome-based reconstructions from the murine knockout suggest involvement of the urate transporter, URAT1 (slc22a12), in novel metabolic pathways. Biochem. Biophys. Rep. 2015, 3, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordbar, A.; Palsson, B.O. Using the reconstructed genome-scale human metabolic network to study physiology and pathology. J. Intern. Med. 2012, 271, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.L.; Patel, T.R.; Chen, K.H.; Joyce, A.R.; Applebee, M.K.; Herring, C.D.; Bui, O.T.; Knight, E.M.; Fong, S.S.; Palsson, B.O. Systems approach to refining genome annotation. Proc. Natl. Acad. Sci. USA 2006, 103, 17480–17484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, C.; Kim, G.B.; Kim, W.J.; Kim, H.U.; Lee, S.Y. Current status and applications of genome-scale metabolic models. Genome Biol. 2019, 20, 121. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.Y.; Sohn, S.B.; Kim, Y.B.; Kim, W.J.; Lee, S.Y. Recent advances in reconstruction and applications of genome-scale metabolic models. Curr. Opin. Biotechnol. 2012, 23, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.L.; Famili, I.; Thiele, I.; Palsson, B.O. Towards multidimensional genome annotation. Nat. Rev. Genet. 2006, 7, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Lewis, N.E.; Nagarajan, H.; Palsson, B.O. Constraining the metabolic genotype-phenotype relationship using a phylogeny of in silico methods. Nat. Rev. Microbiol. 2012, 10, 291–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blais, E.M.; Rawls, K.D.; Dougherty, B.V.; Li, Z.I.; Kolling, G.L.; Ye, P.; Wallqvist, A.; Papin, J.A. Reconciled rat and human metabolic networks for comparative toxicogenomics and biomarker predictions. Nat. Commun. 2017, 8, 14250. [Google Scholar] [CrossRef] [Green Version]
- Jerby, L.; Ruppin, E. Predicting drug targets and biomarkers of cancer via genome-scale metabolic modeling. Clin. Cancer Res. 2012, 18, 5572–5584. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Jamshidi, N.; Eraly, S.A.; Liu, H.C.; Bush, K.T.; Palsson, B.O.; Nigam, S.K. Multispecific drug transporter Slc22a8 (Oat3) regulates multiple metabolic and signaling pathways. Drug Metab. Dispos. Biol. Fate Chem. 2013, 41, 1825–1834. [Google Scholar] [CrossRef] [Green Version]
- Jamshidi, N.; Nigam, S.K. Drug transporters OAT1 and OAT3 have specific effects on multiple organs and gut microbiome as revealed by contextualized metabolic network reconstructions. Sci. Rep. 2022, 12, 18308. [Google Scholar] [CrossRef] [PubMed]
- Tasic, V.; Hynes, A.M.; Kitamura, K.; Cheong, H.I.; Lozanovski, V.J.; Gucev, Z.; Jutabha, P.; Anzai, N.; Sayer, J.A. Clinical and functional characterization of URAT1 variants. PLoS ONE 2011, 6, e28641. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yan, Z.; Tian, J.; Zhang, X.; Han, H.; Ye, F. Urate Transporter URAT1 in Hyperuricemia: New Insights from Hyperuricemic Models. Ann. Clin. Lab. Sci. 2019, 49, 756–762. [Google Scholar] [PubMed]
- Arakawa, H.; Amezawa, N.; Katsuyama, T.; Nakanishi, T.; Tamai, I. Uric acid analogue as a possible xenobiotic marker of uric acid transporter Urat1 in rats. Drug Metab. Pharmacokinet. 2019, 34, 155–158. [Google Scholar] [CrossRef]
- Koepsell, H.; Endou, H. The SLC22 drug transporter family. Pflugers Arch. 2004, 447, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Nieto, C.E.; You, G.; Bush, K.T.; Barros, E.J.; Beier, D.R.; Nigam, S.K. Molecular cloning and characterization of NKT, a gene product related to the organic cation transporter family that is almost exclusively expressed in the kidney. J. Biol. Chem. 1997, 272, 6471–6478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelhart, D.C.; Granados, J.C.; Shi, D.; Saier, M.H., Jr.; Baker, M.E.; Abagyan, R.; Nigam, S.K. Systems Biology Analysis Reveals Eight SLC22 Transporter Subgroups, Including OATs, OCTs, and OCTNs. Int. J. Mol. Sci. 2020, 21, 1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Bush, K.T.; Nigam, S.K. Key Role for the Organic Anion Transporters, OAT1 and OAT3, in the in vivo Handling of Uremic Toxins and Solutes. Sci. Rep. 2017, 2017 7, 4939. [Google Scholar] [CrossRef] [Green Version]
- Rawls, K.D.; Blais, E.M.; Dougherty, B.V.; Vinnakota, K.C.; Pannala, V.R.; Wallqvist, A.; Kolling, G.L.; Papin, J.A. Genome-Scale Characterization of Toxicity-Induced Metabolic Alterations in Primary Hepatocytes. Toxicol. Sci. 2019, 172, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Robinson, J.L.; Kocabas, P.; Gustafsson, J.; Anton, M.; Cholley, P.E.; Huang, S.; Gobom, J.; Svensson, T.; Uhlen, M.; et al. Genome-scale metabolic network reconstruction of model animals as a platform for translational research. Proc. Natl. Acad. Sci. USA 2021, 118, e2102344118. [Google Scholar] [CrossRef] [PubMed]
- Mardinoglu, A.; Agren, R.; Kampf, C.; Asplund, A.; Uhlen, M.; Nielsen, J. Genome-scale metabolic modelling of hepatocytes reveals serine deficiency in patients with non-alcoholic fatty liver disease. Nat. Commun. 2014, 5, 3083. [Google Scholar] [CrossRef] [Green Version]
- Frezza, C.; Zheng, L.; Folger, O.; Rajagopalan, K.N.; MacKenzie, E.D.; Jerby, L.; Micaroni, M.; Chaneton, B.; Adam, J.; Hedley, A.; et al. Haem oxygenase is synthetically lethal with the tumour suppressor fumarate hydratase. Nature 2011, 477, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Jamshidi, N.; Miller, F.J.; Mandel, J.; Evans, T.; Kuo, M.D. Individualized therapy of HHT driven by network analysis of metabolomic profiles. BMC Syst. Biol. 2011, 5, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enomoto, A.; Endou, H. Roles of organic anion transporters (OATs) and a urate transporter (URAT1) in the pathophysiology of human disease. Clin. Exp. Nephrol. 2005, 9, 195–205. [Google Scholar] [CrossRef]
- Granados, J.C.; Bhatnagar, V.; Nigam, S.K. Blockade of Organic Anion Transport in Humans After Treatment With the Drug Probenecid Leads to Major Metabolic Alterations in Plasma and Urine. Clin. Pharmacol. Ther. 2022, 112, 653–664. [Google Scholar] [CrossRef]
- Shin, H.J.; Takeda, M.; Enomoto, A.; Fujimura, M.; Miyazaki, H.; Anzai, N.; Endou, H. Interactions of urate transporter URAT1 in human kidney with uricosuric drugs. Nephrology 2011, 16, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, A.; Kimura, H.; Chairoungdua, A.; Shigeta, Y.; Jutabha, P.; Cha, S.H.; Hosoyamada, M.; Takeda, M.; Sekine, T.; Igarashi, T.; et al. Molecular identification of a renal urate anion exchanger that regulates blood urate levels. Nature 2002, 417, 447–452. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reaction Abbreviation | Subsystem | Biochemical Reaction | Compartment |
|---|---|---|---|
| SPODM | ROS detoxification | 2 hc + 2 o2sc → o2c + h2o2c | Cytosol |
| SPODMe | ROS detoxification | 2 he + 2 o2se → h2o2e + o2e | Extracellular |
| SPODMm | ROS detoxification | 2 hm + 2 o2sm → o2m + h2o2m | Mitochondria |
| SPODMn | ROS detoxification | 2 hn + 2 o2sn → h2o2n + o2n | Nucleus |
| SPODMx | ROS detoxification | 2 hx + 2 o2sx → o2x + h2o2x | Peroxisome |
| ASCBOX1 | Vitamin C metabolism | 3 hc + ascb-Lc + 2 o2sc → 2 h2o2c + dhdascbc | Cytosol |
| DASCBR1 | Vitamin C metabolism | nadphc + dhdascbc → nadpc + ascb-Lc | Cytosol |
| DHAOXc | Vitamin C metabolism | dhdascbc + 2 gthrdc → hc + ascb-Lc + gthoxc | Cytosol |
| Reaction Abbreviation | Subsystem | Biochemical Reaction | Compartment |
|---|---|---|---|
| HMR_4782 | Arginine and proline metabolism | o2c + hc + 4hpro-LTc → 2 h2oc + 1p3h5cc | Cytosol |
| CITL | Citric acid cycle | citc → acc + oaac | Cytosol |
| GPDDA1 | Glycerophospholipid metabolism | h2oc + g3pcc → hc + cholc + glyc3pc | Cytosol |
| HMR_0853 | Glycosphingolipid metabolism | cmpacnac + gd1a-hsc → hc + cmpc + gt1a-hsc | Cytosol |
| RE2675C2 | Glycosphingolipid metabolism | o2c + hc + nadphc + dhcrm-hsc → 2 h2oc + nadpc + crm-hsc | Cytosol |
| PI3P5K | Inositol phosphate metabolism | atpc + pail3p-hsc → hc + adpc+ pail35p-hsc | Cytosol |
| PIK3n | Inositol phosphate metabolism | atpn + pail-hsn → hn + adpn + pail3p-hsn | Nucleus |
| PIK4n | Inositol phosphate metabolism | atpn + pail-hsn → hn+ adpn + pail4p-hsn | Nucleus |
| PIPLC | Inositol phosphate metabolism | h2oc + pail-hsc → hc + dag-hsc + mi1p-Dc | Cytosol |
| GLYOX | Pyruvate metabolism | h2oc + lgt-Sc → hc + gthrdc + lac-Dc | Cytosol |
| SMS | Sphingolipid metabolism | pchol-hsc + crm-hsc → dag-hsc+ sphmyln-hsc | Cytosol |
| RE3050R | Vitamin A metabolism | h2or + nadr + retinal-cis-13r → 2 hr + nadhr + retnr | Endoplasmic reticulum |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jamshidi, N.; Nigam, K.B.; Nigam, S.K. Loss of the Kidney Urate Transporter, Urat1, Leads to Disrupted Redox Homeostasis in Mice. Antioxidants 2023, 12, 780. https://doi.org/10.3390/antiox12030780
Jamshidi N, Nigam KB, Nigam SK. Loss of the Kidney Urate Transporter, Urat1, Leads to Disrupted Redox Homeostasis in Mice. Antioxidants. 2023; 12(3):780. https://doi.org/10.3390/antiox12030780
Chicago/Turabian StyleJamshidi, Neema, Kabir B. Nigam, and Sanjay K. Nigam. 2023. "Loss of the Kidney Urate Transporter, Urat1, Leads to Disrupted Redox Homeostasis in Mice" Antioxidants 12, no. 3: 780. https://doi.org/10.3390/antiox12030780