
Recent Perspective of Lactobacillus in Reducing Oxidative Stress to Prevent Disease
by

Tingting Zhao
1,2,†, 
Haoran Wang
1,2,†,
Zhenjiang Liu
3,
Yang Liu
2,
DeJi
2,
Bin Li
2,* and
Xiaodan Huang
1,2,* 1
School of Public Health, Lanzhou University, Lanzhou 730033, China
2
Institute of Animal Husbandry and Veterinary, Tibet Academy of Agricultural and Animal Husbandry Sciences, Lhasa 850000, China
3
National Engineering Laboratory for AIDS Vaccine, School of Life Sciences, Jilin University, Changchun 130012, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Antioxidants 2023, 12(3), 769; https://doi.org/10.3390/antiox12030769
Submission received: 16 February 2023
/
Revised: 19 March 2023
/
Accepted: 20 March 2023
/
Published: 21 March 2023
(This article belongs to the Special Issue Antioxidant Activity of Fermented Foods and Food Microorganisms)
Abstract
:During oxidative stress, an important factor in the development of many diseases, cellular oxidative and antioxidant activities are imbalanced due to various internal and external factors such as inflammation or diet. The administration of probiotic Lactobacillus strains has been shown to confer a range of antibacterial, anti-inflammatory, antioxidant, and immunomodulatory effects in the host. This review focuses on the potential role of oxidative stress in inflammatory bowel diseases (IBD), cancer, and liver-related diseases in the context of preventive and therapeutic effects associated with Lactobacillus. This article reviews studies in cell lines and animal models as well as some clinical population reports that suggest that Lactobacillus could alleviate basic symptoms and related abnormal indicators of IBD, cancers, and liver damage, and covers evidence supporting a role for the Nrf2, NF-κB, and MAPK signaling pathways in the effects of Lactobacillus in alleviating inflammation, oxidative stress, aberrant cell proliferation, and apoptosis. This review also discusses the unmet needs and future directions in probiotic Lactobacillus research including more extensive mechanistic analyses and more clinical trials for Lactobacillus-based treatments.
1. Introduction
1.1. Oxidative Stress and Disease
Oxidative stress is associated with the accumulation of excess reactive oxygen and reactive nitrogen species (ROS and RNS, respectively, see Table S1). In cells, ROS/RNS are continuously produced through metabolic processes. Low to moderate ROS levels serve as essential secondary messengers in cell signaling, regulating cell proliferation, differentiation, and migration, and participate in triggering cellular survival mechanisms. However, excessive ROS/RNS can cause oxidative damage to a variety of biomolecules such as unsaturated fatty acids, proteins, and DNA [1]. Endogenous ROS are principally produced in mitochondria, the endoplasmic reticulum, and peroxisomes, with the vast majority of ROS generated by the mitochondrial electron transport chain [2,3]. In addition, numerous enzymes are known to participate in catalyzing the production of endogenous ROS/RNS such as peroxidases, nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX), myeloperoxidase (MPO), nitric oxide synthase (NOS), lipoxygenases (LOXs), and cyclooxygenases (COXs) [4,5]. Beyond these enzymatic pathways for ROS/RNS production, exposure to a wide range of exogenous factors such as trans-fatty acids, iron (Fe), and copper (Cu) in foods, ultraviolet radiation, various drugs and xenobiotics as well as chronic infection and inflammatory diseases can also contribute to inducing ROS accumulation [5,6,7,8].
In humans and other mammals, antioxidant defense systems can largely counteract the effects of ROS/RNS. Endogenous antioxidant systems are generally classified as belonging to either the intracellular enzymatic antioxidants, comprising superoxide dismutase (SOD), catalase (CAT), and other such antioxidant enzymes, or the intracellular non-enzymatic antioxidants, which rely on the activity of small molecules in conjunction with enzymes to neutralize ROS/RON. In the latter system, glutathione functions as a significant antioxidant barrier in the gut, along with three related enzymes: glutathione peroxidase (GPX), glutathione reductase (GSR), and glutathione S-transferase (GST) [1,9]. In addition, the melatonin (MEL) and thioredoxin (Trx) system are well-studied, essential, intracellular antioxidants [10,11]. In addition to the above, there are also some extracellular antioxidants such as vitamin A/C or β-carotene obtained from vegetables and fruits, or flavonoids from certain plants [12,13]. Oxidative stress can occur when ROS/RNS generation exceeds the cellular capacity for scavenging activity of the antioxidant systems or as a result of the dysregulation of antioxidant pathways [5], leading to the damage of various physiological systems in the body. Some examples of the pathological damage related to ROS/RNS accumulation include atherosclerosis in the cardiovascular system; neurodegenerative diseases of the nervous system such as Alzheimer’s disease and Parkinson’s disease, rheumatoid arthritis or systemic lupus erythematosus of the autoimmune system, as well as IBD, stomach cancer, colorectal cancer, and other diseases of the digestive system [14,15].
Alleviating oxidative stress is thus essential for treating many diseases and ensuring systemic health. As our understanding of ROS/RNS and antioxidant mechanisms expands, more precise therapeutic interventions can be developed that focus on disease-related sources and targets of ROS/RNS, some of which are currently entering the clinical trial stages of testing [16]. For instance, free iron is a promising target to control the site and extent of highly aggressive hydroxyl radical generation, which could benefit the treatment of oxidative stress-related diseases [8]. There is also potential for the application of more targeted ROS/RNS inhibitors such as site-specific suppressors of superoxide production (i.e., S1QELs) in quinone-mediated reactions [14,17]. In addition, probiotics such as Lactobacillus and Bifidobacterium may serve as effective complementary therapies that enhance antioxidant defenses via ROS/RNS removal, the inhibition of pro-oxidative enzymes, and the synthesis of antioxidant enzymes.
1.2. Antioxidant Activity of Lactic Acid Bacteria
Lactobacillus, a genus comprising Gram-positive, aerotolerant, rod-shaped, non-sporulating species, is commonly found among microbiota in the gastrointestinal tract and is also frequently isolated from many fermented food products. This genus currently includes 253 published, validated species [18], and accounts for an estimated 6.0% of total bacterial cell numbers in the human duodenum and ~0.3% of all bacteria in the colon [19]. The beneficial and functional properties of Lactobacillus in promoting human health have been extensively documented in recent years. Members of Lactobacillus have been shown to adhere to the intestinal epithelium and produce antimicrobial metabolites such as ethanol, hydrogen peroxide (H2O2), acetic acid, lactic acid, and/or succinic acid, which can antagonize other, potentially pathogenic, bacteria [20]. Some Lactobacillus strains can reportedly enhance cellular immune responses via the activation of macrophages, natural killer (NK) cells, and/or antigen-specific cytotoxic T-lymphocytes, or by stimulating the release of various cytokines. Moreover, some Lactobacillus species could also improve the immune response in gut mucosa by promoting the recruitment of IgA(+) cells [21]. Some Lactobacillus species may also exhibit direct antioxidant activity, and a few species are known to possess oxidative stress resistance genes and proteins pivotal for redox mechanisms such as katA expressed by L. sakei, or thioredoxin antioxidant system proteins expressed by some L. plantarum and L. casei strains [18]. Apart from these effects, Lactobacillus species also produce functional products such as exopolysaccharides (EPS), which participate in mitigating oxidative damage or activating the expression of host transcription factors involved in modulating cellular oxidative stress [22,23].
In recent years, several studies have investigated the relationship between Lactobacillus and oxidative stress in the host. Due to the purported beneficial role of Lactobacillus in human health, research has increasingly focused on the positive impacts of these species in ameliorating oxidative stress and related diseases [18]. Moreover, further in-depth exploration of the role of oxidative stress in disease will provide new perspectives that contribute to improving clinical treatments. This review covers the pathological mechanisms of oxidative stress in inflammatory bowel disease, cancer, and liver-related diseases as well as the preventive and therapeutic effects of Lactobacillus in patients with these diseases.
2. Inflammatory Bowel Disease
2.1. Inflammatory Bowel Disease and Oxidative Stress
Inflammatory bowel disease (IBD) including Crohn’s disease (CD) and ulcerative colitis (UC) is characterized by chronic, non-specific intestinal inflammation. CD can affect the entire alimentary tract from the oral cavity to the anus (mainly the terminal ileum and adjacent colon) and include lesions that appear in a segmental or stochastic (i.e., not continuous) distribution and commonly lead to complications such as abscesses, fistula, and stenosis. In contrast, these lesions are absent in UC, which is characterized by mucosal inflammation and is limited to the colon [24]. Previous studies have shown that individual genetic susceptibility, external environment, gut microbiota, immune response, and oxidative stress are all contributing factors that can be firmly linked to the pathogenesis of IBD [25,26], with lipid peroxidation (LPO) and ROS accumulation, in particular, serving as important causes of oxidative stress in the intestine.
Several points have been proposed regarding the relationship between oxidative stress and IBD. First, researchers have identified several genetic risk loci associated with IBD, and mutations in genes encoding antioxidant/biotransformation enzymes can negatively impact their activity and increase the risk of IBD. For example, case-control studies found that inter-individual polymorphisms related to a C609T conversion in NAD(P)H: quinone oxidoreductase 1 (NQO1), an enzyme involved in inflammation and oxidative stress response, might play a significant role in the development of colon cancer and could influence steroid resistance in UC patients [27,28]. In addition, a polymorphism in the promoter region of the Nuclear factor E2-related factor 2 (Nrf2) gene, which participates in regulating the expression of detoxifying and antioxidant proteins in the intestine, has been associated with UC development in a Japanese population [29]. Second, mucosal immune cells and intestinal epithelial cells (IECs) are involved in oxidative stress in the intestine and have been shown to play a central role in the pathogenesis of IBD. During mucosal inflammation, IECs as well as neutrophils and macrophages, produce superoxide and nitric oxide by activating NOX and inducible nitric oxide synthase (iNOS), respectively, both induced by inflammatory cytokines, eventually leading to the production of more ROS/RNS. The ROS/RNS overload alters the structure and function of the intestinal epithelium, damages cytoskeletal proteins, and accelerates cellular damage through lipid peroxidation, ultimately resulting in barrier destruction [5,30,31]. Third, mutual interactions between gut microbiota and oxidative stress have been identified in IBD. Notably, the composition of gut microbial taxa is altered in CD patients compared to that in healthy controls, characterized by increased relative abundance of Bacteroidetes and decreased Firmicutes. In addition, Enterobacteriaceae, especially E. coli, are enriched in CD patients [32]. ROS/RNS are associated with intestinal dysbiosis. During inflammation, gut microbiota can directly generate ROS/RNS, or indirectly induce the production of excessive ROS/RNS by activating neutrophils or gastrointestinal epithelial cells, stimulating an initial inflammatory response via positive feedback, which leads to further ROS/RNS production, aggravating intestinal oxidative stress and damaging tissues [5,33]. Therefore, restoring the normal structure of gut microbiota has positive implications for the alleviation of inflammation and oxidative stress in IBD. Lifestyle-associated factors can also contribute to gastrointestinal oxidative stress. Although the underlying mechanisms remain unclear, many studies have shown that nicotine intake from smoking can play a dual role in CD and UC, and smoking is a risk factor for the occurrence of CD, but has apparently protective effects in UC, according to epidemiological data [34,35]. Similarly, wine consumption may have various effects in IBD patients. Some chemicals such as polyphenols in red wine appear to confer antioxidant properties [36], whereas the alcohol itself can lead to inflammation and oxidative stress in the liver and intestines, and chronic alcohol consumption leads to increased risk of gastric or colon cancer [37].
At present, treatments for IBD are still based on conventional anti-inflammatory agents (e.g., sulfasalazine and mesalazine, etc.) and immune modulators (e.g., thiopurines and cyclosporine, etc.). However, these drugs have severe side effects [38]. The incidence of UC and CD are higher in regions of Asia with high population density and coastal areas of China also have high rates of IBD, indicating that newer, more effective therapeutic interventions for IBD are urgently needed [39]. The beneficial effects of Lactobacillus in IBD have stimulated considerable research attention that has led to increased the clinical application of probiotic Lactobacillus strains as alternative or complementary therapies [40] (Figure 1).
2.2. The Effects Lactobacillus in Inflammatory Bowel Disease
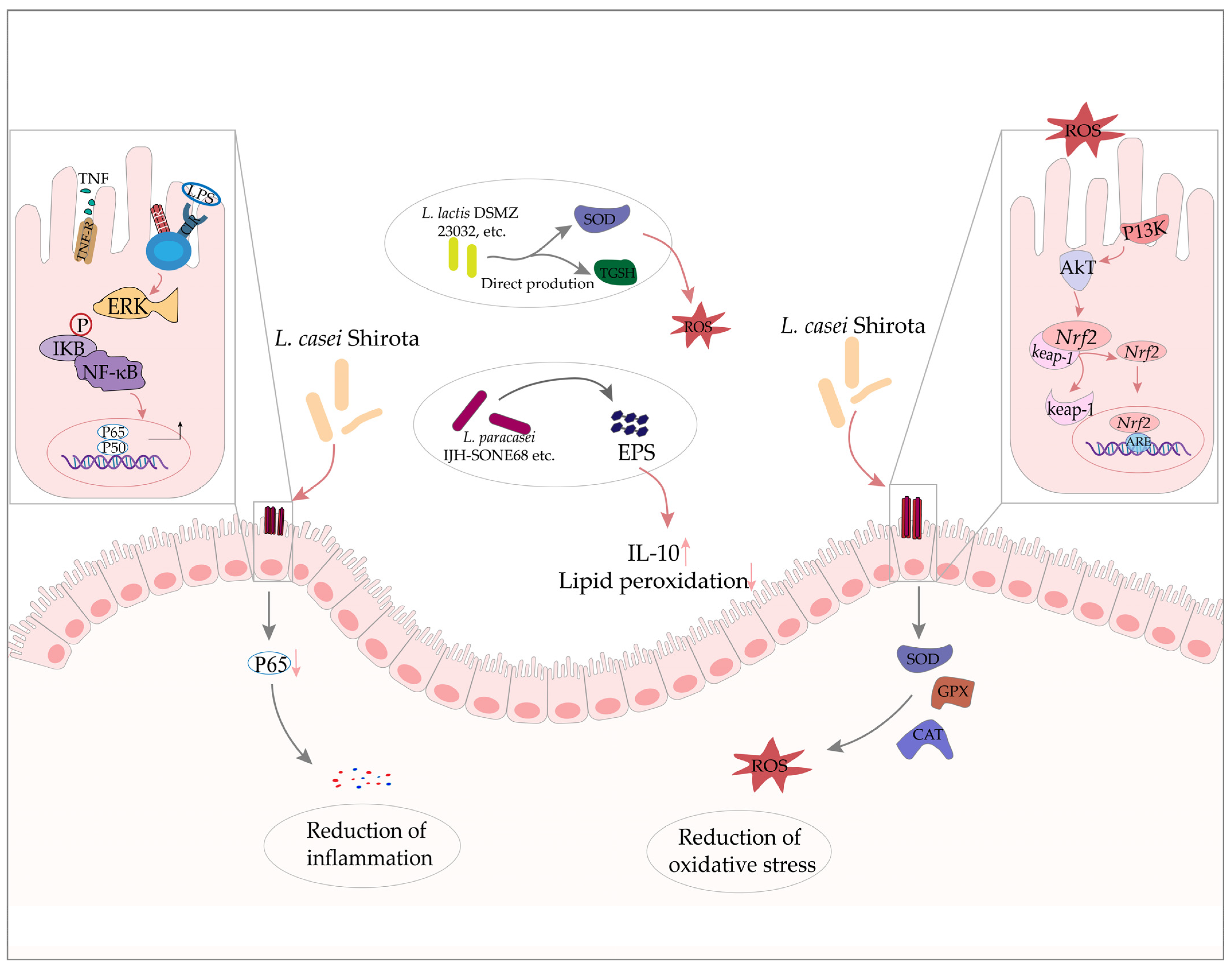
A recent study exploring the effects of EPS-producing probiotic bacteria found that antioxidant enzyme activities were higher and lipid peroxidation was significantly ameliorated in UC model rats treated with probiotics compared with those in untreated UC model rats. More importantly, improvements to oxidative stress were significantly greater in rats administered with a high EPS producing strain than in those treated with a low EPS strain [41]. Similarly, another study showed that administering a novel EPS produced by L. paracasei IJH-SONE68 led to reduced macrophage inflammatory protein 2 mRNA levels, and the increased expression of the anti-inflammatory cytokine, interleukin-10 (IL-10) in dextran sulfate sodium (DSS)-induced UC model mice [42] (Figure 1). These results suggest that EPS should be further examined for application in new treatments for IBD in the future, and Lactobacillus would be among one of the best choices for EPS production.
In the IBD model mice, administration of L. plantarum ZS62 was found to inhibit DSS-induced colonic atrophy. Moreover, mice supplemented with strain ZS62 had lower serum levels of oxidative stress indicators (i.e., Malondialdehyde and MPO) and inflammatory indicators (i.e., IL-1β, IL-6, IL-12, TNF-α, etc.) at both the mRNA and protein levels and elevated protein and mRNA levels of antioxidant enzymes (i.e., CAT, T-SOD) and IL-10 [43]. In addition, Chorawala and colleagues reported that exposure to the cell wall components of L. casei, L. acidophilus, and L. rhamnosus could attenuate lipopolysaccharide (LPS)-induced rats colitis by modulating the inflammatory immune response and oxidative stress [44]. Studies have suggested that Lactobacillus can improve the hosts’ antioxidant level. For example, Erdogan et al. found that feeding fermented Kefir significantly increased the serum total antioxidant status (TAS) levels in Balb/c mice [45]. Similarly, in a healthy volunteer trial (Application No. WO03002131), study subjects who consumed L. fermentum ME-3 in fermented goat milk or who received capsules with L. fermentum ME-3 displayed significantly higher blood total antioxidative activity and TAS than subjects who did not [46]. Furthermore, a pediatric, randomized, placebo-controlled trial demonstrated that a highly concentrated mixture of probiotic bacterial strains (VSL#3) was both safe and effective in maintaining remission among patients with active UC [47]. However, another trial in which CD patients were administered 1 × 109 CFUs of the probiotic strain L. johnsonii LA1 four times daily showed no preventive effects on endoscopic disease recurrence [48].
Assessment of the antioxidant capacity of 34 lactic acid bacteria strains in vitro by Amaretti et al. indicated that strains such as L. lactis DSMZ 23032, L. acidophilus DSMZ 23033, and L. brevis DSMZ 23034 produced relatively high intracellular levels of glutathione and SOD [49], suggesting that LAB might be effective for delivering these antioxidant enzymes to the gut to alleviate oxidative stress in IBD (Figure 1). Furthermore, the co-expression of antioxidant enzyme genes in Lactobacillus through genetic engineering could enhance their antioxidant capacity while also increasing their viability in the host by several-fold, supporting their potential development in therapeutics for IBD [50,51,52].
Finamore et al. showed that pretreatment of the enterocyte-like cell line TC7/human colon carcinoma cell line (Caco-2) with L. casei Shirota (LcS) could prevent membrane barrier disruption and intercellular ROS accumulation, significantly increase the gastrointestinal expression of GPX, and reduce p65 phosphorylation. These findings support the involvement of theNfr2 and Nuclear Factor-Kappa B (NF-κB) pathways in the activation of antioxidant cellular defenses [53] (Figure 1). However, the induction of GPX has also been shown to occur in a Nrf2-independent manner via activation of the transcription factor, STAT3 [54]. In short, some Lactobacillus strains appear to confer positive effects in maintaining intestinal homeostasis via enhanced antioxidant capacity and inflammatory response, although the specific mechanisms through which Lactobacillus can alleviate oxidative stress in IBD remain unclear.
Although Lactobacillus is generally regarded as a safe microorganism, several studies have indicated that certain species may pose risks such as bacteremia and endocarditis [55,56]. Therefore, the application of Lactobacillus for the treatment of IBD raises concerns regarding the presence of virulence genes and antibiotic resistance genes, as their potential transfer to pathogenic microorganisms could constitute a risk [57].
3. Cancer
3.1. ROS and Cancer
Cancer is among the leading causes of death worldwide. According to the Global Cancer Statistics, approximately 19.3 million new cancer cases and nearly 10 million deaths from cancer were reported in 2020 [58]. Tumorigenesis may result from a combination of internal and external factors, some of which influence cancer progression and metastasis through increased ROS production [59]. The generation of ROS in cancer cells is primarily mediated by oxidative phosphorylation in the electron transport chain on the mitochondrial inner membrane. In addition, transition metals such as iron can also generate ROS non-enzymatically via a Fenton reaction, while exposure to various external factors such as air pollutants, radiation, foods, or drugs, etc. can also exogenously induce ROS production [60]. In general, the aberrant and uncontrolled proliferation of cancer cells requires large amounts of ATP as an energy supply, and the resulting metabolic upregulation leads to elevated accumulation of ROS. However, cancer cells have a characteristically high antioxidant capacity due to the Nrf2-mediated activation of various antioxidant response elements (AREs) that can restrict ROS accumulation to levels compatible with cellular biological functions, albeit still higher than that in normal cells [61].
At low to moderate levels, ROS may promote abnormal cell proliferation and differentiation by inducing DNA mutations These mutations mainly include DNA double strand breaks accompanied by abnormalities in their associated repair pathways, or by generating 8-oxo-7-hydrodeoxyguanosine (8-oxodG), which is recognized as an indicator of oxidative damage to DNA [62,63]. Other work has linked Fe-catalyzed ROS accumulation to the development of some cancers. For instance, a multivariate statistical analysis indicated that iron depletion therapy, either through phlebotomy or by maintaining a low-iron diet, could significantly lower the risk of hepatocellular carcinoma compared to that in untreated patients [64,65].
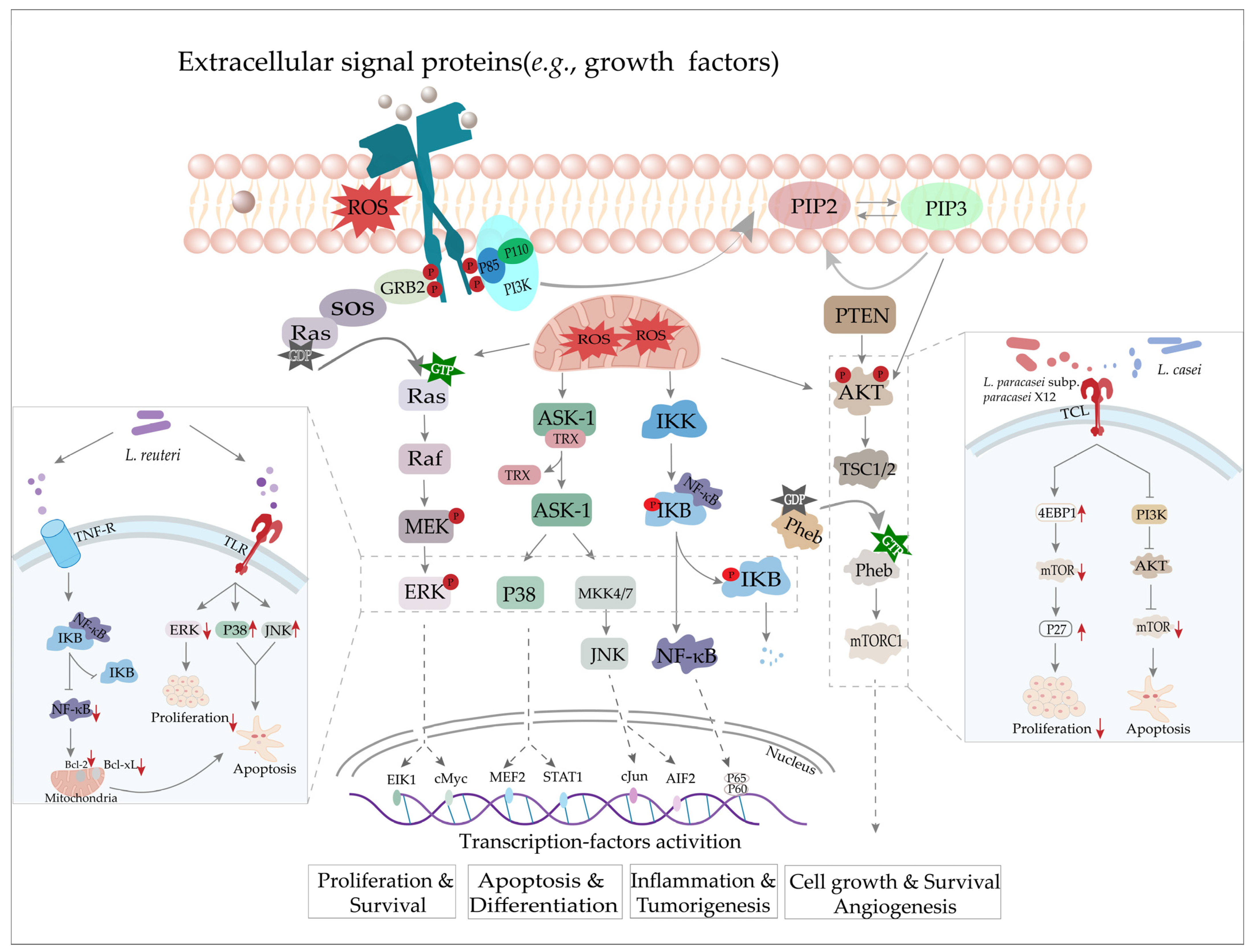
In addition, ROS may also contribute to tumorigenesis through their function as signal molecules (Figure 2). Increased ROS production in cells can induce an enzymatic cascade reaction that results in the activation of extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38 mitogen-activated protein kinases (MAPK), all of which are linked to tumor cell growth and survival [66,67]. Weinberg and colleagues observed that mitochondrial ROS are essential for Kras-mediated tumorigenesis in a murine model of lung carcinoma via ERK-MAPK signaling [59,68]. The phosphatidylinositol-3-kinase (PI3K)/phosphatase and tensin homolog deleted on chromosome 10 (PTEN) signaling pathway is also important for tumorigenesis and cancer metastasis. ROS can stimulate PI3K/protein kinase B (AKT) signaling through the oxidation of a cysteine thiol group on various phosphatases (e.g., PTEN, PTP1B, PP2A) and contribute to dysregulation in a wide range of human cancers (e.g., endometrial, breast, thyroid, and prostate cancers) [69,70]. In addition to the above pathways, NF-κB is increasingly recognized as a key player in many steps of cancer development and progression. Treatment of breast carcinoma cells with IL-1β, TNFα, or sodium arsenite generates H2O2 and O2−, which in turn activate NF-κB and enhance uncontrolled cellular growth [71,72]. Constitutive activation of Nrf2 has been observed in various human cancers such as those affecting the lungs, breasts, and skin [73,74]. In addition, deregulation of the Nrf2–Keap1 pathway has been reported in multiple cancer cells, leading to drug resistance, genomic instability, resistance to apoptosis, metastasis, and metabolic [75].
However, high levels of ROS have contrary effects on cancer, promoting cell death and severe cellular damage. Some chemotherapeutic drugs or cytotoxic agents such as anthracyclines, platinum-based drugs (e.g., cisplatin, carboplatin), and other alkylating drugs exert therapeutic effects by stimulating the high intracellular production of ROS [76,77]. In addition, excessive ROS levels are also positively correlated with apoptosis through the ASK-1/JNK/p38 signaling cascade induced via the H2O2-mediated oxidation of cysteine residues on TRX, which leads to the dissociation of ASK-1, the subsequent activation of JNK and p38 signaling, and ultimately apoptosis [59,78] (Figure 2).
Thus, ROS can serve as a “double-edged sword”, both causing and suppressing cancer [79], and current studies suggest that modulating the oxidative stress response might be an effective, new potential approach for cancer therapies.
3.2. The Role of Lactobacillus in Cancer
Drug resistance is a serious problem in cancer chemotherapy, and thus effective alternative therapeutic options are desperately needed to improve treatment response in many cancer patients. To address this issue, an increasing number of studies has investigated the potential antitumor activity of Lactobacillus and provide evidence supporting its further exploration and development for use in candidate anti-tumor therapies [80,81].
In many cancers, abnormal cell proliferation is tightly linked to the suppression of apoptosis, and many drugs achieve therapeutic efficacy by promoting apoptosis in cancer cells. Notably, Lactobacillus has been shown to induce apoptosis [82,83]. For example, oral pretreatment with the probiotic strain, L. acidophilus NCFM, was found to stimulate apoptosis in CT-26 cell dorsal flank xenograft tumors or in segmental orthotopic colon cancer in mice [84]. Another study showed that the apoptosis levels increased in the LS513 colorectal cancer cells exposed to a combination of the L. acidophilus and L. casei strains in the presence of 5-fluorouracil (5-FU) in a dose-dependent manner. This effect was potentially due to the faster activation of caspase-3 and the downregulation of the p21 protein [85]. Similarly, Altonsy et al. found that the L. rhamnosus GG strain (LGG) could induce caspase-9 and caspase-3 activation, promoting apoptosis in the Caco-2 colon cancer cells [86]. Homogenates of strain LGG were also found to induce the apoptosis pathways in HGC-27 human gastric cancer cells via reduced expression of the anti-apoptotic protein, Bcl-2, and elevated expression of the pro-apoptotic protein, Bax [87]. Similarly, ubiquitination of IKB was suppressed in human chronic myeloid leukemia-derived cells (KBM-5 cells) treated with L. reuteri ATCC PTA 6475, resulting in the downregulation of the NF-κB-dependent gene products (e.g., Bcl-2, Bcl-xL), while pre-treatment with L. reuteri enhanced JNK and p38 phosphorylation, but suppressed ERK1/2 signaling in cells, increasing the apoptosis levels and inhibiting proliferation, respectively [88]. In addition, Hwang and colleagues confirmed that extracts of L. casei could induce apoptosis in gastric cancer cells by suppressing PI3K/AKT signaling [89] (Figure 2). Thus, numerous studies have shown that Lactobacillus strains can regulate apoptosis in cancer cells via the modulation of endogenous signaling pathways or exogenous cues.
Lactobacillus strains and their metabolites (i.e., components of the cell wall and cytoplasm) have been documented to inhibit cancer cell proliferation. For instance, Orlando and colleagues investigated the anti-proliferative effects of the cell wall and cytoplasmic fractions of strain LGG on the HGC-27 cells and DLD-1 human colonic adenocarcinoma cells. They found that both HGC-27 and DLD-1 cells were resistant to the bacterial cell wall fraction, whereas treatment with the cytoplasmic fraction resulted in apparently reduced proliferation [90]. 4E-Binding protein 1 (4EBP1) is a transcriptional inhibitor in the downstream of the mTOR signaling pathway. Treatment with L. paracasei subsp. paracasei X12 could induce cell cycle arrest at G1 in the HT-29 cells through the mTOR-4EBP1-p27 signaling pathway, and the mRNA expression of 4EBP1 was increased several folds after treatment by the X12 strain [91]. Hence, blocking hyperactivation of the PI3K/AKT/mTOR signaling pathway has emerged as a plausible target for cancer therapies due to its involvement in cell growth and proliferation (Figure 2).
Polyamines including putrescine, spermidine, and spermine, etc. can serve as nutrient substrates for cancer cell growth and may thus play an important role in cancer cell proliferation. In gastric cancer cells, probiotics such as Lactobacillus strains can potentially act as anti-neoplastic agents to inhibit proliferation by modulating polyamine contents or function [92]. L. salivarius strains FP25 and FP35, isolated from infant feces, can directly adhere to cancer cells, triggering the biosynthesis of short-chain fatty acids (SCFAs), especially butyric and propionic acids, resulting in the inhibition of colon cancer cell proliferation [93]. In addition, some studies have reported that EPS secreted by Lactobacillus might also exert anti-proliferative effects on cancer cells [94,95,96].
A growing body of evidence supports that Lactobacillus strains can also provide antioxidant functions to prevent oxidative stress-related tumor development. A recently identified probiotic strain, L. salivarius REN, isolated from fecal samples of centenarians by Zhang and colleagues, was found to inhibit 4NQO (4-nitroquioline 1-oxide)-induced oral cancer in rats by reducing oxidative DNA damage and downregulating COX-2 expression [97]. In addition, L. reuteri is capable of regulating intestinal metabolites to promote a growth-repressive effect on cancer and reduce the oxidation level in mice with colorectal cancer [98]. In rats with colon cancer induced by 1,2-dimethylhydrazine (DMH), treatment with L. plantarum AS1 led to significant improvements in lipid peroxidation and antioxidant activities in the colon and plasma, and showed promising antioxidant effects in vitro [99]. Choi et al. found that heat-killed (HK) cells and soluble polysaccharide components of L. acidophilus 606 exhibited potent antioxidative activity, and further reported that the soluble polysaccharide fraction could induce apoptosis in HT-29 cells in vitro, suggesting that soluble polysaccharides of Lactobacillus strains could be explored as possible candidate anticancer agents [100]. Overall, many in vivo and in vitro studies support that Lactobacillus can reduce oxidative damage, attenuating DNA mutation and cancer development.
Lactobacillus can also exert immunomodulatory and cytotoxic anticancer effects. For instance, L. casei strains 9018 and LcS could activate NK cells by inducing the production of cytokines such as IL-12 and IFN-γ, and highly active NK cells are reportedly associated with a lower risk of cancer development [101,102,103]. In addition, L. acidophilus 36YL shows no apparent cytotoxicity toward normal cells, but displays high toxicity toward cancer cell lines such as HeLa and HT-29 [104].
Although several in vivo and in vitro studies have reported that the administration of various Lactobacillus strains leads to the inhibition of cancer development and progression through pro-apoptotic, antioxidant, anti-proliferative, or immunomodulatory pathways, clinical population trials and epidemiological studies are still lacking. Indeed, some issues should be carefully considered in the administration of probiotic Lactobacillus strains for the treatment and prevention of cancers. In particular, the potential for undesirable side effects resulting from host–bacteria interactions, individual resistance to particular strains, or variability in the benefits conferred by different Lactobacillus strains, all warrant further experimental investigation [80].
4. Oxidative Stress-Related Liver Disease
4.1. Oxidative Stress and Liver Disease
Alcoholic Liver Disease (ALD): Long-term and excessive alcohol consumption is the leading cause of ALD, which includes a broad clinical-histological spectrum ranging from simple steatosis and hepatitis, to liver cirrhosis and hepatocellular carcinoma [105]. Disruption of intestinal microbiota, increased intestinal permeability, inflammatory response, liver oxidative stress, and lipid accumulation are all associated with the progression of ALD [106]. In particular, the role of oxidative stress in the occurrence and development of ALD is currently the focus of increasing research attention. Alcohol is mainly metabolized in the liver, leading to elevated ROS/RNS production, weakening antioxidant defense systems, and finally resulting in oxidative stress in the liver due to excess ROS/RNS. It should be noted that oxidative stress in the liver has been reported to play a key role in hepatocellular carcinogenesis [107].
Non-alcoholic fatty liver disease (NAFLD): NAFLD is mainly due to the accumulation of lipids in the liver, or insulin resistance caused by obesity, type 2 diabetes, and lipid metabolism disorders, among other conditions, resulting in hepatic steatosis. In addition, lipid peroxidation, oxidative stress, and pro-inflammatory cytokines are also involved, culminating in hepatocyte infiltration and necrosis. Although NAFLD has multiple clinical manifestations, most patients present with only simple steatosis [108,109].
4.2. The Preventive Effects of Lactobacillus
Preventive effects on ALD: Lactobacillus has been tested in both the treatment and prevention of ALD. Forsyth et al. found that rats fed with alcohol plus L. rhamnosus GG (ALC + LGG) had significantly lower severity of alcoholic steatohepatitis (ASH) than rats fed with alcohol alone, while strain LGG was also associated with significantly less alcohol-induced leakiness in the gut, oxidative stress, and inflammation in both the intestinal and liver tissues [110]. Another study comparing treatments of Lactobacillus strains with the traditional Chinese medicine Hu-Gan-Pian in a rat model of alcohol-induced enteric dysbiosis found that L. rhamnosus CCFM1107 administration could rescue dysbiosis and inhibit the characteristic increase in serum aminotransferase and endotoxin as well as triglyceride (TG) and cholesterol (CHO) levels in the serum and liver. In addition to these findings, the oxidation levels, indicated by the malondialdehyde contents, were lower, while the antioxidant enzyme (GSH/GSH-Px/SOD) levels were elevated in rats treated with L. rhamnosus CCFM1107 [111].
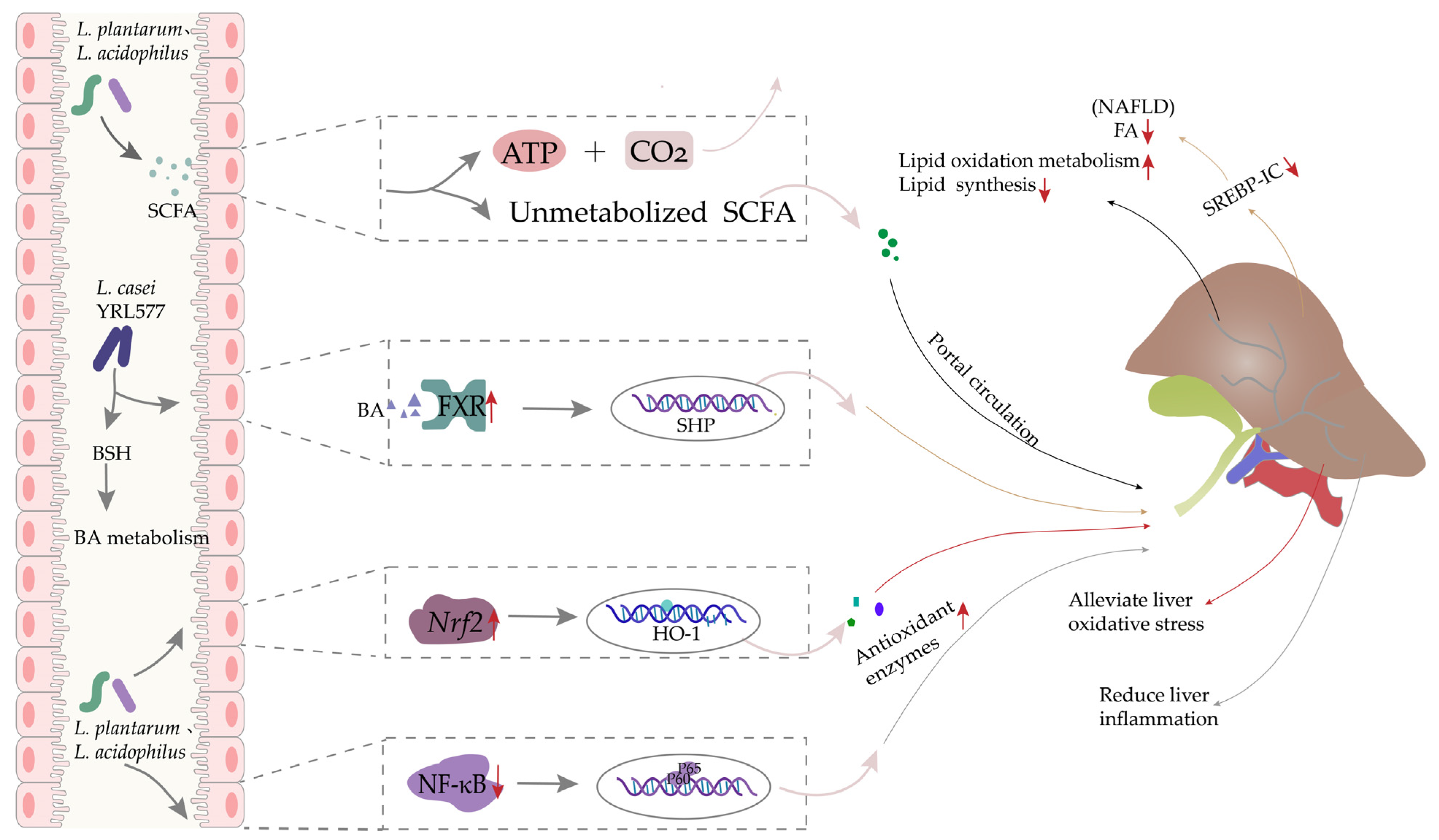
In mice with alcoholic subacute liver injury, treatment with L. plantarum ZS62, isolated from naturally fermented yogurt, could relieve morphological abnormalities in hepatocytes and reduce markers of liver damage (e.g., AST/ALT/hyaluronidase, etc.). Furthermore, inflammation-related genes were significantly downregulated while lipid- and oxidative-metabolism genes were upregulated, implying that L. plantarum ZS62 could potentially function as a beneficial prophylactic supplement for people with frequent alcohol consumption [112]. In a study examining alcoholic liver injury using a zebrafish model, L. plantarum was shown to provide some protective effects via the transcriptional activation of the Keap-Nrf2-ARE signal pathway [113]. In work by Li and coworkers investigating the potential mechanism and effects of Lactobacillus in alleviating ALD symptoms in mice, treatment with a mixture of L. plantarum KLDS1.0344 and L. acidophilus KLDS1.0901 resulted in higher SCFA production by the gut microbiota. These SCFAs could potentially inhibit alcohol-induced lipid accumulation, oxidative stress, and inflammation in the liver through hepatic-intestinal circulation, most likely via activation of the AMPK, Nrf2, and TLR4/NF-κB pathways [106] (Figure 3). Collectively, these studies show that Lactobacillus strains could provide preventive effects against mild ALD, but long-term experiments in animal models are necessary to determine whether these strains could also alleviate end-stage ALD.
Preventive effects of Lactobacillus in NAFLD: One study in rats with induced NAFLD showed that administering a Lactobacillus-rich mixture of probiotic strains including L. acidophilus, L. casei, and L. reuteri, could significantly reverse hepatic and blood triglyceride concentrations and blood glucose levels while suppressing markers of oxidative stress in liver tissue [114]. Zhang et al. isolated L. casei YRL577, which exhibited high bile acid hydrolase (BAH) activity that could regulate bile acid metabolism by increasing the transcription of fibroblast growth factor 15 (FGF15), while downregulating the host mRNA levels of Na-dependent bile acid transporter (ASBT). In addition, the YRL577 strain reduced the de novo synthesis of fatty acids (FA) via the FXR/SHP/SREBP1c pathway. These regulatory effects could possibly contribute to alleviating NAFLD in C57BL/6 mice [109] (Figure 3). Furthermore, a population-based study confirmed that dietary supplementation with probiotic strains Bifidobacterium longum and L. acidophilus could improve some anthropometric, inflammatory, and oxidative indices in patients with NAFLD [115].
Other studies have also examined the mechanisms by which Lactobacillus could affect NAFLD. Nrf2 is a known protective factor against liver damage, and has been implicated in the pathogenesis of chronic conditions such as NAFLD. Notably, a recent study has identified 5-methoxyindoleacetic acid, produced by the human commensal LGG, which can potently activate Nrf2 in both Drosophila liver analog and the murine liver. This activation of Nrf2 has been shown to protect against two models of oxidative liver injury, namely, acetaminophen overdose and acute ethanol toxicity [116]. Chen et al. found that treatment with L. mali APS1 led to reduced hepatic lipid accumulation via regulating the SIRT-1/PGC-1α/SREBP-1 pathway, and promoted hepatic antioxidant activity through the Nrf2/HO-1 pathway [117]. Another study reported that L. plantarum NA136 could improve NAFLD by regulating fatty acid metabolism and oxidative stress defense pathways via AMPK and Nrf2 signaling, respectively [118]. Similarly, another report suggested that L. plantarum NCU116 might ameliorate NAFLD by downregulating lipogenesis while promoting the upregulation of lipolysis and fatty acid oxidation-related gene expression [119]. However, the current body of literature examining the preventive or therapeutic effects of Lactobacillus on NAFLD remains limited. Furthermore, most of the available studies have examined mechanisms related to the regulation of lipid metabolism and the alleviation of oxidative stress, although other mechanisms are likely involved in the impacts of Lactobacillus on NAFLD.
Lactobacillus strains prevent other types of liver damage: In an animal model of carbon tetrachloride (CCl4)-induced liver fibrosis, L. plantarum HFY15 was found to alleviate liver injury through antioxidant, anti-inflammatory, and anti-apoptotic pathways [120]. In addition, selenium-enriched probiotics (SP) might also influence pro- and anti-apoptosis pathways, activate the SIRT1 signaling pathway, or attenuate MAPK signaling, leading to reduced liver fibrosis [121,122]. Lactobacillus strains were also shown to alleviate liver injuries caused by other drugs such as D-galactose (D-Gal)/LPS-induced acute liver injury (ALI) in mice or liver toxicity caused by deoxynivalenol (DON), an extremely common environmental pollutant [123,124] (Table 1).
5. Conclusions and Prospects
In summary, oxidative damage accompanies inflammation, cancer development, and liver injury, whereas the administration of probiotic Lactobacillus strains shows potential for modulating oxidative stress in cells and tissues throughout the human body, especially in the gastrointestinal tract [18]. Treatment with probiotic Lactobacillus supplements can also reportedly prevent or ameliorate IBD by downregulating the expression of inflammatory factors while promoting an increase in related antioxidant enzymes, possibly through the activation of various host signaling pathways.
In cancers, it remains largely controversial as to whether targeting ROS, either by antioxidant supplements or chemical/genetic inhibition, is clinically beneficial or detrimental for cancer treatment. Several studies in animal models support the tumor suppressive effects of Lactobacillus strains, although it also remains uncertain whether these effects are the result of directly alleviating host ROS or due to stimulating the host antioxidant systems. Further mechanistic insights are therefore necessary to better understand the role of ROS in different types of cancer and to develop more effective, targeted probiotic anti-cancer therapies [60]. Treatment with Lactobacillus strains has also been shown to confer prophylactic or therapeutic effects, alleviating inflammation and oxidative damage in ALD, NAFLD, and other types of liver injury in vitro and in vivo.
Since the large majority of the above studies are based on animal and cellular models, multi-level, multi-center population-based trials such as prospective epidemiological studies are necessary for a robust assessment of the effectiveness of probiotic treatments in IBD, cancers, and liver diseases.
In addition, Lactobacillus species share conserved microbe-associated molecular patterns (MAMPs) with other Gram-positive bacteria such as cell wall polysaccharides and lipoteichoic acids, which can be recognized by the human immune system and elicit a host inflammatory response. Advances in probiotic development will thus focus on the discovery or engineering of Lactobacillus strains with lower MAMP-induced immunogenic effects, but with a high antioxidant capacity to maximize their therapeutic benefits in different diseases.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/antiox12030769/s1, Table S1: Abbreviations.
Author Contributions
Conceptualizing the content of manuscript, X.H. and B.L.; Writing—all original draft preparation, T.Z. and H.W.; Revising entire manuscript draft, X.H. Supervising, reviewing and editing final version of article, B.L., Z.L., Y.L. and D. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by the Key Research and Development Program in Tibet Autonomous Region (XZ202201ZY0004N), the Science and Technology Program of Gansu Province (22JR5RA463), and the Special Item of Regional Collaborative Innovation in Tibet Autonomous Region (QYXTZX-LS2021-01).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article or Supplementary Material.
Acknowledgments
We gratefully acknowledge Isaac V. Greenhut, ELS for advice with the language and editorial assistance in manuscript preparation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Alzoghaibi, M.A. Concepts of oxidative stress and antioxidant defense in Crohn’s disease. World J. Gastroenterol. 2013, 19, 6540–6547. [Google Scholar] [CrossRef]
- Li, D.; Ding, Z.; Du, K.; Ye, X.; Cheng, S. Reactive Oxygen Species as a Link between Antioxidant Pathways and Autophagy. Oxidative Med. Cell Longev. 2021, 2021, 5583215. [Google Scholar] [CrossRef]
- Handy, D.E.; Loscalzo, J. Redox regulation of mitochondrial function. Antioxid. Redox Signal. 2012, 16, 1323–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swindle, E.J.; Metcalfe, D.D. The role of reactive oxygen species and nitric oxide in mast cell-dependent inflammatory processes. Immunol. Rev. 2007, 217, 186–205. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Wang, Z.; Zhang, J. Pathomechanisms of Oxidative Stress in Inflammatory Bowel Disease and Potential Antioxidant Therapies. Oxid. Med. Cell Longev. 2017, 2017, 4535194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteng, A.B.; Kersten, S. Mechanisms of Action of trans Fatty Acids. Adv. Nutr. 2020, 11, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: An essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [Green Version]
- Galaris, D.; Barbouti, A.; Pantopoulos, K. Iron homeostasis and oxidative stress: An intimate relationship. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118535. [Google Scholar] [CrossRef]
- Iborra, M.; Moret, I.; Rausell, F.; Bastida, G.; Aguas, M.; Cerrillo, E.; Nos, P.; Beltrán, B. Role of oxidative stress and antioxidant enzymes in Crohn’s disease. Biochem. Soc. Trans. 2011, 39, 1102–1106. [Google Scholar] [CrossRef] [Green Version]
- Bonnefont-Rousselot, D.; Collin, F. Melatonin: Action as antioxidant and potential applications in human disease and aging. Toxicology 2010, 278, 55–67. [Google Scholar] [CrossRef]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.E.; Khodr, H.; Hider, R.C.; Rice-Evans, C.A. Structural dependence of flavonoid interactions with Cu2+ ions: Implications for their antioxidant properties. Biochem. J. 1998, 330, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Hengstermann, S.; Valentini, L.; Schaper, L.; Buning, C.; Koernicke, T.; Maritschnegg, M.; Buhner, S.; Tillinger, W.; Regano, N.; Guglielmi, F.; et al. Altered status of antioxidant vitamins and fatty acids in patients with inactive inflammatory bowel disease. Clin. Nutr. 2008, 27, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Chiurchiù, V.; Maccarrone, M. Chronic inflammatory disorders and their redox control: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 15, 2605–2641. [Google Scholar] [CrossRef]
- Elbatreek, M.H.; Pachado, M.P.; Cuadrado, A.; Jandeleit-Dahm, K.; Schmidt, H. Reactive Oxygen Comes of Age: Mechanism-Based Therapy of Diabetic End-Organ Damage. Trends Endocrinol. Metab. TEM 2019, 30, 312–327. [Google Scholar] [CrossRef] [Green Version]
- Banba, A.; Tsuji, A.; Kimura, H.; Murai, M.; Miyoshi, H. Defining the mechanism of action of S1QELs, specific suppressors of superoxide production in the quinone-reaction site in mitochondrial complex I. J. Biol. Chem. 2019, 294, 6550–6561. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Olejar, K.J.; On, S.L.W.; Chelikani, V. The Potential of Lactobacillus spp. for Modulating Oxidative Stress in the Gastrointestinal Tract. Antioxidants 2020, 9, 610. [Google Scholar] [CrossRef]
- Heeney, D.D.; Gareau, M.G.; Marco, M.L. Intestinal Lactobacillus in health and disease, a driver or just along for the ride? Curr. Opin. Biotechnol. 2018, 49, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Annuk, H.; Shchepetova, J.; Kullisaar, T.; Songisepp, E.; Zilmer, M.; Mikelsaar, M. Characterization of intestinal lactobacilli as putative probiotic candidates. J. Appl. Microbiol. 2003, 94, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, R.; Shah, N.P. Immune system stimulation by probiotic microorganisms. Crit. Rev. Food Sci. Nutr. 2014, 54, 938–956. [Google Scholar] [CrossRef] [PubMed]
- Djavaheri-Mergny, M.; Javelaud, D.; Wietzerbin, J.; Besançon, F. NF-kappaB activation prevents apoptotic oxidative stress via an increase of both thioredoxin and MnSOD levels in TNFalpha-treated Ewing sarcoma cells. FEBS Lett. 2004, 578, 111–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Evivie, S.E.; Lu, J.; Jiao, Y.; Wang, C.; Li, Z.; Liu, F.; Huo, G. Lactobacillus helveticus KLDS1.8701 alleviates d-galactose-induced aging by regulating Nrf-2 and gut microbiota in mice. Food Funct. 2018, 9, 6586–6598. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y. Inflammatory bowel disease: Pathogenesis. World J. Gastroenterol. 2014, 20, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Ramos, G.P.; Papadakis, K.A. Mechanisms of Disease: Inflammatory Bowel Diseases. Mayo Clin. Proc. 2019, 94, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Begleiter, A.; Hewitt, D.; Maksymiuk, A.W.; Ross, D.A.; Bird, R.P. A NAD(P)H:quinone oxidoreductase 1 polymorphism is a risk factor for human colon cancer. Cancer Epidemiol. Biomark. Prev. 2006, 15, 2422–2426. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, T.; Yoshino, J.; Inui, K.; Wakabayashi, T.; Kobayashi, T.; Watanabe, S.; Hayashi, S.; Hirokawa, Y.; Shiraishi, T.; Yamamoto, T.; et al. Involvement of NAD(P)H:quinone oxidoreductase 1 and superoxide dismutase polymorphisms in ulcerative colitis. DNA Cell Biol. 2009, 28, 625–631. [Google Scholar] [CrossRef]
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Yoshioka, D.; Okubo, M.; Sakata, M.; et al. Nrf2 gene promoter polymorphism is associated with ulcerative colitis in a Japanese population. Hepato-Gastroenterol. 2008, 55, 394–397. [Google Scholar]
- Banan, A.; Fields, J.Z.; Decker, H.; Zhang, Y.; Keshavarzian, A. Nitric oxide and its metabolites mediate ethanol-induced microtubule disruption and intestinal barrier dysfunction. J. Pharmacol. Exp. Ther. 2000, 294, 997–1008. [Google Scholar]
- Pastorelli, L.; De Salvo, C.; Mercado, J.R.; Vecchi, M.; Pizarro, T.T. Central role of the gut epithelial barrier in the pathogenesis of chronic intestinal inflammation: Lessons learned from animal models and human genetics. Front. Immunol. 2013, 4, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, E.K.; Kamm, M.A.; Teo, S.M.; Inouye, M.; Wagner, J.; Kirkwood, C.D. Recent advances in characterizing the gastrointestinal microbiome in Crohn’s disease: A systematic review. Inflamm. Bowel Dis. 2015, 21, 1219–1228. [Google Scholar] [PubMed] [Green Version]
- Handa, O.; Naito, Y.; Yoshikawa, T. Helicobacter pylori: A ROS-inducing bacterial species in the stomach. Inflamm. Res. 2010, 59, 997–1003. [Google Scholar] [CrossRef]
- Nakajima, A.; Shibuya, T.; Sasaki, T.; Lu, Y.J.; Ishikawa, D.; Haga, K.; Takahashi, M.; Kaga, N.; Osada, T.; Sato, N.; et al. Nicotine Oral Administration Attenuates DSS-Induced Colitis Through Upregulation of Indole in the Distal Colon and Rectum in Mice. Front. Med. 2021, 8, 789037. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Ko, J.K.; Mei, Q.B.; Cho, C.H. Aggravating effect of cigarette smoke exposure on experimental colitis is associated with leukotriene B4 and reactive oxygen metabolites. Digestion 2001, 63, 180–187. [Google Scholar] [CrossRef]
- Rodrigo, R.; Miranda, A.; Vergara, L. Modulation of endogenous antioxidant system by wine polyphenols in human disease. Clin. Chim. Acta 2011, 412, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Na, H.K.; Lee, J.Y. Molecular Basis of Alcohol-Related Gastric and Colon Cancer. Int. J. Mol. Sci. 2017, 18, 1116. [Google Scholar] [CrossRef] [Green Version]
- Moon, W.; Loftus, E.V. Review article: Recent advances in pharmacogenetics and pharmacokinetics for safe and effective thiopurine therapy in inflammatory bowel disease. Aliment. Pharmacol. Ther. 2016, 43, 863–883. [Google Scholar] [CrossRef] [Green Version]
- Ng, S.C.; Kaplan, G.G.; Tang, W.; Banerjee, R.; Adigopula, B.; Underwood, F.E.; Tanyingoh, D.; Wei, S.; Lin, W.; Lin, H.; et al. Population Density and Risk of Inflammatory Bowel Disease: A Prospective Population-Based Study in 13 Countries or Regions in Asia-Pacific. Am. J. Gastroenterol. 2019, 114, 107–115. [Google Scholar] [CrossRef]
- Kim, E.S.; Tae, C.H.; Jung, S.A.; Park, D.I.; Im, J.P.; Eun, C.S.; Yoon, H.; Jang, B.I.; Ogata, H.; Fukuhara, K.; et al. Perspectives of East Asian patients and physicians on complementary and alternative medicine use for inflammatory bowel disease: Results of a cross-sectional, multinational study. Intest. Res. 2022, 20, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Sengül, N.; Işık, S.; Aslım, B.; Uçar, G.; Demirbağ, A.E. The effect of exopolysaccharide-producing probiotic strains on gut oxidative damage in experimental colitis. Dig. Dis. Sci. 2011, 56, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Noda, M.; Danshiitsoodol, N.; Kanno, K.; Uchida, T.; Sugiyama, M. The Exopolysaccharide Produced by IJH-SONE68 Prevents and Ameliorates Inflammatory Responses in DSS-Induced Ulcerative Colitis. Microorganisms 2021, 9, 2243. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Ning, Y.; Hu, J.; Wang, Z.; Chen, X.; Zhao, X. The Preventive Effect of ZS62 on DSS-Induced IBD by Regulating Oxidative Stress and the Immune Response. Oxidative Med. Cell. Longev. 2021, 2021, 9416794. [Google Scholar] [CrossRef]
- Chorawala, M.R.; Chauhan, S.; Patel, R.; Shah, G. Cell Wall Contents of Probiotics (Lactobacillus species) Protect Against Lipopolysaccharide (LPS)-Induced Murine Colitis by Limiting Immuno-inflammation and Oxidative Stress. Probiotics Antimicrob. Proteins 2021, 13, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, F.S.; Ozarslan, S.; Guzel-Seydim, Z.B.; Kök Taş, T. The effect of kefir produced from natural kefir grains on the intestinal microbial populations and antioxidant capacities of Balb/c mice. Food Res. Int. 2019, 115, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Songisepp, E.; Kals, J.; Kullisaar, T.; Mändar, R.; Hütt, P.; Zilmer, M.; Mikelsaar, M. Evaluation of the functional efficacy of an antioxidative probiotic in healthy volunteers. Nutr. J. 2005, 4, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miele, E.; Pascarella, F.; Giannetti, E.; Quaglietta, L.; Baldassano, R.N.; Staiano, A. Effect of a probiotic preparation (VSL#3) on induction and maintenance of remission in children with ulcerative colitis. Am. J. Gastroenterol. 2009, 104, 437–443. [Google Scholar]
- Marteau, P.; Lémann, M.; Seksik, P.; Laharie, D.; Colombel, J.F.; Bouhnik, Y.; Cadiot, G.; Soulé, J.C.; Bourreille, A.; Metman, E.; et al. Ineffectiveness of Lactobacillus johnsonii LA1 for prophylaxis of postoperative recurrence in Crohn’s disease: A randomised, double blind, placebo controlled GETAID trial. Gut 2006, 55, 842–847. [Google Scholar] [CrossRef]
- Amaretti, A.; di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant properties of potentially probiotic bacteria: In vitro and in vivo activities. Appl. Microbiol. Biotechnol. 2013, 97, 809–817. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; del Carmen, S.; Miyoshi, A.; Azevedo, V.; Sesma, F.; Langella, P.; Bermudez-Humaran, L.G.; Watterlot, L.; Perdigon, G.; de Moreno de LeBlanc, A. Use of superoxide dismutase and catalase producing lactic acid bacteria in TNBS induced Crohn’s disease in mice. J. Biotechnol. 2011, 151, 287–293. [Google Scholar] [CrossRef]
- Del Carmen, S.; de Moreno de LeBlanc, A.; Martin, R.; Chain, F.; Langella, P.; Bermudez-Humaran, L.G.; LeBlanc, J.G. Genetically engineered immunomodulatory Streptococcus thermophilus strains producing antioxidant enzymes exhibit enhanced anti-inflammatory activities. Appl. Environ. Microbiol. 2014, 80, 869–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, H.; Zhai, Z.; Yin, S.; Luo, Y.; Han, B.; Hao, Y. Coexpression of the superoxide dismutase and the catalase provides remarkable oxidative stress resistance in Lactobacillus rhamnosus. J. Agric. Food Chem. 2011, 59, 3851–3856. [Google Scholar] [CrossRef]
- Finamore, A.; Ambra, R.; Nobili, F.; Garaguso, I.; Raguzzini, A.; Serafini, M. Redox Role of Lactobacillus casei Shirota Against the Cellular Damage Induced by 2,2′-Azobis (2-Amidinopropane) Dihydrochloride-Induced Oxidative and Inflammatory Stress in Enterocytes-Like Epithelial Cells. Front. Immunol. 2018, 9, 1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiller, F.; Besselt, K.; Deubel, S.; Brigelius-Flohé, R.; Kipp, A.P. GPx2 Induction Is Mediated Through STAT Transcription Factors During Acute Colitis. Inflamm. Bowel Dis. 2015, 21, 2078–2089. [Google Scholar] [CrossRef] [PubMed]
- Yelin, I.; Flett, K.B.; Merakou, C.; Mehrotra, P.; Stam, J.; Snesrud, E.; Hinkle, M.; Lesho, E.; McGann, P.; McAdam, A.J.; et al. Genomic and epidemiological evidence of bacterial transmission from probiotic capsule to blood in ICU patients. Nat. Med. 2019, 25, 1728–1732. [Google Scholar] [CrossRef]
- Aaron, J.G.; Sobieszczyk, M.E.; Weiner, S.D.; Whittier, S.; Lowy, F.D. Lactobacillus rhamnosus Endocarditis After Upper Endoscopy. Open Forum Infect. Dis. 2017, 4, ofx085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colautti, A.; Arnoldi, M.; Comi, G.; Iacumin, L. Antibiotic resistance and virulence factors in lactobacilli: Something to carefully consider. Food Microbiol. 2022, 103, 103934. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Kirtonia, A.; Sethi, G.; Garg, M. The multifaceted role of reactive oxygen species in tumorigenesis. Cell Mol. Life Sci. 2020, 77, 4459–4483. [Google Scholar] [CrossRef]
- Nakamura, H.; Takada, K. Reactive oxygen species in cancer: Current findings and future directions. Cancer Sci. 2021, 112, 3945–3952. [Google Scholar] [CrossRef]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Sallmyr, A.; Fan, J.; Rassool, F.V. Genomic instability in myeloid malignancies: Increased reactive oxygen species (ROS), DNA double strand breaks (DSBs) and error-prone repair. Cancer Lett. 2008, 270, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shibutani, S.; Takeshita, M.; Grollman, A.P. Insertion of specific bases during DNA synthesis past the oxidation-damaged base 8-oxodG. Nature 1991, 349, 431–434. [Google Scholar] [CrossRef]
- Kato, J.; Miyanishi, K.; Kobune, M.; Nakamura, T.; Takada, K.; Takimoto, R.; Kawano, Y.; Takahashi, S.; Takahashi, M.; Sato, Y.; et al. Long-term phlebotomy with low-iron diet therapy lowers risk of development of hepatocellular carcinoma from chronic hepatitis C. J. Gastroenterol. 2007, 42, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Naguro, I.; Ichijo, H. Iron homeostasis and iron-regulated ROS in cell death, senescence and human diseases. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 1398–1409. [Google Scholar] [CrossRef]
- Son, Y.; Cheong, Y.K.; Kim, N.H.; Chung, H.T.; Kang, D.G.; Pae, H.O. Mitogen-Activated Protein Kinases and Reactive Oxygen Species: How Can ROS Activate MAPK Pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of Reactive Oxygen Species in Cancer Progression: Molecular Mechanisms and Recent Advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberg, F.; Hamanaka, R.; Wheaton, W.W.; Weinberg, S.; Joseph, J.; Lopez, M.; Kalyanaraman, B.; Mutlu, G.M.; Budinger, G.R.S.; Chandel, N.S. Mitochondrial metabolism and ROS generation are essential for Kras-mediated tumorigenicity. Proc. Natl. Acad. Sci. USA 2010, 107, 8788–8793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Goel, V.; Haluska, F.G. PTEN signaling pathways in melanoma. Oncogene 2003, 22, 3113–3122. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.; Lee, S.R.; Yang, K.S.; Ahn, Y.; Kim, Y.J.; Stadtman, E.R.; Rhee, S.G. Reversible oxidation and inactivation of the tumor suppressor PTEN in cells stimulated with peptide growth factors. Proc. Natl. Acad. Sci. USA 2004, 101, 16419–16424. [Google Scholar] [CrossRef] [Green Version]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, M.J.; Liu, Z. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, M.; Soares, M.; Ramalhinho, A.C.; Moutinho, J.F.; Breitenfeld, L.; Pereira, L. The prognostic value of NRF2 in breast cancer patients: A systematic review with meta-analysis. Breast Cancer Res. Treat. 2020, 179, 523–532. [Google Scholar] [CrossRef]
- Rojo de la Vega, M.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef]
- Kerins, M.J.; Ooi, A. A catalogue of somatic NRF2 gain-of-function mutations in cancer. Sci. Rep. 2018, 8, 12846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaitan, D.; Dwarakanath, B.S. Endogenous and induced oxidative stress in multi-cellular tumour spheroids: Implications for improving tumour therapy. Indian J. Biochem. Biophys. 2009, 46, 16–24. [Google Scholar] [PubMed]
- Marullo, R.; Werner, E.; Degtyareva, N.; Moore, B.; Altavilla, G.; Ramalingam, S.S.; Doetsch, P.W. Cisplatin induces a mitochondrial-ROS response that contributes to cytotoxicity depending on mitochondrial redox status and bioenergetic functions. PLoS ONE 2013, 8, e81162. [Google Scholar] [CrossRef] [Green Version]
- Ichijo, H.; Nishida, E.; Irie, K.; ten Dijke, P.; Saitoh, M.; Moriguchi, T.; Takagi, M.; Matsumoto, K.; Miyazono, K.; Gotoh, Y. Induction of apoptosis by ASK1, a mammalian MAPKKK that activates SAPK/JNK and p38 signaling pathways. Science 1997, 275, 90–94. [Google Scholar] [CrossRef]
- Kuo, C.; Ponneri Babuharisankar, A.; Lin, Y.; Lien, H.W.; Lo, Y.K.; Chou, H.; Tangeda, V.; Cheng, L.C.; Cheng, A.N.; Lee, A.Y.-L. Mitochondrial oxidative stress in the tumor microenvironment and cancer immunoescape: Foe or friend? J. Biomed. Sci. 2022, 29, 74. [Google Scholar] [CrossRef]
- Nowak, A.; Paliwoda, A.; Blasiak, J. Anti-proliferative, pro-apoptotic and anti-oxidative activity of Lactobacillus and Bifidobacterium strains: A review of mechanisms and therapeutic perspectives. Crit. Rev. Food Sci. Nutr. 2019, 59, 3456–3467. [Google Scholar] [CrossRef]
- Dasari, S.; Kathera, C.; Janardhan, A.; Praveen Kumar, A.; Viswanath, B. Surfacing role of probiotics in cancer prophylaxis and therapy: A systematic review. Clin. Nutr. 2017, 36, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Daniluk, U.; Alifier, M.; Kaczmarski, M. Probiotic-induced apoptosis and its potential relevance to mucosal inflammation of gastrointestinal tract. Adv. Med. Sci. 2012, 57, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Lin, W.C.; Kong, M.S.; Shi, H.N.; Walker, W.A.; Lin, C.Y.; Huang, C.T.; Lin, Y.C.; Jung, S.M.; Lin, T.Y. Oral inoculation of probiotics Lactobacillus acidophilus NCFM suppresses tumour growth both in segmental orthotopic colon cancer and extra-intestinal tissue. Br. J. Nutr. 2012, 107, 1623–1634. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, C.; Millette, M.; Oth, D.; Ruiz, M.T.; Luquet, F.M.; Lacroix, M. Probiotic Lactobacillus acidophilus and L. casei mix sensitize colorectal tumoral cells to 5-fluorouracil-induced apoptosis. Nutr. Cancer 2010, 62, 371–378. [Google Scholar] [CrossRef]
- Altonsy, M.O.; Andrews, S.C.; Tuohy, K.M. Differential induction of apoptosis in human colonic carcinoma cells (Caco-2) by Atopobium, and commensal, probiotic and enteropathogenic bacteria: Mediation by the mitochondrial pathway. Int. J. Food Microbiol. 2010, 137, 190–203. [Google Scholar] [CrossRef]
- Russo, F.; Orlando, A.; Linsalata, M.; Cavallini, A.; Messa, C. Effects of Lactobacillus rhamnosus GG on the cell growth and polyamine metabolism in HGC-27 human gastric cancer cells. Nutr. Cancer 2007, 59, 106–114. [Google Scholar] [CrossRef]
- Iyer, C.; Kosters, A.; Sethi, G.; Kunnumakkara, A.B.; Aggarwal, B.B.; Versalovic, J. Probiotic Lactobacillus reuteri promotes TNF-induced apoptosis in human myeloid leukemia-derived cells by modulation of NF-kappaB and MAPK signalling. Cell Microbiol. 2008, 10, 1442–1452. [Google Scholar] [CrossRef]
- Hwang, J.W.; Baek, Y.M.; Yang, K.E.; Yoo, H.S.; Cho, C.K.; Lee, Y.W.; Park, J.; Eom, C.Y.; Lee, Z.W.; Choi, J.S.; et al. Lactobacillus casei extract induces apoptosis in gastric cancer by inhibiting NF-κB and mTOR-mediated signaling. Integr. Cancer Ther. 2013, 12, 165–173. [Google Scholar] [CrossRef]
- Orlando, A.; Messa, C.; Linsalata, M.; Cavallini, A.; Russo, F. Effects of Lactobacillus rhamnosus GG on proliferation and polyamine metabolism in HGC-27 human gastric and DLD-1 colonic cancer cell lines. Immunopharmacol. Immunotoxicol. 2009, 31, 108–116. [Google Scholar] [CrossRef]
- Huang, L.; Shan, Y.J.; He, C.X.; Ren, M.H.; Tian, P.J.; Song, W. Effects of L. paracasei subp. paracasei X12 on cell cycle of colon cancer HT-29 cells and regulation of mTOR signalling pathway. J. Funct. Foods 2016, 21, 431–439. [Google Scholar] [CrossRef]
- Russo, F.; Linsalata, M.; Orlando, A. Probiotics against neoplastic transformation of gastric mucosa: Effects on cell proliferation and polyamine metabolism. World J. Gastroenterol. 2014, 20, 13258–13272. [Google Scholar] [CrossRef]
- Thirabunyanon, M.; Hongwittayakorn, P. Potential probiotic lactic acid bacteria of human origin induce antiproliferation of colon cancer cells via synergic actions in adhesion to cancer cells and short-chain fatty acid bioproduction. Appl. Biochem. Biotechnol. 2013, 169, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Sungur, T.; Aslim, B.; Karaaslan, C.; Aktas, B. Impact of Exopolysaccharides (EPSs) of Lactobacillus gasseri strains isolated from human vagina on cervical tumor cells (HeLa). Anaerobe 2017, 47, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Hong, T.; Yu, Q.; Nie, S.; Gong, D.; Xiong, T.; Xie, M. Exopolysaccharides from Lactobacillus plantarum NCU116 induce c-Jun dependent Fas/Fasl-mediated apoptosis via TLR2 in mouse intestinal epithelial cancer cells. Sci. Rep. 2017, 7, 14247. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.F.; Tseng, K.C.; Chiang, S.S.; Lee, B.H.; Hsu, W.H.; Pan, T.M. Immunomodulatory and antioxidant potential of Lactobacillus exopolysaccharides. J. Sci. Food Agric. 2011, 91, 2284–2291. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, F.; Jiang, L.; Liu, R.; Zhang, L.; Lei, X.; Li, J.; Jiang, J.; Guo, H.; Fang, B.; et al. Lactobacillus salivarius REN inhibits rat oral cancer induced by 4-nitroquioline 1-oxide. Cancer Prev. Res. 2013, 6, 686–694. [Google Scholar] [CrossRef] [Green Version]
- Bell, H.N.; Rebernick, R.J.; Goyert, J.; Singhal, R.; Kuljanin, M.; Kerk, S.A.; Huang, W.; Das, N.K.; Andren, A.; Solanki, S.; et al. Reuterin in the healthy gut microbiome suppresses colorectal cancer growth through altering redox balance. Cancer Cell 2022, 40, 185–200.e6. [Google Scholar] [CrossRef]
- Kumar, R.S.; Kanmani, P.; Yuvaraj, N.; Paari, K.A.; Pattukumar, V.; Thirunavukkarasu, C.; Arul, V. Lactobacillus plantarum AS1 isolated from south Indian fermented food Kallappam suppress 1,2-dimethyl hydrazine (DMH)-induced colorectal cancer in male Wistar rats. Appl. Biochem. Biotechnol. 2012, 166, 620–631. [Google Scholar] [CrossRef]
- Choi, S.S.; Kim, Y.; Han, K.S.; You, S.; Oh, S.; Kim, S.H. Effects of Lactobacillus strains on cancer cell proliferation and oxidative stress in vitro. Lett. Appl. Microbiol. 2006, 42, 452–458. [Google Scholar] [CrossRef]
- Hori, T.; Kiyoshima, J.; Yasui, H. Effect of an oral administration of Lactobacillus casei strain Shirota on the natural killer activity of blood mononuclear cells in aged mice. Biosci. Biotechnol. Biochem. 2003, 67, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Asai, Y.; Tamai, R.; Makimura, Y.; Sakamoto, H.; Hashikawa, S.; Yasuda, K. Natural killer cell activities of synbiotic Lactobacillus casei ssp. casei in conjunction with dextran. Clin. Exp. Immunol. 2006, 143, 103–109. [Google Scholar] [CrossRef]
- Shida, K.; Nomoto, K. Probiotics as efficient immunopotentiators: Translational role in cancer prevention. Indian J. Med. Res. 2013, 138, 808–814. [Google Scholar] [PubMed]
- Nami, Y.; Abdullah, N.; Haghshenas, B.; Radiah, D.; Rosli, R.; Khosroushahi, A.Y. Probiotic potential and biotherapeutic effects of newly isolated vaginal Lactobacillus acidophilus 36YL strain on cancer cells. Anaerobe 2014, 28, 29–36. [Google Scholar] [CrossRef]
- Dunn, W.; Shah, V.H. Pathogenesis of Alcoholic Liver Disease. Clin. Liver Dis. 2016, 20, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Shi, J.; Zhao, L.; Guan, J.; Liu, F.; Huo, G.; Li, B. Lactobacillus plantarum KLDS1.0344 and Lactobacillus acidophilus KLDS1.0901 Mixture Prevents Chronic Alcoholic Liver Injury in Mice by Protecting the Intestinal Barrier and Regulating Gut Microbiota and Liver-Related Pathways. J. Agric. Food Chem. 2021, 69, 183–197. [Google Scholar] [CrossRef]
- Ceni, E.; Mello, T.; Galli, A. Pathogenesis of alcoholic liver disease: Role of oxidative metabolism. World J. Gastroenterol. 2014, 20, 17756–17772. [Google Scholar] [CrossRef] [PubMed]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, H.; Zhou, X.; Sun, J.; Liang, X.; Lv, Y.; Bai, L.; Zhang, J.; Gong, P.; Liu, T.; et al. Lactobacillus casei YRL577 ameliorates markers of non-alcoholic fatty liver and alters expression of genes within the intestinal bile acid pathway. Br. J. Nutr. 2020, 125, 521–529. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Farhadi, A.; Jakate, S.M.; Tang, Y.; Shaikh, M.; Keshavarzian, A. Lactobacillus GG treatment ameliorates alcohol-induced intestinal oxidative stress, gut leakiness, and liver injury in a rat model of alcoholic steatohepatitis. Alcohol 2009, 43, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Chi, F.; Wang, G.; Liu, X.; Zhang, Q.; Chen, Y.; Zhang, H.; Chen, W. Lactobacillus rhamnosus CCFM1107 treatment ameliorates alcohol-induced liver injury in a mouse model of chronic alcohol feeding. J. Microbiol. 2015, 53, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Chen, X.; Yi, R.; Zhao, X. Antioxidative and Anti-Inflammatory Effects of ZS62 on Alcohol-Induced Subacute Hepatic Damage. Oxidative Med. Cell Longev. 2021, 2021, 7337988. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, X.; Wang, Y.; Yi, C.; Tian, J.; Liu, K.; Chu, J. Protective effect of lactobacillus plantarum on alcoholic liver injury and regulating of keap-Nrf2-ARE signaling pathway in zebrafish larvae. PLoS ONE 2019, 14, e0222339. [Google Scholar] [CrossRef] [Green Version]
- Azarang, A.; Farshad, O.; Ommati, M.M.; Jamshidzadeh, A.; Heydari, R.; Abootalebi, S.N.; Gholami, A. Protective Role of Probiotic Supplements in Hepatic Steatosis: A Rat Model Study. Biomed. Res. Int. 2020, 2020, 5487659. [Google Scholar] [CrossRef] [PubMed]
- Javadi, L.; Khoshbaten, M.; Safaiyan, A.; Ghavami, M.; Abbasi, M.M.; Gargari, B.P. Pro- and prebiotic effects on oxidative stress and inflammatory markers in non-alcoholic fatty liver disease. Asia Pac. J. Clin. Nutr. 2018, 27, 1031–1039. [Google Scholar] [PubMed]
- Saeedi, B.J.; Liu, K.H.; Owens, J.A.; Hunter-Chang, S.; Camacho, M.C.; Eboka, R.U.; Chandrasekharan, B.; Baker, N.F.; Darby, T.M.; Robinson, B.S.; et al. Gut-Resident Lactobacilli Activate Hepatic Nrf2 and Protect Against Oxidative Liver Injury. Cell Metab. 2020, 31, 956–968.e5. [Google Scholar] [CrossRef]
- Chen, Y.T.; Lin, Y.C.; Lin, J.S.; Yang, N.S.; Chen, M.J. Sugary Kefir Strain Lactobacillus mali APS1 Ameliorated Hepatic Steatosis by Regulation of SIRT-1/Nrf-2 and Gut Microbiota in Rats. Mol. Nutr. Food Res. 2018, 62, e1700903. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, C.; Zhang, L.; Zhao, Y.; Duan, C.; Zhang, X.; Gao, L.; Li, S. Lactobacillus plantarum NA136 improves the non-alcoholic fatty liver disease by modulating the AMPK/Nrf2 pathway. Appl. Microbiol. Biotechnol. 2019, 103, 5843–5850. [Google Scholar] [CrossRef]
- Li, C.; Nie, S.P.; Zhu, K.X.; Ding, Q.; Li, C.; Xiong, T.; Xie, M.Y. Lactobacillus plantarum NCU116 improves liver function, oxidative stress and lipid metabolism in rats with high fat diet induced non-alcoholic fatty liver disease. Food Funct. 2014, 5, 3216–3223. [Google Scholar] [CrossRef]
- Long, X.; Wang, P.; Zhou, Y.; Wang, Q.; Ren, L.; Li, Q.; Zhao, X. Preventive effect of Lactobacillus plantarum HFY15 on carbon tetrachloride (CCl)-induced acute liver injury in mice. J. Food Sci. 2022, 87, 2626–2639. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Q.; Hesketh, J.; Huang, D.; Gan, F.; Hao, S.; Tang, S.; Guo, Y.; Huang, K. Protective effects of selenium-glutathione-enriched probiotics on CCl-induced liver fibrosis. J. Nutr. Biochem. 2018, 58, 138–149. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Q.; Ye, G.; Khan, A.; Liu, J.; Gan, F.; Zhang, X.; Kumbhar, S.; Huang, K. Protective effects of Selenium-enriched probiotics on carbon tetrachloride-induced liver fibrosis in rats. J. Agric. Food Chem. 2015, 63, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Ma, K.; Li, J.; Li, J.; Bi, C.; Shan, A. Deoxynivalenol exposure induces liver damage in mice: Inflammation and immune responses, oxidative stress, and protective effects of Lactobacillus rhamnosus GG. Food Chem. Toxicol. 2021, 156, 112514. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Si, J.; Tan, F.; Park, K.Y.; Zhao, X. Lactobacillus plantarum KSFY06 Prevents Inflammatory Response and Oxidative Stress in Acute Liver Injury Induced by D-Gal/LPS in Mice. Drug Des. Dev. Ther. 2021, 15, 37–50. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The role of Lactobacillus in IBD. Lactobacillus can produce EPS or/and related antioxidant enzymes to alleviate intestinal inflammation and oxidative stress, and ameliorate IBD by regulating the NF-κB, Nrf2 signaling pathway.
Figure 1.
The role of Lactobacillus in IBD. Lactobacillus can produce EPS or/and related antioxidant enzymes to alleviate intestinal inflammation and oxidative stress, and ameliorate IBD by regulating the NF-κB, Nrf2 signaling pathway.

Figure 2.
ROS induces the activation of the MAPK, NF-κB, and PI3K/PTEN signaling pathways to promote cancer development, progression, and apoptosis, etc. and aa possible target of LAB [59].
Figure 2.
ROS induces the activation of the MAPK, NF-κB, and PI3K/PTEN signaling pathways to promote cancer development, progression, and apoptosis, etc. and aa possible target of LAB [59].

Figure 3.
The effects of Lactobacillus treatment in oxidative stress-related liver disease. Some Lactobacillus regulate hepatic lipid metabolism and synthesis via SCFA production and the FXR/SHP/SREBP1c pathway, and some others reduce liver inflammation and oxidative stress through Nrf2 and NF-κB pathways.
Figure 3.
The effects of Lactobacillus treatment in oxidative stress-related liver disease. Some Lactobacillus regulate hepatic lipid metabolism and synthesis via SCFA production and the FXR/SHP/SREBP1c pathway, and some others reduce liver inflammation and oxidative stress through Nrf2 and NF-κB pathways.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the cited Lactobacillus strains and their function in disease.
| Lactobacillus Strain | Diseases Involved | Reported Functions | Reference |
|---|---|---|---|
| L. delbrueckii subsp. bulgaricus B3, L. delbrueckii subsp. bulgaricus A13 | IBD | Produces EPS and antioxidant | [41] |
| L. paracasei IJH-SONE68 | UC | Produces EPS | [42] |
| L. plantarum ZS62 | IBD | Regulates oxidative stress and immune response | [43] |
| L. casei, L. acidophilus, L. rhamnosus | IBD | Modulates inflammatory response and oxidative stress. | [44] |
| L. fermentum ME-3 | /(a healthy volunteer trial) | Antioxidant activity in healthy volunteers | [46] |
| Probiotic bacterial strains VSL#3 | UC | Maintain remission among patients with active UC | [47] |
| Engineered L. casei BL23 | CD | Candidate strain for genetic engineering | [50] |
| L. delbrueckii subsp. Bulgaricus, S. thermophilus CRL 807 | Colitis | Anti-inflammatory effects | [51] |
| L. casei Shirota | Cellular (caco-2 cells) inflammatory damage and oxidative stress induced by DPPH | Alleviate oxidative stress and inflammation via Nrf2 and NF-κB pathway | [53] |
| L. acidophilus NCFM | Orthotopic colon cancers | Attenuates tumor growth and pro-apoptotic effects | [84] |
| L. acidophilus, L. casei | Colorectal cancer | Enhances apoptosis | [85] |
| LGG | Colon cancer | promotes apoptosis | [86] |
| LGG | HGC-27 human gastric cancer cells | Anti-proliferative effects | [87] |
| L. reuteri ATCC PTA 6475 | Human chronic myeloid leukemia-derived cells | Pro-apoptotic and anti-proliferation effects | [88] |
| L. casei | Gastric cancer cells | Pro-apoptotic effects | [89] |
| LGG | Gastric and colonic neoplasms | Anti-proliferative effects | [90] |
| L. paracasei subsp. paracasei X12 | HT-29 colon cancer cells | Induce cell cycle arrest at G1 (anti-proliferative) | [91] |
| L. salivarius FP25, L. salivarius FP35 | Colon cancer cells | Anti-proliferative effects | [93] |
| L. gasseri strains | Cervical cancer cells (HeLa) | Inhibits cancer cell growth; Modulates immune response | [94] |
| L. plantarum NCU116 | Colorectal cell line CT26 | Regulates cancer cell proliferation and apoptosis by EPS | [95] |
| L. salivarius REN | Oral cancer | Induces apoptosis and protects against oxidative DNA damage | [97] |
| L. plantarum AS1 | Colorectal cancer | Antioxidant effects | [99] |
| L. acidophilus 606 | HT-29 colon cancer cells | Induces apoptosis; Antioxidant effects | [100] |
| L. casei Shirota | Normal blood mononuclear cells and splenocytes | Enhances NK cell activity | [101] |
| L. casei ssp. casei | Normal mice | Elicits NK cell activities. | [102] |
| L. casei Shirota | Cancer | Immunomodulatory effects | [103] |
| L. acidophilus 36YL | Human cancer cell lines (AGS, HeLa, MCF-7, and HT-29) | Cytoxicity toward cancer cells | [104] |
| L. plantarum KLDS1.0344; L. acidophilus KLDS1.0901 | ALD | Inhibits liver lipid accumulation, oxidative stress, and inflammation. Regulates gut microbiota | [106] |
| L. casei YRL577 | NAFLD | Modulates genes in intestinal bile acid pathway | [109] |
| LGG | ALD | Reduces oxidative stress and inflammation | [110] |
| L. rhamnosus CCFM1107 | ALD | Reduces oxidative stress; Restores the intestinal microbiota | [111] |
| L. plantarum ZS62 | Alcohol-induced subacute hepatic damage | Reduces inflammation; Enhances antioxidative | [112] |
| L. plantarum | ALD | Antioxidant effects via activation of Keap-Nrf2-ARE pathway | [113] |
| L. acidophilus, L. casei, L. reuteri | NAFLD | Antioxidant effects | [114] |
| Bifidobacterium longum; L. acidophilus | NAFLD | Mitigates oxidative stress and inflammatory response; Improves lipid profiles | [115] |
| LGG | NAFLD | Active Nrf2 | [116] |
| L. mali APS1 | NAFLD | Modulates lipid metabolism and antioxidant activity | [117] |
| L. plantarum NA136 | NAFLD | Regulates the fatty acid metabolism and defends against oxidative stress via AMPK and Nrf2 pathways | [118] |
| L. plantarum NCU116 | NAFLD | Regulates microbiota and lipid metabolism | [119] |
| L. plantarum HFY15 | Liver fibrosis | Antioxidant; Anti-inflammatory; Anti-apoptotic effects | [120] |
| Se-enriched probiotics | Liver fibrosis | Attenuate hepatic oxidative stress, ER stress, and inflammation | [121] |
| Se-enriched probiotics | Liver fibrosis | Attenuate hepatic oxidative stress and inflammation; Induce apoptosis of hepatic stellate cells. | [122] |
| LGG | Deoxynivalenol exposure induces liver damage | Reduces DON toxicity | [123] |
| L. plantarum KSFY06 | Acute liver injury induced by D-Gal/LPS | Anti-oxidant and anti-inflammatory activities | [124] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, T.; Wang, H.; Liu, Z.; Liu, Y.; DeJi; Li, B.; Huang, X. Recent Perspective of Lactobacillus in Reducing Oxidative Stress to Prevent Disease. Antioxidants 2023, 12, 769. https://doi.org/10.3390/antiox12030769
AMA Style
Zhao T, Wang H, Liu Z, Liu Y, DeJi, Li B, Huang X. Recent Perspective of Lactobacillus in Reducing Oxidative Stress to Prevent Disease. Antioxidants. 2023; 12(3):769. https://doi.org/10.3390/antiox12030769
Chicago/Turabian StyleZhao, Tingting, Haoran Wang, Zhenjiang Liu, Yang Liu, DeJi, Bin Li, and Xiaodan Huang. 2023. "Recent Perspective of Lactobacillus in Reducing Oxidative Stress to Prevent Disease" Antioxidants 12, no. 3: 769. https://doi.org/10.3390/antiox12030769
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.