
Structural Features Governing the Metabolic Stability of Tetraethyl-Substituted Nitroxides in Rat Liver Microsomes

Abstract
:1. Introduction
2. Material and Methods
2.1. Reagents
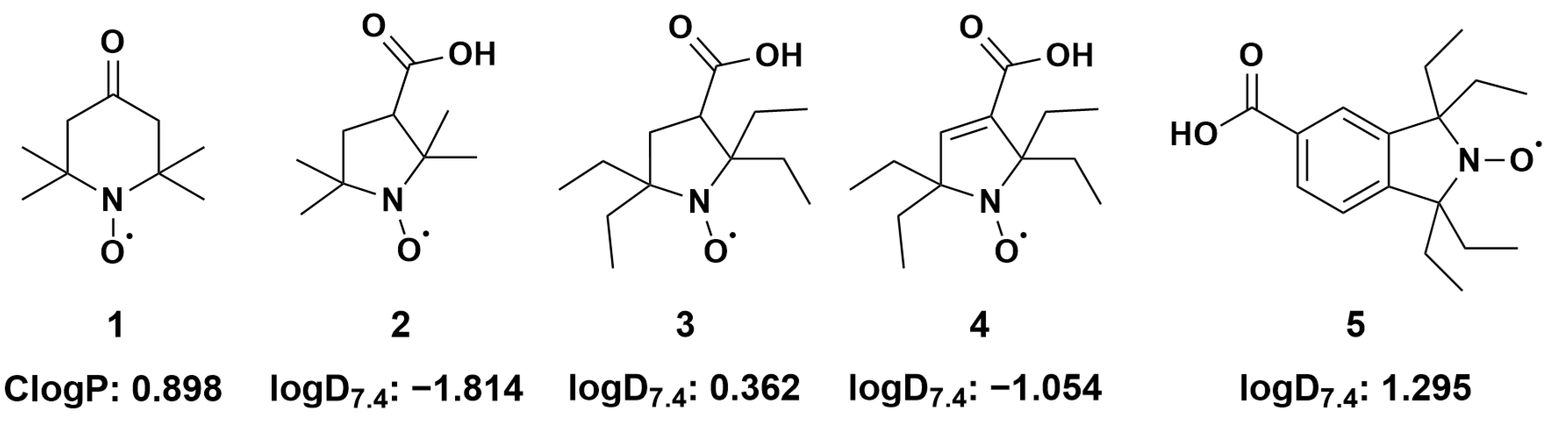
2.2. Estimation of Lipophilicity
2.3. Rat Liver Microsome Preparation
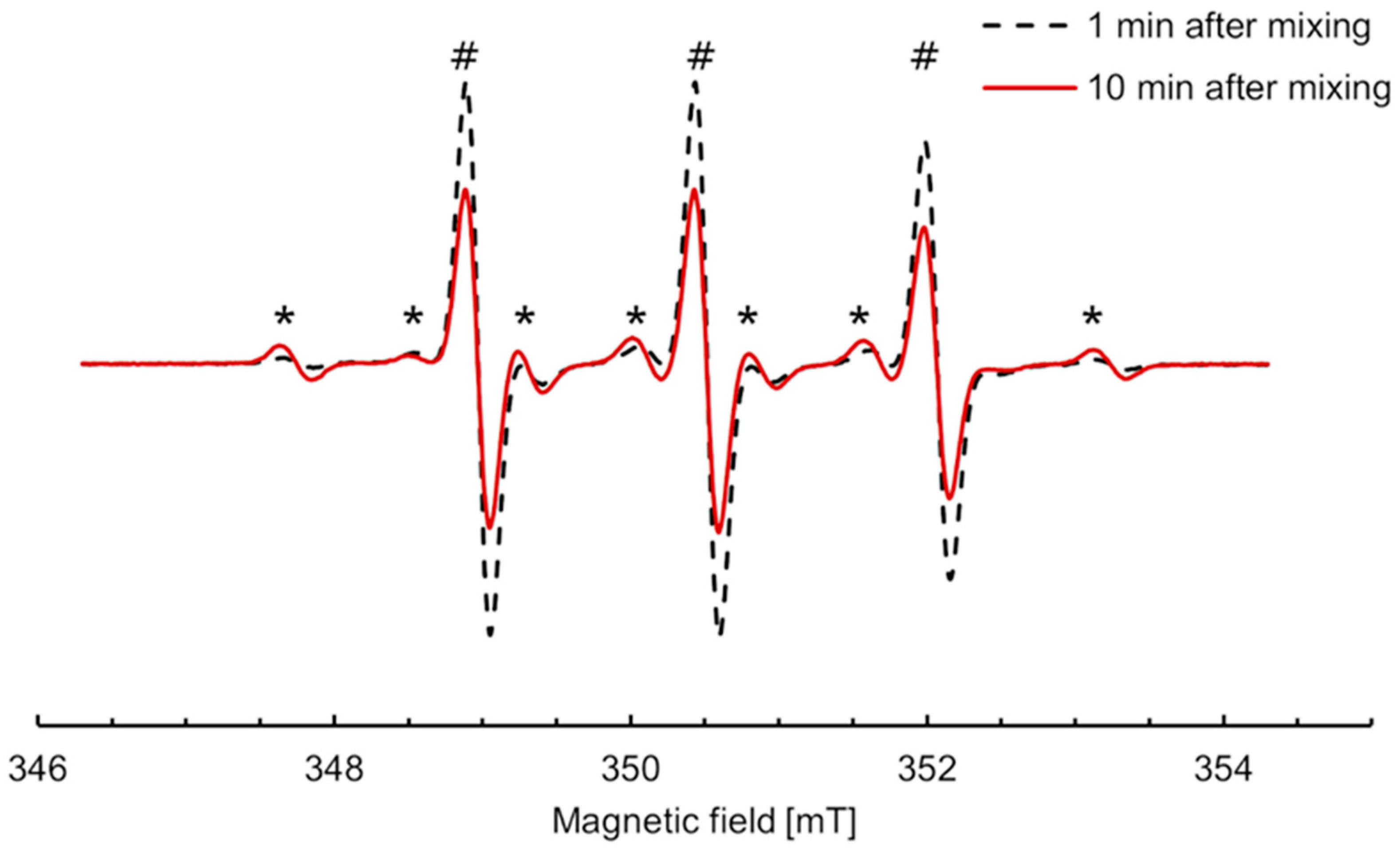
2.4. Reactions of Nitroxides with Ascorbic Acid Monitored by EPR Spectroscopy
2.5. Reactions of Nitroxides with RLM Monitored by EPR Spectroscopy
2.6. Calibration and Simulation of EPR Spectra
2.7. Statistical Analyses
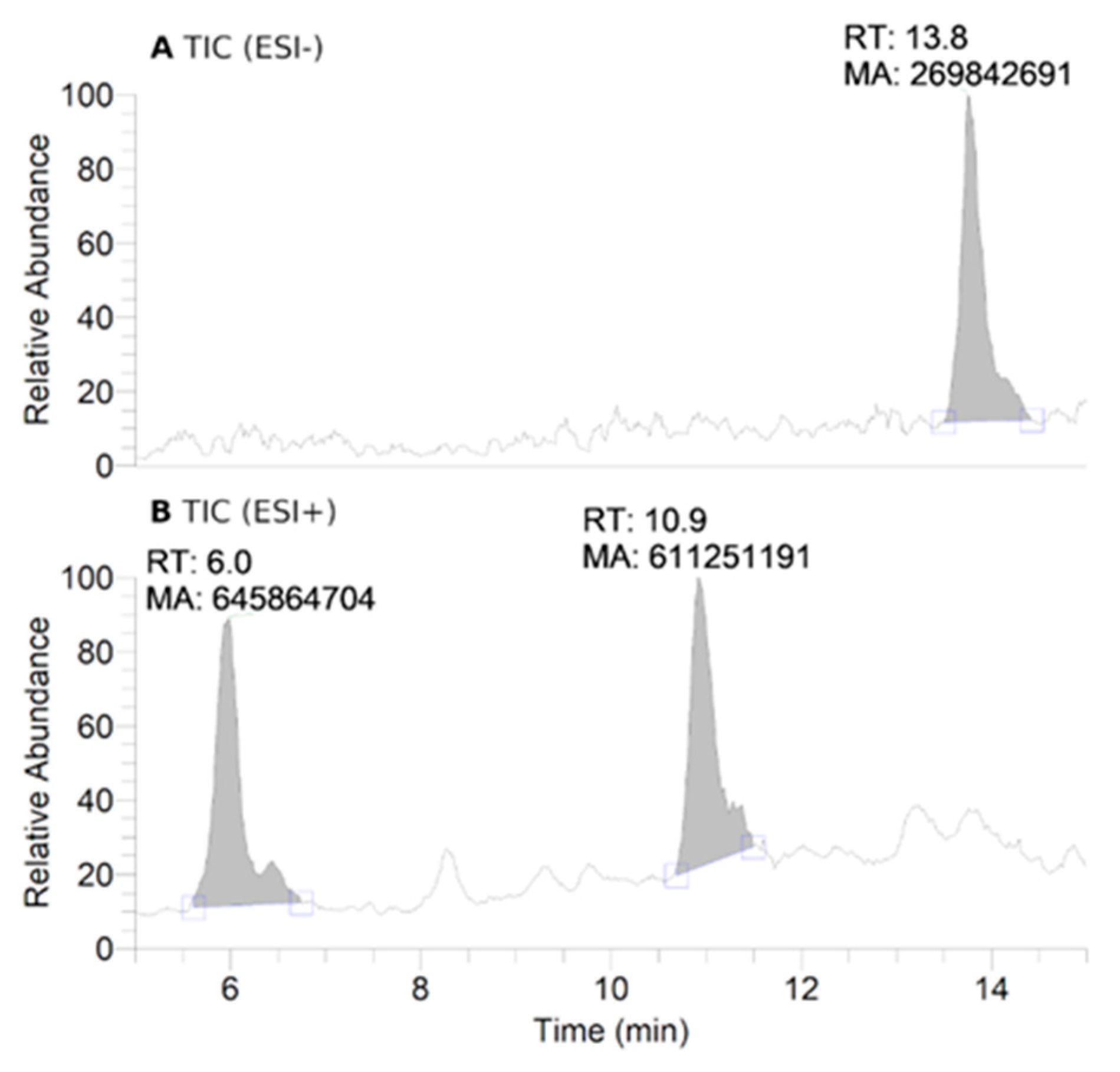
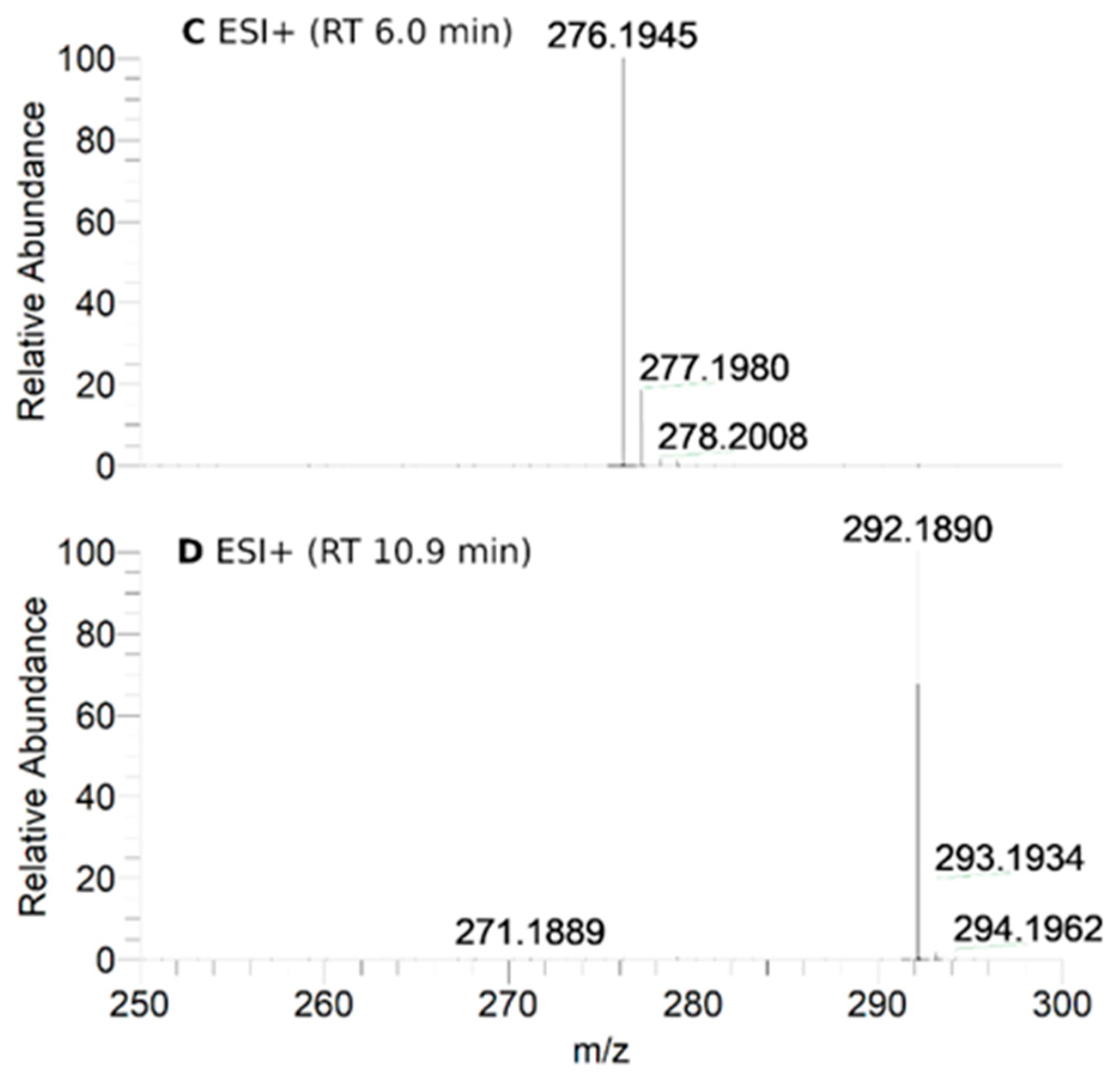
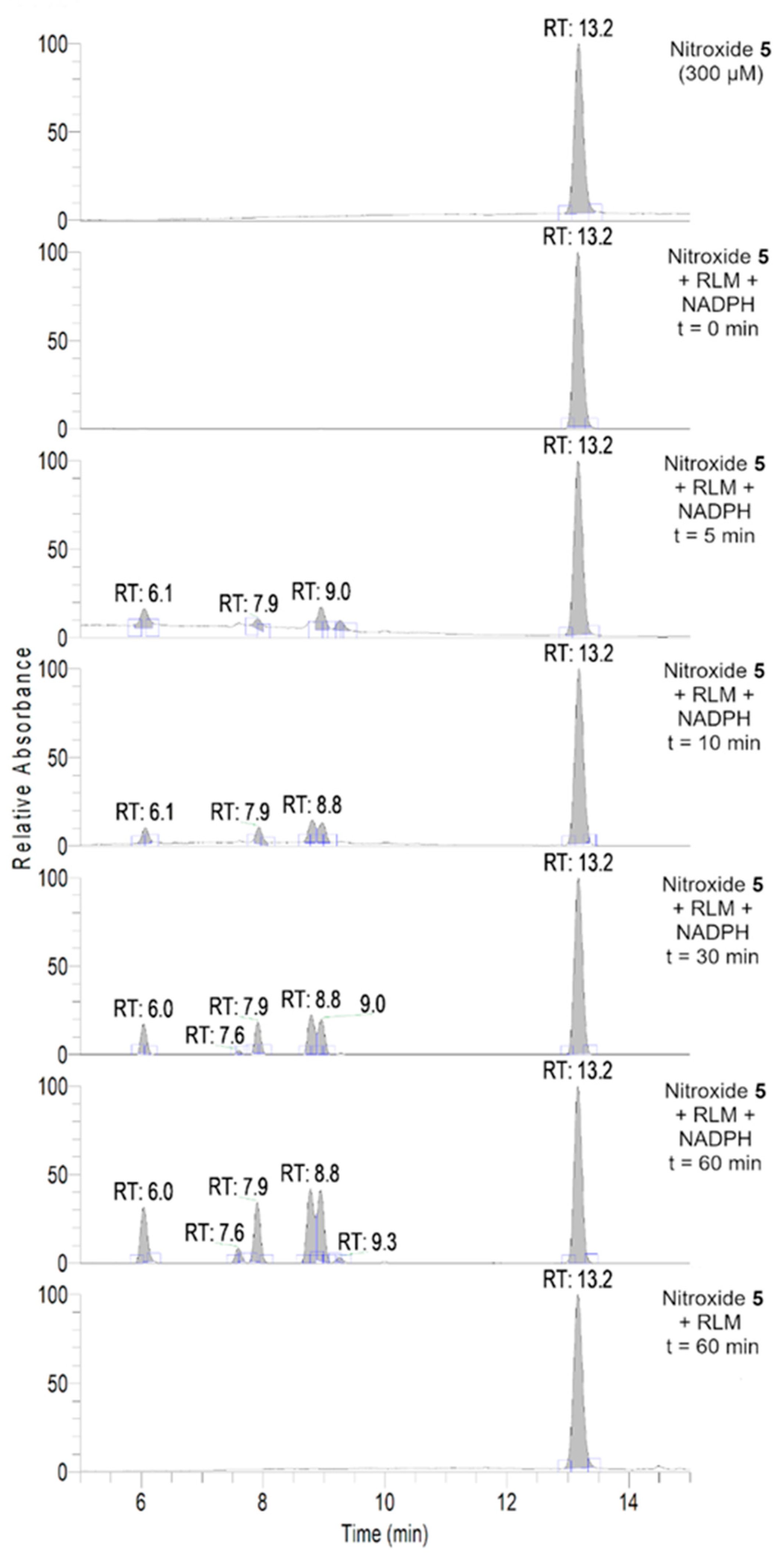
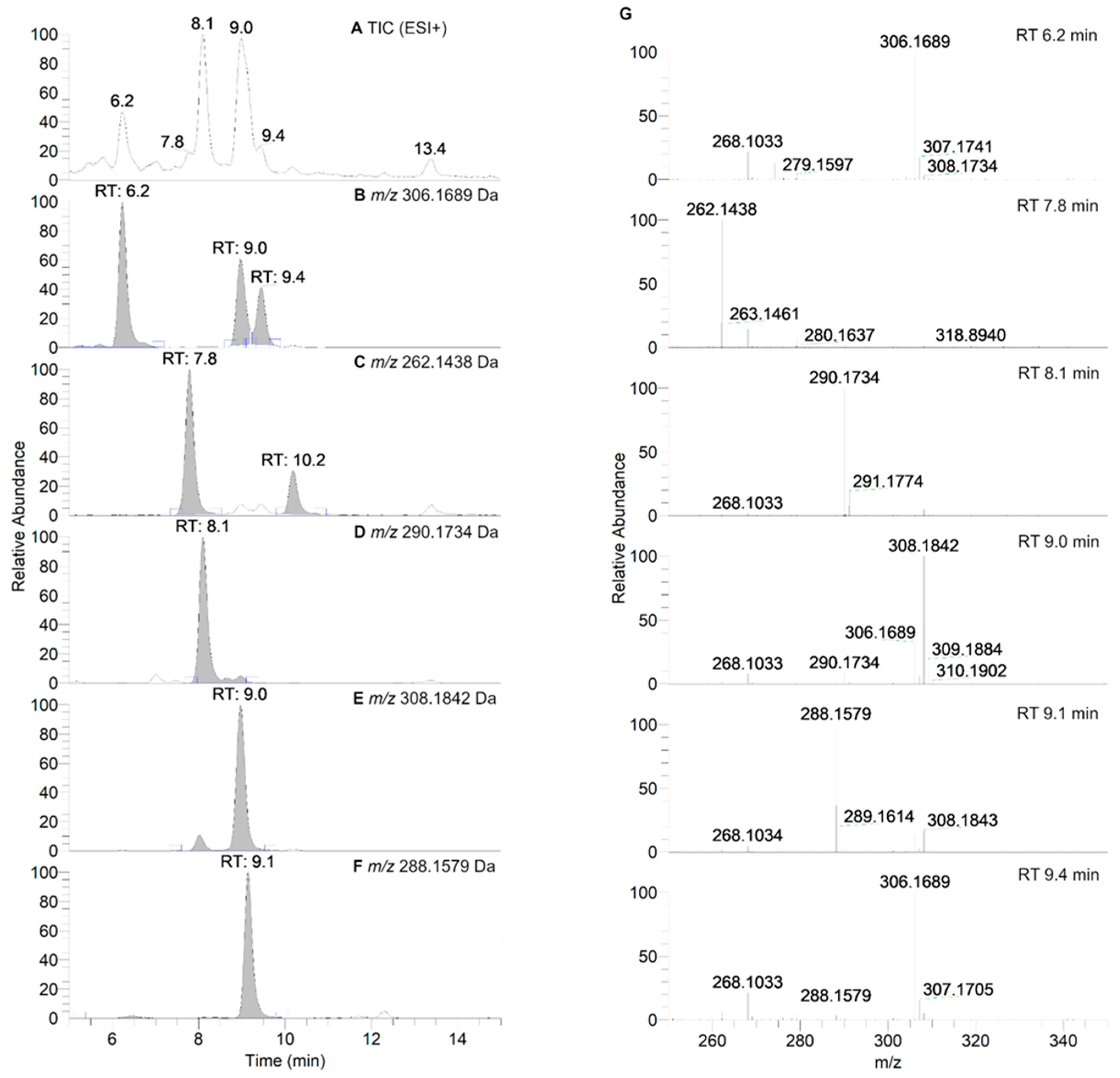
2.8. HPLC-HRMS Study of Isoindoline Nitroxide with RLM
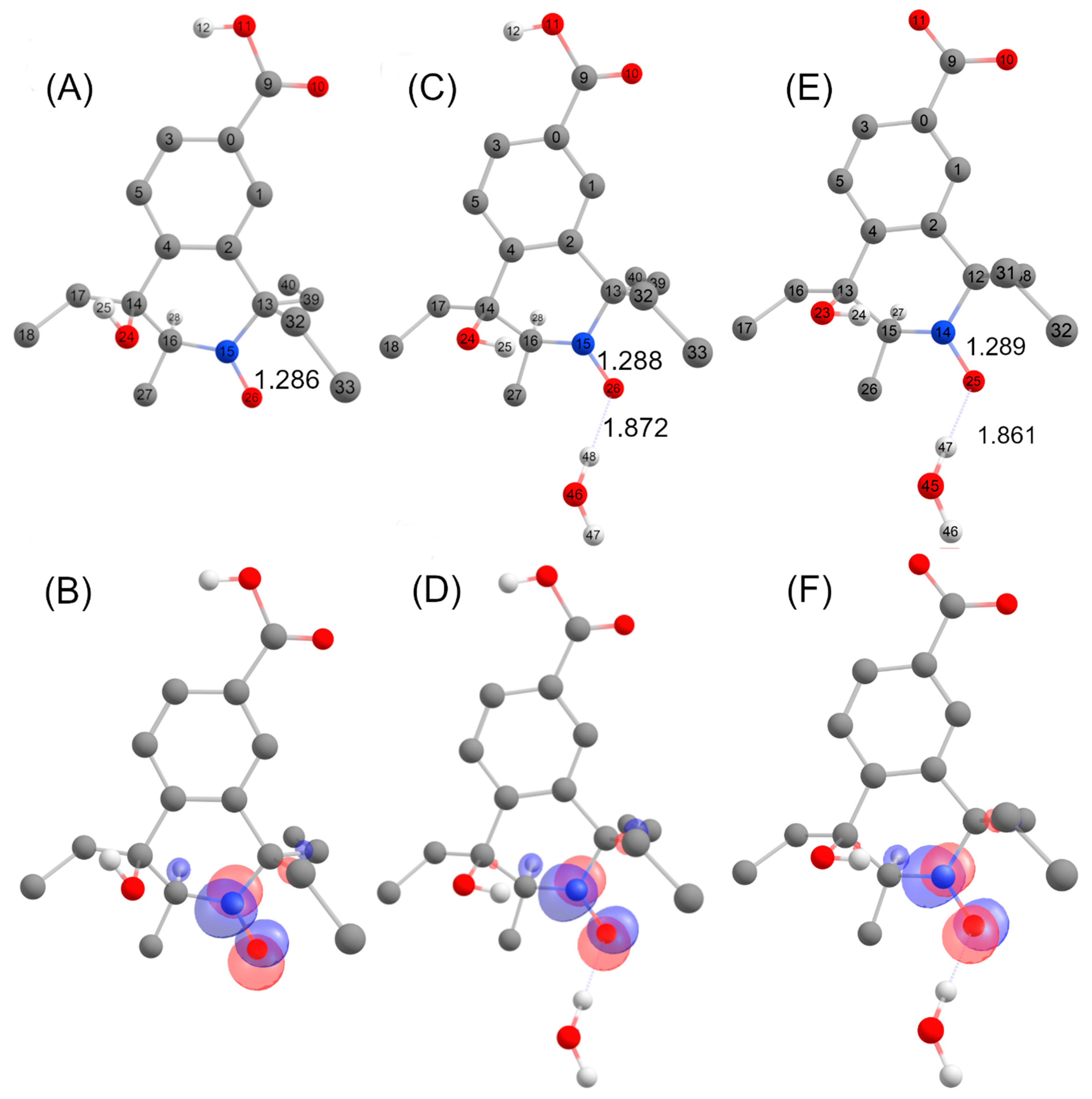
2.9. Computational Investigations
3. Results
3.1. Reactivity of Nitroxides with Ascorbate
3.2. Anaerobic Study of Nitroxides Incubated with RLM
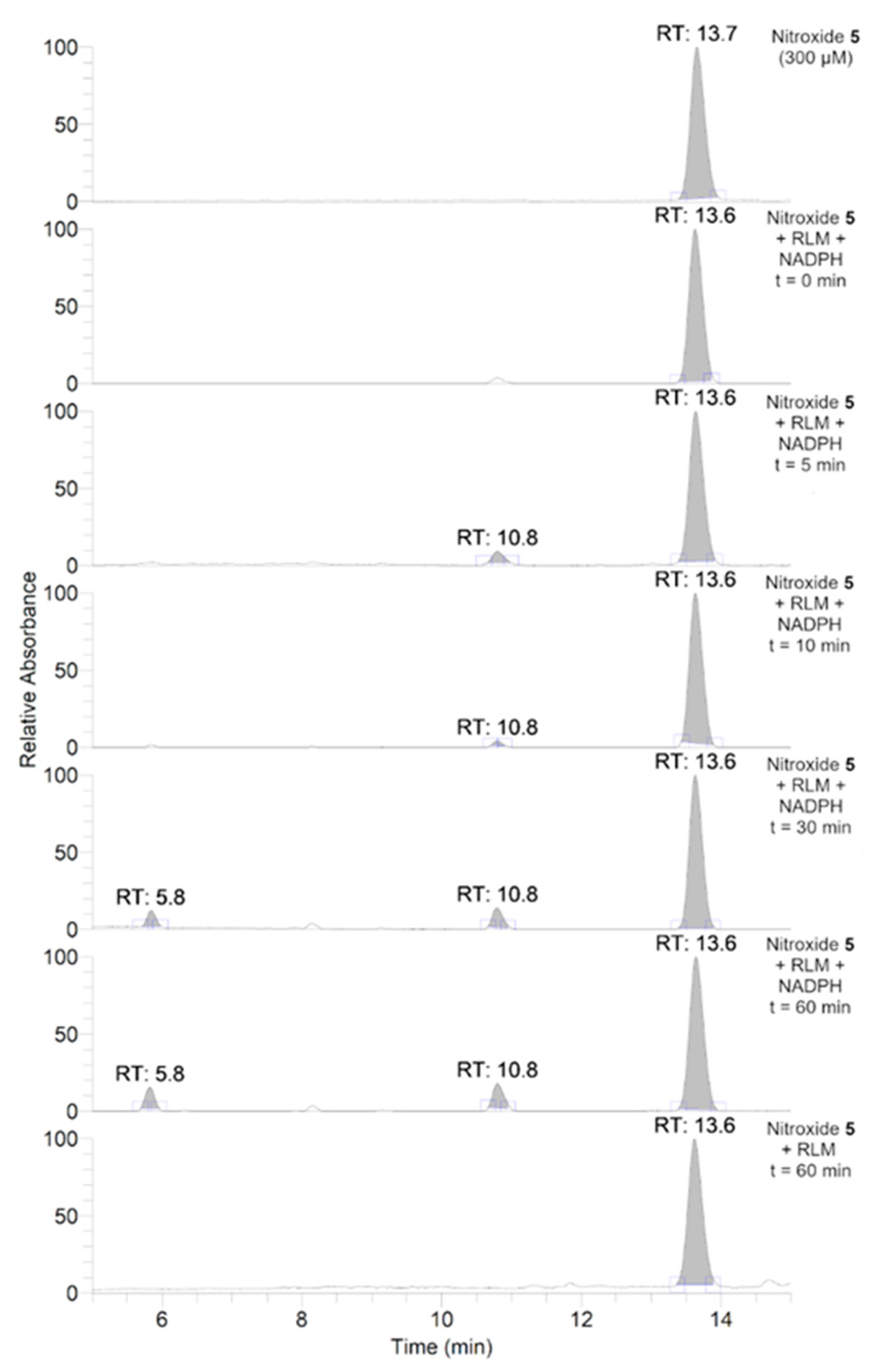
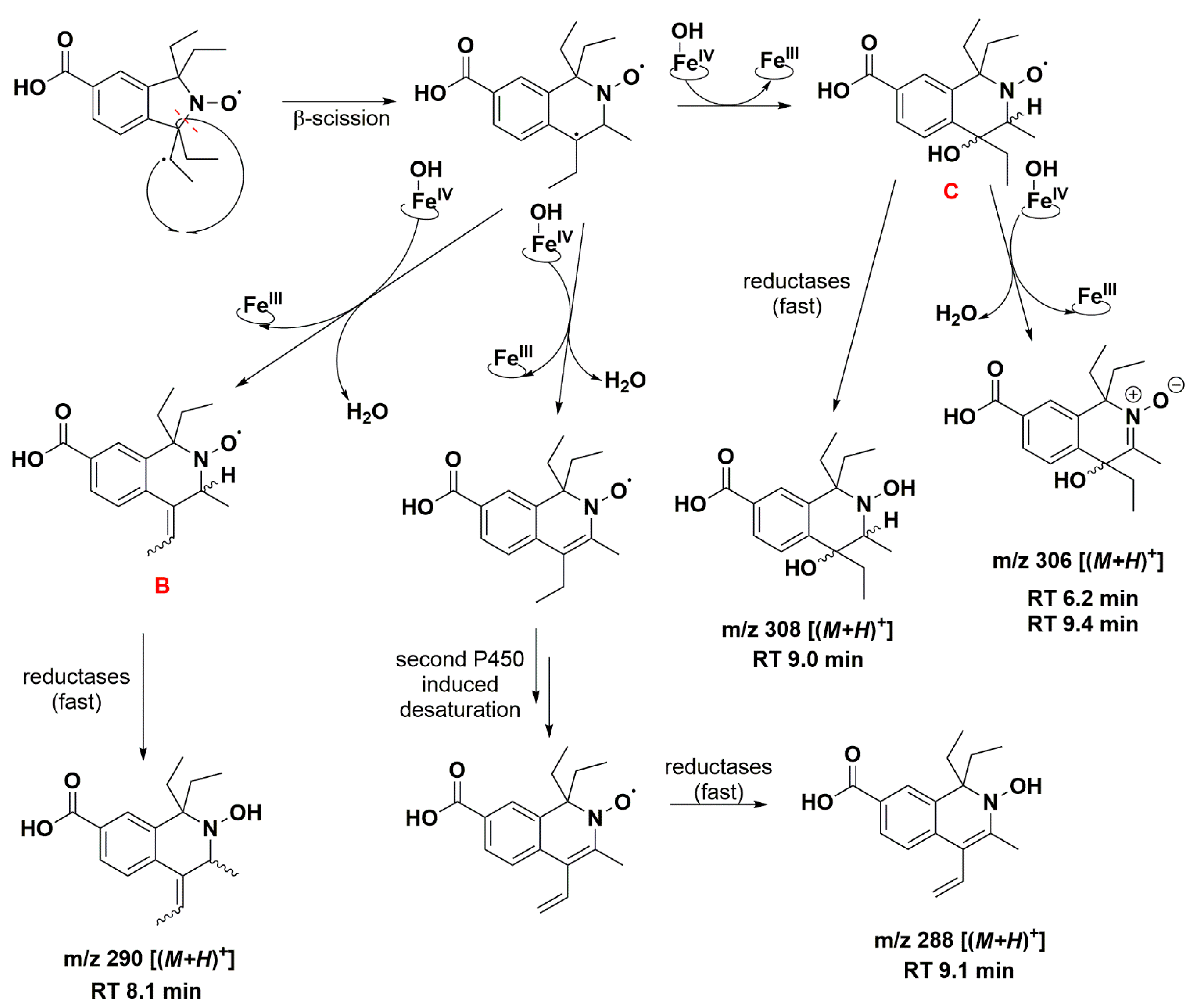
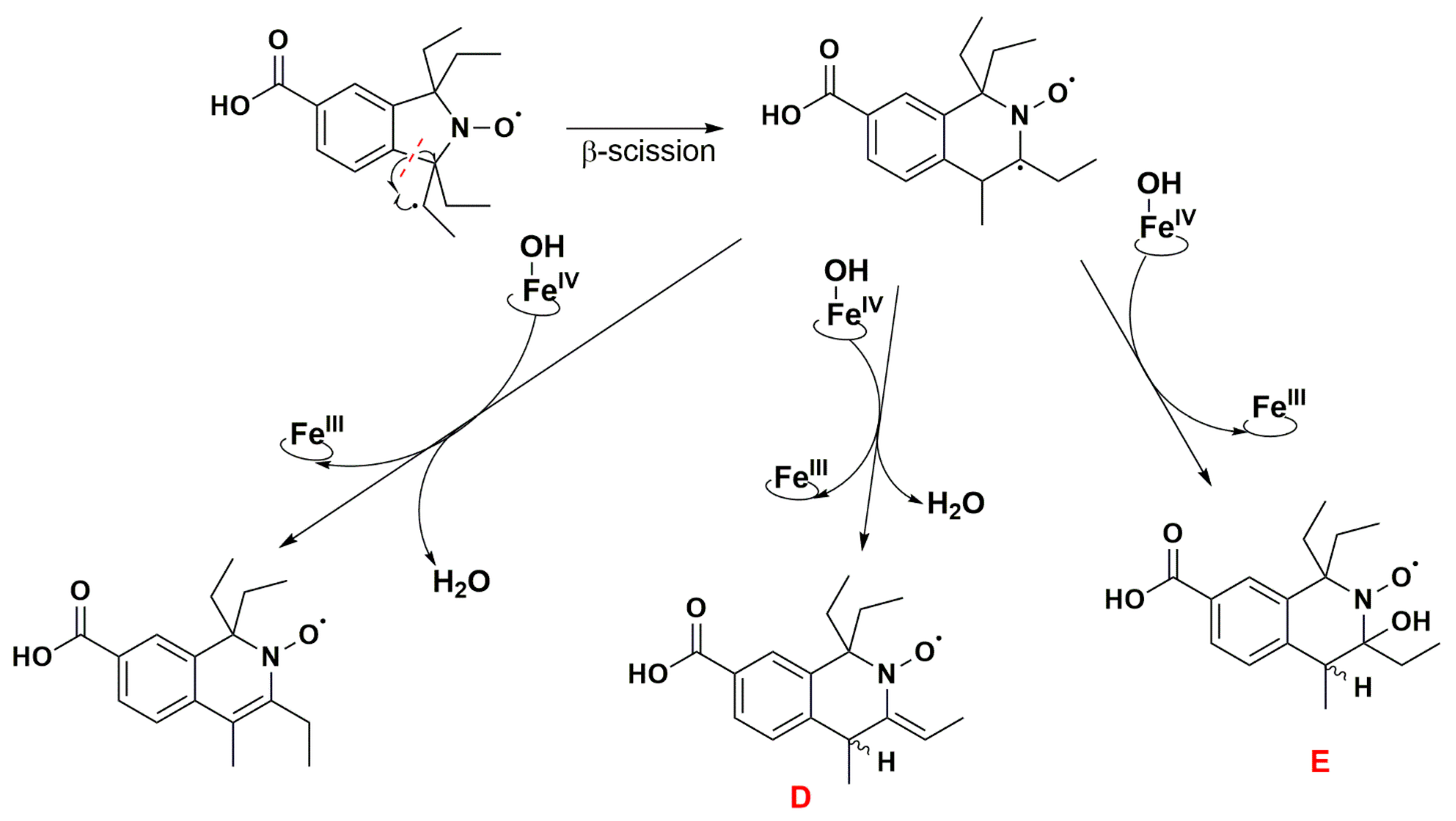
3.3. Aerobic Study of Nitroxides in Incubation with RLM
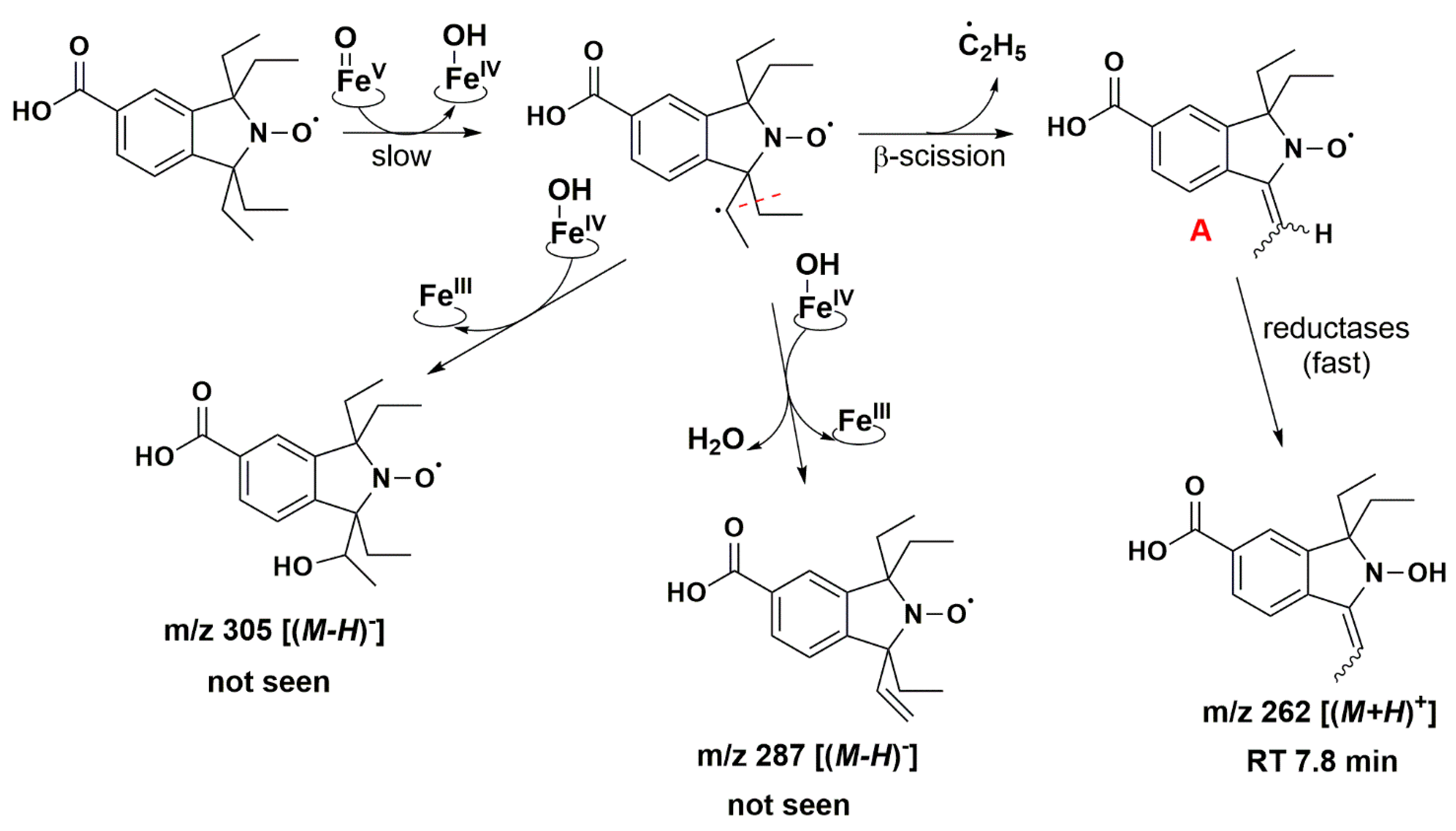
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Samuni, Y.; Gamson, J.; Samuni, A.; Yamada, K.; Russo, A.; Krishna, M.C.; Mitchell, J.B. Factors Influencing Nitroxide Reduction and Cytotoxicity In Vitro. Antioxid. Redox Signal. 2004, 6, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, K.; Chen, P.; Lavin, F.M.; Bottle, E.S. The Impact of Carboxy Nitroxide Antioxidants on Irradiated Ataxia Telangiectasia Cells. Free Radic. Biol. Med. 2004, 37, 946–952. [Google Scholar] [CrossRef]
- Kocherginsky, N.; Swartz, H.M. Nitroxide Spin Labels: Reactions in Biology and Chemistry; CRC Press: Boca Raton, FL, USA, 1995; ISBN 978-0-8493-4204-2. [Google Scholar]
- Zamora, P.L.; Villamena, F.A. Clinical Probes for ROS and Oxidative Stress. In Measuring Oxidants and Oxidative Stress in Biological Systems; Berliner, L.J., Parinandi, N.L., Eds.; Biological Magnetic Resonance; Springer: Cham, Germany, 2020; pp. 13–38. ISBN 978-3-030-47318-1. [Google Scholar]
- Fujii, H.; Emoto, M.; Sato-Akaba, H. Brain Redox Imaging Using In Vivo Electron Paramagnetic Resonance Imaging and Nitroxide Imaging Probes. Magnetochemistry 2019, 5, 11. [Google Scholar] [CrossRef]
- Ilangovan, G.; Li, H.; Zweier, J.L.; Kuppusamy, P. In Vivo Measurement of Tumor Redox Environment Using EPR Spectroscopy. In Oxygen/Nitrogen Radicals: Cell Injury and Disease; Vallyathan, V., Shi, X., Castranova, V., Eds.; Springer: Boston, MA, USA, 2002; pp. 393–398. ISBN 978-1-4613-5388-1. [Google Scholar]
- Elas, M.; Ichikawa, K.; Halpern, H.J. Oxidative Stress Imaging in Live Animals with Techniques Based on Electron Paramagnetic Resonance. Radiat. Res. 2012, 177, 514–523. [Google Scholar] [CrossRef]
- Bačić, G.; Pavićević, A.; Peyrot, F. In Vivo Evaluation of Different Alterations of Redox Status by Studying Pharmacokinetics of Nitroxides Using Magnetic Resonance Techniques. Redox Biol. 2016, 8, 226–242. [Google Scholar] [CrossRef]
- Babić, N.; Peyrot, F. Molecular Probes for Evaluation of Oxidative Stress by In Vivo EPR Spectroscopy and Imaging: State-of-the-Art and Limitations. Magnetochemistry 2019, 5, 13. [Google Scholar] [CrossRef]
- Ahmad, R.; Kuppusamy, P. Theory, Instrumentation, and Applications of Electron Paramagnetic Resonance Oximetry. Chem. Rev. 2010, 110, 3212–3236. [Google Scholar] [CrossRef]
- Kirilyuk, I.A.; Bobko, A.A.; Khramtsov, V.V.; Grigor’ev, I.A. Nitroxides with Two PK Values—Useful Spin Probes for pH Monitoring within a Broad Range. Org. Biomol. Chem. 2005, 3, 1269–1274. [Google Scholar] [CrossRef]
- Kovaleva, E.G.; Molochnikov, L.S.; Golovkina, E.L.; Hartmann, M.; Kirilyuk, I.A.; Grigor’ev, I.A. Dynamics of pH-Sensitive Nitroxide Radicals in Water Adsorbed in Ordered Mesoporous Molecular Sieves by EPR Spectroscopy. Microporous Mesoporous Mater. 2013, 179, 258–264. [Google Scholar] [CrossRef]
- Torricella, F.; Pierro, A.; Mileo, E.; Belle, V.; Bonucci, A. Nitroxide Spin Labels and EPR Spectroscopy: A Powerful Association for Protein Dynamics Studies. Biochim. Biophys. Acta BBA Proteins Proteom. 2021, 1869, 140653. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Nakanishi, I.; Zhelev, Z.; Bakalova, R.; Aoki, I. Nitroxyl Radical as a Theranostic Contrast Agent in Magnetic Resonance Redox Imaging. Antioxid. Redox Signal. 2022, 36, 95–121. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Hyodo, F.; Matsumoto, A.; Koretsky, A.P.; Sowers, A.L.; Mitchell, J.B.; Krishna, M.C. High-Resolution Mapping of Tumor Redox Status by Magnetic Resonance Imaging Using Nitroxides as Redox-Sensitive Contrast Agents. Clin. Cancer Res. 2006, 12, 2455–2462. [Google Scholar] [CrossRef] [PubMed]
- Hyodo, F.; Chuang, K.-H.; Goloshevsky, A.G.; Sulima, A.; Griffiths, G.L.; Mitchell, J.B.; Koretsky, A.P.; Krishna, M.C. Brain Redox Imaging Using Blood—Brain Barrier-Permeable Nitroxide MRI Contrast Agent. J. Cereb. Blood Flow Metab. 2008, 28, 1165–1174. [Google Scholar] [CrossRef]
- Hodgson, J.L.; Namazian, M.; Bottle, S.E.; Coote, M.L. One-Electron Oxidation and Reduction Potentials of Nitroxide Antioxidants: A Theoretical Study. J. Phys. Chem. A 2007, 111, 13595–13605. [Google Scholar] [CrossRef] [PubMed]
- Krishna, M.C.; Russo, A.; Mitchell, J.B.; Goldstein, S.; Dafni, H.; Samuni, A. Do Nitroxide Antioxidants Act as Scavengers of O2.− or as SOD Mimics? J. Biol. Chem. 1996, 271, 26026–26031. [Google Scholar] [CrossRef] [PubMed]
- Krishna, M.C.; Grahame, D.A.; Samuni, A.; Mitchell, J.B.; Russo, A. Oxoammonium Cation Intermediate in the Nitroxide-Catalyzed Dismutation of Superoxide. Proc. Natl. Acad. Sci. USA 1992, 89, 5537–5541. [Google Scholar] [CrossRef]
- Yamada, K.-I.; Kuppusamy, P.; English, S.; Yoo, J.; Irie, A.; Subramanian, S.; Mitchell, J.B.; Krishna, M.C. Feasibility and Assessment of Non-Invasive in Vivo Redox Status Using Electron Paramagnetic Resonance Imaging. Acta Radiol. 2002, 43, 433–440. [Google Scholar] [CrossRef]
- Emoto, M.C.; Sato-Akaba, H.; Hirata, H.; Fujii, H.G. Dynamic Changes in the Distribution and Time Course of Blood–Brain Barrier-Permeative Nitroxides in the Mouse Head with EPR Imaging: Visualization of Blood Flow in a Mouse Model of Ischemia. Free Radic. Biol. Med. 2014, 74, 222–228. [Google Scholar] [CrossRef]
- Bobko, A.A.; Kirilyuk, I.A.; Grigor’ev, I.A.; Zweier, J.L.; Khramtsov, V.V. Reversible Reduction of Nitroxides to Hydroxylamines: Roles for Ascorbate and Glutathione. Free Radic. Biol. Med. 2007, 42, 404–412. [Google Scholar] [CrossRef]
- Emoto, M.; Mito, F.; Yamasaki, T.; Yamada, K.-I.; Sato-Akaba, H.; Hirata, H.; Fujii, H. A Novel Ascorbic Acid-Resistant Nitroxide in Fat Emulsion Is an Efficient Brain Imaging Probe for in Vivo EPR Imaging of Mouse. Free Radic. Res. 2011, 45, 1325–1332. [Google Scholar] [CrossRef]
- Swartz, H.M.; Sentjurc, M.; Morse, P.D. Cellular Metabolism of Water-Soluble Nitroxides: Effect on Rate of Reduction of Cell/Nitroxide Ratio, Oxygen Concentrations and Permeability of Nitroxides. Biochim. Biophys. Acta BB Mol. Cell Res. 1986, 888, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Swartz, H.M.; Chen, K.; Pals, M.; Sentjurc, M.; Morse, P.D. Hypoxia-Sensitive NMR Contrast Agents. Magn. Reson. Med. 1986, 3, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Azuma, R.; Yamasaki, T.; Emoto, M.C.; Sato-Akaba, H.; Sano, K.; Munekane, M.; Fujii, H.G.; Mukai, T. Effect of Relative Configuration of TEMPO-Type Nitroxides on Ascorbate Reduction. Free Radic. Biol. Med. 2023, 194, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, G.; Bonucci, A.; Casano, G.; Gerbaud, G.; Abel, S.; Thomé, V.; Kodjabachian, L.; Magalon, A.; Guigliarelli, B.; Belle, V.; et al. A Bioresistant Nitroxide Spin Label for In-Cell EPR Spectroscopy: In Vitro and In Oocytes Protein Structural Dynamics Studies. Angew. Chem. Int. Ed. 2018, 57, 1366–1370. [Google Scholar] [CrossRef]
- Braun, T.S.; Widder, P.; Osswald, U.; Groß, L.; Williams, L.; Schmidt, M.; Helmle, I.; Summerer, D.; Drescher, M. Isoindoline-Based Nitroxides as Bioresistant Spin Labels for Protein Labeling through Cysteines and Alkyne-Bearing Noncanonical Amino Acids. ChemBioChem 2020, 21, 958–962. [Google Scholar] [CrossRef] [PubMed]
- Dobrynin, S.A.; Usatov, M.S.; Zhurko, I.F.; Morozov, D.A.; Polienko, Y.F.; Glazachev, Y.I.; Parkhomenko, D.A.; Tyumentsev, M.A.; Gatilov, Y.V.; Chernyak, E.I.; et al. A Simple Method of Synthesis of 3-Carboxy-2,2,5,5-Tetraethylpyrrolidine-1-Oxyl and Preparation of Reduction-Resistant Spin Labels and Probes of Pyrrolidine Series. Molecules 2021, 26, 5761. [Google Scholar] [CrossRef]
- Paletta, J.T.; Pink, M.; Foley, B.; Rajca, S.; Rajca, A. Synthesis and Reduction Kinetics of Sterically Shielded Pyrrolidine Nitroxides. Org. Lett. 2012, 14, 5322–5325. [Google Scholar] [CrossRef]
- Kinoshita, Y.; Yamada, K.-I.; Yamasaki, T.; Sadasue, H.; Sakai, K.; Utsumi, H. Development of Novel Nitroxyl Radicals for Controlling Reactivity with Ascorbic Acid. Free Radic. Res. 2009, 43, 565–571. [Google Scholar] [CrossRef]
- Kirilyuk, I.A.; Bobko, A.A.; Grigor’ev, I.A.; Khramtsov, V.V. Synthesis of the Tetraethyl Substituted pH-Sensitive Nitroxides of Imidazole Series with Enhanced Stability towards Reduction. Org. Biomol. Chem. 2004, 2, 1025. [Google Scholar] [CrossRef]
- Baranczewski, P.; Stańczak, A.; Sundberg, K.; Svensson, R.; Wallin, A.; Jansson, J.; Garberg, P.; Postlind, H. Introduction to in Vitro Estimation of Metabolic Stability and Drug Interactions of New Chemical Entities in Drug Discovery and Development. Pharmacol. Rep. PR 2006, 58, 453–472. [Google Scholar]
- Testa, B. Metabolism in Drug Development. In Drug Metabolism Prediction; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 1–26. ISBN 978-3-527-67326-1. [Google Scholar]
- Shen, J.; Bottle, S.; Khan, N.; Grinberg, O.; Reid, D.; Micallef, A.; Swartz, H. Development of Isoindoline Nitroxides for EPR Oximetry in Viable Systems. Appl. Magn. Reson. 2002, 22, 357–368. [Google Scholar] [CrossRef]
- Quintanilha, A.T.; Packer, L. Surface Localization of Sites of Reduction of Nitroxide Spin-Labeled Molecules in Mitochondria. Proc. Natl. Acad. Sci. USA 1977, 74, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Glockner, J.F.; Morse, P.D.I.; Swartz, H.M. Effects of Oxygen on the Metabolism of Nitroxide Spin Labels in Cells. Biochemistry 1989, 28, 2496–2501. [Google Scholar] [CrossRef] [PubMed]
- Ueda, A.; Nagase, S.; Yokoyama, H.; Tada, M.; Noda, H.; Ohya, H.; Kamada, H.; Hirayama, A.; Koyama, A. Importance of Renal Mitochondria in the Reduction of TEMPOL, a Nitroxide Radical. Mol. Cell. Biochem. 2003, 244, 119–124. [Google Scholar] [CrossRef]
- Keana, J.F.; Pou, S.; Rosen, G.M. Nitroxides as Potential Contrast Enhancing Agents for MRI Application: Influence of Structure on the Rate of Reduction by Rat Hepatocytes, Whole Liver Homogenate, Subcellular Fractions, and Ascorbate. Magn. Reson. Med. 1987, 5, 525–536. [Google Scholar] [CrossRef]
- Iannone, A.; Tomasi, A.; Vannini, V.; Swartz, H.M. Metabolism of Nitroxide Spin Labels in Subcellular Fraction of Rat Liver. Biochim. Biophys. Acta BBA Gen. Subj. 1990, 1034, 285–289. [Google Scholar] [CrossRef]
- Babić, N.; Orio, M.; Peyrot, F. Unexpected Rapid Aerobic Transformation of 2,2,6,6-Tetraethyl-4-Oxo(Piperidin-1-Yloxyl) Radical by Cytochrome P450 in the Presence of NADPH: Evidence against a Simple Reduction of the Nitroxide Moiety to the Hydroxylamine. Free Radic. Biol. Med. 2020, 156, 144–156. [Google Scholar] [CrossRef]
- Jagtap, A.P.; Krstic, I.; Kunjir, N.C.; Hänsel, R.; Prisner, T.F.; Sigurdsson, S.T. Sterically Shielded Spin Labels for In-Cell EPR Spectroscopy: Analysis of Stability in Reducing Environment. Free Radic. Res. 2015, 49, 78–85. [Google Scholar] [CrossRef]
- Kajer, T.B.; Fairfull-Smith, K.E.; Yamasaki, T.; Yamada, K.; Fu, S.; Bottle, S.E.; Hawkins, C.L.; Davies, M.J. Inhibition of Myeloperoxidase- and Neutrophil-Mediated Oxidant Production by Tetraethyl and Tetramethyl Nitroxides. Free Radic. Biol. Med. 2014, 70, 96–105. [Google Scholar] [CrossRef]
- Kinoshita, Y.; Yamada, K.; Yamasaki, T.; Mito, F.; Yamato, M.; Kosem, N.; Deguchi, H.; Shirahama, C.; Ito, Y.; Kitagawa, K.; et al. In Vivo Evaluation of Novel Nitroxyl Radicals with Reduction Stability. Free Radic. Biol. Med. 2010, 49, 1703–1709. [Google Scholar] [CrossRef]
- Griffiths, P.G.; Rizzardo, E.; Solomon, D.H. Quantitative Studies on Free Radical Reactions with the Scavenger 1,1,3,3-Tetramethylisoindolinyl-2-Oxy. Tetrahedron Lett. 1982, 23, 1309–1312. [Google Scholar] [CrossRef]
- Khan, N.; Blinco, J.P.; Bottle, S.E.; Hosokawa, K.; Swartz, H.M.; Micallef, A.S. The Evaluation of New and Isotopically Labeled Isoindoline Nitroxides and an Azaphenalene Nitroxide for EPR Oximetry. J. Magn. Reson. 2011, 211, 170–177. [Google Scholar] [CrossRef]
- Kuppusamy, P.; Chzhan, M.; Zweier, J.L. Principles of Imaging: Theory and Instrumentation. In In Vivo EPR (ESR); Berliner, L.J., Ed.; Biological Magnetic Resonance; Springer: Boston, MA, USA, 2003; Volume 18, pp. 99–152. ISBN 978-1-4613-4906-8. [Google Scholar]
- Wang, Y.; Paletta, J.T.; Berg, K.; Reinhart, E.; Rajca, S.; Rajca, A. Synthesis of Unnatural Amino Acids Functionalized with Sterically Shielded Pyrroline Nitroxides. Org. Lett. 2014, 16, 5298–5300. [Google Scholar] [CrossRef]
- Hatano, B.; Araya, H.; Yoshimura, Y.; Sato, H.; Ito, T.; Ogata, T.; Kijima, T. Facile Synthesis of 3-Methoxycarbonyl-2,2,5,5-Tetra-Methylpyrrolidine-1-Oxyl and Derivatives. Heterocycles 2010, 81, 349. [Google Scholar] [CrossRef]
- Fairfull-Smith, K.E.; Brackmann, F.; Bottle, S.E. The Synthesis of Novel Isoindoline Nitroxides Bearing Water-Solubilising Functionality. Eur. J. Org. Chem. 2009, 2009, 1902–1915. [Google Scholar] [CrossRef]
- Smith, C.D.; Bartley, J.P.; Bottle, S.E.; Micallef, A.S.; Reid, D.A. Electrospray Ionization Mass Spectrometry of Stable Nitroxide Free Radicals and Two Isoindoline Nitroxide Dimers. J. Mass Spectrom. 2000, 35, 607–611. [Google Scholar] [CrossRef]
- Scherrer, R.A.; Howard, S.M. Use of Distribution Coefficients in Quantitative Structure-Activity Relations. J. Med. Chem. 1977, 20, 53–58. [Google Scholar] [CrossRef]
- Kremers, P.; Beaune, P.; Cresteil, T.; Graeve, J.; Columelli, S.; Leroux, J.-P.; Gielen, J.E. Cytochrome P-450 Monooxygenase Activities in Human and Rat Liver Microsomes. Eur. J. Biochem. 1981, 118, 599–606. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Vermilion, J.L.; Coon, M.J. Purified Liver Microsomal NADPH-Cytochrome P-450 Reductase. Spectral Characterization of Oxidation-Reduction States. J. Biol. Chem. 1978, 253, 2694–2704. [Google Scholar] [CrossRef]
- Omura, T.; Sato, R. The Carbon Monoxide-Binding Pigment of Liver Microsomes. J. Biol. Chem. 1964, 239, 2370–2378. [Google Scholar] [CrossRef] [PubMed]
- Bézière, N.; Hardy, M.; Poulhès, F.; Karoui, H.; Tordo, P.; Ouari, O.; Frapart, Y.-M.; Rockenbauer, A.; Boucher, J.-L.; Mansuy, D.; et al. Metabolic Stability of Superoxide Adducts Derived from Newly Developed Cyclic Nitrone Spin Traps. Free Radic. Biol. Med. 2014, 67, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Babić, N.; Peyrot, F. New Synthetic Route to 2,2,6,6-Tetraethylpiperidin-4-One: A Key-Intermediate towards Tetraethyl Nitroxides. Tetrahedron Lett. 2019, 60, 151207. [Google Scholar] [CrossRef]
- Stoll, S.; Schweiger, A. EasySpin, a Comprehensive Software Package for Spectral Simulation and Analysis in EPR. J. Magn. Reson. 2006, 178, 42–55. [Google Scholar] [CrossRef]
- Neese, F. The ORCA Program System. WIREs Comput. Mol. Sci. 2012, 2, 73–78. [Google Scholar] [CrossRef]
- Becke, A.D. A New Mixing of Hartree–Fock and Local Density-functional Theories. J. Chem. Phys. 1993, 98, 1372–1377. [Google Scholar] [CrossRef]
- Lee, C.; Yang, W.; Parr, R.G. Development of the Colle-Salvetti Correlation-Energy Formula into a Functional of the Electron Density. Phys. Rev. B 1988, 37, 785–789. [Google Scholar] [CrossRef]
- Csonka, G.I. Proper Basis Set for Quantum Mechanical Studies of Potential Energy Surfaces of Carbohydrates. J. Mol. Struct. 2002, 584, 1–4. [Google Scholar] [CrossRef]
- Boese, A.D.; Martin, J.M.L.; Handy, N.C. The Role of the Basis Set: Assessing Density Functional Theory. J. Chem. Phys. 2003, 119, 3005–3014. [Google Scholar] [CrossRef]
- Mackie, I.D.; DiLabio, G.A. Accurate Dispersion Interactions from Standard Density-Functional Theory Methods with Small Basis Sets. Phys. Chem. Chem. Phys. 2010, 12, 6092–6098. [Google Scholar] [CrossRef]
- Ho, J.; Ertem, M.Z. Calculating Free Energy Changes in Continuum Solvation Models. J. Phys. Chem. B 2016, 120, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Franchi, P.; Lucarini, M.; Pedrielli, P.; Pedulli, G.F. Nitroxide Radicals as Hydrogen Bonding Acceptors. An Infrared and EPR Study. ChemPhysChem 2002, 3, 789–793. [Google Scholar] [CrossRef] [PubMed]
- Owenius, R.; Engström, M.; Lindgren, M.; Huber, M. Influence of Solvent Polarity and Hydrogen Bonding on the EPR Parameters of a Nitroxide Spin Label Studied by 9-GHz and 95-GHz EPR Spectroscopy and DFT Calculations. J. Phys. Chem. A 2001, 105, 10967–10977. [Google Scholar] [CrossRef]
- Hermosilla, L.; Vega, J.G.D.L.; Sieiro, C.; Calle, P. DFT Calculations of Isotropic Hyperfine Coupling Constants of Nitrogen Aromatic Radicals: The Challenge of Nitroxide Radicals. J. Chem. Theory Comput. 2011, 7, 169–179. [Google Scholar] [CrossRef]
- Dikalov, S.; Skatchkov, M.; Bassenge, E. Spin Trapping of Superoxide Radicals and Peroxynitrite by 1-Hydroxy-3-Carboxy-Pyrrolidine and 1-Hydroxy-2,2,6,6-Tetramethyl-4-Oxo-Piperidine and the Stability of Corresponding Nitroxyl Radicals Towards Biological Reductants. Biochem. Biophys. Res. Commun. 1997, 231, 701–704. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, H.; Paletta, J.T.; Rajca, S.; Rajca, A. Reduction Kinetics and Electrochemistry of Tetracarboxylate Nitroxides. Free Radic. Res. 2018, 52, 327–334. [Google Scholar] [CrossRef]
- Tew, D.G. Inhibition of Cytochrome P450 Reductase by the Diphenyliodonium Cation. Kinetic Analysis and Covalent Modifications. Biochemistry 1993, 32, 10209–10215. [Google Scholar] [CrossRef]
- Crowley, P.D.; Gallagher, H.C. Clotrimazole as a Pharmaceutical: Past, Present and Future. J. Appl. Microbiol. 2014, 117, 611–617. [Google Scholar] [CrossRef]
- Lewis, D.F.V.; Pratt, J.M. The P450 Catalytic Cycle and Oxygenation Mechanism. Drug Metab. Rev. 1998, 30, 739–786. [Google Scholar] [CrossRef]
- Amunom, I.; Srivastava, S.; Prough, R.A. Aldehyde Reduction by Cytochrome P450. Curr. Protoc. Toxicol. 2011, 48, 4.37.1–4.37.15. [Google Scholar] [CrossRef]
- Guengerich, F.P. Oxidation-Reduction Properties of Rat Liver Cytochromes P-450 and NADPH-Cytochrome P-450 Reductase Related to Catalysis in Reconstituted Systems. Biochemistry 1983, 22, 2811–2820. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Johnson, W.W.; Ueng, Y.-F.; Shimada, T.; Guengerich, F.P. Lack of Electron Transfer from Cytochrome B5 in Stimulation of Catalytic Activities of Cytochrome P450 3A4: Characterization of a Reconstituted Cytochrome P450 3A4/NADPH-Cytochrome P450 Reductase System and Studies With APO-Cytochrome B5*. J. Biol. Chem. 1996, 271, 27438–27444. [Google Scholar] [CrossRef] [PubMed]
- Correia, M.A.; Ortiz de Montellano, P.R. Inhibition of Cytochrome P450 Enzymes. In Cytochrome P450: Structure, Mechanism, and Biochemistry; Ortiz de Montellano, P.R., Ed.; Springer: Boston, MA, USA, 2005; pp. 247–322. ISBN 978-0-387-27447-8. [Google Scholar]
- Guengerich, F.P. Common and Uncommon Cytochrome P450 Reactions Related to Metabolism and Chemical Toxicity. Chem. Res. Toxicol. 2001, 14, 611–650. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.F.V.; Jacobs, M.N.; Dickins, M. Compound Lipophilicity for Substrate Binding to Human P450s in Drug Metabolism. Drug Discov. Today 2004, 9, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Mehlhorn, R.J.; Swanson, C.E. Nitroxide-Stimulated H2O2 Decomposition by Peroxidases and Pseudoperoxidases. Free Radic. Res. Commun. 1992, 17, 157–175. [Google Scholar] [CrossRef]
- Krishna, M.C.; Samuni, A.; Taira, J.; Goldstein, S.; Mitchell, J.B.; Russo, A. Stimulation by Nitroxides of Catalase-like Activity of Hemeproteins: Kinetics and Mechanism. J. Biol. Chem. 1996, 271, 26018–26025. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Nakano, T.; Kimoto, E. Oxidation of Nitroxide Radicals by the Reaction of Hemoglobin with Hydrogen Peroxide. Biochem. Biophys. Res. Commun. 1984, 120, 534–539. [Google Scholar] [CrossRef]
- Bono, D.D.; Yang, W.-D.; Symons, M.C.R. The Effect of Myoglobin on the Stability of the Hydroxyl-Radical Adducts of 5, 5 Dimethyl-1-Pyrolline-N-Oxide (DMPO), 3, 3,5, 5 Tetramethyl-1-Pyrolline-N-Oxide(TMPO) and 1-Alpha-Phenyl-Tert-Butyl Nitrone (PBN) in the Presence of Hydrogen Peroxide. Free Radic. Res. 1994, 20, 327–332. [Google Scholar] [CrossRef]
- Kroll, C.; Borchert, H.-H. Metabolism of the Stable Nitroxyl Radical 4-Oxo-2,2,6,6-Tetramethylpiperidine-N-Oxyl (TEMPONE). Eur. J. Pharm. Sci. 1999, 8, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Isin, E.M.; Guengerich, F.P. Complex Reactions Catalyzed by Cytochrome P450 Enzymes. Biochim. Biophys. Acta BBA Gen. Subj. 2007, 1770, 314–329. [Google Scholar] [CrossRef]
- Mittra, K.; Green, M.T. Reduction Potentials of P450 Compounds I and II: Insight into the Thermodynamics of C–H Bond Activation. J. Am. Chem. Soc. 2019, 141, 5504–5510. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Mechanisms of Cytochrome P450-Catalyzed Oxidations. ACS Catal. 2018, 8, 10964–10976. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, Y.; Bossant, M.-J.; Bertrand, M.; Walther, B. 5.10—In Vitro Studies of Drug Metabolism. In Comprehensive Medicinal Chemistry II; Taylor, J.B., Triggle, D.J., Eds.; Elsevier: Oxford, UK, 2007; pp. 231–257. ISBN 978-0-08-045044-5. [Google Scholar]
- Iannone, A.; Bini, A.; Swartz, H.M.; Tomasi, A.; Vannini, V. Metabolism in Rat Liver Microsomes of the Nitroxide Spin Probe Tempol. Biochem. Pharmacol. 1989, 38, 2581–2586. [Google Scholar] [CrossRef]
- Zhitkovich, A. Nuclear and Cytoplasmic Functions of Vitamin C. Chem. Res. Toxicol. 2020, 33, 2515–2526. [Google Scholar] [CrossRef] [PubMed]
- Hoch, M.J.R.; Ewert, U. Resolution in EPR Imaging*. In EPR Imaging and In Vivo EPR; CRC Press: Boca Raton, FL, USA, 1983; ISBN 978-1-351-07188-8. [Google Scholar]
- Valvis, I.I.; Lischick, D.; Shen, D.; Sofer, S.S. In Vitro Synthesis of Nitroxide Free Radicals by Hog Liver Microsomes. Free Radic. Biol. Med. 1990, 9, 345–353. [Google Scholar] [CrossRef]
- Olsen, L.; Rydberg, P.; Rod, T.H.; Ryde, U. Prediction of Activation Energies for Hydrogen Abstraction by Cytochrome P450. J. Med. Chem. 2006, 49, 6489–6499. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nitroxide | k0 (M−1·s−1) | Reference |
|---|---|---|
| 1 | 6.9 ± 0.1 | This work |
| 6.9 ± 0.7 | [57] | |
| 5.75 ± 0.04 | [42] | |
| 7.2 ± 0.8 a | [70] | |
| 2 | 0.072 ± 0.003 | This work |
| 0.11 ± 0.01 | [70] | |
| 0.079 ± 0.001 | [42] | |
| 0.060 ± 0.003 a | [71] | |
| 3 | 0.0011 ± 0.0002 | This work |
| ≤0.001 | [30] | |
| 0.002 ± 0.001 | [42] | |
| 0.0011 ± 0.0002 a | [71] | |
| 4 | 0.0077 ± 0.0009 | This work |
| 0.00122 ± 0.00003 a | [71] | |
| 5 | 0.02 ± 0.01 | This work |
| Nitroxide | Anaerobic | ||||
|---|---|---|---|---|---|
| WS | -RLM | -NADPH | +DPI | + Clo | |
| TEMPONE 1 | 19.3 ± 0.9 (5.0 ± 0.5) | 0.0 ** | 0.0 ** | 2.7 ± 0.3 ** (0.70 ± 0.08) | 4.3 ± 0.4 (1.1 ± 0.2) |
| 2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| 3 | 2.2 ± 0.6 (0.56 ± 0.08) | 0.0 | 0.0 | 0.0 | 0.0 |
| 4 | 1.8 ± 0.2 (0.47 ± 0.05) | 0.0 * | 0.0 * | 0.0 * | 0.0 * |
| 5 | 5.2 ± 0.5 (1.35 ± 0.07) | 0.0 *** | 0.0 *** | 0.0 *** | 0.0 *** |
| Nitroxide | Aerobic | ||||
|---|---|---|---|---|---|
| WS | -RLM | -NADPH | +DPI | + Clo | |
| TEMPONE 1 | 11.1 ± 0.8 * (2.8 ± 0.2) | 0.0 *** | 0.0 *** | 5.1 ± 0.3 ** (1.31 ± 0.07) | 1.2 ± 0.3 *** (0.30 ± 0.09) |
| 2 | 4.8 ± 0.8 ** (1.2 ± 0.1) | 0.0 ** | 0.0 ** | 2.1 ± 0.3 * (0.54 ± 0.07) | 0.94 ± 0.09 * (0.24 ± 0.03) |
| 3 | 1.8 ± 0.2 (0.46 ± 0.05) | 0.0 ** | 0.0 ** | 0.0 ** | 0.0 ** |
| 4 | 0.84 ± 0.09 ** (0.22 ± 0.03) | 0.0 ** | 0.0 ** | 0.0 ** | 0.0 ** |
| 5 | 3.4 ± 0.2 * (0.88 ± 0.05) | 0.0 *** | 0.0 *** | 0.0 *** | 0.0 *** |
| RT [min] | Positive ESI Mode | Suggested Modification | ||
|---|---|---|---|---|
| m/z [Da] | Chemical Formula | |||
| Observed | Calculated | |||
| 6.2 | 306.1689 | 306.1700 | C17H24NO4, [(M+H)+] | +O, −H |
| 7.8 | 262.1438 | 262.1438 | C15H20NO3, [(M+H)+] | −C2H6, +H |
| 8.1 | 290.1734 | 290.1751 | C17H24NO3, [(M+H)+] | −H2, +H |
| 9.0 | 308.1842 | 308.1856 | C17H26NO4, [(M+H)+] | +O, +H |
| 9.1 | 288.1579 | 288.1594 | C17H22NO3, [(M+H)+] | −2H2, +H |
| 9.4 | 306.1689 | 306.1700 | C17H24NO4, [(M+H)+] | +O, −H |
| Compound | AN (MHz/mT) | AH (MHz/mT) |
|---|---|---|
| 5 (carboxylic acid form) | 37.2/1.33 | - |
| 5 (carboxylic acid form) + H2O | 41.6/1.49 | - |
| 5 (carboxylate form) + H2O | 42.1/1.50 | - |
| 5 (Expt.) | 43.4/1.55 | - |
| C trans (carboxylic acid form) | 41.3/1.47 | 70.3/2.51 |
| C trans (carboxylic acid form) + H2O | 42.5/1.52 | 73.4/2.62 |
| C trans (carboxylate form) + H2O | 42.9/1.53 | 73.9/2.64 |
| 6-line intermediate (Expt.) | 44.3/1.58 | 66.1/2.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rančić, A.; Babić, N.; Orio, M.; Peyrot, F. Structural Features Governing the Metabolic Stability of Tetraethyl-Substituted Nitroxides in Rat Liver Microsomes. Antioxidants 2023, 12, 402. https://doi.org/10.3390/antiox12020402
Rančić A, Babić N, Orio M, Peyrot F. Structural Features Governing the Metabolic Stability of Tetraethyl-Substituted Nitroxides in Rat Liver Microsomes. Antioxidants. 2023; 12(2):402. https://doi.org/10.3390/antiox12020402
Chicago/Turabian StyleRančić, Aleksandra, Nikola Babić, Maylis Orio, and Fabienne Peyrot. 2023. "Structural Features Governing the Metabolic Stability of Tetraethyl-Substituted Nitroxides in Rat Liver Microsomes" Antioxidants 12, no. 2: 402. https://doi.org/10.3390/antiox12020402