
The NADPH Oxidase Inhibitors Apocynin and Diphenyleneiodonium Protect Rats from LPS-Induced Pulmonary Inflammation

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animal Care
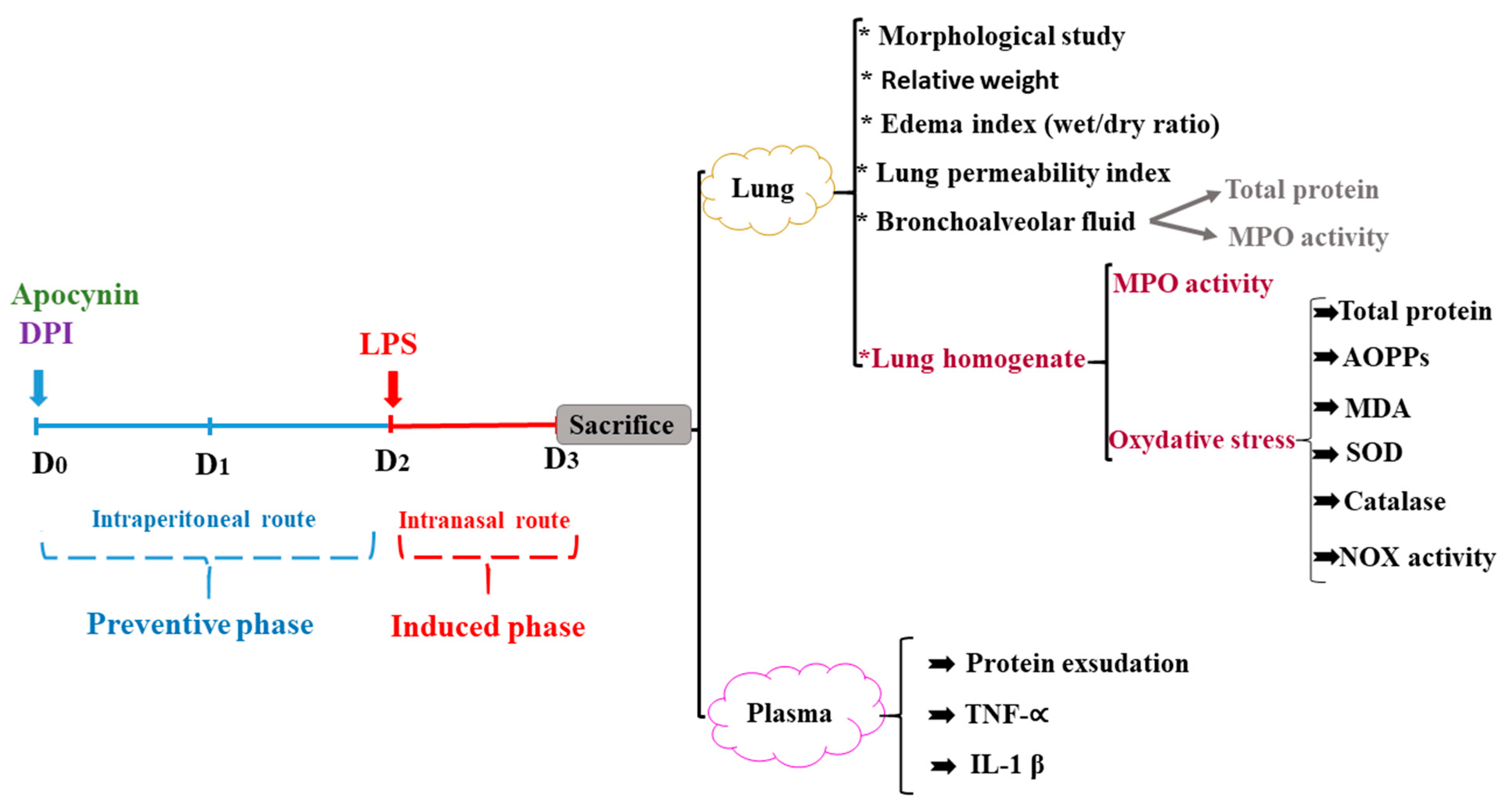
2.3. In Vivo Experimental Protocol
2.4. Body Weight Changes and Organ Measurement
2.5. Bronchoalveolar Liquid (BAL) and Lung Permeability Index Determination
2.6. Lung Wet/Dry Weight Ratio
2.7. Histological Study
2.8. Lung Homogenate and Protein Determination
2.9. NOX Activity
2.10. Advanced Oxidation of Protein
2.11. Lipid Peroxidation Was Measured as Malondialdehyde
2.12. Superoxide Dismutase (SOD) and Catalase Activity
2.13. Assessment of Myeloperoxidase (MPO) Activity and Pro-Inflammatory Cytokines (IL-1β, TNF-α) Levels
2.14. Statistical Analysis
3. Results
3.1. Apocynin and DPI, at 10 mg/kg and 100 ng/kg Respectively, Are Not Toxic for Rats
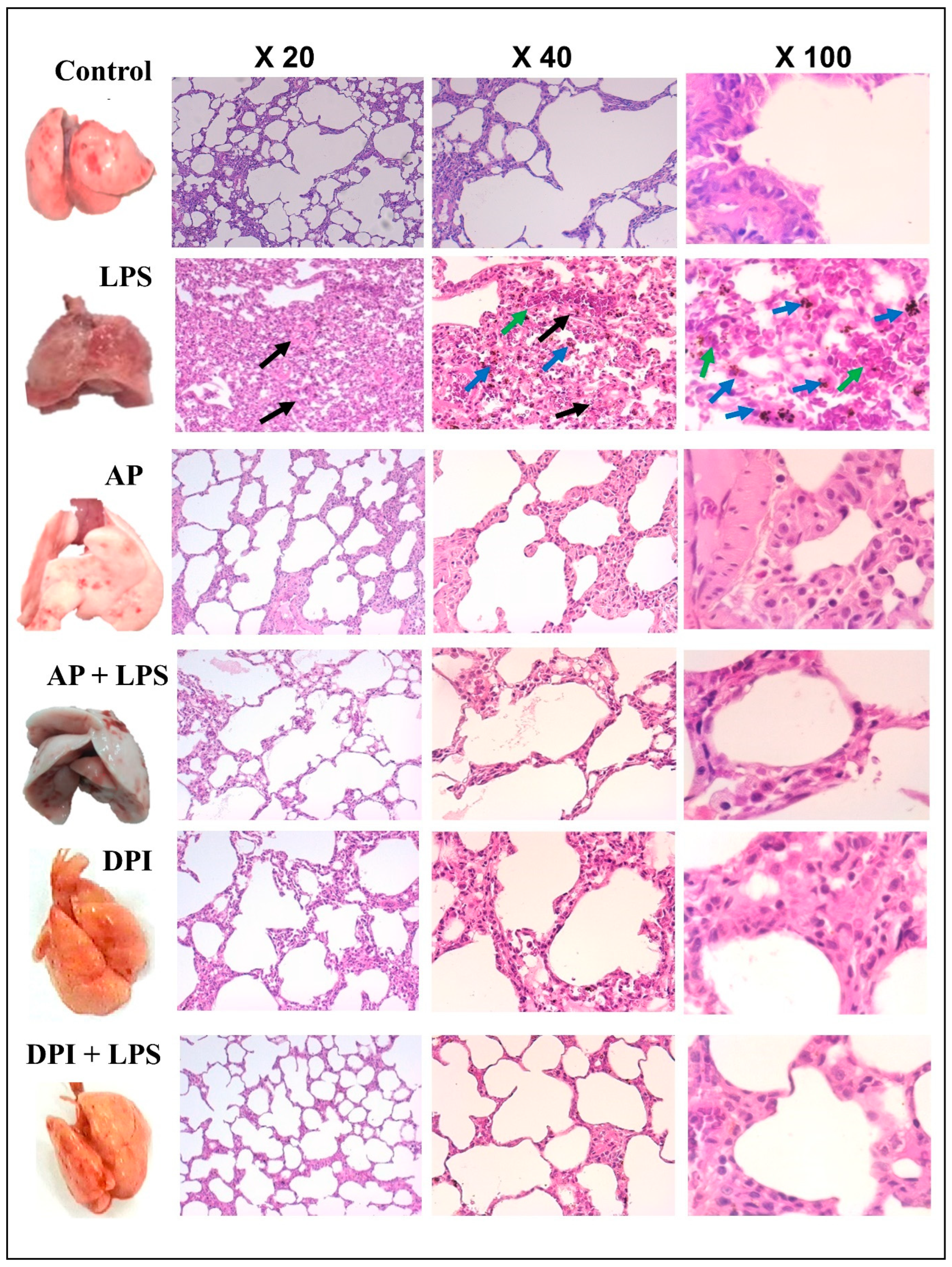
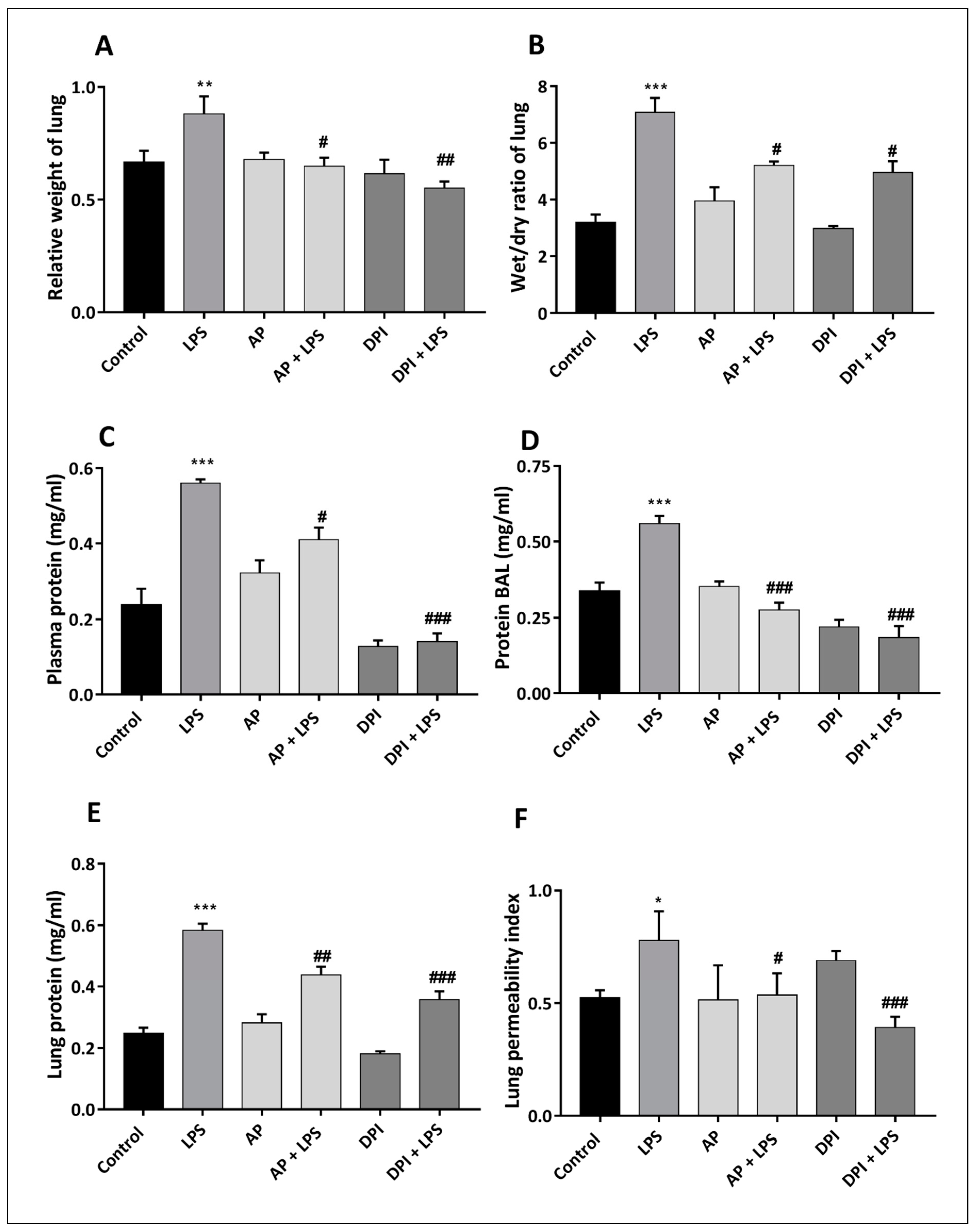
3.2. Morphological and Histological Evaluation of the Effects of Apocynin and DPI on Rat Lung in the Absence or Presence of LPS
3.3. Apocynin and DPI Reduce LPS-Induced Oxidative Stress in Rat
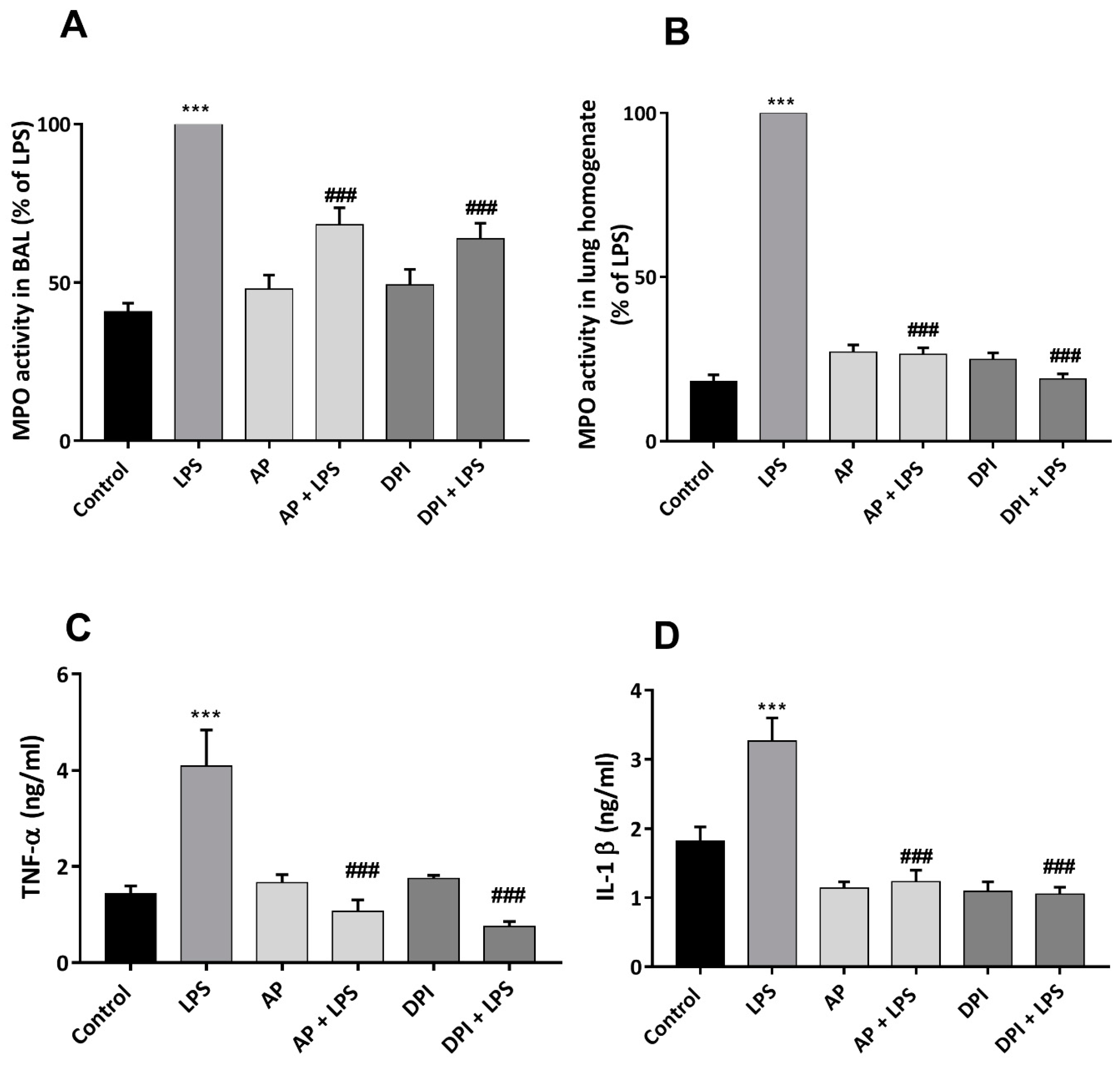
3.4. Apocynin and DPI Reduce LPS-Induced Increase in Mediators of Inflammation Levels in Rat
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AOPP | Advanced oxidation protein products |
| AP | Apocynin |
| ATSAL | Tunisian Association of Laboratory Animals Science |
| BAL | Broncho alveolar fluid |
| BHT | Butylated hydroxytoluene |
| BSA | Bovine serum albumin |
| DMSO | Dimethyl sulfoxide |
| DPI | Diphenyleneiodonium |
| HRP | Horseradish peroxidase |
| HTAB | Hexadecyltrimethylammonuim bromide |
| i.p. | intraperitoneal |
| KI | Potassium iodide |
| LPS | Lipopolysaccharide |
| Luminol | (5-amino2,3-dihydro-1,4-phtalazinedione) |
| MDA | Malondialdehyde |
| MPO | Myeloperoxidase activity |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NOXs | NADPH oxidases |
| PBS | Phosphate-buffered saline |
| ROS | Reactive Oxygen Species |
| SOD | Superoxide dismutase |
| TBA | Thiobarbituric acid |
| TCA | Trichloroacetic acid |
References
- Kumar, V. Pulmonary Innate Immune Response Determines the Outcome of Inflammation During Pneumonia and Sepsis-Associated Acute Lung Injury. Front. Immunol. 2020, 11, 1722. [Google Scholar] [CrossRef] [PubMed]
- Boskabadi, J.; Askari, V.R.; Hosseini, M.; Boskabady, M.H. Immunomodulatory properties of captopril, an ACE inhibitor, on LPS-induced pulmonary inflammation and fibrosis and on oxidative stress. Inflammopharmacology 2019, 27, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Stearns-Kurosawa, D.J.; Osuchowski, M.F.; Valentine, C.; Kurosawa, S.; Remick, D.G. The pathogenesis of sepsis. Annu. Rev. Pathol. 2011, 6, 19–48. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Li, H.; Tang, H.; Jin, Y.; Li, W.; Sun, Y.; Feng, P.; Sun, X.; Xia, Z. Hydrogen inhalation ameliorates lipopolysaccharide-induced acute lung injury in mice. Int. Immunopharmacol. 2011, 11, 2130–2137. [Google Scholar] [CrossRef]
- Matthay, M.A.; Ware, L.B.; Zimmerman, G.A. The acute respiratory distress syndrome. J. Clin. Investig. 2012, 122, 2731–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dröge, W.; Schipper, H.M. Oxidative stress and aberrant signaling in aging and cognitive decline. Aging Cell 2007, 6, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Griffith, B.; Pendyala, S.; Hecker, L.; Lee, P.J.; Natarajan, V.; Thannickal, V.J. NOX enzymes and pulmonary disease. Antioxid. Redox Signal. 2009, 11, 2505–2516. [Google Scholar] [CrossRef] [Green Version]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.C.; Gougerot-Pocidalo, M.A.; Dang, P.M. Priming of the neutrophil respiratory burst: Role in host defense and inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef]
- Segal, A.W. The NADPH oxidase and chronic granulomatous disease. Mol. Med. Today 1996, 2, 129–135. [Google Scholar] [CrossRef]
- Rahman, I.; MacNee, W. Oxidative stress and regulation of glutathione in lung inflammation. Eur. Respir. J. 2000, 16, 534–554. [Google Scholar] [CrossRef]
- Dang, P.M.; Rolas, L.; El-Benna, J. The Dual Role of Reactive Oxygen Species-Generating Nicotinamide Adenine Dinucleotide Phosphate Oxidases in Gastrointestinal Inflammation and Therapeutic Perspectives. Antioxid. Redox Signal. 2020, 33, 354–373. [Google Scholar] [CrossRef] [PubMed]
- Wind, S.; Beuerlein, K.; Eucker, T.; Müller, H.; Scheurer, P.; Armitage, M.E.; Ho, H.; Schmidt, H.H.; Wingler, K. Comparative pharmacology of chemically distinct NADPH oxidase inhibitors. Br. J. Pharmacol. 2010, 161, 885–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Benna, J.; Dang, P.M.; Périanin, A. Peptide-based inhibitors of the phagocyte NADPH oxidase. Biochem. Pharmacol. 2010, 80, 778–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ’t Hart, B.A.; Copray, S.; Philippens, I. Apocynin, a low molecular oral treatment for neurodegenerative disease. Biomed. Res. Int. 2014, 2014, 298020. [Google Scholar] [CrossRef] [PubMed]
- Heumüller, S.; Wind, S.; Barbosa-Sicard, E.; Schmidt, H.H.; Busse, R.; Schröder, K.; Brandes, R.P. Apocynin is not an inhibitor of vascular NADPH oxidases but an antioxidant. Hypertension 2008, 51, 211–217. [Google Scholar] [CrossRef]
- Cross, A.R.; Jones, O.T. The effect of the inhibitor diphenyleneiodonium on the superoxide-generating system of neutrophils. Specific labelling of a component polypeptide of the oxidase. Biochem. J. 1986, 237, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Doussière, J.; Vignais, P.V. Diphenyleniodonium as an inhibitor of the NADPH oxidase complex of bovine neutrophils. Factors controlling the inhibitory potency of diphenyleneiodonium in a cell-free system of oxidase activation. Eur. J. Biochem. 1992, 208, 61–71. [Google Scholar] [CrossRef]
- Wang, Q.; Chu, C.H.; Oyarzabal, E.; Jiang, L.; Chen, S.H.; Wilson, B.; Qian, L.; Hong, J.S. Subpicomolar diphenyleneiodonium inhibits microglial NADPH oxidase with high specificity and shows great potential as a therapeutic agent for neurodegenerative diseases. Glia 2014, 62, 2034–2043. [Google Scholar] [CrossRef] [Green Version]
- Stuehr, D.J.; Fasehun, O.A.; Kwon, N.S.; Gross, S.S.; Gonzalez, J.A.; Levi, R.; Nathan, C.F. Inhibition of macrophage and endothelial cell nitric oxide synthase by diphenyleneiodonium and its analogues. FASEB J. 1991, 5, 98–103. [Google Scholar] [CrossRef] [Green Version]
- Holland, P.C.; Clark, M.G.; Bloxham, D.P.; Lardy, H.A. Mechanism of action of the hypoglycemic agent diphenyleneiodonium. J. Biol. Chem. 1973, 248, 6050–6056. [Google Scholar] [CrossRef]
- Kuai, Y.; Liu, H.; Liu, D.; Liu, Y.; Sun, Y.; Xi, J.; Sun, J.; Fang, Y.; Pan, H.; Han, W. An ultralow dose of the NADPH oxidase inhibitor diphenyleneiodonium (DPI) is an economical and effective therapeutic agent for the treatment of colitis-associated colorectal cancer. Theranostics 2020, 10, 6743–6757. [Google Scholar] [CrossRef] [PubMed]
- El-Alfy, T.S.; Hetta, M.H.; Yassin, N.A.; Rahman, R.F.; Kadry, E.M. Estrogenic activity of Citrus medica L. leaves growing in Egypt. J. Appl. Pharm. Sci. 2012, 2, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Li, Y.; Wan, S.; Wang, Y.; Yu, P. Protective effects of apocynin nitrone on acute lung injury induced by lipopolysaccharide in rats. Int. Immunopharmacol. 2014, 20, 377–382. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 93, 265–275. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Friedlander, M.; Capeillère-Blandin, C.; Nguyen-Khoa, T.; Nguyen, A.T.; Zingraff, J.; Jungers, P.; Descamps-Latscha, B. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int. 1996, 49, 1304–1313. [Google Scholar] [CrossRef] [Green Version]
- Yagi, K. Simple Fluorometric Assay for lipoperoxyde in blood plasma. Biochem. Med. 1976, 49, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Goldblum, S.E.; Wu, K.M.; Jay, M. Lung myeloperoxidase as a measure of pulmonary leukostasis in rabbits. J. Appl. Physiol. 1985, 59, 1978–1985. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Mazzon, E.; Bauer, M.; Bauer, I.; Delbosc, S.; Cristol, J.P.; Patel, N.S.; Cuzzocrea, S.; Thiemermann, C. Inhibitors of NADPH oxidase reduce the organ injury in hemorrhagic shock. Shock 2005, 23, 107–114. [Google Scholar] [CrossRef]
- Kumari, A.; Tyagi, N.; Dash, D.; Singh, R. Intranasal curcumin ameliorates lipopolysaccharide-induced acute lung injury in mice. Inflammation 2015, 38, 1103–1112. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Liu, G.Q.; Zeng, Y.Y.; Zhu, J.J.; Liu, Z.Y.; Zhang, X.; Huang, J.A. Role of IL-17 in LPS-induced acute lung injury: An in vivo study. Oncotarget 2017, 8, 93704–93711. [Google Scholar] [CrossRef] [Green Version]
- Li-Mei, W.; Jie, T.; Shan-He, W.; Dong-Mei, M.; Peng-Jiu, Y. Anti-inflammatory and Anti-oxidative Effects of Dexpanthenolon Lipopolysaccharide Induced Acute Lung Injury in Mice. Inflammation 2016, 39, 1757–1763. [Google Scholar] [CrossRef]
- Bernard, K.; Hecker, L.; Luckhardt, T.R.; Cheng, G.; Thannickal, V.J. NADPH oxidases in lung health and disease. Antioxid. Redox Signal. 2014, 20, 2838–2853. [Google Scholar] [CrossRef] [Green Version]
- Augsburger, F.; Filippova, A.; Rasti, D.; Seredenina, T.; Lam, M.; Maghzal, G.; Mahiout, Z.; Jansen-Dürr, P.; Knaus, U.G.; Doroshow, J.; et al. Pharmacological characterization of the seven human NOX isoforms and their inhibitors. Redox Biol. 2019, 26, 101272. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; Moochhala, S. Role of inflammatory mediators in the pathophysiology of acute respiratory distress syndrome. J. Pathol. 2004, 202, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Jin, S.; Wang, J.; Zhang, G.; Kawanokuchi, J.; Kuno, R.; Sonobe, Y.; Mizuno, T.; Suzumura, A. Tumor necrosis factor-alpha induces neurotoxicity via glutamate release from hemichannels of activated microglia in an autocrine manner. J. Biol. Chem. 2006, 281, 21362–21368. [Google Scholar] [CrossRef] [Green Version]
- Gouwy, M.; Struyf, S.; Proost, P.; Van Damme, J. Synergy in cytokine and chemokine networks amplifies the inflammatory response. Cytokine Growth Factor Rev. 2005, 16, 561–580. [Google Scholar] [CrossRef]
- Kolb, M.; Margetts, P.J.; Anthony, D.C.; Pitossi, F.; Gauldie, J. Transient expression of IL-1beta induces acute lung injury and chronic repair leading to pulmonary fibrosis. J. Clin. Investig. 2001, 107, 1529–1536. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, A.C.; Dos Santos Vilela, M.M.; Condino-Neto, A.; Ximenes, V.F. The importance of myeloperoxidase in apocynin-mediated NADPH oxidase inhibition. ISRN Inflamm. 2012, 22, 260453. [Google Scholar] [CrossRef] [Green Version]
- Dodd-o, J.M.; Welsh, L.E.; Salazar, J.D.; Walinsky, P.L.; Peck, E.A.; Shake, J.G.; Caparrelli, D.J.; Ziegelstein, R.C.; Zweier, J.L.; Baumgartner, W.A.; et al. Effect of NADPH oxidase inhibition on cardiopulmonary bypass-induced lung injury. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H927–H936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chian, C.F.; Chiang, C.H.; Yuan-Jung, C.; Chuang, C.H.; Liu, S.L.; Yi-Han, J.; Zhang, H.; Ryu, J.H. Apocynin attenuates lipopolysaccharide-induced lung injury in an isolated and perfused rat lung model. Shock 2012, 38, 196–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, H.; Matsumura, T.; Ishii, N.; Fukuda, K.; Senokuchi, T.; Motoshima, H.; Kondo, T.; Taketa, K.; Kawasaki, S.; Hanatani, S.; et al. Apocynin suppresses the progression of atherosclerosis in apoE-deficient mice by inactivation of macrophages. Biochem. Biophys. Res. Commun. 2013, 431, 124–130. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Control | LPS | AP | AP + LPS | DPI | DPI + LPS | |
|---|---|---|---|---|---|---|---|
| Phase | |||||||
| Preventive phase | Day 1 | 215.05 ± 5.7 | 226.3 ± 6.7 | 196.71 ± 2.4 | 206.45 ± 8.9 | 232.53 ± 7.5 | 198.73 ± 1.6 |
| Day 2 | 208.25 ± 6.1 | 223.19 ± 5.1 | 196.05 ± 1.9 | 203.3 ± 9.33 | 220.13 ± 6.2 | 189.95 ± 2.05 * | |
| Induced phase | Day 3 | 202.33 ± 5.8 | 218.18 ± 3.9 | 191.44 ± 2.3 | 194.88 ± 8.9 | 212.98 ± 5.7 | 187.93 ± 2.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kouki, A.; Ferjani, W.; Ghanem-Boughanmi, N.; Ben-Attia, M.; Dang, P.M.-C.; Souli, A.; El-Benna, J. The NADPH Oxidase Inhibitors Apocynin and Diphenyleneiodonium Protect Rats from LPS-Induced Pulmonary Inflammation. Antioxidants 2023, 12, 770. https://doi.org/10.3390/antiox12030770
Kouki A, Ferjani W, Ghanem-Boughanmi N, Ben-Attia M, Dang PM-C, Souli A, El-Benna J. The NADPH Oxidase Inhibitors Apocynin and Diphenyleneiodonium Protect Rats from LPS-Induced Pulmonary Inflammation. Antioxidants. 2023; 12(3):770. https://doi.org/10.3390/antiox12030770
Chicago/Turabian StyleKouki, Ahmed, Wafa Ferjani, Néziha Ghanem-Boughanmi, Mossadok Ben-Attia, Pham My-Chan Dang, Abdelaziz Souli, and Jamel El-Benna. 2023. "The NADPH Oxidase Inhibitors Apocynin and Diphenyleneiodonium Protect Rats from LPS-Induced Pulmonary Inflammation" Antioxidants 12, no. 3: 770. https://doi.org/10.3390/antiox12030770