
Dietary Guanidine Acetic Acid Improves Ruminal Antioxidant Capacity and Alters Rumen Fermentation and Microflora in Rapid-Growing Lambs
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics Statement
2.2. Guanidinoacetic Acid Products
2.3. Experimental Animals, Diets, and Design
2.4. Sample Collection and Analyses
2.4.1. Rumen Fluid Sampling
2.4.2. Rumen Fermentation Index Measurement
2.4.3. Ruminal GAA, Creatine, Enzyme Activity, and Antioxidant Capacity Related to GAA Metabolism
2.4.4. Ruminal Microorganism DNA Extraction, PCR Amplification, and Sequencing
2.4.5. Processing of Sequencing Data
2.5. Statistical Analysis
3. Results
3.1. Rumen Fermentation
3.2. Ruminal GAA, Creatine, ATP, and Related Metabolic Enzymes
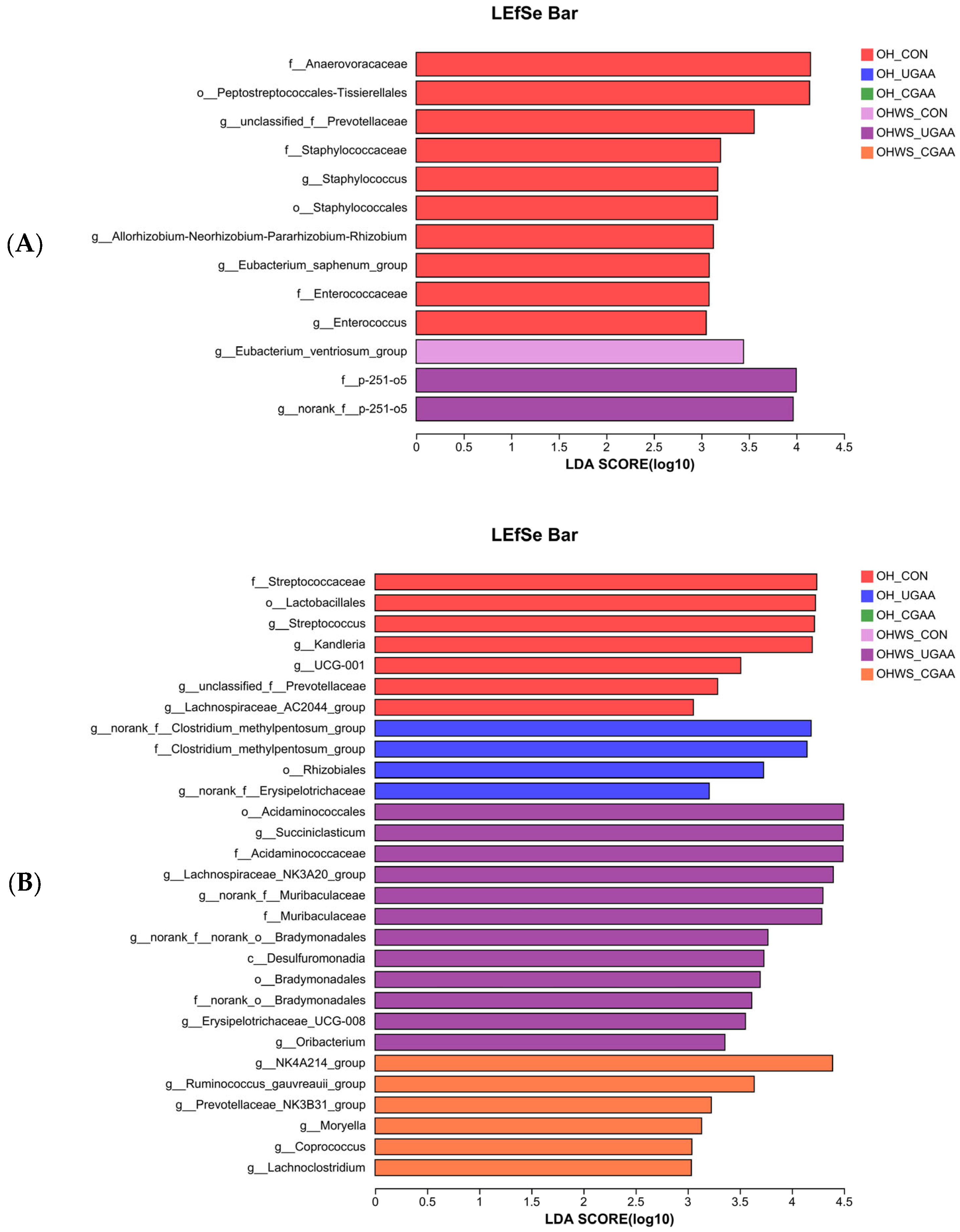
3.3. Ruminal Microbiota
3.4. Rumen Antioxidant Capacity
4. Discussion
4.1. Rumen Fermentation
4.2. Ruminal GAA, Creatine, ATP, and Related Metabolic Enzymes
4.3. Ruminal Microbiota
4.4. Rumen Antioxidant Capacity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
| Ingredients | Stage 1 | Stage 2 | ||
|---|---|---|---|---|
| OH | OHWS | OH | OHWS | |
| Wheat silage 1 | 0 | 90 | 0 | 70 |
| Oat hay 2 | 250 | 160 | 200 | 130 |
| Concentrate 3 | 750 | 750 | 800 | 800 |
| Nutrient level of TMR (g/kg, as Dry Matter) | ||||
| Organic matter | 893.3 | 901.8 | 905.6 | 910.5 |
| Crude protein | 191.9 | 193.7 | 184.6 | 174.5 |
| Ether extract | 36.6 | 35.1 | 38.0 | 37.5 |
| Neutral detergent fiber | 307.6 | 293.8 | 274.5 | 271.2 |
| Acid detergent fiber | 118.4 | 117.1 | 108.0 | 99.7 |
| Net energy for gain (MJ/kg) | 4.60 | 4.64 | 4.79 | 4.81 |
| Stages | Temperature | Time | Cycles |
|---|---|---|---|
| Initial denaturation | 95 °C | 3 min | 1 |
| Denaturation | 95 °C | 30 s | 27 |
| Annealing | 55 °C | 30 s | 1 |
| Extension | 72 °C | 45 s | 1 |
| Single extension | 72 °C | 10 min | 1 |
| Items | Volume |
|---|---|
| 5 × FastPfu Buffer | 4 μL |
| 2.5 mmol/L dNTPs | 2 μL |
| Forward primer (5 mmol/L) | 0.8 μL |
| Reverse primer (5 mmol/L) | 0.8 μL |
| FastPfu Polymerase | 0.4 μL |
| Template DNA | 10 ng |
| ddH2O | 12 μL |
| Total | 20 μL |
References
- Ostojic, S.M. Tackling guanidinoacetic acid for advanced cellular bioenergetics. Nutrition 2017, 34, 55–57. [Google Scholar] [CrossRef]
- Zhu, Z.; Gu, C.; Hu, S.; Li, B.; Zeng, X.; Yin, J. Dietary guanidinoacetic acid supplementation improved carcass characteristics, meat quality and muscle fibre traits in growing-finishing gilts. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1454–1461. [Google Scholar] [CrossRef]
- Majdeddin, M.; Braun, U.; Lemme, A.; Golian, A.; Kermanshahi, H.; Smet, D.S.; Michiels, J. Guanidinoacetic acid supplementation improves feed conversion in broilers subjected to heat stress associated with muscle creatine loading and arginine sparing. Poult. Sci. 2020, 99, 4442–4453. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, C.; Wu, Z.; Liu, Q.; Guo, G.; Huo, W.; Zhang, J.; Chen, L.; Zhang, Y.; Pei, C.; et al. Effects of guanidinoacetic acid supplementation on growth performance, nutrient digestion, rumen fermentation and blood metabolites in Angus bulls. Animal 2020, 14, 2535–2542. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, X.; Song, P.; Zhao, J.; Zhang, J.; Zhao, J. Skeletal muscle mass, meat quality and antioxidant status in growing lambs supplemented with guanidinoacetic acid. Meat Sci. 2022, 192, 108906. [Google Scholar] [CrossRef] [PubMed]
- Newbold, C.J.; Ramos-Morales, E. Review: Ruminal microbiome and microbial metabolome: Effects of diet and ruminant host. Animal 2020, 14, s78–s86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, M.; Webb, M.; Ghimire, S.; Blair, A.; Olson, K.; Fenske, G.J.; Fonder, A.T.; Christopher-Hennings, J.; Brake, D.; Scaria, J. Metagenomic characterization of the effect of feed additives on the gut microbiome and antibiotic resistome of feedlot cattle. Sci. Rep. 2017, 7, 12257. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Wu, H.; Liu, S.; Chai, S.; Meng, Q.; Zhou, Z. Dynamic alterations in yak rumen bacteria community and metabolome characteristics in response to feed type. Front. Microbiol. 2019, 10, 1116. [Google Scholar] [CrossRef] [Green Version]
- Speer, H.F.; Pearl, K.A.; Titgemeyer, E.C. Relative bioavailability of guanidinoacetic acid delivered ruminally or abomasally to cattle. J. Anim. Sci. 2020, 98, skaa282. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Juniper, D.T. Revisiting Oxidative Stress and the Use of Organic Selenium in Dairy Cow Nutrition. Animals 2019, 9, 462. [Google Scholar] [CrossRef] [Green Version]
- Bezerra, H.V.A.; Buarque, V.L.M.; Silva, L.S.B.; Leme, P.R.P.; Vidal, A.M.C.; Vaz, A.C.N.; Gallo, S.B.; Silva, S.L.; Leme, P.R. Effect of Castor and Cashew Nut Shell Oils, Selenium and Vitamin E as Antioxidants on the Health and Meat Stability of Lambs Fed a High-Concentrate Diet. Antioxidants 2020, 9, 1298. [Google Scholar] [CrossRef] [PubMed]
- Izuddin, W.I.; Humam, A.M.; Loh, T.C.; Foo, H.L.; Samsudin, A.A. Dietary Postbiotic Lactobacillus plantarum Improves Serum and Ruminal Antioxidant Activity and Upregulates Hepatic Antioxidant Enzymes and Ruminal Barrier Function in Post-Weaning Lambs. Antioxidants 2020, 9, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mion, B.; Ogilvie, L.; Van Winters, B.; Spricigo, J.F.W.; Anan, S.; Duplessis, M.; McBride, B.W.; LeBlanc, S.J.; Steele, M.A.; Ribeiro, E.S. Effects of replacing inorganic salts of trace minerals with organic trace minerals in the pre- and postpartum diets on mineral status, antioxidant biomarkers, and health of dairy cows. J. Anim. Sci. 2023, 3, skad041. [Google Scholar] [CrossRef] [PubMed]
- Gázquez, A.; Sánchez-Campillo, M.; Arnao, M.B.; Barranco, A.; Rueda, R.; Jensen, S.K.; Chan, J.P.; Kuchan, M.J.; Larqué, E. Natural vitamin E supplementation during pregnancy in rats increases RRR-α-tocopherol stereoisomer proportion and enhances fetal antioxidant capacity, compared to synthetic vitamin E administration. Ann. Nutr. Metab. 2023. [Google Scholar] [CrossRef]
- Hiramatsu, M. A role for guanidino compounds in the brain. Mol. Cell. Biochem. 2003, 244, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Arazi, H.; Eghbali, E.; Suzuki, K. Creatine Supplementation, Physical Exercise and Oxidative Stress Markers: A Review of the Mechanisms and Effectiveness. Nutrients 2021, 13, 869. [Google Scholar] [CrossRef] [PubMed]
- Sestili, P.; Martinelli, C.; Colombo, E.; Barbieri, E.; Potenza, L.; Sartini, S.; Fimognari, C. Creatine as an antioxidant. Amino Acids 2011, 40, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Berikol, G.B.; Berikol, G.; Ayrik, C.; Kose, A.; Babus, S.B.; Gumus, L.T.; Arpaci, R.B.; Dag, A.; Ayan, E.; Gorur, A. Antioxidant and Neuroprotective Effects of L-arginine Administration After Traumatic Brain Injury and Hemorrhagic Shock in Rats. Turk. Neurosurg. 2022. [Google Scholar] [CrossRef]
- An, X.; Zhang, L.; Luo, J.; Zhao, S.; Jiao, T. Effects of Oat Hay Content in Diets on Nutrient Metabolism and the Rumen Microflora in Sheep. Animals 2020, 10, 2341. [Google Scholar] [CrossRef]
- Mc Geough, E.J.; O’Kiely, P.; Hart, K.J.; Moloney, A.P.; Boland, T.M.; Kenny, D.A. Methane emissions, feed intake, performance, digestibility, and rumen fermentation of finishing beef cattle offered whole-crop wheat silages differing in grain content. J. Anim. Sci. 2010, 88, 2703–2716. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Toxicity Testing in the 21st Century: A Vision and a Strategy; The National Academies Press: Washington, DC, USA, 2007. [Google Scholar]
- Zhang, D.; Yang, H. Combination effects of nitrocompounds, pyromellitic diimide, and 2-bromoethanesulfonate on in vitro ruminal methane production and fermentation of a grain-rich feed. J. Agric. Food. Chem. 2012, 60, 364–371. [Google Scholar] [CrossRef]
- Bremner, M.; Keeney, D.R. Distillation Methods for Determination of Ammonium Nitrate and Nitrite. Anal. Chim. Acta 1965, 32, 485–495. [Google Scholar] [CrossRef]
- Perez, J.F.; Balcells, J.; Cebrian, J.A.; Martin, S.M. Excretion of endogenous and exogenous purine derivatives in sheep: Effect of increased concentrate intake. Br. J. Nutr. 1998, 79, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Wada, T.; Shimbo, H.; Osaka, H. A simple screening method using ion chromatography for the diagnosis of cerebral creatine deficiency syndromes. Amino Acids 2012, 43, 993–997. [Google Scholar] [CrossRef]
- Han, H.; Zhang, L.; Shang, Y.; Wang, M.; Phillips, C.J.C.; Wang, Y.; Su, C.; Lian, H.; Fu, T.; Gao, T. Replacement of Maize Silage and Soyabean Meal with Mulberry Silage in the Diet of Hu Lambs on Growth, Gastrointestinal Tissue Morphology, Rumen Fermentation Parameters and Microbial Diversity. Animals 2022, 12, 1406. [Google Scholar] [CrossRef] [PubMed]
- Institute SAS Inc. SAS User’s Guide: Statistics; Version 9.2; SAS Institute Inc.: Cary, NC, USA, 2003. [Google Scholar]
- van der Pol, A.; van Gilst, W.H.; Voors, A.A.; van der Meer, P. Treating oxidative stress in heart failure: Past, present and future. Eur. J. Heart Fail. 2019, 21, 425–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celi, P. The role of oxidative stress in small ruminants’ health and production. Rev. Bras. Zootec. 2010, 39, 348–363. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, J.; Wang, C.; Guo, G.; Huo, W.; Xia, C.; Chen, L.; Zhang, Y.; Pei, C.; Liu, Q. Effects of guanidinoacetic acid supplementation on lactation performance, nutrient digestion and rumen fermentation in Holstein dairy cows. J. Sci. Food Agric. 2023, 103, 1522–1529. [Google Scholar] [CrossRef]
- Castillo-Gonzalez, A.R.; Burrola-Barraza, M.E.; Dominguez-Viveros, J.; Chavez-Martinez, A. Rumen microorganisms and fermentation. Arch. De Med. Vet. 2014, 46, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Li, W.; Wang, W.; Wang, Y.; Zhang, F.; Lv, L.; Yang, H. Foxtail millet (Setaria italica L.) silage compared peanut vine hay (Arachis hypogaea L.) exhibits greater feed efficiency via enhancing nutrient digestion and promoting rumen fermentation more efficiently in feedlotting lambs. Small Rumin. Res. 2022, 215, 106704. [Google Scholar] [CrossRef]
- Li, W.; Wu, Q.; Cui, Z.; Jiang, Y.; Aisikaer, A.; Zhang, F.; Chen, H.; Wang, W.; Wang, Y.; Lv, L.; et al. Guanidine acetic acid exhibited greater growth performance in younger (13–30 kg) than in older (30–50 kg) lambs under high-concentrate feedlotting pattern. Front. Vet. Sci. 2022, 9, 954675. [Google Scholar] [CrossRef]
- Mourino, F.; Akkarawongsa, R.A.; Weimer, P.J. Initial pH as a determinant of cellulose digestion rate by mixed ruminal microorganisms in vitro. J. Dairy Sci. 2001, 84, 848–859. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zang, C.; Pan, J.; Ma, C.; Wang, C.; Li, X.; Cai, W.; Yang, K. Effects of dietary guanidinoacetic acid on growth performance, guanidinoacetic acid absorption and creatine metabolism of lambs. PLoS ONE 2022, 17, e0264864. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Sun, Q. Laboratory Diagnosis of Cerebral Creatine Deficiency Syndromes by Determining Creatine and Guanidinoacetate in Plasma and Urine. Methods Mol. Biol. 2022, 2546, 129–140. [Google Scholar] [PubMed]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Xing, T.; Li, J.; Zhang, L.; Jiang, Y.; Gao, F. Guanidinoacetic acid supplementation improves intestinal morphology, mucosal barrier function of broilers subjected to chronic heat stress. J. Anim. Sci. 2022, 25, skac355. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Global Rumen Census Collaborators; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Zhu, W.; Zhu, W.; Liu, J.; Mao, S. Effect of dietary forage sources on rumen microbiota, rumen fermentation and biogenic amines in dairy cows. J. Sci. Food Agric. 2014, 94, 1886–1895. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Li, F.; Li, C.; Li, G.; Zhang, D.; Song, Q.; Li, X.; Zhao, Y.; Wang, W. Characterization of the rumen microbiota and its relationship with residual feed intake in sheep. Animal 2021, 15, 100161. [Google Scholar] [CrossRef]
- Flint, H.J. Encyclopedia of Food Microbiology, 2nd ed.; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- McCann, J.C.; Wiley, L.M.; Forbes, T.D.; Rouquette, F.M., Jr.; Tedeschi, L.O. Relationship between the rumen microbiome and residual feed intake-efficiency of Brahman bulls stocked on bermudagrass pastures. PLoS ONE 2014, 9, e91864. [Google Scholar] [CrossRef] [Green Version]
- Alzahal, O.; Li, F.; Le, L.G.; Walker, N.D.; Mcbride, B.W. Factors influencing ruminal bacterial community diversity and composition and microbial fibrolytic enzyme abundance in lactating dairy cows with a focus on the role of active dry yeast. J. Dairy Sci. 2017, 100, 4377–4393. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Chen, H.; Zhang, F.; Wang, W.; Xiong, F.; Liu, Y.; Lv, L.; Li, W.; Bo, Y.; Yang, H. Cysteamine Supplementation In Vitro Remarkably Promoted Rumen Fermentation Efficiency towards Propionate Production via Prevotella Enrichment and Enhancing Antioxidant Capacity. Antioxidants 2022, 11, 2233. [Google Scholar] [CrossRef]
- Zhang, K.; Qian, Q.; Mao, Y.; Xu, Y.; Yang, Y.; Chen, Y.; Wang, X. Characterization of growth phenotypes and gastrointestinal tract microbiota in sheep fed with caragana. J. Appl. Microbiol. 2021, 131, 2763–2779. [Google Scholar] [CrossRef]
- Ma, T.; Villot, C.; Renaud, D.; Skidmore, A.; Chevaux, E.; Steele, M.; Guan, L.L. Linking perturbations to temporal changes in diversity, stability, and compositions of neonatal calf gut microbiota: Prediction of diarrhea. ISME J. 2020, 14, 2223–2235. [Google Scholar] [CrossRef]
- Shen, Y.; Jiang, Y.; Zhang, S.; Zou, J.; Gao, X.; Song, Y.; Zhang, Y.; Hu, Y.; Huang, Y.; Jiang, Q. The Effect of Dietary Supplementation with Resveratrol on Growth Performance, Carcass and Meat Quality, Blood Lipid Levels and Ruminal Microbiota in Fattening Goats. Foods 2022, 11, 598. [Google Scholar] [CrossRef]
- Himelbloom, B.H.; Canale-Parola, E. Clostridium methylpentosum sp. nov.: A ring-shaped intestinal bacterium that ferments only methylpentoses and pentoses. Arch. Microbiol. 1989, 151, 287–293. [Google Scholar] [CrossRef]
- Kaakoush, N.O. Insights into the Role of Erysipelotrichaceae in the Human Host. Front. Cell. Infect. Microbiol. 2015, 5, 84. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yang, M.; Wu, Q.; Chen, J.; Deng, S.; Chen, L.; Wei, D.; Liang, F. Improvement of intestinal flora: Accompany with the antihypertensive effect of electroacupuncture on stage 1 hypertension. Chin. Med. 2021, 16, 7. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, J.; Keller, D.; Verbruggen, S.; Abboud, K.Y.; Venema, K. A blend of 3 mushrooms dose-dependently increases butyrate production by the gut microbiota. Benef. Microbes 2021, 12, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.M.A.E.; Bast, A.; Vanhoutvin, S.A.L.W.; Fischer, M.A.J.G.; Kodde, A.; Troost, F.J.; Venema, K.; Brummer, R.J.M. Butyrate modulates oxidative stress in the colonic mucosa of healthy humans. Clin. Nutr. 2009, 28, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.M.; Hosseini, A.; Zhou, Z.; Alharthi, A.; Trevisi, E.; Osorio, J.S.; Loor, J.J. Reticulo-rumen mass, epithelium gene expression, and systemic biomarkers of metabolism and inflammation in Holstein dairy cows fed a high-energy diet. J. Dairy Sci. 2017, 100, 9352–9360. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, H.; Zhang, T.; Zhang, R.; Liu, R.; Chen, Y. Molecular mechanism on cadmium-induced activity changes of catalase and superoxide dismutase. Int. J. Biol. Macromol. 2015, 77, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Shukla, R.; Madhu, S.V.; Gambhir, J.K.; Prabhu, K.M. Antioxidant status, lipid peroxidation and nitric oxide end products in patients of type 2 diabetes mellitus with nephropathy. Clin. Biochem. 2003, 36, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Nauseef, W.M. Detection of superoxide anion and hydrogen peroxide production by cellular NADPH oxidases. Biochim. Biophys. Acta 2014, 1840, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [Green Version]
- Ostojic, S.M.; Stojanovic, M.D.; Olcina, G. Oxidant-Antioxidant Capacity of Dietary Guanidinoacetic Acid. Ann. Nutr. Metab. 2015, 67, 243–246. [Google Scholar] [CrossRef]
- Lawler, J.M.; Barnes, W.S.; Wu, G.; Song, W.; Demaree, S. Direct antioxidant properties of creatine. Biochem. Biophys. Res. Commun. 2002, 290, 47–52. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, Y.; Zha, X.; Ma, Y.; Liu, X.; Elsabagh, M.; Wang, H.; Wang, M. Dietary L-Arginine or N-Carbamylglutamate Alleviates Colonic Barrier Injury, Oxidative Stress, and Inflammation by Modulation of Intestinal Microbiota in Intrauterine Growth-Retarded Suckling Lambs. Antioxidants 2022, 11, 2251. [Google Scholar] [CrossRef]
- Ma, X.; Fan, P.X.; Li, L.S.; Qiao, S.Y.; Zhang, G.L.; Li, D.F. Butyrate promotes the recovering of intestinal wound healing through its positive effect on the tight junctions. J. Anim. Sci. 2012, 90 (Suppl. 4), 266–268. [Google Scholar] [CrossRef] [Green Version]

| Items | Forage | GAA Addition | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Control | UGAA | CGAA | Forage | GAA | Forage × GAA | |||
| Stage 1 (60 d) | ||||||||
| Total VFA, mmol/L | OH | 78.79 b | 84.20 a | 86.21 a | 0.78 | 0.245 | <0.001 | 0.010 |
| OHWS | 82.39 b | 83.79 ab | 85.27 a | |||||
| Acetate, % | OH | 56.11 b | 56.69 a | 56.17 b | 0.16 | 0.111 | 0.089 | 0.071 |
| OHWS | 55.93 | 56.07 | 56.32 | |||||
| Propionate, % | OH | 32.28 a | 31.26 b | 32.43 a | 0.18 | 0.084 | 0.009 | 0.001 |
| OHWS | 32.34 | 32.32 | 32.07 | |||||
| Butyrate, % | OH | 9.06 b | 9.09 b | 9.57 a | 0.16 | 0.508 | 0.257 | 0.132 |
| OHWS | 9.12 | 9.21 | 9.14 | |||||
| A:P | OH | 1.74 b | 1.81 a | 1.73 b | 0.01 | 0.059 | 0.011 | 0.001 |
| OHWS | 1.73 | 1.74 | 1.76 | |||||
| pH | OH | 6.86 | 6.77 | 6.80 | 0.03 | 0.240 | 0.023 | 0.917 |
| OHWS | 6.82 | 6.74 | 6.78 | |||||
| NH3-N, mg/dL | OH | 9.51 | 9.89 | 9.83 | 0.16 | 0.592 | 0.025 | 0.873 |
| OHWS | 9.39 b | 9.92 a | 9.70 ab | |||||
| MCP, mg/mL | OH | 42.51 | 43.24 | 43.25 | 0.33 | 0.394 | 0.015 | 0.914 |
| OHWS | 42.58 b | 43.56 a | 43.56 a | |||||
| Stage 2 (120 d) | ||||||||
| Total VFA, mmol/L | OH | 83.11 b | 86.74 a | 87.42 a | 0.59 | 0.018 | <0.001 | 0.007 |
| OHWS | 86.70 | 86.93 | 87.54 | |||||
| Acetate, % | OH | 54.40 | 54.91 | 54.86 | 0.22 | 0.066 | 0.575 | 0.003 |
| OHWS | 54.98 a | 54.07 b | 54.10 b | |||||
| Propionate, % | OH | 33.95 a | 32.95 b | 33.32 b | 0.24 | 0.817 | 0.133 | 0.067 |
| OHWS | 33.34 | 33.37 | 33.63 | |||||
| Butyrate, % | OH | 8.94 | 9.29 | 9.48 | 0.19 | 0.334 | 0.008 | 0.787 |
| OHWS | 8.97 b | 9.61 a | 9.58 a | |||||
| A:P | OH | 1.60 b | 1.67 a | 1.65 a | 0.01 | 0.311 | 0.416 | 0.003 |
| OHWS | 1.65 | 1.62 | 1.61 | |||||
| pH | OH | 6.89 a | 6.69 b | 6.77 ab | 0.05 | 0.561 | 0.082 | 0.364 |
| OHWS | 6.83 | 6.79 | 6.81 | |||||
| NH3-N, mg/dL | OH | 9.96 | 10.39 | 10.28 | 0.35 | 0.158 | 0.160 | 0.773 |
| OHWS | 10.09 | 10.97 | 10.83 | |||||
| MCP, mg/mL | OH | 43.44 | 44.30 | 44.10 | 0.30 | 0.825 | 0.011 | 0.978 |
| OHWS | 43.46 b | 44.43 a | 44.12 ab | |||||
| Items | Forage | GAA Addition | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Control | UGAA | CGAA | Forage | GAA | Forage × GAA | |||
| Stage 1 (60 d) | ||||||||
| GAA, μg/mL | OH | 57.56 c | 99.90 a | 86.45 b | 1.97 | 0.899 | <0.001 | 0.983 |
| OHWS | 57.87 c | 99.70 a | 86.97 b | |||||
| Creatine, μg/mL | OH | 34.80 | 35.18 | 36.83 | 1.17 | 0.099 | 0.341 | 0.634 |
| OHWS | 36.12 | 38.01 | 37.49 | |||||
| GAMT, U/L | OH | 7.96 b | 9.65 a | 9.58 a | 0.40 | 0.973 | <0.001 | 0.873 |
| OHWS | 7.97 b | 9.87 a | 9.38 ab | |||||
| AGAT, U/L | OH | 13.56 a | 13.30 a | 12.68 b | 0.31 | 0.973 | 0.025 | 0.998 |
| OHWS | 13.55 a | 13.31 ab | 12.65 b | |||||
| ATP, μmol/L | OH | 3.61 | 3.66 | 3.64 | 0.02 | 0.571 | 0.014 | 0.725 |
| OHWS | 3.60 b | 3.65 a | 3.65 a | |||||
| Stage 2 (120 d) | ||||||||
| GAA, μg/mL | OH | 62.34 c | 100.03 a | 87.15 b | 1.59 | 0.856 | <0.001 | 0.838 |
| OHWS | 61.24 c | 99.64 a | 87.93 b | |||||
| Creatine, μg/mL | OH | 36.64 | 37.69 | 37.60 | 1.41 | 0.205 | 0.724 | 0.949 |
| OHWS | 38.29 | 39.51 | 38.57 | |||||
| GAMT, U/L | OH | 11.08 | 11.13 | 11.57 | 0.24 | 0.943 | 0.430 | 0.594 |
| OHWS | 11.18 | 11.33 | 11.31 | |||||
| AGAT, U/L | OH | 14.13 | 14.54 | 14.70 | 0.45 | 0.429 | 0.157 | 0.870 |
| OHWS | 14.27 | 14.65 | 15.16 | |||||
| ATP, μmol/L | OH | 3.62 b | 3.66 ab | 3.69 a | 0.02 | 0.388 | 0.001 | 0.937 |
| OHWS | 3.61 b | 3.65 ab | 3.67 a | |||||
| Items | Forage | GAA Addition | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Control | UGAA | CGAA | Forage | GAA | Forage × GAA | |||
| Stage 1 (60 d) | ||||||||
| Firmicutes | OH | 61.91 | 59.20 | 52.22 | 6.07 | 0.112 | 0.612 | 0.297 |
| OHWS | 68.25 | 58.79 | 71.10 | |||||
| Bacteroidota | OH | 28.11 ab | 19.23 b | 41.50 a | 5.31 | 0.747 | 0.447 | 0.026 |
| OHWS | 26.56 | 33.82 | 24.21 | |||||
| Synergistota | OH | 0.28 | 14.93 | 0.48 | 5.25 | 0.277 | 0.291 | 0.314 |
| OHWS | 0.40 | 0.59 | 0.30 | |||||
| Patescibacteria | OH | 1.73 | 1.45 | 2.79 | 0.66 | 0.671 | 0.331 | 0.686 |
| OHWS | 1.27 | 1.88 | 2.12 | |||||
| Actinobacteriota | OH | 5.20 | 0.64 | 0.38 | 1.32 | 0.556 | 0.092 | 0.189 |
| OHWS | 1.73 | 2.01 | 0.55 | |||||
| Proteobacteria | OH | 0.84 | 3.05 | 0.88 | 0.92 | 0.163 | 0.448 | 0.364 |
| OHWS | 0.47 | 0.41 | 0.61 | |||||
| Stage 2 (120 d) | ||||||||
| Firmicutes | OH | 44.31 b | 61.95 a | 54.79 a | 4.13 | 0.039 | 0.213 | 0.036 |
| OHWS | 65.31 | 61.63 | 56.68 | |||||
| Bacteroidota | OH | 48.79 a | 31.32 b | 38.8 ab | 4.54 | 0.091 | 0.267 | 0.047 |
| OHWS | 28.05 | 32.49 | 38.50 | |||||
| Actinobacteriota | OH | 3.11 | 3.58 | 1.39 | 1.38 | 0.627 | 0.628 | 0.675 |
| OHWS | 1.52 | 2.69 | 2.20 | |||||
| Patescibacteria | OH | 1.09 | 1.00 | 2.41 | 0.67 | 0.507 | 0.463 | 0.168 |
| OHWS | 2.13 a | 0.72 b | 0.77 b | |||||
| Spirochaetota | OH | 0.89 | 0.28 | 1.55 | 0.51 | 0.261 | 0.909 | 0.140 |
| OHWS | 1.47 | 1.77 | 0.92 | |||||
| Items | Forage | GAA Addition | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Control | UGAA | CGAA | Forage | GAA | Forage × GAA | |||
| Stage 1 (60 d) | ||||||||
| Ruminococcus | OH | 6.77 | 28.54 | 15.75 | 8.73 | 0.682 | 0.286 | 0.236 |
| OHWS | 13.94 | 14.39 | 31.64 | |||||
| Prevotella | OH | 12.39 b | 9.69 b | 26.86 a | 4.22 | 0.313 | 0.134 | 0.108 |
| OHWS | 13.10 | 12.72 | 12.39 | |||||
| Rikenellaceae_RC9_gut_group | OH | 7.87 | 1.93 | 5.35 | 2.60 | 0.583 | 0.761 | 0.104 |
| OHWS | 5.04 | 9.89 | 3.79 | |||||
| NK4A214_group | OH | 6.02 a | 2.29 b | 4.23 ab | 0.97 | 0.999 | 0.269 | 0.040 |
| OHWS | 4.20 | 5.39 | 2.96 | |||||
| CAG-352 | OH | 0.06 | 2.38 | 5.26 | 3.78 | 0.395 | 0.404 | 0.061 |
| OHWS | 13.71 | 1.55 | 0.54 | |||||
| Christensenellaceae_R-7_group | OH | 5.09 | 2.17 | 3.12 | 1.08 | 0.489 | 0.107 | 0.095 |
| OHWS | 4.75 | 5.64 | 1.87 | |||||
| Succiniclasticum | OH | 8.50 a | 0.46 b | 3.10 ab | 1.89 | 0.368 | 0.049 | 0.246 |
| OHWS | 3.87 | 2.40 | 1.53 | |||||
| Fretibacterium | OH | 0.22 | 14.93 | 0.42 | 5.26 | 0.279 | 0.287 | 0.314 |
| OHWS | 0.32 | 0.59 | 0.27 | |||||
| norank_f__norank_o__Clostridia_UCG-014 | OH | 1.75 a | 0.25 b | 0.87 ab | 3.14 | 0.189 | 0.521 | 0.522 |
| OHWS | 2.77 | 2.04 | 8.57 | |||||
| Stage 2 (120 d) | ||||||||
| Ruminococcus | OH | 5.14 | 16.03 | 12.43 | 4.76 | 0.502 | 0.770 | 0.027 |
| OHWS | 23.85 a | 7.74 b | 10.00 ab | |||||
| Prevotella | OH | 30.06 a | 12.19 b | 11.06 b | 4.15 | 0.214 | 0.075 | 0.042 |
| OHWS | 12.78 | 11.86 | 15.58 | |||||
| Rikenellaceae_RC9_gut_group | OH | 6.58 | 7.76 | 15.04 | 2.97 | 0.424 | 0.089 | 0.769 |
| OHWS | 6.17 | 6.69 | 10.58 | |||||
| Christensenellaceae_R-7_group | OH | 3.29 | 7.46 | 8.47 | 1.45 | 0.584 | 0.119 | 0.303 |
| OHWS | 5.29 | 5.74 | 6.20 | |||||
| NK4A214_group | OH | 3.49 | 4.05 | 5.43 | 1.02 | 0.008 | 0.040 | 0.578 |
| OHWS | 4.74 | 7.25 | 8.46 | |||||
| F082 | OH | 2.03 | 6.91 | 5.04 | 1.52 | 0.358 | 0.275 | 0.315 |
| OHWS | 3.34 | 3.45 | 3.67 | |||||
| Lachnospiraceae_NK3A20_group | OH | 1.00 | 3.61 | 1.18 | 1.16 | 0.038 | 0.105 | 0.966 |
| OHWS | 3.23 | 5.39 | 3.54 | |||||
| Succiniclasticum | OH | 2.93 a | 0.97 b | 2.17 a | 1.05 | 0.062 | 0.226 | 0.006 |
| OHWS | 1.29 | 6.88 | 2.99 | |||||
| norank_f__norank_o__Clostridia_UCG-014 | OH | 2.31 | 1.25 | 2.20 | 1.36 | 0.171 | 0.490 | 0.170 |
| OHWS | 1.31 | 5.61 | 3.58 | |||||
| Items | Forage | GAA Addition | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Control | UGAA | CGAA | Forage | GAA | Forage × GAA | |||
| Stage 1 (60 d) | ||||||||
| T-AOC, U/mL | OH | 11.29 b | 11.91 b | 12.75 a | 0.25 | 0.596 | <0.001 | 0.274 |
| OHWS | 11.35 b | 12.46 a | 12.48 a | |||||
| SOD, U/mL | OH | 97.07 c | 123.57 b | 176.26 a | 2.54 | 0.061 | <0.001 | <0.001 |
| OHWS | 96.79 c | 184.66 a | 127.49 b | |||||
| CAT, U/mL | OH | 8.60 b | 8.97 b | 10.06 a | 0.21 | 0.259 | <0.001 | <0.001 |
| OHWS | 8.45 c | 10.31 a | 9.46 b | |||||
| GSH-Px, U/mL | OH | 806.07 c | 840.07 b | 924.25 a | 8.78 | 0.898 | <0.001 | <0.001 |
| OHWS | 800.20 c | 924.27 a | 848.73 b | |||||
| GSH, nmol/mL | OH | 9.23 b | 9.69 b | 10.84 a | 0.21 | 0.123 | <0.001 | 0.060 |
| OHWS | 9.32 b | 10.55 a | 10.71 a | |||||
| MDA, nmol/mL | OH | 5.82 a | 5.33 a | 5.20 b | 0.17 | 0.305 | <0.001 | 0.274 |
| OHWS | 5.84 a | 4.87 b | 5.21 b | |||||
| Stage 2 (120 d) | ||||||||
| T-AOC, U/mL | OH | 13.87 c | 14.60 b | 15.22 a | 0.19 | 0.097 | <0.001 | 0.001 |
| OHWS | 14.12 c | 15.61 a | 14.75 b | |||||
| SOD, U/mL | OH | 107.31 b | 121.5 b | 169.68 a | 5.35 | 0.179 | <0.001 | <0.001 |
| OHWS | 95.68 c | 183.83 a | 137 b | |||||
| CAT, U/mL | OH | 10.46 | 10.75 | 11.00 | 0.40 | 0.322 | 0.048 | 0.110 |
| OHWS | 10.32 b | 12.08 a | 10.81 ab | |||||
| GSH-Px, U/mL | OH | 931.47 b | 948.30 b | 1007.26 a | 12.54 | 0.619 | <0.001 | <0.001 |
| OHWS | 909.69 c | 1031.11 a | 961.74 b | |||||
| GSH, nmol/mL | OH | 11.81 | 12.16 | 12.63 | 0.26 | 0.370 | 0.009 | 0.845 |
| OHWS | 11.50 b | 12.12 ab | 12.40 a | |||||
| MDA, nmol/mL | OH | 6.15 a | 6.16 a | 5.30 b | 0.12 | 0.454 | <0.001 | <0.001 |
| OHWS | 6.29 a | 5.48 b | 5.61 b | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Cui, Z.; Jiang, Y.; Aisikaer, A.; Wu, Q.; Zhang, F.; Wang, W.; Bo, Y.; Yang, H. Dietary Guanidine Acetic Acid Improves Ruminal Antioxidant Capacity and Alters Rumen Fermentation and Microflora in Rapid-Growing Lambs. Antioxidants 2023, 12, 772. https://doi.org/10.3390/antiox12030772
Li W, Cui Z, Jiang Y, Aisikaer A, Wu Q, Zhang F, Wang W, Bo Y, Yang H. Dietary Guanidine Acetic Acid Improves Ruminal Antioxidant Capacity and Alters Rumen Fermentation and Microflora in Rapid-Growing Lambs. Antioxidants. 2023; 12(3):772. https://doi.org/10.3390/antiox12030772
Chicago/Turabian StyleLi, Wenjuan, Zhaoyang Cui, Yaowen Jiang, Ailiyasi Aisikaer, Qichao Wu, Fang Zhang, Weikang Wang, Yukun Bo, and Hongjian Yang. 2023. "Dietary Guanidine Acetic Acid Improves Ruminal Antioxidant Capacity and Alters Rumen Fermentation and Microflora in Rapid-Growing Lambs" Antioxidants 12, no. 3: 772. https://doi.org/10.3390/antiox12030772