
SARS-CoV-2 Main Protease Targets Host Selenoproteins and Glutathione Biosynthesis for Knockdown via Proteolysis, Potentially Disrupting the Thioredoxin and Glutaredoxin Redox Cycles


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Peptide Cleavage Materials
2.2. Expression and Purification of TXNRD1 Protein
2.3. Cleavage of Recombinant Peptides
2.4. Cleavage of TXNRD1 Protein
2.5. Cleavage Analysis via UPLC-MS
3. Results
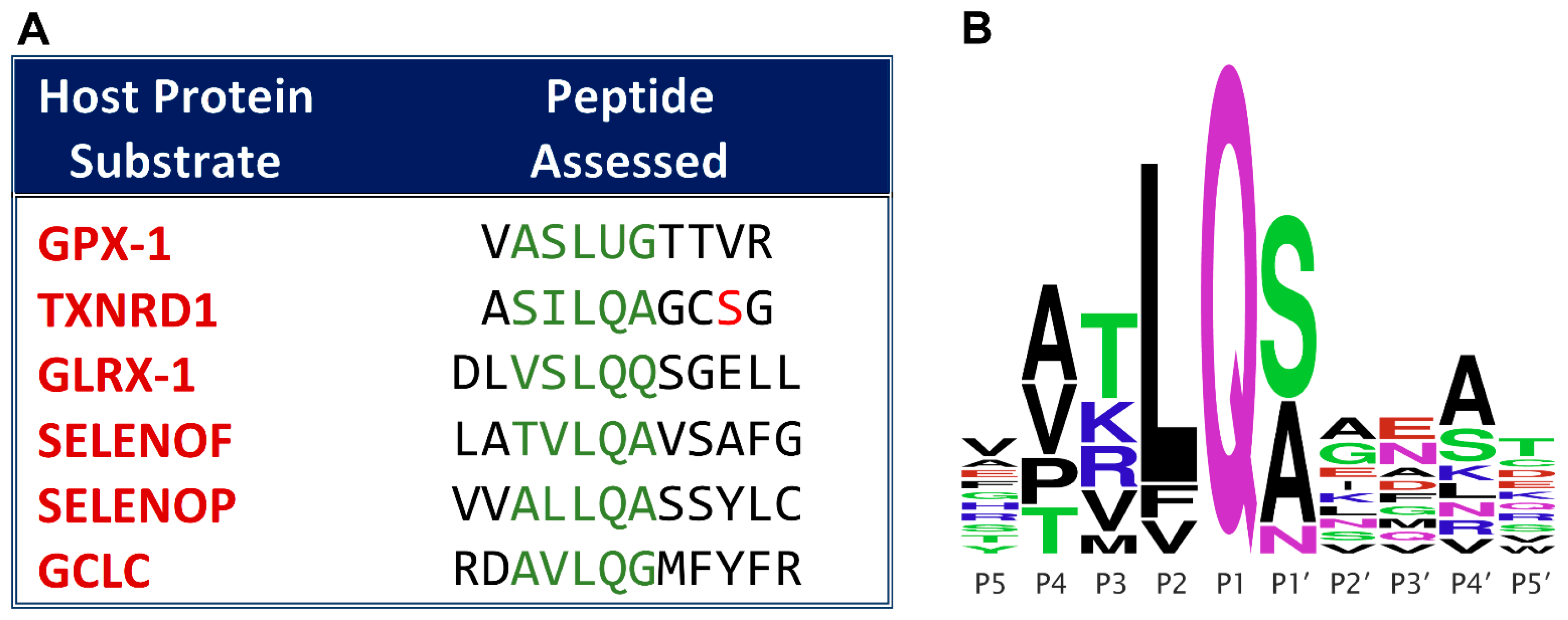
3.1. Validation of Proposed Cleavage Sites
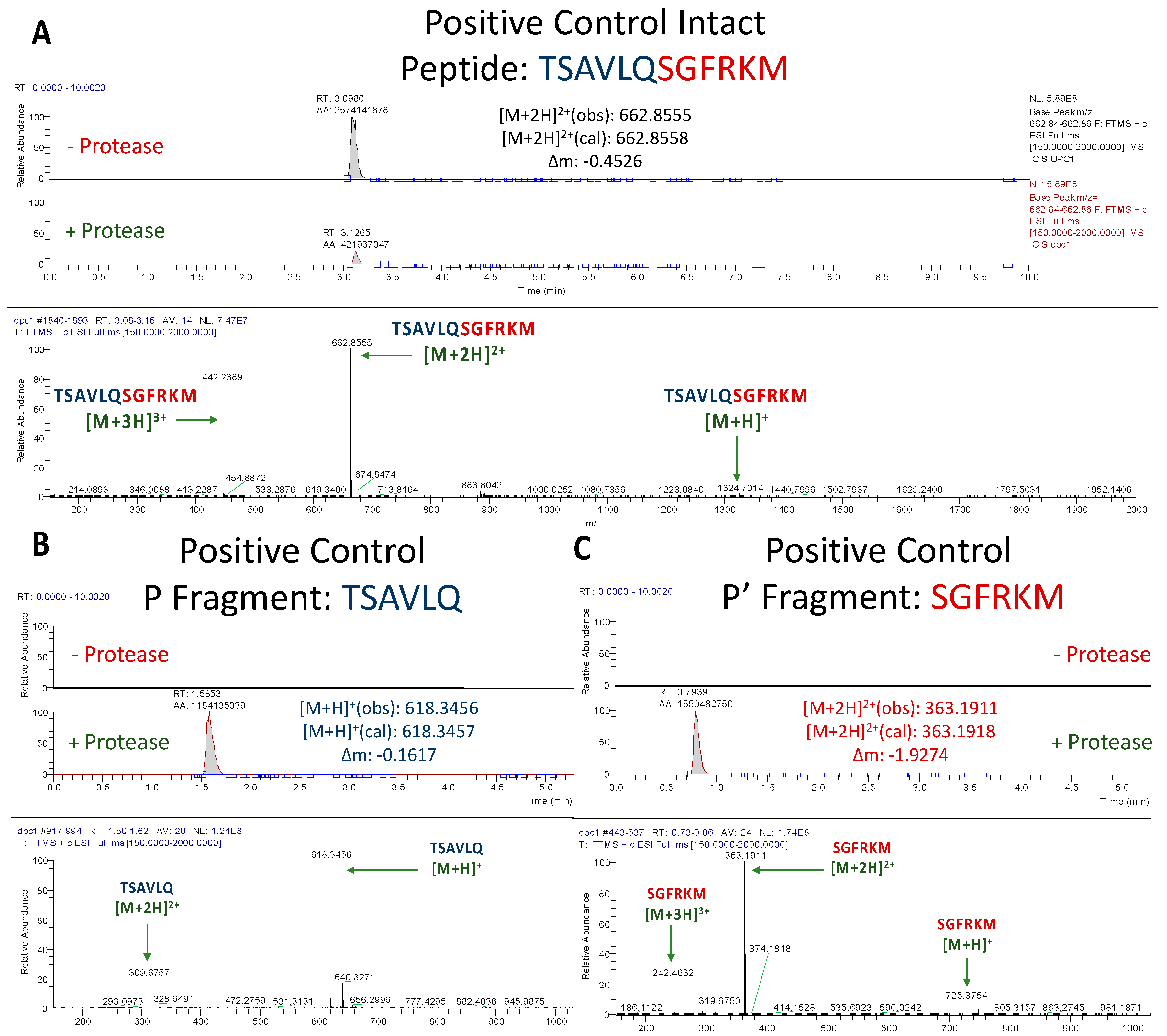
3.1.1. Assessment of Positive Control Peptide Cleavage
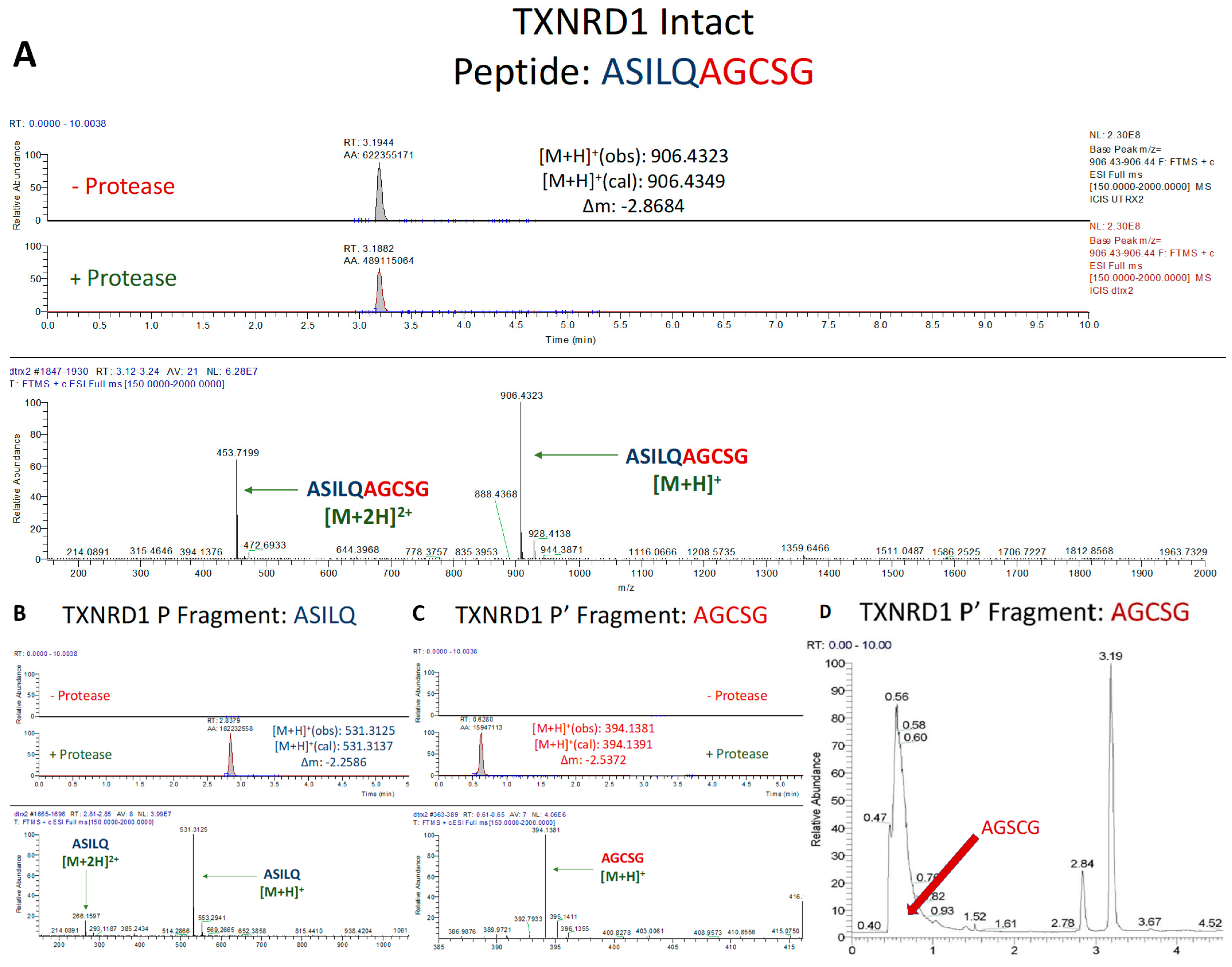
3.1.2. Assessment of TXNRD1 Peptide Cleavage
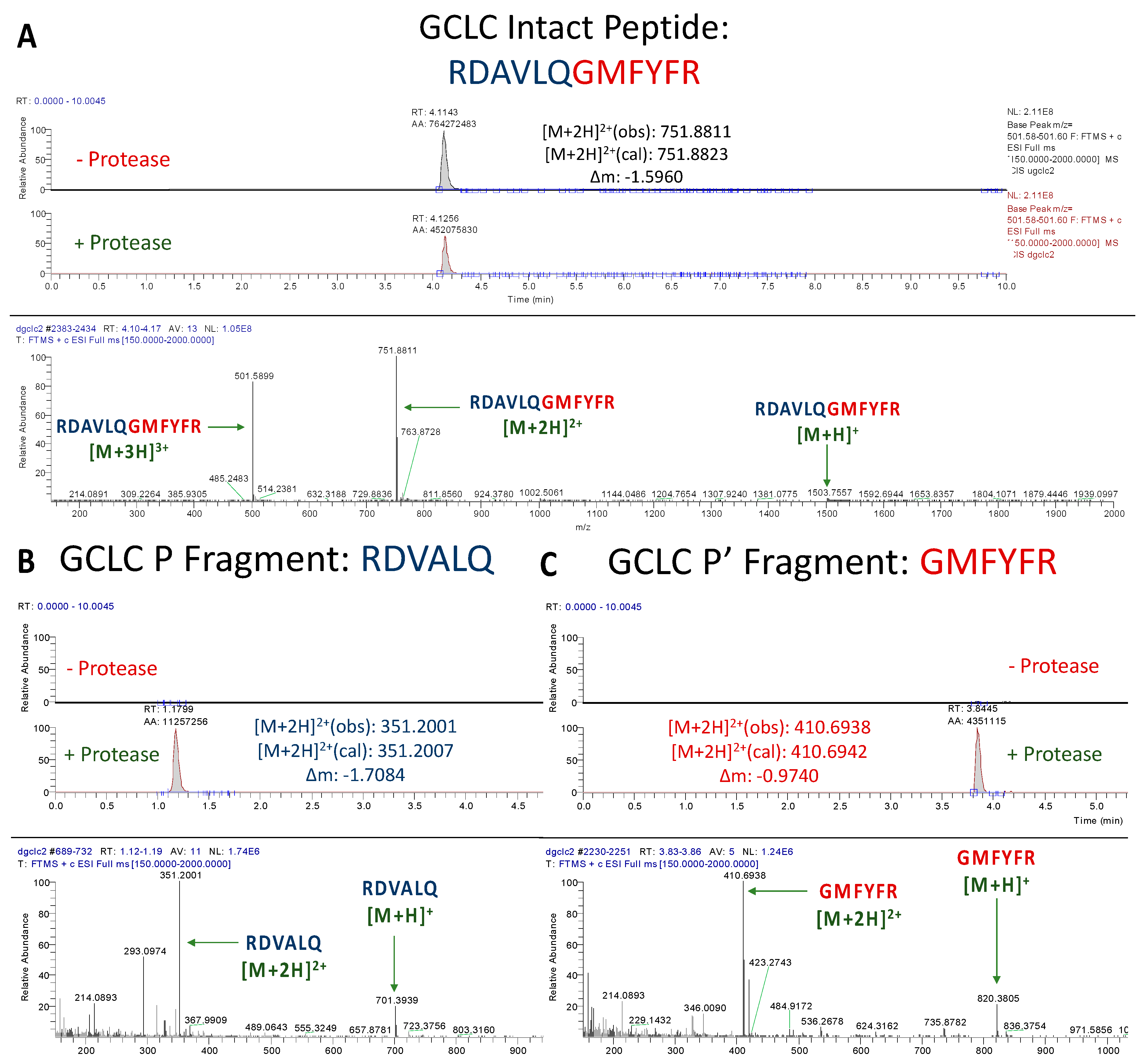
3.1.3. Assessment of GCLC Peptide Cleavage
3.1.4. Assessment of SelenoP Peptide Cleavage
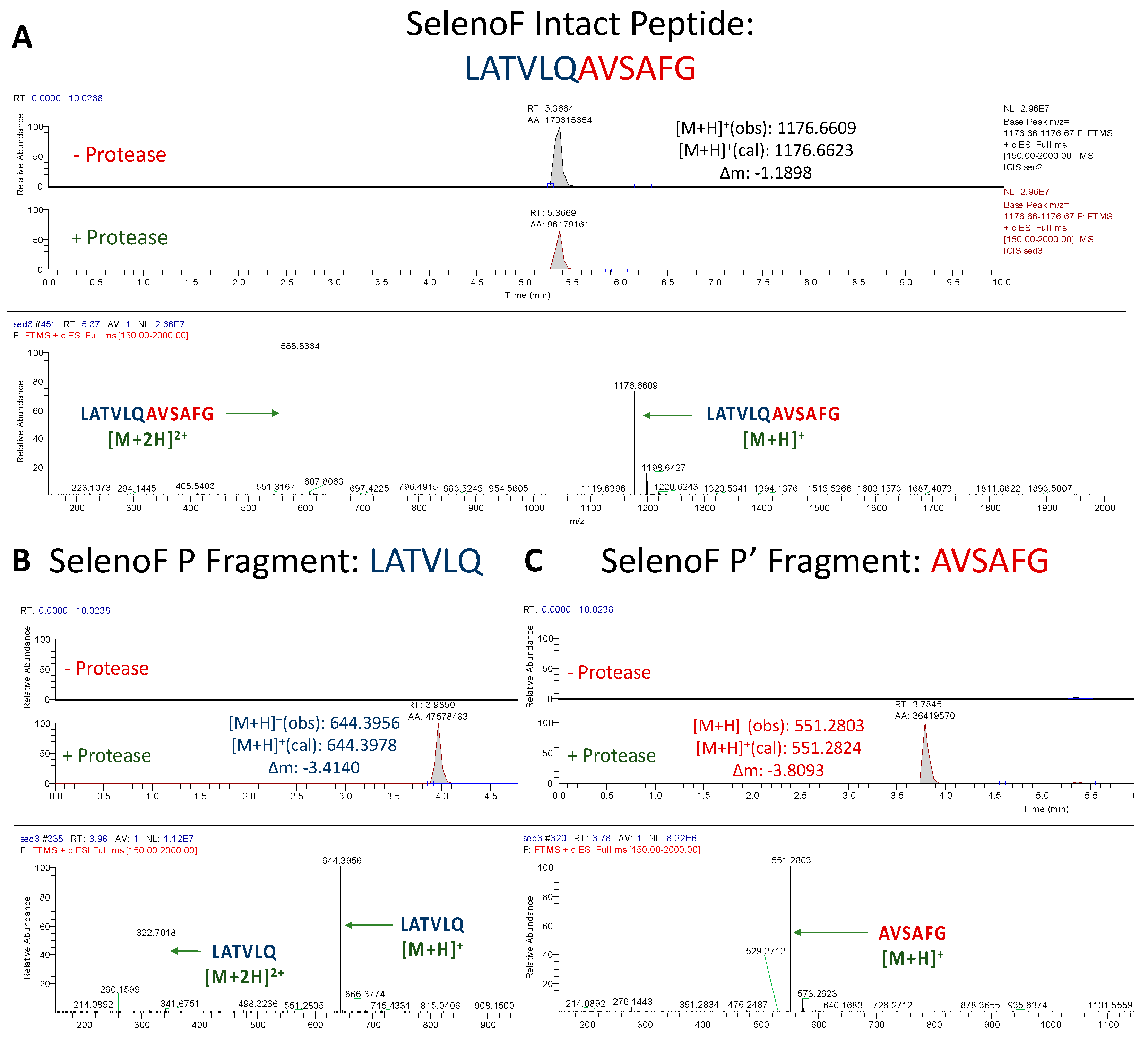
3.1.5. Assessment of SelenoF Peptide Cleavage
3.1.6. Assessment of GLRX-1 Peptide Cleavage
3.1.7. Assessment of GPX1 Peptide Cleavage
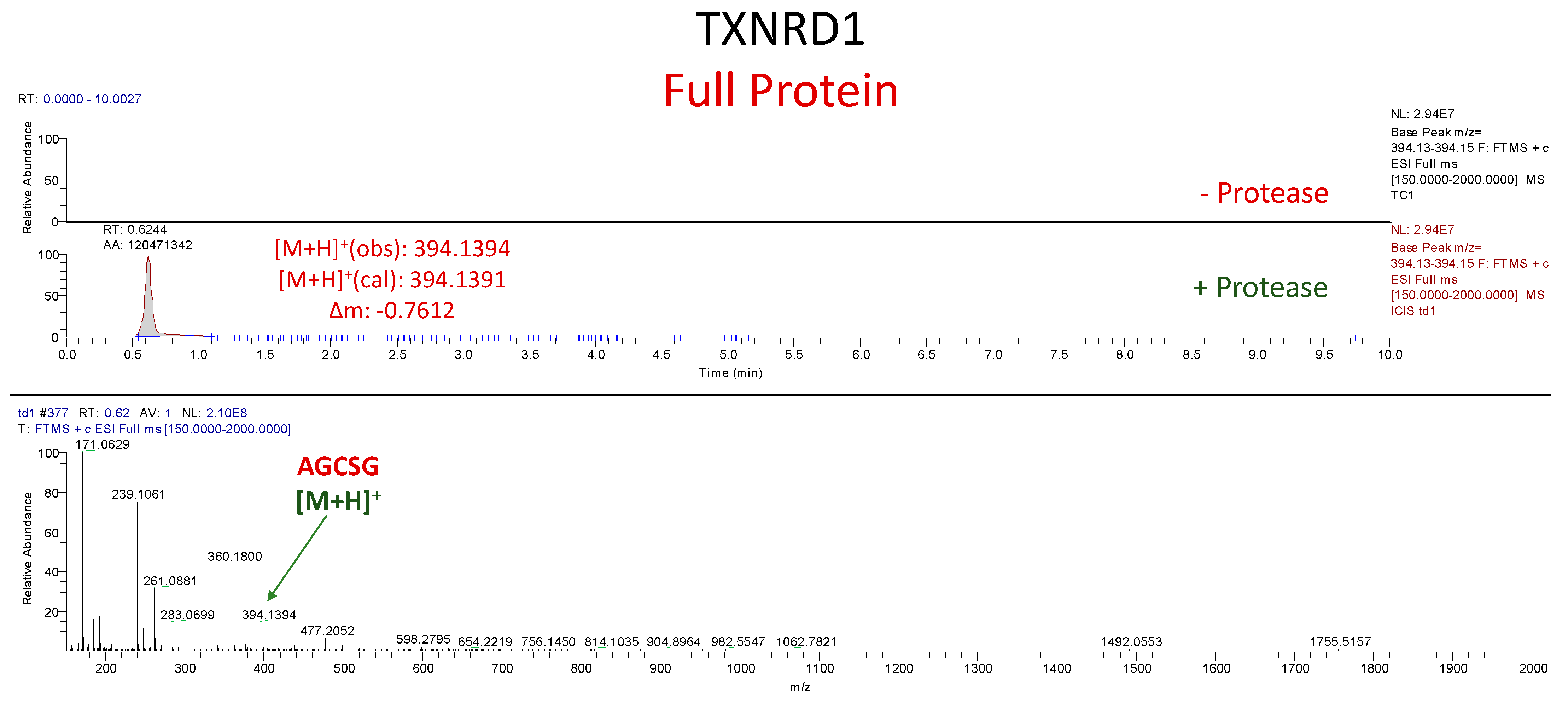
3.2. In Vitro Cleavage of Recombinant TXNRD1 Protein by SARS-CoV-2 Mpro
Assessment of TXNRD1 Protein Cleavage
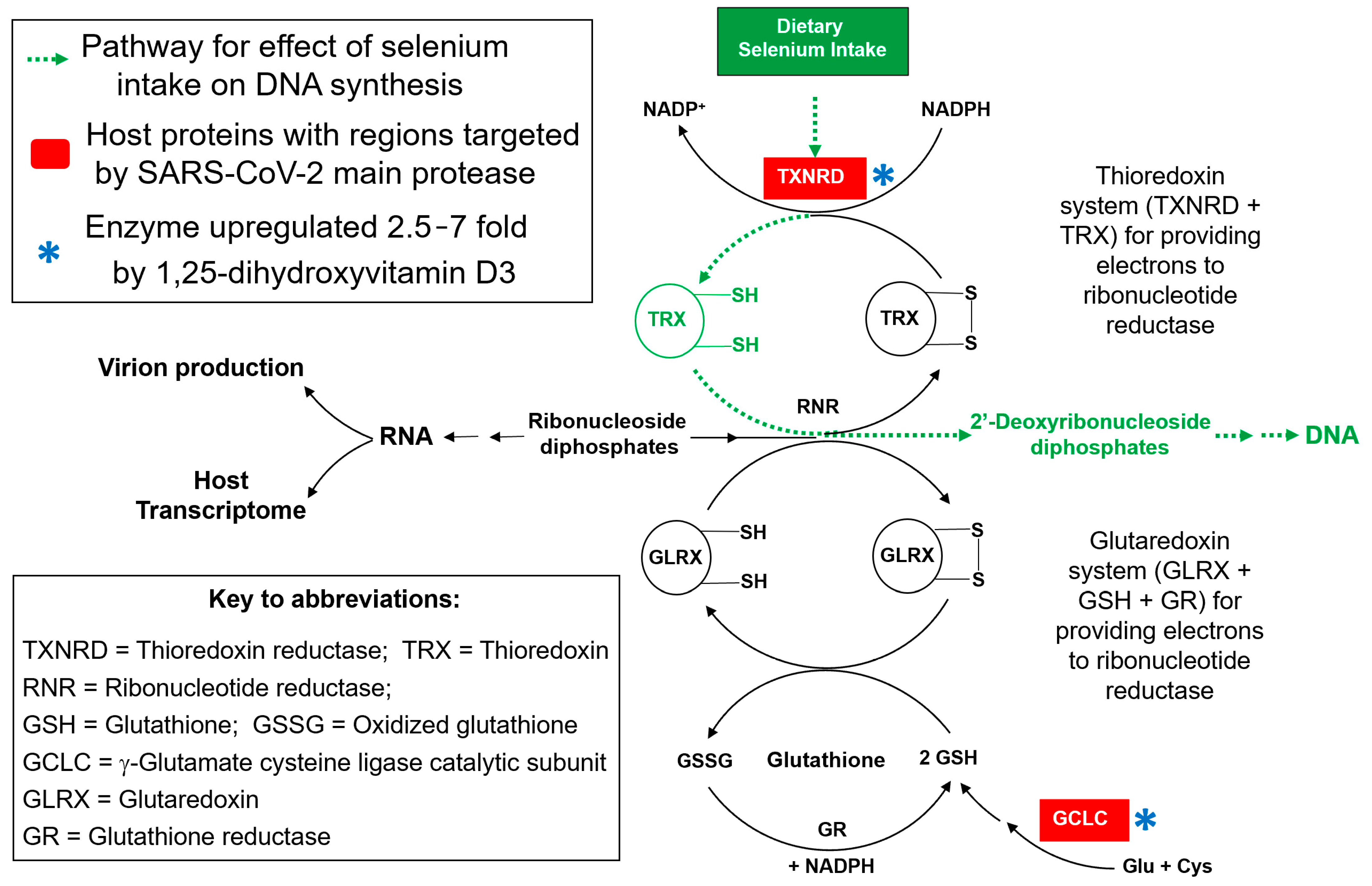
4. Discussion

5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int (accessed on 3 May 2021).
- Wilhelm, A.; Widera, M.; Grikscheit, K.; Toptan, T.; Schenk, B.; Pallas, C.; Metzler, M.; Kohmer, N.; Hoehl, S.; Marschalek, R.; et al. Limited Neutralisation of the SARS-CoV-2 Omicron Subvariants BA.1 and BA.2 by Convalescent and Vaccine Serum and Monoclonal Antibodies. eBioMedicine 2022, 82, 104158. [Google Scholar] [CrossRef] [PubMed]
- Bellanti, J.A. The Long COVID Syndrome: A Conundrum for the Allergist/Immunologist. Allergy Asthma Proc. 2022, 43, 368–374. [Google Scholar] [CrossRef] [PubMed]
- De Niet, S.; Trémège, M.; Coffiner, M.; Rousseau, A.-F.; Calmes, D.; Frix, A.-N.; Gester, F.; Delvaux, M.; Dive, A.-F.; Guglielmi, E.; et al. Positive Effects of Vitamin D Supplementation in Patients Hospitalized for COVID-19: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2022, 14, 3048. [Google Scholar] [CrossRef] [PubMed]
- Dror, A.A.; Morozov, N.; Daoud, A.; Namir, Y.; Yakir, O.; Shachar, Y.; Lifshitz, M.; Segal, E.; Fisher, L.; Mizrachi, M.; et al. Pre-Infection 25-Hydroxyvitamin D3 Levels and Association with Severity of COVID-19 Illness. PLoS ONE 2022, 17, e0263069. [Google Scholar] [CrossRef] [PubMed]
- Merzon, E.; Tworowski, D.; Gorohovski, A.; Vinker, S.; Golan Cohen, A.; Green, I.; Frenkel-Morgenstern, M. Low Plasma 25(OH) Vitamin D Level Is Associated with Increased Risk of COVID-19 Infection: An Israeli Population-based Study. FEBS J. 2020, 287, 3693–3702. [Google Scholar] [CrossRef]
- Jolliffe, D.A.; Holt, H.; Greenig, M.; Talaei, M.; Perdek, N.; Pfeffer, P.; Vivaldi, G.; Maltby, S.; Symons, J.; Barlow, N.L.; et al. Effect of a Test-and-Treat Approach to Vitamin D Supplementation on Risk of All Cause Acute Respiratory Tract Infection and COVID-19: Phase 3 Randomised Controlled Trial (CORONAVIT). BMJ 2022, 378, e071230. [Google Scholar] [CrossRef]
- Brunvoll, S.H.; Nygaard, A.B.; Ellingjord-Dale, M.; Holland, P.; Istre, M.S.; Kalleberg, K.T.; Søraas, C.L.; Holven, K.B.; Ulven, S.M.; Hjartåker, A.; et al. Prevention of COVID-19 and Other Acute Respiratory Infections with Cod Liver Oil Supplementation, a Low Dose Vitamin D Supplement: Quadruple Blinded, Randomised Placebo Controlled Trial. BMJ 2022, 378, e071245. [Google Scholar] [CrossRef]
- Zhang, J.; Taylor, E.W.; Bennett, K.; Saad, R.; Rayman, M.P. Association between Regional Selenium Status and Reported Outcome of COVID-19 Cases in China. Am. J. Clin. Nutr. 2020, 111, 1297–1299. [Google Scholar] [CrossRef]
- Moghaddam, A.; Heller, R.; Sun, Q.; Seelig, J.; Cherkezov, A.; Seibert, L.; Hackler, J.; Seemann, P.; Diegmann, J.; Pilz, M.; et al. Selenium Deficiency Is Associated with Mortality Risk from COVID-19. Nutrients 2020, 12, 2098. [Google Scholar] [CrossRef]
- Im, J.H.; Je, Y.S.; Baek, J.; Chung, M.-H.; Kwon, H.Y.; Lee, J.-S. Nutritional Status of Patients with COVID-19. Int. J. Infect. Dis. 2020, 100, 390–393. [Google Scholar] [CrossRef]
- Bermano, G.; Méplan, C.; Mercer, D.K.; Hesketh, J.E. Selenium and Viral Infection: Are There Lessons for COVID-19? Br. J. Nutr. 2021, 125, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Hiffler, L.; Rakotoambinina, B. Selenium and RNA Virus Interactions: Potential Implications for SARS-CoV-2 Infection (COVID-19). Front. Nutr. 2020, 7, 164. [Google Scholar] [CrossRef]
- Zhang, J.; Saad, R.; Taylor, E.W.; Rayman, M.P. Selenium and Selenoproteins in Viral Infection with Potential Relevance to COVID-19. Redox Biol. 2020, 37, 101715. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, X.; Ma, J.; Mu, Y.; Wang, Y.; Yang, S.; Wu, Y.; Wu, F.; Zhou, Y. Selenium (Se) Plays a Key Role in the Biological Effects of Some Viruses: Implications for COVID-19. Environ. Res. 2021, 196, 110984. [Google Scholar] [CrossRef]
- Martinez, S.S.; Huang, Y.; Acuna, L.; Laverde, E.; Trujillo, D.; Barbieri, M.A.; Tamargo, J.; Campa, A.; Baum, M.K. Role of Selenium in Viral Infections with a Major Focus on SARS-CoV-2. IJMS 2021, 23, 280. [Google Scholar] [CrossRef] [PubMed]
- Fakhrolmobasheri, M.; Mazaheri-Tehrani, S.; Kieliszek, M.; Zeinalian, M.; Abbasi, M.; Karimi, F.; Mozafari, A.M. COVID-19 and Selenium Deficiency: A Systematic Review. Biol. Trace Elem. Res. 2022, 200, 3945–3956. [Google Scholar] [CrossRef]
- Schomburg, L. Selenium Deficiency in COVID-19—A Possible Long-Lasting Toxic Relationship. Nutrients 2022, 14, 283. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P.; Taylor, E.W.; Zhang, J. The Relevance of Selenium to Viral Disease with Special Reference to SARS-CoV-2 and COVID-19. Proc. Nutr. Soc. 2022, 19, 1–12. [Google Scholar] [CrossRef]
- Sheffy, B.E.; Schultz, R.D. Nutrition and the Immune Response. Cornell Vet. 1978, 68 (Suppl. S7), 48–61. [Google Scholar] [PubMed]
- Spallholz, J.E. Selenium and Glutathione Peroxidase: Essential Nutrient and Antioxidant Component of the Immune System. Adv. Exp. Med. Biol. 1990, 262, 145–158. [Google Scholar] [CrossRef]
- Turner, R.J.; Finch, J.M. Selenium and the Immune Response. Proc. Nutr. Soc. 1991, 50, 275–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenzie, R.C.; Rafferty, T.S.; Beckett, G.J. Selenium: An Essential Element for Immune Function. Immunol. Today 1998, 19, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Hoffmann, P.R. Selenoproteins as Regulators of T Cell Proliferation, Differentiation, and Metabolism. Semin. Cell Dev. Biol. 2021, 115, 54–61. [Google Scholar] [CrossRef]
- Avery, J.; Hoffmann, P. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Huang, J.; Sun, Y.; Stubbs, D.; He, J.; Li, W.; Wang, F.; Liu, Z.; Ruzicka, J.A.; Taylor, E.W.; et al. SARS-CoV-2 Suppresses MRNA Expression of Selenoproteins Associated with Ferroptosis, Endoplasmic Reticulum Stress and DNA Synthesis. Food Chem. Toxicol. 2021, 153, 112286. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.W.; Radding, W. Understanding Selenium and Glutathione as Antiviral Factors in COVID-19: Does the Viral Mpro Protease Target Host Selenoproteins and Glutathione Synthesis? Front. Nutr. 2020, 7, 143. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, N.D.; Aceves, N.M.; Liu, J.L.; Compton, J.R.; Leary, D.H.; Freitas, B.T.; Pegan, S.D.; Doctor, K.Z.; Wu, F.Y.; Hu, X.; et al. The SARS-CoV-2 SSHHPS Recognized by the Papain-like Protease. ACS Infect. Dis. 2021, 7, 1483–1502. [Google Scholar] [CrossRef]
- Miczi, M.; Golda, M.; Kunkli, B.; Nagy, T.; Tőzsér, J.; Mótyán, J.A. Identification of Host Cellular Protein Substrates of SARS-COV-2 Main Protease. Int. J. Mol. Sci. 2020, 21, 9523. [Google Scholar] [CrossRef]
- Moustaqil, M.; Ollivier, E.; Chiu, H.-P.; Van Tol, S.; Rudolffi-Soto, P.; Stevens, C.; Bhumkar, A.; Hunter, D.J.B.; Freiberg, A.N.; Jacques, D.; et al. SARS-CoV-2 Proteases PLpro and 3CLpro Cleave IRF3 and Critical Modulators of Inflammatory Pathways (NLRP12 and TAB1): Implications for Disease Presentation across Species. Emerg. Microbes Infect. 2021, 10, 178–195. [Google Scholar] [CrossRef]
- Ruzicka, J.A. Identification of the Antithrombotic Protein S as a Potential Target of the SARS-CoV-2 Papain-like Protease. Throm. Res. 2020, 196, 257–259. [Google Scholar] [CrossRef]
- Blanco, R.; Carrasco, L.; Ventoso, I. Cell Killing by HIV-1 Protease. J. Biol. Chem. 2003, 278, 1086–1093. [Google Scholar] [CrossRef] [Green Version]
- Taylor, E.W. RNA Viruses vs. DNA Synthesis: A General Viral Strategy That May Contribute to the Protective Antiviral Effects of Selenium. Preprints 2020, 2020060069. [Google Scholar] [CrossRef]
- Kiemer, L.; Lund, O.; Brunak, S.; Blom, N. Coronavirus 3CLpro Proteinase Cleavage Sites: Possible Relevance to SARS Virus Pathology. BMC Bioinform. 2004, 5, 72. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Leier, A.; Liu, Q.; Wang, Y.; Xiang, D.; Akutsu, T.; Webb, G.I.; Smith, A.I.; Marquez-Lago, T.; Li, J.; et al. Procleave: Predicting Protease-Specific Substrate Cleavage Sites by Combining Sequence and Structural Information. Genom. Proteom. Bioinform. 2020, 18, 52–64. [Google Scholar] [CrossRef]
- Song, J.; Tan, H.; Perry, A.J.; Akutsu, T.; Webb, G.I.; Whisstock, J.C.; Pike, R.N. PROSPER: An Integrated Feature-Based Tool for Predicting Protease Substrate Cleavage Sites. PLoS ONE 2012, 7, e50300. [Google Scholar] [CrossRef] [Green Version]
- Lima, S.T.; Fallon, T.R.; Cordoza, J.L.; Chekan, J.R.; Delbaje, E.; Hopiavuori, A.R.; Alvarenga, D.O.; Wood, S.M.; Luhavaya, H.; Baumgartner, J.T.; et al. Biosynthesis of Guanitoxin Enables Global Environmental Detection in Freshwater Cyanobacteria. J. Am. Chem. Soc. 2022, 144, 9372–9379. [Google Scholar] [CrossRef]
- Fan, K.; Ma, L.; Han, X.; Liang, H.; Wei, P.; Liu, Y.; Lai, L. The Substrate Specificity of SARS Coronavirus 3C-like Proteinase. Biochem. Biophys. Res. Commun. 2005, 329, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Grum-Tokars, V.; Ratia, K.; Begaye, A.; Baker, S.C.; Mesecar, A.D. Evaluating the 3C-like Protease Activity of SARS-Coronavirus: Recommendations for Standardized Assays for Drug Discovery. Virus Res. 2008, 133, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Mengist, H.M.; Dilnessa, T.; Jin, T. Structural Basis of Potential Inhibitors Targeting SARS-CoV-2 Main Protease. Front. Chem. 2021, 9, 622898. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 Protein Interaction Map Reveals Targets for Drug Repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Orian, L.; Mauri, P.; Roveri, A.; Toppo, S.; Benazzi, L.; Bosello-Travain, V.; De Palma, A.; Maiorino, M.; Miotto, G.; Zaccarin, M.; et al. Selenocysteine Oxidation in Glutathione Peroxidase Catalysis: An MS-Supported Quantum Mechanics Study. Free Rad. Biol. Med. 2015, 87, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-W.; Lin, K.-H.; Hsieh, T.-H.; Shiu, S.-Y.; Li, J.-Y. Severe Acute Respiratory Syndrome Coronavirus 3C-like Protease-Induced Apoptosis. FEMS Immonol. Med. Microbiol. 2006, 46, 375–380. [Google Scholar] [CrossRef] [Green Version]
- Schomburg, L. Selenoprotein P—Selenium Transport Protein, Enzyme and Biomarker of Selenium Status. Free Rad. Biol. Med. 2022, 191, 150–163. [Google Scholar] [CrossRef]
- Kumar, P.; Osahon, O.; Vides, D.B.; Hanania, N.; Minard, C.G.; Sekhar, R.V. Severe Glutathione Deficiency, Oxidative Stress and Oxidant Damage in Adults Hospitalized with COVID-19: Implications for GlyNAC (Glycine and N-Acetylcysteine) Supplementation. Antioxidants 2021, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Silvagno, F.; Vernone, A.; Pescarmona, G.P. The Role of Glutathione in Protecting against the Severe Inflammatory Response Triggered by COVID-19. Antioxidants 2020, 9, 624. [Google Scholar] [CrossRef] [PubMed]
- Labarrere, C.A.; Kassab, G.S. Glutathione Deficiency in the Pathogenesis of SARS-CoV-2 Infection and Its Effects upon the Host Immune Response in Severe COVID-19 Disease. Front. Microbiol. 2022, 13, 979719. [Google Scholar] [CrossRef] [PubMed]
- Polonikov, A. Endogenous Deficiency of Glutathione as the Most Likely Cause of Serious Manifestations and Death in COVID-19 Patients. ACS Infect. Dis. 2020, 6, 1558–1562. [Google Scholar] [CrossRef]
- Ghezzi, P. Role of Glutathione in Immunity and Inflammation in the Lung. IJGM 2011, 4, 105. [Google Scholar] [CrossRef] [Green Version]
- Fraternale, A.; Paoletti, M.; Casabianca, A.; Oiry, J.; Clayette, P.; Vogel, J., Jr.; Cinatl, J.; Palamara, A.; Sgarbanti, R.; Garaci, E.; et al. Antiviral and Immunomodulatory Properties of New Pro-Glutathione (GSH) Molecules. CMC 2006, 13, 1749–1755. [Google Scholar] [CrossRef]
- Bartolini, D.; Stabile, A.M.; Bastianelli, S.; Giustarini, D.; Pierucci, S.; Busti, C.; Vacca, C.; Gidari, A.; Francisci, D.; Castronari, R.; et al. SARS-CoV2 Infection Impairs the Metabolism and Redox Function of Cellular Glutathione. Redox Biol. 2021, 45, 102041. [Google Scholar] [CrossRef]
- van der Horst, D.; Carter-Timofte, M.; van Grevenynghe, J.; Laguette, N.; Dinkova-Kostova, A.; Olagnier, D. Regulation of Innate Immunity by Nrf2. Curr. Opin. Immunol. 2022, 78, 102247. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Liu, M.; Ni, J.; Tian, J. Role of Selenoprotein F in Protein Folding and Secretion: Potential Involvement in Human Disease. Nutrients 2018, 10, 1619. [Google Scholar] [CrossRef] [Green Version]
- Grishin, A.M.; Dolgova, N.V.; Landreth, S.; Fisette, O.; Pickering, I.J.; George, G.N.; Falzarano, D.; Cygler, M. Disulfide Bonds Play a Critical Role in the Structure and Function of the Receptor-Binding Domain of the SARS-CoV-2 Spike Antigen. J. Mol. Biol. 2022, 434, 167357. [Google Scholar] [CrossRef] [PubMed]
- Murae, M.; Shimizu, Y.; Yamamoto, Y.; Kobayashi, A.; Houri, M.; Inoue, T.; Irie, T.; Gemba, R.; Kondo, Y.; Nakano, Y.; et al. The Function of SARS-CoV-2 Spike Protein Is Impaired by Disulfide-Bond Disruption with Mutation at Cysteine-488 and by Thiol-Reactive N-Acetyl-Cysteine and Glutathione. Biochem. Biophys. Res. Commun. 2022, 597, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Manček-Keber, M.; Hafner-Bratkovič, I.; Lainšček, D.; Benčina, M.; Govednik, T.; Orehek, S.; Plaper, T.; Jazbec, V.; Bergant, V.; Grass, V.; et al. Disruption of Disulfides within RBD of SARS-CoV-2 Spike Protein Prevents Fusion and Represents a Target for Viral Entry Inhibition by Registered Drugs. FASEB J. 2021, 35, e21651. [Google Scholar] [CrossRef]
- Gibbons, J.B.; Norton, E.C.; McCullough, J.S.; Meltzer, D.O.; Lavigne, J.; Fiedler, V.C.; Gibbons, R.D. Association between Vitamin D Supplementation and COVID-19 Infection and Mortality. Sci. Rep. 2022, 12, 19397. [Google Scholar] [CrossRef]
- Aquilano, K.; Baldelli, S.; Ciriolo, M.R. Glutathione: New Roles in Redox Signaling for an Old Antioxidant. Front. Pharmacol. 2014, 5, 196. [Google Scholar] [CrossRef] [Green Version]
- Dröge, W.; Breitkreutz, R. Glutathione and Immune Function. Proc. Nutr. Soc. 2000, 59, 595–600. [Google Scholar] [CrossRef]
- Jia, J.; Xu, G.; Zhu, D.; Liu, H.; Zeng, X.; Li, L. Advances in the Functions of Thioredoxin System in Central Nervous System Diseases. Antioxid. Redox Signal. 2022, 38, 425–441. [Google Scholar] [CrossRef]
- Stancill, J.S.; Hansen, P.A.; Mathison, A.J.; Schmidt, E.E.; Corbett, J.A. Deletion of Thioredoxin Reductase Disrupts Redox Homeostasis and Impairs β-Cell Function. Function 2022, 3, zqac034. [Google Scholar] [CrossRef]
- Resnick, S.J.; Iketani, S.; Hong, S.J.; Zask, A.; Liu, H.; Kim, S.; Melore, S.; Lin, F.-Y.; Nair, M.S.; Huang, Y.; et al. Inhibitors of Coronavirus 3CL Proteases Protect Cells from Protease-Mediated Cytotoxicity. J. Virol. 2021, 95, e02374-20. [Google Scholar] [CrossRef]
- Kovalenko, P.L.; Zhang, Z.; Cui, M.; Clinton, S.K.; Fleet, J.C. 1,25 Dihydroxyvitamin D-Mediated Orchestration of Anticancer, Transcript-Level Effects in the Immortalized, Non-Transformed Prostate Epithelial Cell Line, RWPE1. BMC Genom. 2010, 11, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, C.A.; Kawai, K.; Kupka, R.; Fawzi, W.W. Role of Selenium in HIV Infection. Nutr. Rev. 2010, 68, 671–681. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Al-Quraishy, S.; Dkhil, M.A.; Wunderlich, F.; Sies, H. Dietary Selenium in Adjuvant Therapy of Viral and Bacterial Infections. Adv. Nutr. 2015, 6, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Guillin, O.; Vindry, C.; Ohlmann, T.; Chavatte, L. Selenium, Selenoproteins and Viral Infection. Nutrients 2019, 11, 2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, E.W.; Ruzicka, J.A.; Premadasa, L.; Zhao, L. Cellular Selenoprotein mRNA Tethering via Antisense Interactions with Ebola and HIV-1 mRNAs May Impact Host Selenium Biochemistry. Curr. Top. Med. Chem. 2016, 16, 1530–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Premadasa, L.; Dailey, G.; Ruzicka, J.A.; Taylor, E.W. Selenium-Dependent Read Through of the Conserved 3′-Terminal UGA Stop Codon of HIV-1. Nef. Am. J. Biopharm. Pharm. Sci. 2021, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Dailey, G.P.; Premadasa, L.S.; Ruzicka, J.A.; Taylor, E.W. Inhibition of Selenoprotein Synthesis by Zika Virus May Contribute to Congenital Zika Syndrome and Microcephaly by Mimicking SELENOP Knockout and the Genetic Disease PCCA. BBA Adv. 2021, 1, 100023. [Google Scholar] [CrossRef]
- Gallardo, I.A. Identification of SARS-CoV-2 Main Protease Cleavage Sites in Host Cellular Selenoproteins and Glutathione-Related Proteins. Master’s Thesis, The University of North Carolina at Greensboro, Greensboro, NC, USA, 2022. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Composition |
|---|---|
| Blank 1 | 50 µL of buffer solution |
| SARS-CoV-2 Mpro Sample | 30 µL of SARS-CoV-2 Mpro solution and 20 µL of buffer solution |
| Positive Control Sample w/o Mpro | 10 µL of the positive control solution and 40 µL of the buffer solution |
| Positive Control Sample w/ Mpro | 10 µL of the positive control solution, 30 µL of the SARS-CoV-2 Mpro solution, and 10 µL of buffer |
| Proposed Peptide w/o Mpro | 10 µL of the proposed peptide solution and 40 µL of the buffer solution |
| Proposed Peptide w/ Mpro | 10 µL of the proposed peptide solution, 30 µL of the SARS-CoV-2 Mpro solution, and 10 µL of buffer |
| Blank 2 | 50 µL of buffer solution |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallardo, I.A.; Todd, D.A.; Lima, S.T.; Chekan, J.R.; Chiu, N.H.; Taylor, E.W. SARS-CoV-2 Main Protease Targets Host Selenoproteins and Glutathione Biosynthesis for Knockdown via Proteolysis, Potentially Disrupting the Thioredoxin and Glutaredoxin Redox Cycles. Antioxidants 2023, 12, 559. https://doi.org/10.3390/antiox12030559
Gallardo IA, Todd DA, Lima ST, Chekan JR, Chiu NH, Taylor EW. SARS-CoV-2 Main Protease Targets Host Selenoproteins and Glutathione Biosynthesis for Knockdown via Proteolysis, Potentially Disrupting the Thioredoxin and Glutaredoxin Redox Cycles. Antioxidants. 2023; 12(3):559. https://doi.org/10.3390/antiox12030559
Chicago/Turabian StyleGallardo, Ignacio A., Daniel A. Todd, Stella T. Lima, Jonathan R. Chekan, Norman H. Chiu, and Ethan Will Taylor. 2023. "SARS-CoV-2 Main Protease Targets Host Selenoproteins and Glutathione Biosynthesis for Knockdown via Proteolysis, Potentially Disrupting the Thioredoxin and Glutaredoxin Redox Cycles" Antioxidants 12, no. 3: 559. https://doi.org/10.3390/antiox12030559