
Impact of Bariatric Surgery on the Stability of the Genetic Material, Oxidation, and Repair of DNA and Telomere Lengths
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Methods
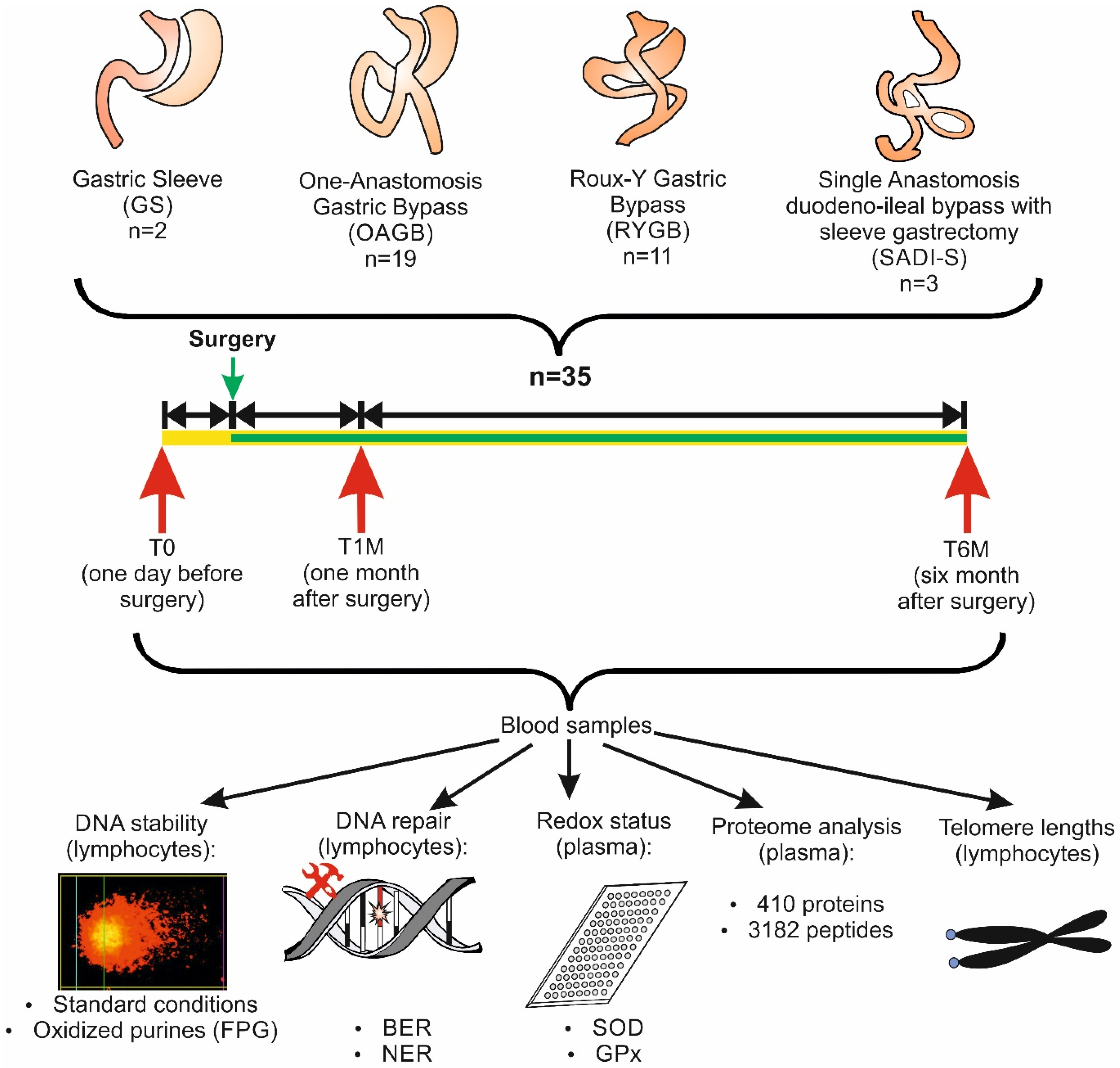
2.1. Recruitment of the Participants
2.2. Isolation of Plasma and Lymphocytes
2.3. SCGE Experiments with Lymphocytes
2.4. Measurement of BER and NER
2.5. Measurement of GPx and SOD
2.6. Measurement of Malondiadehlyde in Plasma
2.7. Measurement of the Telomere Lengths
2.8. Proteome Analyses of the Plasma Samples
2.9. Statistical Analyses
3. Results
3.1. Description of the Study Group
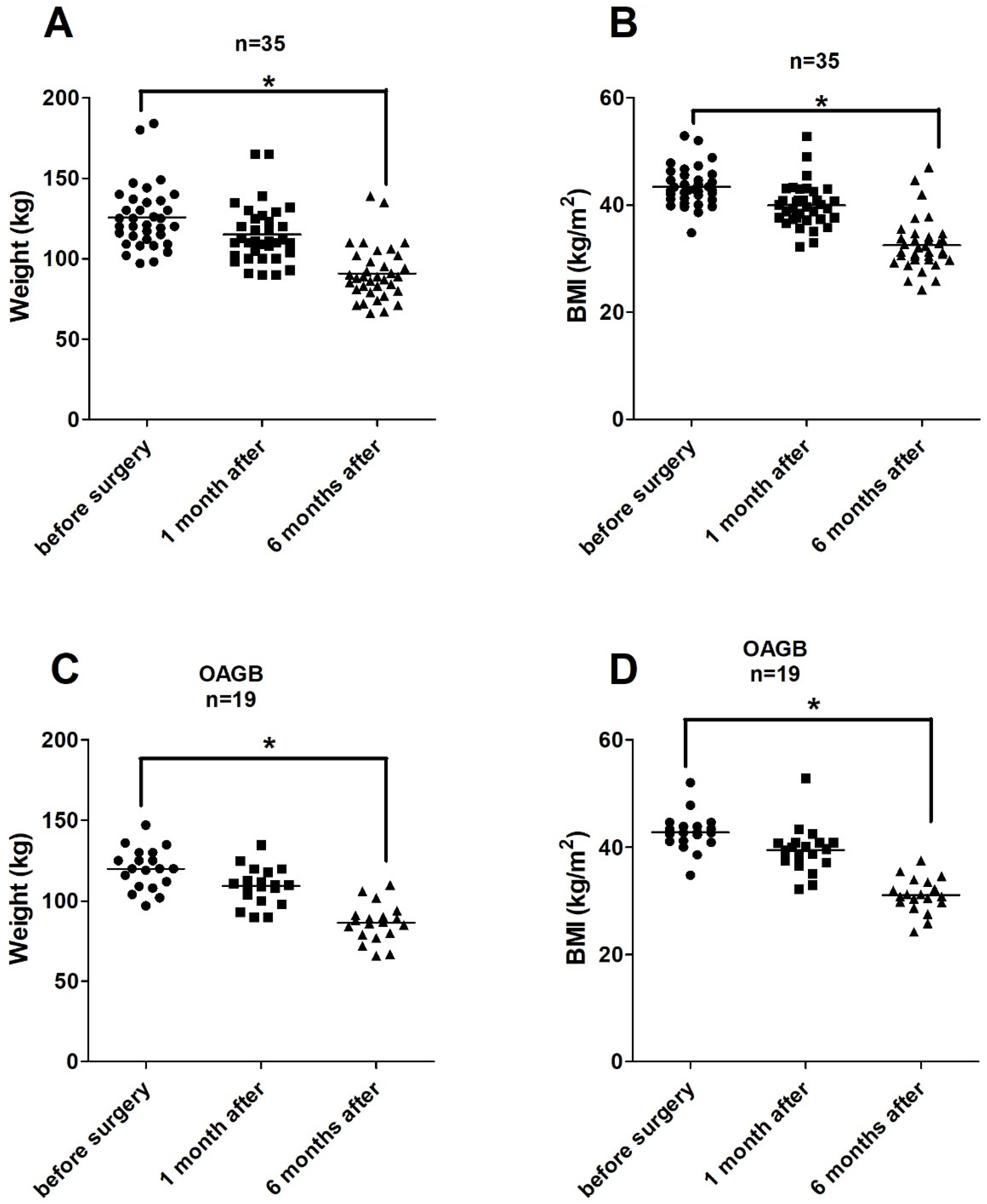
3.2. Impact of BS on Weights and BMIs
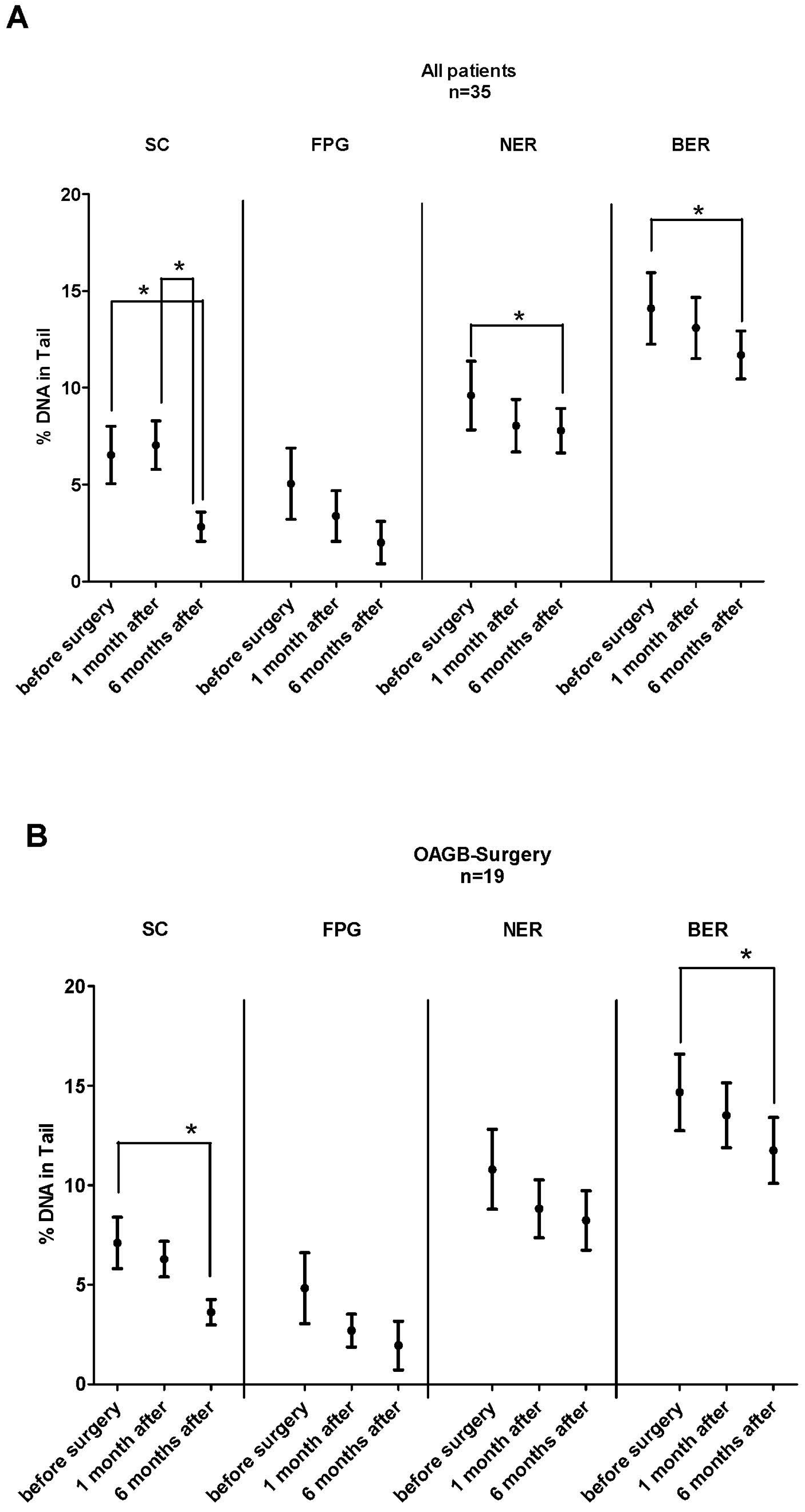
3.3. Impact of BS on DNA Stability and Oxidative DNA Damage and Repair
3.4. Alterations of the Activities of Antioxidant Enzymes
3.5. Results of Malondialdehyde Measurements
3.6. Alterations of the Telomere Lengths
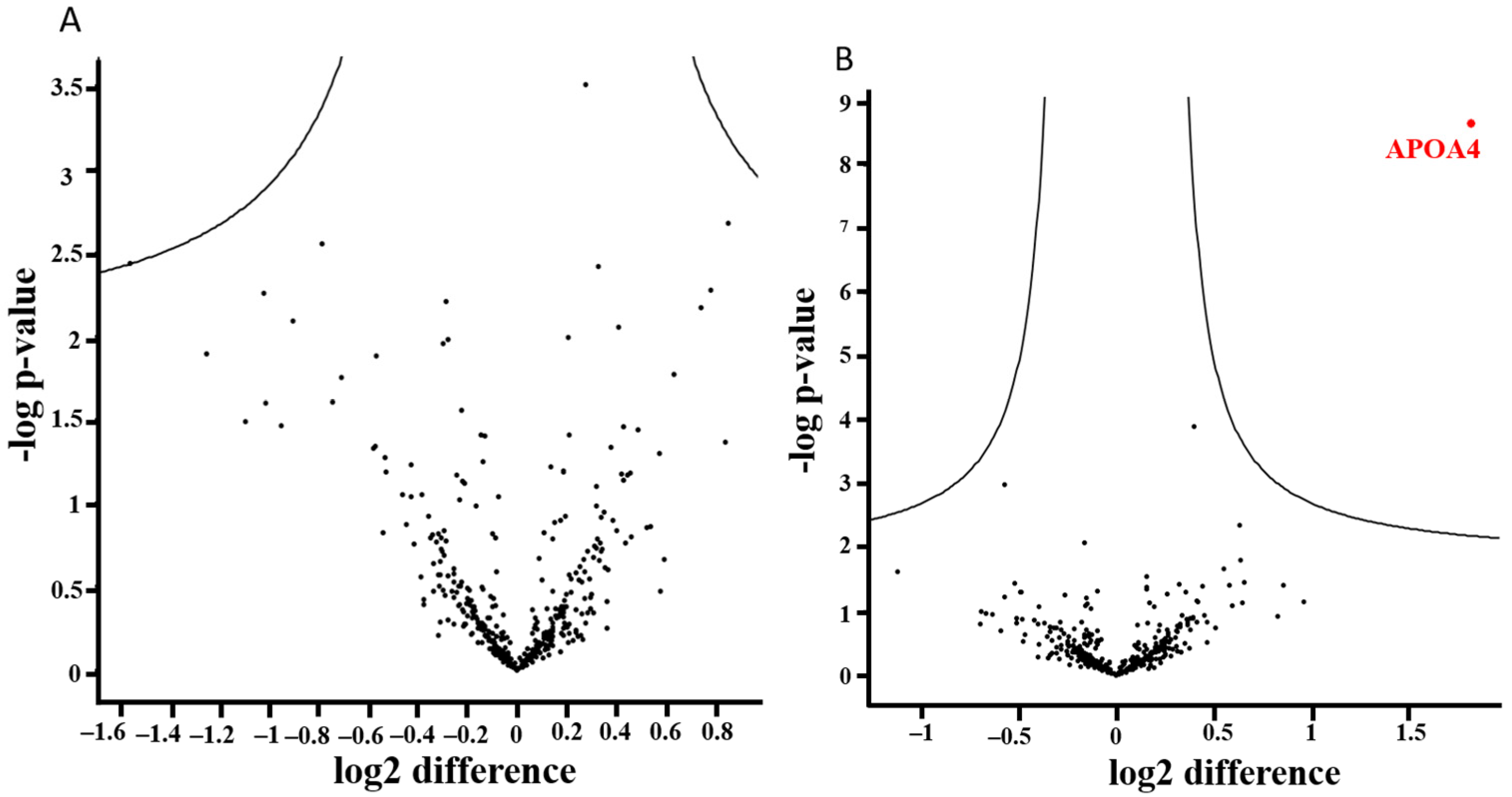
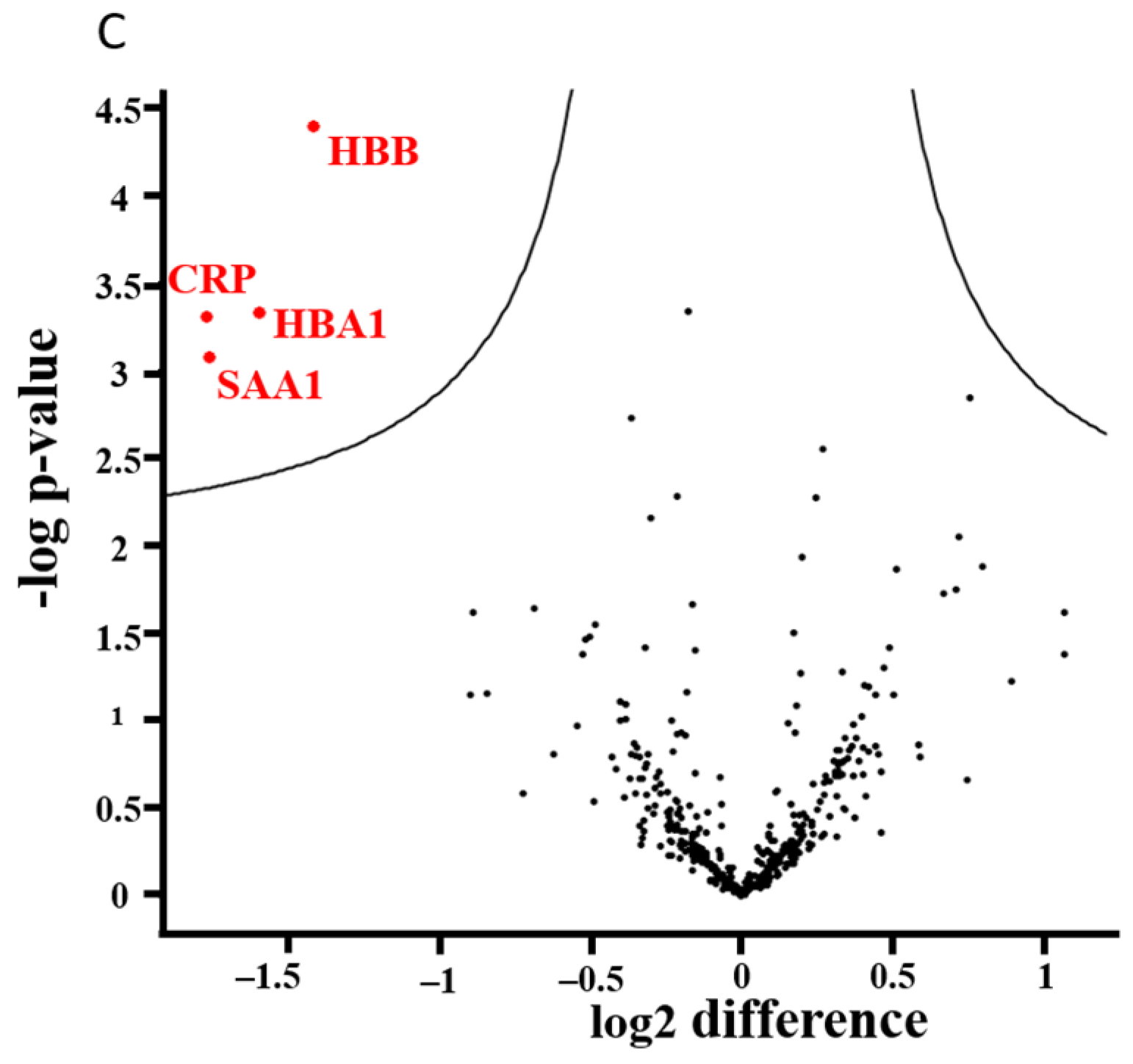
3.7. Alterations of the Proteome Profile
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACN | acetonitrile |
| APOA4 | apolipoprotein A-IV |
| BCA | bicinchoninic acid assay |
| BER | base excision repair |
| BMI | body mass index |
| BS | bariatric surgery |
| BW | body weight |
| CI | confidence interval |
| CRP | C-reactive protein |
| EFSA | European Food Safety Authority |
| FA | formic acid |
| FDR | false discovery rate |
| FPG | formamidopyrimidine glycosylase |
| GPx | glutathione peroxidase |
| GS | gastric sleeve |
| LFQ | label-free quantification |
| LMPA | low melting point agarose |
| LP | lipid peroxidation |
| MDA | malondialdehyde |
| NER | nucleotide excision repair |
| NMPA | normal melting point agarose |
| OAGB | one-anastomosis gastric bypass |
| PASEF | parallel accumulation-serial fragmentation |
| relTL | relative telomere length |
| ROS | reactive oxygen species |
| RYGB SAA1 | Roux-en-Y gastric bypass serum amyloid A1 |
| SADI-S | single-anastomosis duodeno-ileal bypass with sleeve gastrectomy |
| SC | standard conditions |
| SCGE | single-cell gel electrophoresis |
| SD | standard deviation |
| SOD | superoxide dismutase |
References
- WHO. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 2 June 2022).
- Wolfe, B.M.; Kvach, E.; Eckel, R.H. Treatment of Obesity: Weight Loss and Bariatric Surgery. Circ. Res. 2016, 118, 1844–1855. [Google Scholar] [CrossRef] [PubMed]
- Arterburn, D.E.; Telem, D.A.; Kushner, R.F.; Courcoulas, A.P. Benefits and Risks of Bariatric Surgery in Adults: A Review. JAMA 2020, 324, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Luo, Y.; Dai, H.; Deng, Z. Effects of Bariatric Surgery on Cancer Risk: Evidence from Meta-analysis. Obes. Surg. 2020, 30, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Carbajo, M.A.; Luque-de-Leon, E.; Jimenez, J.M.; Ortiz-de-Solorzano, J.; Perez-Miranda, M.; Castro-Alija, M.J. Laparoscopic One-Anastomosis Gastric Bypass: Technique, Results, and Long-Term Follow-Up in 1200 Patients. Obes. Surg. 2017, 27, 1153–1167. [Google Scholar] [CrossRef] [Green Version]
- Gloy, V.L.; Briel, M.; Bhatt, D.L.; Kashyap, S.R.; Schauer, P.R.; Mingrone, G.; Bucher, H.C.; Nordmann, A.J. Bariatric surgery versus non-surgical treatment for obesity: A systematic review and meta-analysis of randomised controlled trials. BMJ 2013, 347, f5934. [Google Scholar] [CrossRef] [Green Version]
- Sheng, B.; Truong, K.; Spitler, H.; Zhang, L.; Tong, X.; Chen, L. The Long-Term Effects of Bariatric Surgery on Type 2 Diabetes Remission, Microvascular and Macrovascular Complications, and Mortality: A Systematic Review and Meta-Analysis. Obes. Surg. 2017, 27, 2724–2732. [Google Scholar] [CrossRef]
- van Veldhuisen, S.L.; Gorter, T.M.; van Woerden, G.; de Boer, R.A.; Rienstra, M.; Hazebroek, E.J.; van Veldhuisen, D.J. Bariatric surgery and cardiovascular disease: A systematic review and meta-analysis. Eur. Heart J. 2022, 43, 1955–1969. [Google Scholar] [CrossRef] [PubMed]
- Setayesh, T.; Nersesyan, A.; Misik, M.; Ferk, F.; Langie, S.; Andrade, V.M.; Haslberger, A.; Knasmuller, S. Impact of obesity and overweight on DNA stability: Few facts and many hypotheses. Mutat. Res. 2018, 777, 64–91. [Google Scholar] [CrossRef]
- Azqueta, A.; Collins, A.R. The essential comet assay: A comprehensive guide to measuring DNA damage and repair. Arch. Toxicol. 2013, 87, 949–968. [Google Scholar] [CrossRef]
- Azqueta, A.; Ladeira, C.; Giovannelli, L.; Boutet-Robinet, E.; Bonassi, S.; Neri, M.; Gajski, G.; Duthie, S.; Del Bo, C.; Riso, P.; et al. Application of the comet assay in human biomonitoring: An hCOMET perspective. Mutat. Res. 2020, 783, 108288. [Google Scholar] [CrossRef]
- Bonassi, S.; Ceppi, M.; Moller, P.; Azqueta, A.; Milic, M.; Neri, M.; Brunborg, G.; Godschalk, R.; Koppen, G.; Langie, S.A.S.; et al. DNA damage in circulating leukocytes measured with the comet assay may predict the risk of death. Sci. Rep. 2021, 11, 16793. [Google Scholar] [CrossRef]
- Moller, P.; Stopper, H.; Collins, A.R. Measurement of DNA damage with the comet assay in high-prevalence diseases: Current status and future directions. Mutagenesis 2020, 35, 5–18. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products; Nutrition and Allergies (EFSA NDA Panel); Turck, D.; Bresson, J.L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; et al. Guidance for the scientific requirements for health claims related to antioxidants, oxidative damage and cardiovascular health: (Revision 1). EFSA J. 2018, 16, e05136. [Google Scholar] [PubMed] [Green Version]
- Maynard, S.; Schurman, S.H.; Harboe, C.; de Souza-Pinto, N.C.; Bohr, V.A. Base excision repair of oxidative DNA damage and association with cancer and aging. Carcinogenesis 2009, 30, 2–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krwawicz, J.; Arczewska, K.D.; Speina, E.; Maciejewska, A.; Grzesiuk, E. Bacterial DNA repair genes and their eukaryotic homologues: 1. Mutations in genes involved in base excision repair (BER) and DNA end processors and their implication in mutagenesis and human disease. Acta Biochim. Pol. 2007, 54, 413–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia-Melo, C.; Hewitt, G.; Passos, J.F. Telomeres, oxidative stress and inflammatory factors: Partners in cellular senescence? Longev. Healthspan 2014, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Vaiserman, A.; Krasnienkov, D. Telomere Length as a Marker of Biological Age: State-of-the-Art, Open Issues, and Future Perspectives. Front. Genet. 2020, 11, 630186. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.; Akinloye, O. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Johnson, F.; Giulivi, C. Superoxide dismutases and their impact upon human health. Mol. Asp. Med. 2005, 26, 340–352. [Google Scholar] [CrossRef]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [Green Version]
- Marnett, L.J. Oxyradicals and DNA damage. Carcinogenesis 2000, 21, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [Green Version]
- Bankoglu, E.E.; Seyfried, F.; Arnold, C.; Soliman, A.; Jurowich, C.; Germer, C.T.; Otto, C.; Stopper, H. Reduction of DNA damage in peripheral lymphocytes of obese patients after bariatric surgery-mediated weight loss. Mutagenesis 2018, 33, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Bankoglu, E.E.; Gerber, J.; Kodandaraman, G.; Seyfried, F.; Stopper, H. Influence of bariatric surgery induced weight loss on oxidative DNA damage. Mutat. Res. 2020, 853, 503194. [Google Scholar] [CrossRef]
- Carlsson, E.R.; Fenger, M.; Henriksen, T.; Kjaer, L.K.; Worm, D.; Hansen, D.L.; Madsbad, S.; Poulsen, H.E. Reduction of oxidative stress on DNA and RNA in obese patients after Roux-en-Y gastric bypass surgery-An observational cohort study of changes in urinary markers. PLoS ONE 2020, 15, e0243918. [Google Scholar] [CrossRef] [PubMed]
- Kocael, A.; Erman, H.; Zengin, K.; Kocael, P.C.; Korkmaz, G.G.; Gelisgen, R.; Taskin, M.; Ersan, Y.; Uzun, H. The effects on oxidative DNA damage of laparoscopic gastric band applications in morbidly obese patients. Can. J. Surg. 2014, 57, 183–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monzo-Beltran, L.; Vazquez-Tarragon, A.; Cerda, C.; Garcia-Perez, P.; Iradi, A.; Sanchez, C.; Climent, B.; Tormos, C.; Vazquez-Prado, A.; Girbes, J.; et al. One-year follow-up of clinical, metabolic and oxidative stress profile of morbid obese patients after laparoscopic sleeve gastrectomy. 8-oxo-dG as a clinical marker. Redox Biol. 2017, 12, 389–402. [Google Scholar] [CrossRef]
- Habermann, N.; Makar, K.W.; Abbenhardt, C.; Xiao, L.; Wang, C.Y.; Utsugi, H.K.; Alfano, C.M.; Campbell, K.L.; Duggan, C.; Foster-Schubert, K.E.; et al. No effect of caloric restriction or exercise on radiation repair capacity. Med. Sci. Sport. Exerc. 2015, 47, 896–904. [Google Scholar] [CrossRef] [Green Version]
- Pena, E.; Leon-Mengibar, J.; Powell, T.R.; Caixas, A.; Cardoner, N.; Rosa, A. Telomere length in patients with obesity submitted to bariatric surgery: A systematic review. Eur. Eat. Disord. Rev. 2021, 29, 842–853. [Google Scholar] [CrossRef]
- Schmatz, R.; Bitencourt, M.R.; Patias, L.D.; Beck, M.; da, C. Alvarez, G.; Zanini, D.; Gutierres, J.M.; Diehl, L.N.; Pereira, L.B.; Leal, C.A.; et al. Evaluation of the biochemical, inflammatory and oxidative profile of obese patients given clinical treatment and bariatric surgery. Clin. Chim. Acta 2017, 465, 72–79. [Google Scholar] [CrossRef]
- Abad-Jimenez, Z.; Lopez-Domenech, S.; Gomez-Abril, S.A.; Perianez-Gomez, D.; de Maranon, A.M.; Banuls, C.; Morillas, C.; Victor, V.M.; Rocha, M. Effect of Roux-en-Y Bariatric Bypass Surgery on Subclinical Atherosclerosis and Oxidative Stress Markers in Leukocytes of Obese Patients: A One-Year Follow-Up Study. Antioxidants 2020, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Nersesyan, A.; Hoelzl, C.; Ferk, F.; Mišík, M.; Knasmueller, S. Comet assays in dietary intervention trials. In The Comet Assay in Toxicology; Royal Society of Chemistry: Cambridge, UK, 2009; pp. 267–296. [Google Scholar]
- Hoelzl, C.; Knasmuller, S.; Misik, M.; Collins, A.; Dusinska, M.; Nersesyan, A. Use of single-cell gel electrophoresis assays for the detection of DNA protective effects of dietary factors in humans: Recent results and trends. Mutat. Res. 2009, 681, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Bal, B.S.; Finelli, F.C.; Shope, T.R.; Koch, T.R. Nutritional deficiencies after bariatric surgery. Nat. Rev. Endocrinol. 2012, 8, 544–556. [Google Scholar] [CrossRef]
- Azqueta, A.; Muruzabal, D.; Boutet-Robinet, E.; Milic, M.; Dusinska, M.; Brunborg, G.; Moller, P.; Collins, A.R. Technical recommendations to perform the alkaline standard and enzyme-modified comet assay in human biomonitoring studies. Mutat. Res. 2019, 843, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Koppen, G.; Azqueta, A.; Pourrut, B.; Brunborg, G.; Collins, A.R.; Langie, S.A.S. The next three decades of the comet assay: A report of the 11th International Comet Assay Workshop. Mutagenesis 2017, 32, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Azqueta, A.; Shaposhnikov, S.; Collins, A. Detection of Oxidised DNA Using DNA Repair Enzymes. In Comet Assay in Toxicology; Dhawan, A., Anderson, D., Eds.; Royal Society of Chemistry: Cambridge, UK, 2009; Volume 5, pp. 57–78. [Google Scholar]
- Ramel, A.; Wagner, K.H.; Elmadfa, I. Plasma antioxidants and lipid oxidation after submaximal resistance exercise in men. Eur. J. Nutr. 2004, 43, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Ferk, F.; Kundi, M.; Brath, H.; Szekeres, T.; Al-Serori, H.; Misik, M.; Saiko, P.; Marculescu, R.; Wagner, K.H.; Knasmueller, S. Gallic Acid Improves Health-Associated Biochemical Parameters and Prevents Oxidative Damage of DNA in Type 2 Diabetes Patients: Results of a Placebo-Controlled Pilot Study. Mol. Nutr. Food Res. 2018, 62, 1700482. [Google Scholar] [CrossRef]
- Ferk, F.; Misik, M.; Nersesyan, A.; Pichler, C.; Jager, W.; Szekeres, T.; Marculescu, R.; Poulsen, H.E.; Henriksen, T.; Bono, R.; et al. Impact of xanthohumol (a prenylated flavonoid from hops) on DNA stability and other health-related biochemical parameters: Results of human intervention trials. Mol. Nutr. Food Res. 2016, 60, 773–786. [Google Scholar] [CrossRef]
- Wultsch, G.; Nersesyan, A.; Kundi, M.; Wagner, K.H.; Ferk, F.; Jakse, R.; Knasmueller, S. Impact of exposure to wood dust on genotoxicity and cytotoxicity in exfoliated buccal and nasal cells. Mutagenesis 2015, 30, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Cawthon, R.M. Telomere length measurement by a novel monochrome multiplex quantitative PCR method. Nucleic Acids Res. 2009, 37, e21. [Google Scholar] [CrossRef] [Green Version]
- Wultsch, G.; Setayesh, T.; Kundi, M.; Al-Serori, H.; Kreilmeier-Berger, T.; Ropek, N.; Holzmann, K.; Nersesyan, A.; Knasmuller, S. Impact of extended working periods on genomic and telomeric DNA and on inflammatory markers: Results of an intervention study with office workers and carpenters. Mutat. Res. 2018, 836, 78–81. [Google Scholar] [CrossRef]
- Zougman, A.; Selby, P.J.; Banks, R.E. Suspension trapping (STrap) sample preparation method for bottom-up proteomics analysis. Proteomics 2014, 14, 1006-1000. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Julve, J.; Pardina, E.; Perez-Cuellar, M.; Ferrer, R.; Rossell, J.; Baena-Fustegueras, J.A.; Fort, J.M.; Lecube, A.; Blanco-Vaca, F.; Sanchez-Quesada, J.L.; et al. Bariatric surgery in morbidly obese patients improves the atherogenic qualitative properties of the plasma lipoproteins. Atherosclerosis 2014, 234, 200–205. [Google Scholar] [CrossRef]
- Iqbal, Z.; Bashir, B.; Adam, S.; Ho, J.H.; Dhage, S.; Azmi, S.; Ferdousi, M.; Yusuf, Z.; Donn, R.; Malik, R.A.; et al. Glycated apolipoprotein B decreases after bariatric surgery in people with and without diabetes: A potential contribution to reduction in cardiovascular risk. Atherosclerosis 2022, 346, 10–17. [Google Scholar] [CrossRef]
- Tan, H.C.; Hsu, J.W.; Tai, E.S.; Chacko, S.; Wu, V.; Lee, C.F.; Kovalik, J.P.; Jahoor, F. De Novo Glycine Synthesis Is Reduced in Adults With Morbid Obesity and Increases Following Bariatric Surgery. Front. Endocrinol. 2022, 13, 900343. [Google Scholar] [CrossRef]
- Chambers, A.P.; Jessen, L.; Ryan, K.K.; Sisley, S.; Wilson-Perez, H.E.; Stefater, M.A.; Gaitonde, S.G.; Sorrell, J.E.; Toure, M.; Berger, J.; et al. Weight-independent changes in blood glucose homeostasis after gastric bypass or vertical sleeve gastrectomy in rats. Gastroenterology 2011, 141, 950–958. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, N.B.; Jacobsen, S.H.; Dirksen, C.; Bojsen-Moller, K.N.; Naver, L.; Hvolris, L.; Clausen, T.R.; Wulff, B.S.; Worm, D.; Lindqvist Hansen, D.; et al. Acute and long-term effects of Roux-en-Y gastric bypass on glucose metabolism in subjects with Type 2 diabetes and normal glucose tolerance. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E122–E131. [Google Scholar] [CrossRef] [Green Version]
- Lingvay, I.; Guth, E.; Islam, A.; Livingston, E. Rapid improvement in diabetes after gastric bypass surgery: Is it the diet or surgery? Diabetes Care 2013, 36, 2741–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefater, M.A.; Wilson-Perez, H.E.; Chambers, A.P.; Sandoval, D.A.; Seeley, R.J. All bariatric surgeries are not created equal: Insights from mechanistic comparisons. Endocr. Rev. 2012, 33, 595–622. [Google Scholar] [CrossRef] [Green Version]
- Bentham, J.; Di Cesare, M.; Bilano, V.; Bixby, H.; Zhou, B.; Stevens, G.A.; Riley, L.M.; Taddei, C.; Hajifathalian, K.; Lu, Y.; et al. Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: A pooled analysis of 2416 population-based measurement studies in 128·9 million children, adolescents, and adults. Lancet 2017, 390, 2627–2642. [Google Scholar] [CrossRef] [Green Version]
- Rutledge, R. The mini-gastric bypass: Experience with the first 1,274 cases. Obes. Surg. 2001, 11, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Nedeljkovic-Arsenovic, O.; Banovic, M.; Radenkovic, D.; Rancic, N.; Polovina, S.; Micic, D.; Nedeljkovic, I. The Amount of Weight Loss Six Months after Bariatric Surgery: It Makes a Difference. Obes. Facts 2019, 12, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lee, C.H.; Lee, H.M.; Yin, W.Y.; Chin, W.L.; Lee, M.H.; Chen, J.H. Check point to get adequate weight loss within 6-months after laparoscopic sleeve gastrectomy for morbid obesity in Asian population. Sci. Rep. 2020, 10, 12788. [Google Scholar] [CrossRef]
- Kermansaravi, M.; Pishgahroudsari, M.; Kabir, A.; Abdolhosseini, M.R.; Pazouki, A. Weight loss after one-anastomosis/mini-gastric bypass—The impact of biliopancreatic limb: A retrospective cohort study. J. Res. Med. Sci. 2020, 25, 5. [Google Scholar] [CrossRef]
- Heilbronn, L.K.; de Jonge, L.; Frisard, M.I.; DeLany, J.P.; Larson-Meyer, D.E.; Rood, J.; Nguyen, T.; Martin, C.K.; Volaufova, J.; Most, M.M.; et al. Effect of 6-month calorie restriction on biomarkers of longevity, metabolic adaptation, and oxidative stress in overweight individuals: A randomized controlled trial. JAMA 2006, 295, 1539–1548. [Google Scholar] [CrossRef]
- Ozvald, I.; Bozicevic, D.; Duh, L.; Vinkovic Vrcek, I.; Domijan, A.M.; Milic, M. Changes in anthropometric, biochemical, oxidative, and DNA damage parameters after 3-weeks-567-kcal-hospital-controlled-VLCD in severely obese patients with BMI ≥ 35 kg m−2. Clin. Nutr. ESPEN 2022, 49, 319–327. [Google Scholar] [CrossRef]
- Benassi-Evans, B.; Clifton, P.M.; Noakes, M.; Keogh, J.B.; Fenech, M. High protein-high red meat versus high carbohydrate weight loss diets do not differ in effect on genome stability and cell death in lymphocytes of overweight men. Mutagenesis 2009, 24, 271–277. [Google Scholar] [CrossRef]
- Moller, P.; Loft, S. Interventions with antioxidants and nutrients in relation to oxidative DNA damage and repair. Mutat. Res. 2004, 551, 79–89. [Google Scholar] [CrossRef]
- Gunasekarana, V.; Raj, G.V.; Chand, P. A comprehensive review on clinical applications of comet assay. J. Clin. Diagn. Res. 2015, 9, GE01–GE05. [Google Scholar] [CrossRef]
- Bukowska, B.; Karwowski, B.T. Actual state of knowledge in the field of diseases related with defective nucleotide excision repair. Life Sci. 2018, 195, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Tyson, J.; Caple, F.; Spiers, A.; Burtle, B.; Daly, A.K.; Williams, E.A.; Hesketh, J.E.; Mathers, J.C. Inter-individual variation in nucleotide excision repair in young adults: Effects of age, adiposity, micronutrient supplementation and genotype. Br. J. Nutr. 2009, 101, 1316–1323. [Google Scholar] [CrossRef] [Green Version]
- Parrott, J.; Frank, L.; Rabena, R.; Craggs-Dino, L.; Isom, K.A.; Greiman, L. American Society for Metabolic and Bariatric Surgery Integrated Health Nutritional Guidelines for the Surgical Weight Loss Patient 2016 Update: Micronutrients. Surg. Obes. Relat. Dis. 2017, 13, 727–741. [Google Scholar] [CrossRef]
- Osland, E.; Powlesland, H.; Guthrie, T.; Lewis, C.A.; Memon, M.A. Micronutrient management following bariatric surgery: The role of the dietitian in the postoperative period. Ann. Transl. Med. 2020, 8, S9. [Google Scholar] [CrossRef] [PubMed]
- Basten, G.P.; Duthie, S.J.; Pirie, L.; Vaughan, N.; Hill, M.H.; Powers, H.J. Sensitivity of markers of DNA stability and DNA repair activity to folate supplementation in healthy volunteers. Br. J. Cancer 2006, 94, 1942–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarnieri, S.; Loft, S.; Riso, P.; Porrini, M.; Risom, L.; Poulsen, H.E.; Dragsted, L.O.; Moller, P. DNA repair phenotype and dietary antioxidant supplementation. Br. J. Nutr. 2008, 99, 1018–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brevik, A.; Karlsen, A.; Azqueta, A.; Tirado, A.E.; Blomhoff, R.; Collins, A. Both base excision repair and nucleotide excision repair in humans are influenced by nutritional factors. Cell Biochem. Funct. 2011, 29, 36–42. [Google Scholar] [CrossRef]
- Guan, B.; Chen, Y.; Chong, T.H.; Peng, J.; Mak, T.K.; Wang, C.; Yang, J. Effect of Bariatric Surgery on Serum Enzyme Status in Obese Patients. Obes. Surg. 2020, 30, 2700–2707. [Google Scholar] [CrossRef]
- João Cabrera, E.; Valezi, A.C.; Delfino, V.D.A.; Lavado, E.L.; Barbosa, D.S. Reduction in plasma levels of inflammatory and oxidative stress indicators after Roux-en-Y gastric bypass. Obes. Surg. 2010, 20, 42–49. [Google Scholar] [CrossRef]
- Rubino, F. Bariatric surgery: Effects on glucose homeostasis. Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 497–507. [Google Scholar] [CrossRef]
- Othman, E.M.; Leyh, A.; Stopper, H. Insulin mediated DNA damage in mammalian colon cells and human lymphocytes in vitro. Mutat. Res. 2013, 745–746, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, S.A.; Rajoka, M.I.; Nagra, S.A.; Rehman, Z.U. Plasma homocysteine and DNA damage profiles in normal and obese subjects in the Pakistani population. Mol. Biol. Rep. 2010, 37, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Kim, O.Y.; Ryu, H.J.; Kim, J.Y.; Song, S.H.; Ordovas, J.M.; Lee, J.H. Visceral fat accumulation determines postprandial lipemic response, lipid peroxidation, DNA damage, and endothelial dysfunction in nonobese Korean men. J. Lipid Res. 2003, 44, 2356–2364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaman, A.; Aydin, H.; Geckinli, B.; Cetinkaya, A.; Karaman, S. DNA damage is increased in lymphocytes of patients with metabolic syndrome. Mutat. Res. 2015, 782, 30–35. [Google Scholar] [CrossRef]
- Liu, A.L.; Lu, W.Q.; Wang, Z.Z.; Chen, W.H.; Lu, W.H.; Yuan, J.; Nan, P.H.; Sun, J.Y.; Zou, Y.L.; Zhou, L.H.; et al. Elevated levels of urinary 8-hydroxy-2 -deoxyguanosine, lymphocytic micronuclei, and serum glutathione S-transferase in workers exposed to coke oven emissions. Environ. Health Perspect. 2006, 114, 673–677. [Google Scholar] [CrossRef] [Green Version]
- Laimer, M.; Melmer, A.; Lamina, C.; Raschenberger, J.; Adamovski, P.; Engl, J.; Ress, C.; Tschoner, A.; Gelsinger, C.; Mair, L.; et al. Telomere length increase after weight loss induced by bariatric surgery: Results from a 10 year prospective study. Int. J. Obes. 2016, 40, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Muezzinler, A.; Zaineddin, A.K.; Brenner, H. Body mass index and leukocyte telomere length in adults: A systematic review and meta-analysis. Obes. Rev. 2014, 15, 192–201. [Google Scholar] [CrossRef]
- Wewer Albrechtsen, N.J.; Geyer, P.E.; Doll, S.; Treit, P.V.; Bojsen-Moller, K.N.; Martinussen, C.; Jorgensen, N.B.; Torekov, S.S.; Meier, F.; Niu, L.; et al. Plasma Proteome Profiling Reveals Dynamics of Inflammatory and Lipid Homeostasis Markers after Roux-En-Y Gastric Bypass Surgery. Cell Syst. 2018, 7, 601–612.e603. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.A.; de Jersey, S.; Seymour, M.; Hopkins, G.; Hickman, I.; Osland, E. Iron, Vitamin B12, Folate and Copper Deficiency After Bariatric Surgery and the Impact on Anaemia: A Systematic Review. Obes. Surg. 2020, 30, 4542–4591. [Google Scholar] [CrossRef]
- Ansar, W.; Ghosh, S. C-reactive protein and the biology of disease. Immunol. Res. 2013, 56, 131–142. [Google Scholar] [CrossRef]
- Sun, L.; Ye, R.D. Serum amyloid A1: Structure, function and gene polymorphism. Gene 2016, 583, 48–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askarpour, M.; Khani, D.; Sheikhi, A.; Ghaedi, E.; Alizadeh, S. Effect of Bariatric Surgery on Serum Inflammatory Factors of Obese Patients: A Systematic Review and Meta-Analysis. Obes. Surg. 2019, 29, 2631–2647. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Values 1 |
|---|---|
| Average age (years) | |
| All | 41.3 ± 13.3 |
| OAGB | 40.1 ± 12.2 |
| RYGB | 43.6 ± 13.9 |
| GS | 50.5 ± 10.6 |
| SADI-S | 33.3 ± 20.0 |
| Gender | |
| All | 29 F, 6 M |
| OAGB | 18 F, 1 M |
| RYGB | 8 F, 3 M |
| GS | 2 M |
| SADI-S | 3 F |
| Smoking | |
| All | 13 (12 F, 1 M) |
| OAGB | 5 (5 F) |
| RYGB | 4 (4 F) |
| GS | 1 (1 M) |
| SADI-S | 3 (3 F) |
| Initial weight (kg) | |
| All | 125.5 ± 19.6 |
| OAGB | 120.0 ± 12.8 |
| RYGB | 128.6 ± 23.5 |
| GS | 130.5 ± 13.4 |
| SADI-S | 145.3 ± 35.2 |
| Initial BMI (kg/m2) | |
| All | 45.4 ± 6.6 |
| OAGB | 43.6 ± 4.3 |
| RYGB | 47.1 ± 8.0 |
| GS | 40.2 ± 2.2 |
| SADI-S | 54.1 ± 7.5 |
| Parameters | before Surgery | 1 Month after Surgery | ∆ (%) (T0 vs. 1M) | p-Values (T0 vs. 1M) | 6 Months after Surgery | ∆ (%) (T0 vs. 6M) | p-Values | ∆ (%) (1M vs. 6M) | p-Values |
|---|---|---|---|---|---|---|---|---|---|
| SOD | |||||||||
| (% inhibition of tetrazolium salt formation) | |||||||||
| All (n = 35) | 55.8 ± 9.6 | 52.2 ± 9.4 | −6.4 | 0.158 | 54.7 ± 8.5 | −1.9 | 0.562 | +4.7 | 0.244 |
| OAGB (n = 19) | 54.3 ± 9.4 | 52.3 ± 10.5 | −3.6 | 0.653 | 53.7 ± 8.5 | −1.1 | 0.854 | +2.6 | 0.694 |
| GPx (mU/mL) | |||||||||
| All (n = 35) | 719.5 ± 163.6 | 667.0 ± 132.6 | −7.3 | 0.228 | 701.1 ± 139.5 | −2.5 | 0.787 | +5.0 | 0.151 |
| OAGB (n = 19) | 690.9 ± 168.0 | 642.7 ± 116.9 | −6.9 | 0.437 | 669.2 ± 123.2 | +1.2 | 0.825 | +4.1 | 0.407 |
| Telomere (T/S Ratio) (in Lymphocytes) | before Surgery | 1 Month after Surgery | ∆ (%) (T0 vs. 1M) | p-Values | 6 Months after Surgery | ∆ (%) (T0 vs. 6M) | p-Values | ∆ (%) (1M vs. 6M) | p-Values |
|---|---|---|---|---|---|---|---|---|---|
| relTL-ALB | |||||||||
| All (n = 34) 2 | 1.23 ± 1.19 | 0.99 ± 0.46 | −19.5 | 0.353 | 1.11 ± 0.42 | −9.7 | 0.512 | +12.1 | 0.022 * |
| OAGB (n = 19) | 1.24 ± 0.83 | 1.00 ± 0.39 | −19.3 | 0.157 | 1.18 ± 0.44 | −4.8 | 0.712 | +18.0 | 0.046 * |
| relTL-36B4 | |||||||||
| All (n = 34) 2 | 1.05 ± 0.47 | 1.02 ± 0.44 | −2.8 | 0.797 | 1.08 ± 0.43 | +2.8 | 0.447 | +5.8 | 0.300 |
| OAGB (n = 19) | 1.16 ± 0.47 | 1.05 ± 0.44 | −9.4 | 0.220 | 1.18 ± 0.38 | +1.7 | 0.667 | +12.3 | 0.125 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferk, F.; Mišík, M.; Ernst, B.; Prager, G.; Bichler, C.; Mejri, D.; Gerner, C.; Bileck, A.; Kundi, M.; Langie, S.; et al. Impact of Bariatric Surgery on the Stability of the Genetic Material, Oxidation, and Repair of DNA and Telomere Lengths. Antioxidants 2023, 12, 760. https://doi.org/10.3390/antiox12030760
Ferk F, Mišík M, Ernst B, Prager G, Bichler C, Mejri D, Gerner C, Bileck A, Kundi M, Langie S, et al. Impact of Bariatric Surgery on the Stability of the Genetic Material, Oxidation, and Repair of DNA and Telomere Lengths. Antioxidants. 2023; 12(3):760. https://doi.org/10.3390/antiox12030760
Chicago/Turabian StyleFerk, Franziska, Miroslav Mišík, Benjamin Ernst, Gerhard Prager, Christoph Bichler, Doris Mejri, Christopher Gerner, Andrea Bileck, Michael Kundi, Sabine Langie, and et al. 2023. "Impact of Bariatric Surgery on the Stability of the Genetic Material, Oxidation, and Repair of DNA and Telomere Lengths" Antioxidants 12, no. 3: 760. https://doi.org/10.3390/antiox12030760