
Biofloc Technology in Fish Aquaculture: A Review

Abstract
:1. The Necessity of Biofloc Technology in Fish Aquaculture
2. Survival Rate and Growth Performance of Fish Raised in Biofloc

{kind=link}
| Species | Carbon Source | C:N Ratio | Period | Change of Survival Rate | Reference | |
|---|---|---|---|---|---|---|
| Freshwater | Oreochromis niloticus | Molasses | 15:1 | 30 days | + | [16] |
| Starch | + | |||||
| Barley flour | + | |||||
| Corn | + | |||||
| Molasses | 8.4:1 | 12 weeks | × | [22] | ||
| Rice bran | 15:1 | 10 weeks | × | [28] | ||
| Wheat-milling by-product | × | |||||
| Sucrose | >10:1 | 87 days | × | [25] | ||
| Glucose | 15:1 | 8 weeks | × | [29] | ||
| Wheat flour (200 fish/m3) | 15:1 | 90 days | × | [30] | ||
| Wheat flour (250 fish/m3) | × | |||||
| Wheat flour (300 fish/m3) | × | |||||
| Wheat flour (350 fish/m3) | × | |||||
| Molasses | 10:1 | 14 days | + | [10] | ||
| 100% molasses | 15:1, 20:1 | 8 weeks | × | [26] | ||
| 100% wheat flour | × | |||||
| 75% molasses + 25% wheat flour | × | |||||
| 50% molasses + 50% wheat flour | × | |||||
| 25% molasses + 75% wheat flour | × | |||||
| Molasses | 15:1 | 37 days | + | [17] | ||
| Starch | + | |||||
| Barley flour | + | |||||
| Corn | + | |||||
| Molasses (40 fish/m3) | 15:1 | 112 days | × | [31] | ||
| Molasses (80 fish/m3) | × | |||||
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | × | [32] | |
| Sugar (12 kg/m3) | × | |||||
| Rice bran | 20:1 | 60 days | × | [33] | ||
| Sugarcane molasses | - | |||||
| Rice bran + sugarcane molasses | × | |||||
| Corn starch | 15:1 | 60 days | × | [34] | ||
| Cyprinus carpio L. | Sugar beet molasses | 20:1 | 70 days | × | [35] | |
| Sugar | × | |||||
| Corn starch | × | |||||
| Molasses | 20:1 | 30 days | × | [36] | ||
| Clarias gariepinus | Tapioca (4 fish/L) | 10:1 | 20 days | + | [19] | |
| Tapioca (6 fish/L) | - | |||||
| Tapioca (8 fish/L) | - | |||||
| Glycerol | 15:1 | 8 weeks | × | [37] | ||
| Sucrose | 15:1 | 6 weeks | + | [20] | ||
| Glycerol | + | |||||
| Rice bran | - | |||||
| Carassius auratus | Starch | 15:1, 20:1 | 56 days | × | [27] | |
| Mugil cephalus | Sucrose | 15:1 | 60 days | + | [21] | |
| Heteropneustes fossilis | Sugarcane molasses | 10:1 | 120 days | × | [38] | |
Lemon fin barb hybrid (Hypsibarbus wetmorei  × Barboides gonionotus × Barboides gonionotus  ) ) | Glycerol | 15:1 | 8 weeks | × | [37] | |
| Lepomis macrochirus | Corn starch | 15:1 | 32 days | - | [39] | |
| Sucrose-sugar | - | |||||
| Labeo rohita | Wheat flour | 10:1 | 90 days | × | [40] | |
| Seawater | Oreochromis niloticus sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | × | [13] |
| Cornmeal + molasses (240 fish/m3) | × | |||||
| Species | Carbon Source | C:N Ratio | Period | Response C:N Ratio | Response | Reference | |
|---|---|---|---|---|---|---|---|
| Growth performance | |||||||
| Freshwater | Oreochromis niloticus | Molasses | 8.4:1 | 12 weeks | 8.4:1 | + | [23] |
| Molasses (salinity level 4 g/L) | 6:1 | 70 days | 6:1 | + | [23] | ||
| Molasses (salinity level 8 g/L) | 6:1 | + | |||||
| Molasses (salinity level 12 g/L) | 6:1 | - | |||||
| Molasses (salinity level 16 g/L) | 6:1 | - | |||||
| Glycerol | 15:1 | 12 weeks | 15:1 | + | [24] | ||
| Mannan oligosaccharides | 15:1 | + | |||||
| Glucose (166 organisms/m3) | 15:1 | 120 days | 15:1 | + | [41] | ||
| Glucose (333 organisms/m3) | 15:1 | + | |||||
| Glucose (600 organisms/m3) | 15:1 | + | |||||
| Glucose | 10:1, 15:1, 20:1 | 120 days | 10:1, 15:1 | + | [42] | ||
| Rice bran and molasses (1:1) (60 fish/m3) | 15:1 | 20 weeks | 15:1 | + | [43] | ||
| Rice bran and molasses (1:1) (80 fish/m3) | 15:1 | + | |||||
| Molasses | 15:1 | 30 days | 15:1 | + | [16] | ||
| Starch | 15:1 | + | |||||
| Barley flour | 15:1 | + | |||||
| Corn | 15:1 | + | |||||
| Rice bran | 15:1 | 10 weeks | 15:1 | + | [28] | ||
| Wheat-milling by-product | 15:1 | + | |||||
| Wheat flour (200 fish/m3) | 15:1 | 90 days | 15:1 | + | [30] | ||
| Wheat flour (250 fish/m3) | 15:1 | + | |||||
| Wheat flour (300 fish/m3) | 15:1 | + | |||||
| Wheat flour (350 fish/m3) | 15:1 | + | |||||
| Sucrose | >10:1 | 87 days | > 10:1 | + | [25] | ||
| Glucose | 15:1 | 8 weeks | 15:1 | + | [29] | ||
| Molasses | 10:1 | 14 days | - | × | [18] | ||
| 100% molasses | 15:1, 20:1 | 8 weeks | 15:1, 20:1 | + | [26] | ||
| 100% wheat flour | 15:1, 20:1 | + | |||||
| 75% molasses + 25% wheat flour | 15:1, 20:1 | + | |||||
| 50% molasses + 50% wheat flour | 15:1, 20:1 | + | |||||
| 25% molasses + 75% wheat flour | 15:1, 20:1 | + | |||||
| Molasses | 15:1 | 37 days | 15:1 | + | [17] | ||
| Starch | 15:1 | + | |||||
| Barley flour | 15:1 | + | |||||
| Corn | 15:1 | + | |||||
| Molasses (40 fish/m3) | 15:1 | 112 days | 15:1 | + | [31] | ||
| Molasses (80 fish/m3) | 15:1 | + | |||||
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | 15:1 | + | [32] | |
| Sugar (12 kg/m3) | 15:1 | + | |||||
| Glucose | 20:1 | 8 weeks | 20:1 | + | [8] | ||
| Corn starch | 15:1 | 60 days | - | × | [34] | ||
| Rice bran (4.5 kg/m3) | 20:1 | 60 days | 20:1 | + | [33] | ||
| Sugarcane molasses (4.5 kg/m3) | - | × | |||||
| Rice bran + sugarcane molasses (4.5 kg/m3) | 20:1 | + | |||||
| Cyprinus carpio | Sugar beet molasses | 20:1 | 70 days | - | × | [35] | |
| Sugar | - | × | |||||
| Corn starch | - | × | |||||
| Sugar beet molasses | 20:1 | 10 weeks | - | × | [44] | ||
| Sugar | - | × | |||||
| Corn starch | - | × | |||||
| Molasses | 20:1 | 30 days | 20:1 | + | [36] | ||
| Clarias gariepinus | Tapioca (4 fish/L) | 10:1 | 20 days | 10:1 | + | [19] | |
| Tapioca (6 fish/L) | 10:1 | + | |||||
| Tapioca (8 fish/L) | 10:1 | + | |||||
| Sucrose | 15:1 | 6 weeks | - | × | [20] | ||
| Glycerol | - | × | |||||
| Rice bran | - | × | |||||
| Glycerol | 15:1 | 8 weeks | - | × | [37] | ||
| Carassius auratus | Starch | 15:1, 20:1, 25:1 | 56 days | 20:1, 25:1 | + | [27] | |
| Carassius auratus gibelio | Glucose | 20:1 | 8 weeks | 20:1 | + | [8] | |
| Mugil cephalus | Sucrose | 15:1 | 60 days | 15:1 | + | [21] | |
| Heteropneustes fossilis | Sugarcane molasses | 10:1 | 120 days | 10:1 | + | [29] | |
| Lemon fin barb hybrid (Hypsibarbus wetmorei × Barboides gonionotus ) | Glycerol | 15:1 | 8 weeks | 15:1 | + | [37] | |
| Lepomis macrochirus | Corn starch | 15:1 | 32 days | 15:1 | - | [39] | |
| Sucrose-sugar | 15:1 | - | |||||
| Labeo rohita | Tapioca | 15:1 | 60 days | 15:1 | + | [45] | |
| Wheat | 15:1 | + | |||||
| Corn | 15:1 | + | |||||
| Sugar bagasse | 15:1 | + | |||||
| Seawater | Paralichthys olivaceus | Glucose | <10:1 | 4 months | 10:1 | + | [27] |
| Oreochromis niloticus sp. | Cornmeal + molasses 120 fish/m3) | 15:1 | 7 weeks | - | × | [13] | |
| Cornmeal + molasses (240 fish/m3) | - | × | |||||
| Feed conversion rate (FCR) | |||||||
| Freshwater | Oreochromis niloticus | Molasses | 8.4:1 | 12 weeks | 8.4:1 | - | [22] |
| Molasses (salinity level 4 g/L) | 6:1 | 70 days | - | × | [23] | ||
| Molasses (salinity level 8 g/L) | - | × | |||||
| Molasses (salinity level 12 g/L) | - | × | |||||
| Molasses (salinity level 16 g/L) | 6:1 | + | |||||
| Glycerol | 15:1 | 12 weeks | 15:1 | - | [24] | ||
| Mannan oligosaccharides | 15:1 | - | |||||
| Glucose (166 organisms/m3) | 15:1 | 120 days | 15:1 | - | [41] | ||
| Glucose (333 organisms/m3) | 15:1 | - | |||||
| Glucose (600 organisms/m3) | 15:1 | - | |||||
| Glucose | 10:1, 15:1, 20:1 | 120 days | 10:1, 15:1 | + | [42] | ||
| Rice bran and molasses (1:1) (60 fish/m3) | 15:1 | 20 weeks | 15:1 | - | [43] | ||
| Rice bran and molasses (1:1) (80 fish/m3) | 15:1 | - | |||||
| Rice bran | 15:1 | 10 weeks | 15:1 | - | [28] | ||
| Wheat-milling by-product | 15:1 | - | |||||
| Sucrose | > 10:1 | 87 days | > 10:1 | - | [25] | ||
| Glucose | 15:1 | 8 weeks | 15:1 | - | [43] | ||
| 100% molasses | 15:1, 20:1 | 8 weeks | 15:1, 20:1 | - | [26] | ||
| 100% wheat flour | 15:1, 20:1 | - | |||||
| 75% molasses + 25% wheat flour | 15:1, 20:1 | - | |||||
| 50% molasses + 50% wheat flour | 15:1, 20:1 | - | |||||
| 25% molasses + 75% wheat flour | 15:1, 20:1 | - | |||||
| Molasses | 15:1 | 37 days | 15:1 | + | [17] | ||
| Starch | 15:1 | + | |||||
| Barley flour | 15:1 | + | |||||
| Corn | 15:1 | + | |||||
| Molasses (40 fish/m3) | 15:1 | 112 days | 15:1 | + | [33] | ||
| Molasses (80 fish/m3) | 15:1 | + | |||||
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | 15:1 | - | [32] | |
| Sugar (12 kg/m3) | 15:1 | - | |||||
| Glucose | 20:1 | 8 weeks | 20:1 | - | [8] | ||
| Corn starch | 15:1 | 60 days | 15:1 | - | [34] | ||
| Rice bran | 20:1 | 60 days | 20:1 | - | [33] | ||
| Sugarcane molasses | - | × | |||||
| Rice bran + sugarcane molasses | 20:1 | - | |||||
| Sugar beet molasses | 20:1 | 70 days | 20:1 | - | [35] | ||
| Sugar | 20:1 | - | |||||
| Corn starch | 20:1 | - | |||||
| Sugar beet molasses | 20:1 | 10 weeks | 20:1 | - | [44] | ||
| Sugar | 20:1 | - | |||||
| Corn starch | 20:1 | - | |||||
| Clarias gariepinus | Tapioca (4 fish/L) | 10:1 | 20 days | 10:1 | - | [19] | |
| Tapioca (6 fish/L) | 10:1 | - | |||||
| Tapioca (8 fish/L) | 10:1 | - | |||||
| Glycerol | 15:1 | 8 weeks | - | × | [37] | ||
| Carassius auratus | Starch | 15:1, 20:1, 25:1 | 56 days | 20:1, 25:1 | - | [27] | |
| Carassius auratus gibelio | Glucose | 20:1 | 8 weeks | 20:1 | - | [8] | |
| Mugil cephalus | Sucrose | 15:1 | 60 days | 15:1 | - | [21] | |
| Heteropneustes fossilis | Sugarcane molasses | 10:1 | 120 days | 10:1 | - | [38] | |
| Lemon fin barb hybrid (Hypsibarbus wetmorei × Barboides gonionotus ) | Glycerol | 15:1 | 8 weeks | 15:1 | - | [37] | |
| Labeo rohita | Tapioca | 15:1 | 60 days | 15:1 | - | [45] | |
| Wheat | 15:1 | - | |||||
| Corn | 15:1 | - | |||||
| Sugar bagasse | 15:1 | - | |||||
| Seawater | Oreochromis niloticus sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | - | × | [13] |
| Cornmeal + molasses (240 fish/m3) | - | × | |||||
3. Hematological Parameters
| Species | Carbon Source | C:N Ratio | Period | Target Organ | Response C/N Ratio | Response | Reference | |
|---|---|---|---|---|---|---|---|---|
| Red blood cell (RBC) | ||||||||
| Freshwater | Oreochromis niloticus | Glycerol (140 fish/m3) | 15:1 | 98 days | Blood | 15:1 | + | [47] |
| Glycerol (280 fish/m3) | 15:1 | + | ||||||
| Molasses (140 fish/m3) | 15:1 | 98 days | Blood | 15:1 | + | |||
| Molasses (280 fish/m3) | 15:1 | + | ||||||
| Starch (140 fish/m3) | 15:1 | 98 days | Blood | 15:1 | + | |||
| Starch (280 fish/m3) | 15:1 | + | ||||||
| Glucose | 15:1 | 8 weeks | Blood | - | × | [29] | ||
| Molasses (40 fish/m3) | 15:1 | 112 days | Blood | - | × | [31] | ||
| Molasses (80 fish/m3) | - | × | ||||||
| Cyprinus carpio | Rice bran | 20:1 | 60 days | Blood | - | × | [33] | |
| Sugarcane molasses | - | × | ||||||
| Rice bran + sugarcane molasses | 20:1 | + | ||||||
| Corn starch | 15:1 | 60 days | Blood | - | × | [34] | ||
| Corn starch (10% of daily feed deducted) | - | × | ||||||
| Sugar beet molasses | 20:1 | 10 weeks | Blood | - | × | [44] | ||
| Sugar | - | × | ||||||
| corn starch | - | × | ||||||
| Clarias gariepinus | Tapioca (4 fish/L) | 10:1 | 20 days | Blood | - | × | [19] | |
| Tapioca (6 fish/L) | 10:1 | + | ||||||
| Tapioca (8 fish/L) | - | × | ||||||
| Heteropneustes fossilis | Sugarcane molasses | 10:1 | 120 days | Blood | 10:1 | + | [38] | |
| White blood cell (WBC) | ||||||||
| Freshwater | Oreochromis niloticus | Rice bran | 15:1 | 10 weeks | Blood | 15:1 | + | [28] |
| Wheat-milling by-product | 15:1 | + | ||||||
| Glycerol (140 fish/m3) | 15:1 | 98 days | Blood | 15:1 | - | [47] | ||
| Glycerol (280 fish/m3) | 15:1 | - | ||||||
| Molasses (140 fish/m3) | 15:1 | 98 days | Blood | 15:1 | - | |||
| Molasses (280 fish/m3) | 15:1 | - | ||||||
| Starch (140 fish/m3) | 15:1 | 98 days | Blood | 15:1 | - | |||
| Starch (280 fish/m3) | - | × | ||||||
| Glucose | 15:1 | 8 weeks | Blood | - | × | [29] | ||
| Cyprinus carpio | Rice bran | 20:1 | 60 days | Blood | - | × | [33] | |
| Sugarcane molasses | - | × | ||||||
| Rice bran + sugarcane molasses | - | × | ||||||
| Corn starch | 15:1 | 60 days | Blood | - | × | [34] | ||
| Sugar beet molasses | 20:1 | 10 weeks | Blood | - | × | [44] | ||
| Sugar | - | × | ||||||
| corn starch | - | × | ||||||
| Clarias gariepinus | Tapioca (4 fish/L) | 10:1 | 20 days | Blood | 10:1 | + | [19] | |
| Tapioca (6 fish/L) | 10:1 | + | ||||||
| Tapioca (8 fish/L) | 10:1 | + | ||||||
| Heteropneustes fossilis | Sugarcane molasses | 10:1 | 120 days | Blood | 10:1 | + | [38] | |
| Hematocrit (Ht) | ||||||||
| Freshwater | Oreochromis niloticus | Rice bran | 15:1 | 10 weeks | Blood | 15:1 | + | [28] |
| Wheat-milling by-product | 15:1 | + | ||||||
| Glucose | 15:1 | 8 weeks | Blood | - | × | [29] | ||
| Cyprinus carpio | Sugar beet molasses | 20:1 | 10 weeks | Blood | - | × | [44] | |
| Sugar | - | × | ||||||
| Corn starch | - | × | ||||||
| Corn starch | 15:1 | 60 days | Blood | - | × | [34] | ||
| Clarias gariepinus | Tapioca (4 fish/L) | 10:1 | 20 days | Blood | - | × | [19] | |
| Tapioca (6 fish/L) | - | × | ||||||
| Tapioca (8 fish/L) | - | × | ||||||
| Seawater | Paralichthys olivaceus | Glucose | <10:1 | 4 months | Blood | - | × | [27] |
| Hemoglobin (Hb) | ||||||||
| Freshwater | Oreochromis niloticus | Glycerol (140 fish/m3) | 15:1 | 98 days | Blood | 15:1 | + | [47] |
| Glycerol (280 fish/m3) | 15:1 | + | ||||||
| Molasses (140 fish/m3) | 15:1 | 98 days | Blood | 15:1 | + | |||
| Molasses (280 fish/m3) | 15:1 | + | ||||||
| Starch (140 fish/m3) | 15:1 | 98 days | Blood | 15:1 | + | |||
| Starch (280 fish/m3) | 15:1 | + | ||||||
| Glucose | 15:1 | 8 weeks | Blood | - | × | [29] | ||
| Molasses (40 fish/m3) | 15:1 | 112 days | Blood | 15:1 | + | [31] | ||
| Molasses (80 fish/m3) | - | × | ||||||
| Cyprinus carpio | Rice bran (4.5 kg/m3) | 20:1 | 60 days | Blood | - | × | [33] | |
| Sugarcane molasses (4.5 kg/m3) | 20:1 | - | ||||||
| Rice bran + sugarcane molasses (4.5 kg/m3) | - | × | ||||||
| Clarias gariepinus | Tapioca (4 fish/L) | 10:1 | 20 days | Blood | 10:1 | + | [19] | |
| Tapioca (6 fish/L) | - | × | ||||||
| Tapioca (8 fish/L) | 10:1 | + | ||||||
| Heteropneustes fossilis | Sugarcane molasses | 10:1 | 120 days | Blood | 10:1 | + | [38] | |
| Pangasianodon hypophthalmus | Tapioca | 15:1 | 90 days | Blood | 15:1 | + | [54] | |
| Sorghum | 15:1 | + | ||||||
| Pearl millet | 15:1 | + | ||||||
| Finger millet | 15:1 | + | ||||||
| Seawater | Paralichthys olivaceus | Glucose | <10:1 | 4 months | Blood | - | × | [27] |
| Glucose | ||||||||
| Freshwater | Oreochromis niloticus | Glycerol (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | - | [48] |
| Glycerol (280 fish/m3) | 15:1 | - | ||||||
| Molasses (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | - | |||
| Molasses (280 fish/m3) | 15:1 | - | ||||||
| Starch (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | - | |||
| Starch (280 fish/m3) | 15:1 | - | ||||||
| Sucrose | >10:1 | 87 days | Serum | - | × | [25] | ||
| Wheat flour (200 fish/m3) | 15:1 | 90 days | Serum | 15:1 | - | [30] | ||
| Wheat flour (250 fish/m3) | 15:1 | - | ||||||
| Wheat flour (300 fish/m3) | 15:1 | - | ||||||
| Wheat flour (350 fish/m3) | 15:1 | - | ||||||
| Molasses (40 fish/m3) | 15:1 | 112 days | Plasma | 15:1 | - | [31] | ||
| Molasses (80 fish/m3) | - | × | ||||||
| Genetically Improved Farmed Tilapia | Spentwash | 10:1 | 180 days | Serum | 10:1 | - | [73] | |
| Labeo rohita | Tapioca | 15:1 | 20 days | Serum | 15:1 | - | [45] | |
| 40 days | 15:1 | - | ||||||
| 60 days | 15:1 | - | ||||||
| Wheat | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | 15:1 | - | ||||||
| 60 days | 15:1 | - | ||||||
| Corn | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | 15:1 | - | ||||||
| 60 days | - | × | ||||||
| Sugar bagasse | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | 15:1 | - | ||||||
| 60 days | 15:1 | - | ||||||
| Molasses | 15:1 | 16 weeks | Serum | - | × | [74] | ||
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | 15:1 | - | [32] | |
| Sugar (12 kg/m3) | 15:1 | - | ||||||
| Rice bran (4.5 kg/m3) | 20:1 | 60 days | Serum | - | × | [33] | ||
| Sugarcane molasses (4.5 kg/m3) | 20:1 | + | ||||||
| Rice bran + Sugarcane molasses (4.5 kg/m3) | - | × | ||||||
| Sugar beet molasses | 20:1 | 10 weeks | Serum | - | × | [44] | ||
| Sugar | - | × | ||||||
| Corn starch | - | × | ||||||
| Clarias gariepinus | Sucrose | 15:1 | 6 weeks | Plasma | - | × | [20] | |
| Glycerol | - | × | ||||||
| Rice bran | - | × | ||||||
| Pangasianodon hypophthalmus | Tapioca | 15:1 | 90 days | Serum | 15:1 | - | [55] | |
| Sorghum | 15:1 | - | ||||||
| Pearl millet | 15:1 | - | ||||||
| Finger millet | 15:1 | - | ||||||
| Brackish water | Mugil cephalus | Sucrose | 15:1 | 60 days | Serum | - | × | [30] |
| Chanos chanos | Sorghum | 15:1 | 45 days | Serum | 15:1 | - | [58] | |
| 90 days | 15:1 | - | ||||||
| Potato | 15:1 | 45 days | Serum | 15:1 | - | |||
| 90 days | 15:1 | - | ||||||
| Yam | 15:1 | 45 days | Serum | 15:1 | - | |||
| 90 days | 15:1 | - | ||||||
| Glucose | 15:1 | 45 days | Serum | 15:1 | - | |||
| 90 days | 15:1 | - | ||||||
| Seawater | Paralichthys olivaceus | Glucose | <10:1 | 4 months | Plasma | <10:1 | - | [27] |
| Oreochromis sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | Plasma | - | × | [13] | |
| Cornmeal + molasses (240 fish/m3) | - | × | ||||||
| Cholesterol | ||||||||
| Freshwater | Oreochromis niloticus | Molasses (40 fish/m3) | 15:1 | 112 days | Plasma | - | × | [31] |
| Molasses (80 fish/m3) | 15:1 | - | ||||||
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | - | × | [32] | |
| Sugar (12 kg/m3) | 15:1 | + | ||||||
| Rice bran (4.5 kg/m3) | 20:1 | 60 days | Serum | - | × | [33] | ||
| Sugarcane molasses (4.5 kg/m3) | - | × | ||||||
| Rice bran + sugarcane molasses (4.5 kg/m3) | - | × | ||||||
| Corn starch | 15:1 | 60 days | Plasma | - | × | [34] | ||
| Sugar beet molasses | 20:1 | 10 weeks | Serum | - | × | [44] | ||
| Sugar | - | × | ||||||
| Corn starch | - | × | ||||||
| Seawater | Paralichthys olivaceus | Glucose | <10:1 | 4 months | Plasma | - | × | [27] |
| Oreochromis sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | Plasma | 15:1 | - | [13] | |
| Cornmeal + molasses (240 fish/m3) | - | × | ||||||
| Total protein | ||||||||
| Freshwater | Oreochromis niloticus | Rice bran | 15:1 | 10 weeks | Plasma | 15:1 | + | [28] |
| Wheat-milling by-product | 15:1 | + | ||||||
| Glycerol (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | + | [47] | ||
| Glycerol (280 fish/m3) | - | × | ||||||
| Molasses (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | + | |||
| Molasses (280 fish/m3) | 15:1 | + | ||||||
| Starch (140 fish/m3) | 15:1 | 98 days | Serum | - | × | |||
| Starch (280 fish/m3) | 15:1 | + | ||||||
| Glucose | 15:1 | 8 weeks | Serum | - | × | [29] | ||
| Mannan oligosaccharides | 15:1 | 12 weeks | Serum | 15:1 | + | [24] | ||
| Glycerol | 15:1 | + | ||||||
| 100% molasses | 15:1, 20:1 | 8 weeks | Serum | 15:1, 20:1 | + | [26] | ||
| 100% wheat flour | 15:1, 20:1 | + | ||||||
| 75% molasses + 25% wheat flour | 15:1, 20:1 | + | ||||||
| 50% molasses + 50 wheat flour | 15:1, 20:1 | + | ||||||
| 25% molasses + 75% wheat flour | 15:1, 20:1 | + | ||||||
| Molasses (40 fish/m3) | 15:1 | 112 days | Plasma | - | × | [31] | ||
| Molasses (80 fish/m3) | 15:1 | - | ||||||
| Genetically Improved Farmed Tilapia | Spentwash | 10:1 | 180 days | Serum | 10:1 | + | [73] | |
| Labeo rohita | Tapioca | 15:1 | 20 days | Serum | - | × | [45] | |
| 40 days | 15:1 | + | ||||||
| 60 days | 15:1 | + | ||||||
| Wheat | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | - | × | ||||||
| 60 days | - | × | ||||||
| Corn | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | - | × | ||||||
| 60 days | 15:1 | - | ||||||
| Sugar bagasse | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | - | × | ||||||
| 60 days | - | × | ||||||
| Molasses | 15:1 | 16 weeks | Serum | - | × | [74] | ||
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | - | × | [32] | |
| Sugar (12 kg/m3) | 15:1 | + | ||||||
| Rice bran (4.5 kg/m3) | 20:1 | 60 days | Serum | - | × | [33] | ||
| Sugarcane molasses (4.5 kg/m3) | 20:1 | + | ||||||
| Rice bran + sugarcane molasses (4.5 kg/m3) | 20:1 | + | ||||||
| Corn starch | 15:1 | 60 days | Plasma | - | × | [34] | ||
| Sugar beet molasses | 20:1 | 10 weeks | Serum | 20:1 | + | [44] | ||
| Sugar | 20:1 | + | ||||||
| Corn starch | 20:1 | + | ||||||
| Pangasianodon hypophthalmus | Tapioca | 15:1 | 90 days | Serum | 15:1 | + | [55] | |
| Sorghum | 15:1 | + | ||||||
| Pearl millet | 15:1 | + | ||||||
| Finger millet | 15:1 | + | ||||||
| Brackish water | Mugil cephalus | Sucrose | 15:1 | 60 days | Serum | - | × | [21] |
| Chanos chanos | Sorghum | 15:1 | 45 days | Serum | 15:1 | + | [58] | |
| 90 days | 15:1 | + | ||||||
| Potato | 15:1 | 45 days | Serum | - | × | |||
| 90 days | 15:1 | + | ||||||
| Yam | 15:1 | 45 days | Serum | 15:1 | + | |||
| 90 days | 15:1 | + | ||||||
| Glucose | 15:1 | 45 days | Serum | 15:1 | + | |||
| 90 days | 15:1 | + | ||||||
| Seawater | Paralichthys olivaceus | Glucose | <10:1 | 4 months | Plasma | - | × | [28] |
| Oreochromis sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | Plasma | 15:1 | - | [13] | |
| Cornmeal + molasses (240 fish/m3) | - | × | ||||||
| Albumin | ||||||||
| Freshwater | Oreochromis niloticus | Rice bran | 15:1 | 10 weeks | Plasma | 15:1 | + | [28] |
| Wheat-milling by-product | 15:1 | + | ||||||
| Glycerol (140 fish/m3) | 15:1 | 98 days | Serum | - | × | [47] | ||
| Glycerol (280 fish/m3) | - | × | ||||||
| Molasses (140 fish/m3) | 15:1 | 98 days | Serum | - | × | |||
| Molasses (280 fish/m3) | - | × | ||||||
| Starch (140 fish/m3) | 15:1 | 98 days | Serum | - | × | |||
| Starch (280 fish/m3) | 15:1 | + | ||||||
| Mannan oligosaccharides | 15:1 | 12 weeks | Serum | 15:1 | + | [24] | ||
| Glycerol | 15:1 | + | ||||||
| 100% molasses | 15:1, 20:1 | 8 weeks | Serum | - | × | [26] | ||
| 100% wheat flour | 15:1, 20:1 | + | ||||||
| 75% molasses + 25% wheat flour | - | × | ||||||
| 50% molasses + 50 wheat flour | - | × | ||||||
| 25% molasses + 75% wheat flour | - | × | ||||||
| Labeo rohita | Tapioca | 15:1 | 20 days | Serum | 15:1 | - | [45] | |
| 40 days | 15:1 | + | ||||||
| 60 days | - | × | ||||||
| Wheat | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | - | × | ||||||
| 60 days | 15:1 | - | ||||||
| Corn | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | 15:1 | + | ||||||
| 60 days | 15:1 | + | ||||||
| Sugar bagasse | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | - | × | ||||||
| 60 days | - | × | ||||||
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | 15:1 | - | [32] | |
| Sugar (12 kg/m3) | 15:1 | + | ||||||
| Rice bran (4.5 kg/m3) | 20:1 | 60 days | Serum | - | × | [33] | ||
| Sugarcane molasses (4.5 kg/m3) | - | × | ||||||
| Rice bran + sugarcane molasses (4.5 kg/m3) | - | × | ||||||
| Corn starch | 15:1 | 60 days | Plasma | - | × | [34] | ||
| Sugar beet molasses | 20:1 | 10 weeks | Serum | - | × | [44] | ||
| Sugar | - | × | ||||||
| Corn starch | - | × | ||||||
| Pangasianodon hypophthalmus | Tapioca | 15:1 | 90 days | Serum | 15:1 | + | [55] | |
| Sorghum | 15:1 | + | ||||||
| Pearl millet | 15:1 | + | ||||||
| Finger millet | 15:1 | + | ||||||
| Brackish water | Mugil cephalus | Sucrose | 15:1 | 60 days | Serum | - | × | [21] |
| Chanos chanos | Sorghum | 15:1 | 45 days | Serum | 15:1 | + | [58] | |
| 90 days | 15:1 | + | ||||||
| Potato | 15:1 | 45 days | Serum | 15:1 | + | |||
| 90 days | 15:1 | + | ||||||
| Yam | 15:1 | 45 days | Serum | 15:1 | + | |||
| 90 days | 15:1 | + | ||||||
| Glucose | 15:1 | 45 days | Serum | 15:1 | + | |||
| 90 days | 15:1 | + | ||||||
| Sea water | Oreochromis sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | Plasma | 15:1 | - | [8] |
| Cornmeal + molasses (240 fish/m3) | 15:1 | + | ||||||
| Globulin | ||||||||
| Freshwater | Oreochromis niloticus | Rice bran | 15:1 | 10 weeks | Plasma | - | × | [28] |
| Wheat-milling by-product | 15:1 | + | ||||||
| Glycerol (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | + | [47] | ||
| Glycerol (280 fish/m3) | - | × | ||||||
| Molasses (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | + | |||
| Molasses (280 fish/m3) | 15:1 | + | ||||||
| Starch (140 fish/m3) | 15:1 | 98 days | Serum | - | × | |||
| Starch (280 fish/m3) | 15:1 | + | ||||||
| Mannan oligosaccharides | 15:1 | 12 weeks | Serum | 15:1 | + | [24] | ||
| Glycerol | 15:1 | + | ||||||
| 100% molasses | 15:1, 20:1 | 8 weeks | Serum | - | × | [26] | ||
| 100% wheat flour | 15:1, 20:1 | + | ||||||
| 75% molasses + 25% wheat flour | - | × | ||||||
| 50% molasses + 50% wheat flour | 15:1 | + | ||||||
| 25% molasses + 75% wheat flour | 15:1, 20:1 | + | ||||||
| Labeo rohita | Tapioca | 15:1 | 20 days | Serum | 15:1 | + | [45] | |
| 40 days | - | × | ||||||
| 60 days | 15:1 | + | ||||||
| Wheat | 15:1 | 20 days | Serum | 15:1 | + | |||
| 40 days | - | × | ||||||
| 60 days | - | × | ||||||
| Corn | 15:1 | 20 days | Serum | - | × | |||
| 40 days | 15:1 | - | ||||||
| 60 days | - | × | ||||||
| Sugar bagasse | 15:1 | 20 days | Serum | - | × | |||
| 40 days | - | × | ||||||
| 60 days | - | × | ||||||
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | - | × | [32] | |
| Sugar (12 kg/m3) | 15:1 | + | ||||||
| Rice bran (4.5 kg/m3) | 20:1 | 60 days | Serum | 20:1 | + | [33] | ||
| Sugarcane molasses (4.5 kg/m3) | 20:1 | + | ||||||
| Rice bran + sugarcane molasses (4.5 kg/m3) | 20:1 | + | ||||||
| Pangasianodon hypophthalmus | Tapioca | 15:1 | 90 days | Serum | 15:1 | + | [55] | |
| Sorghum | 15:1 | + | ||||||
| Pearl millet | 15:1 | + | ||||||
| Finger millet | 15:1 | + | ||||||
| Brackish water | Chanos chanos | Sorghum | 15:1 | 45 days | Serum | 15:1 | + | [58] |
| 90 days | 15:1 | + | ||||||
| Potato | 15:1 | 45 days | Serum | - | × | |||
| 90 days | 15:1 | + | ||||||
| Yam | 15:1 | 45 days | Serum | 15:1 | + | |||
| 90 days | 15:1 | + | ||||||
| Glucose | 15:1 | 45 days | Serum | 15:1 | + | |||
| 90 days | 15:1 | + | ||||||
| Sea water | Oreochromis sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | Plasma | 15:1 | + | [13] |
| Cornmeal + molasses (240 fish/m3) | 15:1 | - | ||||||
| Triglyceride | ||||||||
| Freshwater | Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | - | × | [32] |
| Sugar (12 kg/m3) | 15:1 | + | ||||||
| Rice bran (4.5 kg/m3) | 20:1 | 60 days | Serum | - | × | [33] | ||
| Sugarcane molasses (4.5 kg/m3) | - | × | ||||||
| Rice bran + sugarcane molasses (4.5 kg/m3) | - | × | ||||||
| Sugar beet molasses | 20:1 | 10 weeks | Serum | - | × | [44] | ||
| Sugar | - | × | ||||||
| Corn starch | - | × | ||||||
| Clarias gariepinus | Sucrose | 15:1 | 6 weeks | Plasma | - | × | [20] | |
| Glycerol | 15:1 | + | ||||||
| Rice bran | - | × | ||||||
| Aspartate aminotransferase (AST) | ||||||||
| Freshwater | Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | 15:1 | - | [32] |
| Sugar (12 kg/m3) | 15:1 | - | ||||||
| Clarias gariepinus | Sucrose | 15:1 | 6 weeks | Plasma | - | × | [20] | |
| Glycerol | - | × | ||||||
| Rice bran | - | × | ||||||
| Opsariichthys kaopingensis | Glucose | 15:1, 20:1, 25:1 | 28 days | Serum | 15:1, 20:1, 25:1 | - | [68] | |
| Sea water | Paralichthys olivaceus | Glucose | <10:1 | 4 months | Plasma | <10:1 | - | [28] |
| Alanine aminotransminase (ALT) | ||||||||
| Freshwater | Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | 15:1 | - | [32] |
| Sugar (12 kg/m3) | 15:1 | - | ||||||
| Clarias gariepinus | Sucrose | 15:1 | 6 weeks | Plasma | - | × | [20] | |
| Glycerol | - | × | ||||||
| Rice bran | - | × | ||||||
| Opsariichthys kaopingensis | Glucose | 15:1, 20:1, 25:1 | 28 days | Serum | 20:1 | - | [68] | |
| Sea water | Paralichthys olivaceus | Glucose | <10:1 | 4 months | Plasma | <10:1 | - | [28] |
| Alkaline phosphatase (ALP) | ||||||||
| Freshwater | Oreochromis niloticus | Sucrose | >10:1 | 87 days | Serum | - | × | [25] |
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | - | × | [32] | |
| Sugar (12 kg/m3) | 15:1 | + | ||||||
| Seawater | Paralichthys olivaceus | Glucose | <10:1 | 4 months | Plasma | - | × | [28] |
| Cortisol | ||||||||
| Freshwater | Oreochromis niloticus | Wheat flour (+35% crude protein) | 8.4:1 | 12 weeks | Plasma | - | × | [22] |
| Glycerol (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | - | [31] | ||
| Glycerol (280 fish/m3) | 15:1 | - | ||||||
| Molasses (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | - | |||
| Molasses (280 fish/m3) | 15:1 | - | ||||||
| Starch (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | - | |||
| Starch (280 fish/m3) | 15:1 | - | ||||||
| Wheat flour (200 fish/m3) | 15:1 | 90 days | Serum | 15:1 | - | [63] | ||
| Wheat flour (250 fish/m3) | 15:1 | - | ||||||
| Wheat flour (300 fish/m3) | 15:1 | - | ||||||
| Wheat flour (350 fish/m3) | 15:1 | - | ||||||
| Labeo rohita | Tapioca | 15:1 | 20 days | Serum | 15:1 | - | [45] | |
| 40 days | - | × | ||||||
| 60 days | 15:1 | - | ||||||
| Wheat | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | 15:1 | - | ||||||
| 60 days | 15:1 | - | ||||||
| Corn | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | 15:1 | - | ||||||
| 60 days | 15:1 | + | ||||||
| Sugar bagasse | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | - | × | ||||||
| 60 days | 15:1 | - | ||||||
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | - | × | [32] | |
| Sugar (12 kg/m3) | 15:1 | - | ||||||
| Corn starch | 15:1 | 60 days | Plasma | 15:1 | - | [34] | ||
| Pangasianodon hypophthalmus | Tapioca | 15:1 | 90 days | Serum | 15:1 | - | [55] | |
| Sorghum | 15:1 | - | ||||||
| Pearl millet | 15:1 | - | ||||||
| Finger millet | 15:1 | - | ||||||
| Brackish water | Mugil cephalus | Sucrose | 15:1 | 60 days | Serum | - | × | [21] |
| Chanos chanos | Sorghum | 15:1 | 45 days | Serum | 15:1 | - | [58] | |
| 90 days | 15:1 | - | ||||||
| Potato | 15:1 | 45 days | Serum | 15:1 | - | |||
| 90 days | 15:1 | - | ||||||
| Yam | 15:1 | 45 days | Serum | 15:1 | - | |||
| 90 days | 15:1 | - | ||||||
| Glucose | 15:1 | 45 days | Serum | 15:1 | - | |||
| 90 days | 15:1 | - | ||||||
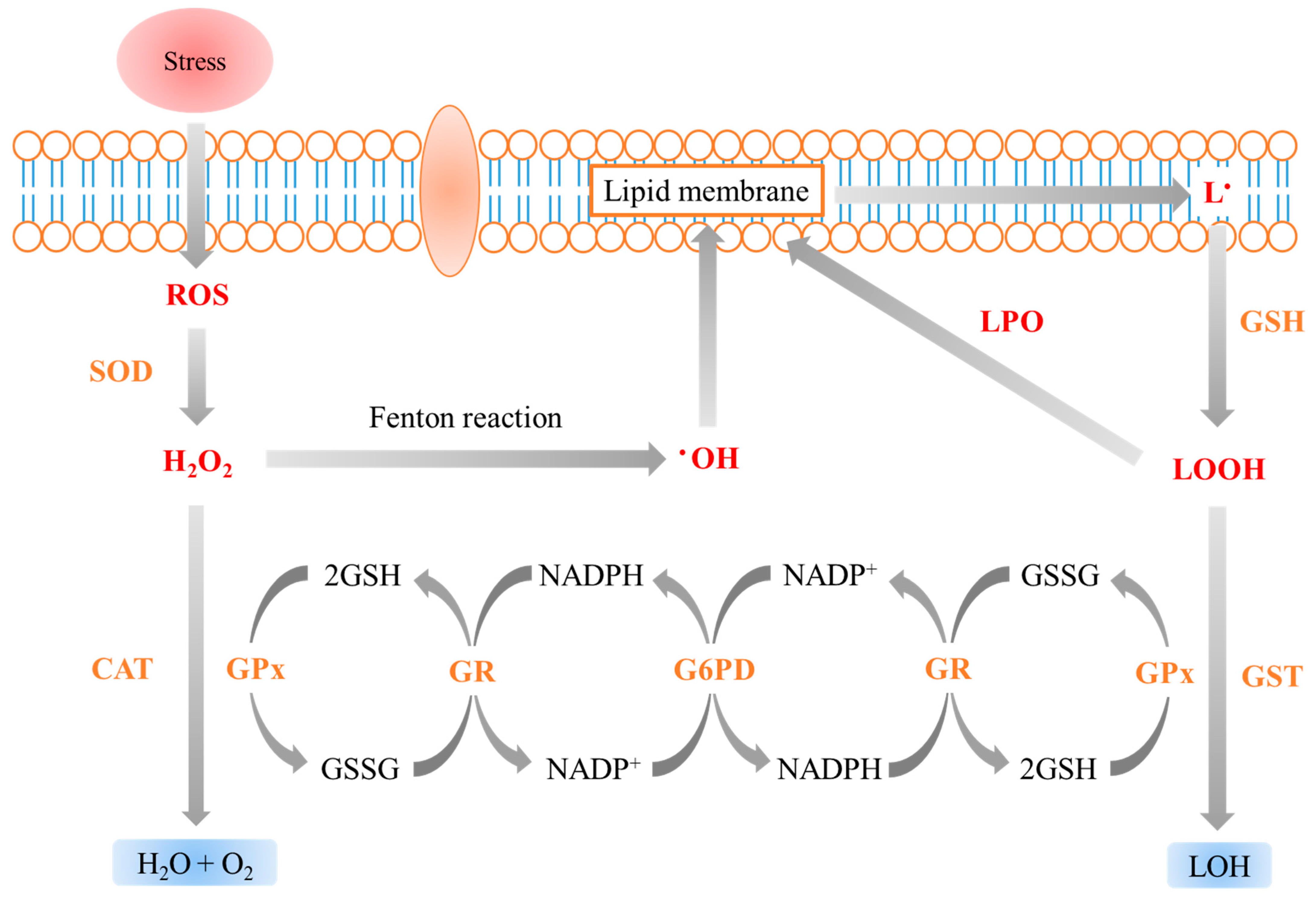
4. Antioxidant Responses
| Species | Carbon Source | C:N Ratio | Period | Target Organs | Response C:N Ratio | Response | Reference | |
|---|---|---|---|---|---|---|---|---|
| Total antioxidant capacity (TAC) | ||||||||
| Freshwater | Cyprinus carpio | Sugar beet molasses | 20:1 | 10 weeks | Serum | 20:1 | + | [44] |
| Sugar | 20:1 | + | ||||||
| Corn starch | 20:1 | + | ||||||
| Opsariichthys kaopingensis | Glucose | 15:1, 20:1, 25:1 | 28 days | Gills | 20:1, 25:1 | + | [68] | |
| Kidney | 15:1, 20:1 25:1 | + | ||||||
| Brain | 20:1 | + | ||||||
| Liver | 20:1 | + | ||||||
| Gut | 15:1, 20:1, 25:1 | + | ||||||
| Serum | 20:1, 25:1 | + | ||||||
| Carassius auratus | Anhydrous glucose | 10:1, 15:1, 20:1, 25:1 | 8 weeks | Gut | 15:1, 20:1, 25:1 | + | [79] | |
| Kidney | 10:1, 15:1, 20:1, 25:1 | + | ||||||
| Liver | 15:1, 20:1, 25:1 | + | ||||||
| Channa argus | Glucose | 10:1, 15:1, 20:1 | 8 weeks | Liver | 15:1 | + | [80] | |
| Intestine | 15:1 | + | ||||||
| Supero×ide dismutase (SOD) | ||||||||
| Freshwater | Oreochromis niloticus | Wheat-milling by-product | 15:1 | 10 weeks | Plasma | 15:1 | + | [28] |
| Rice bran | 15:1 | + | ||||||
| Glycerol (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | + | [47] | ||
| Glycerol (280 fish/m3) | 15:1 | + | ||||||
| Molasses (140 fish/m3) | 15:1 | 98 days | Serum | - | × | |||
| Molasses (280 fish/m3) | 15:1 | + | ||||||
| Starch (140 fish/m3) | 15:1 | 98 days | Serum | - | × | |||
| Starch (280 fish/m3) | - | × | ||||||
| Glucose (166 organisms/m3) | 15:1 | 120 days | Liver | 15:1 | + | [41] | ||
| Glucose (333 organisms/m3) | 15:1 | + | ||||||
| Glucose (600 organisms/m3) | - | × | ||||||
| Wheat flour (200 fish/m3) | 15:1 | 90 days | Liver | 15:1 | - | [30] | ||
| Wheat flour (250 fish/m3) | 15:1 | - | ||||||
| Wheat flour (300 fish/m3) | 15:1 | - | ||||||
| Wheat flour (350 fish/m3) | 15:1 | - | ||||||
| Glucose | 10:1, 15:1, 20:1 | 120 days | Liver | 10:1, 15:1 | + | [42] | ||
| 20:1 | - | |||||||
| Spentwash | 10:1 | 180 days | Serum | 10:1 | + | [73] | ||
| Molasses | 14:1, 17:1, 20:1 | 62 days | Liver | - | × | [86] | ||
| Cyprinus carpio | Sugar beet molasses | 20:1 | 10 weeks | Serum | - | × | [44] | |
| Sugar | - | × | ||||||
| Corn starch | - | × | ||||||
| Sugar (6 kg/m3) | 15:1 | 49 days | Serum | - | × | [32] | ||
| Sugar (12 kg/m3) | - | × | ||||||
| Rice bran | 20:1 | 60 days | Serum | - | × | [33] | ||
| Sugarcane molasses | 20:1 | + | ||||||
| Rice bran + sugarcane molasses | 20:1 | + | ||||||
| Opsariichthys kaopingensis | Glucose | 15:1, 20:1, 25:1 | 28 days | Gills | 20:1 | + | [68] | |
| Kidney | 15:1, 20:1 25:1 | + | ||||||
| Brain | 15:1, 20:1 | + | ||||||
| Liver | 20:1 | + | ||||||
| Liver | 15:1, 25:1 | - | ||||||
| Gut | 15:1, 20:1, 25:1 | + | ||||||
| Serum | 15:1, 20:1, 25:1 | + | ||||||
| Carassius auratus | Anhydrous glucose | 10:1, 15:1, 20:1, 25:1 | 8 weeks | Gut | 10:1, 15:1, 20:1, 25:1 | + | [79] | |
| Kidney | 15:1, 20:1, 25:1 | + | ||||||
| Liver | 15:1, 20:1, 25:1 | + | ||||||
| Pangasianodon hypophthalmus | Tapioca | 15:1 | 90 days | Serum | 15:1 | + | [55] | |
| Sorghum | 15:1 | + | ||||||
| Pearl millet | 15:1 | + | ||||||
| Finger millet | 15:1 | + | ||||||
| Channa argus | Glucose | 10:1, 15:1, 20:1 | 8 weeks | Liver | 15:1, 20:1 | + | [80] | |
| Intestine | 10:1, 15:1, 20:1 | + | ||||||
| Brackish water | Chanos chanos | Sorghum | 15:1 | 45 days | Liver | 15:1 | - | [58] |
| 90 days | - | × | ||||||
| Potato | 15:1 | 45 days | Liver | - | × | |||
| 90 days | - | × | ||||||
| Yam | 15:1 | 45 days | Liver | 15:1 | - | |||
| 90 days | 15:1 | - | ||||||
| Glucose | 15:1 | 45 days | Liver | 15:1 | - | |||
| 90 days | - | × | ||||||
| Seawater | Oreochromis niloticus sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | Liver | 15:1 | + | [13] |
| Cornmeal + molasses (240 fish/m3) | - | × | ||||||
| Catalase (CAT) | ||||||||
| Freshwater | Oreochromis niloticus | Wheat-milling by-product | 15:1 | 10 weeks | Plasma | 15:1 | + | [28] |
| Rice bran | 15:1 | + | ||||||
| Glycerol (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | + | [47] | ||
| Glycerol (280 fish/m3) | 15:1 | + | ||||||
| Molasses (140 fish/m3) | 15:1 | 98 days | Serum | - | × | |||
| Molasses (280 fish/m3) | 15:1 | + | ||||||
| Starch (140 fish/m3) | 15:1 | 98 days | Serum | - | × | |||
| Starch (280 fish/m3) | 15:1 | + | ||||||
| Wheat flour (200 fish/m3) | 15:1 | 90 days | Liver | 15:1 | - | [63] | ||
| Wheat flour (250 fish/m3) | 15:1 | - | ||||||
| Wheat flour (300 fish/m3) | 15:1 | - | ||||||
| Wheat flour (350 fish/m3) | 15:1 | - | ||||||
| Spentwash | 10:1 | 180 days | Serum | 10:1 | + | [73] | ||
| Molasses | 14:1, 17:1, 20:1 | 62 days | Liver | - | × | [86] | ||
| Molasses (15% food reduction) (500 fish/m3) | 15:1 | 53 days | Skin mucus | - | × | [87] | ||
| Molasses (30% food reduction) (500 fish/m3) | 15:1 | - | ||||||
| Molasses (45% food reduction) (500 fish/m3) | 15:1 | - | ||||||
| Molasses (100% food reduction) (500 fish/m3) | 15:1 | - | ||||||
| Molasses (15% food reduction) (1000 fish/m3) | 15:1 | 53 days | Skin mucus | - | × | |||
| Molasses (30% food reduction) (1000 fish/m3) | 15:1 | - | ||||||
| Molasses (45% food reduction) (1000 fish/m3) | 15:1 | - | ||||||
| Molasses (100% food reduction) (1000 fish/m3) | 15:1 | - | ||||||
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | 15:1 | + | [32] | |
| Sugar (12 kg/m3) | 15:1 | - | ||||||
| Rice bran | 20:1 | 60 days | Serum | 20:1 | + | [33] | ||
| Sugarcane molasses | 20:1 | + | ||||||
| Rice bran + sugarcane molasses | 20:1 | + | ||||||
| Opsariichthys kaopingensis | Glucose | 15:1, 20:1, 25:1 | 28 days | Gills | 20:1 | + | [68] | |
| Kidney | 20:1, 25:1 | + | ||||||
| Brain | 15:1, 20:1, 25:1 | + | ||||||
| Liver | 20:1, 25:1 | + | ||||||
| Gut | 15:1, 20:1, 25:1 | + | ||||||
| Serum | 20:1, 25:1 | + | ||||||
| Carassius auratus | Anhydrous glucose | 10:1, 15:1, 20:1, 25:1 | 8 weeks | Gut | 10:1, 15:1, 20:1, 25:1 | + | [79] | |
| Kidney | 15:1, 20:1, 25:1 | + | ||||||
| Liver | 10:1, 15:1, 20:1, 25:1 | + | ||||||
| Pangasianodon hypophthalmus | Tapioca | 15:1 | 90 days | Serum | 15:1 | + | [55] | |
| Sorghum | 15:1 | + | ||||||
| Pearl millet | 15:1 | + | ||||||
| Finger millet | 15:1 | + | ||||||
| Channa argus | Glucose | 10:1, 15:1, 20:1 | 8 weeks | Liver | 10:1, 15:1, 20:1 | + | [80] | |
| Intestine | 15:1, 20:1 | + | ||||||
| Brackish water | Chanos chanos | Sorghum | 15:1 | 45 days | Liver | 15:1 | - | [58] |
| 90 days | - | × | ||||||
| Potato | 15:1 | 45 days | Liver | 15:1 | - | |||
| 90 days | - | × | ||||||
| Yam | 15:1 | 45 days | Liver | 15:1 | - | |||
| 90 days | 15:1 | - | ||||||
| Glucose | 15:1 | 45 days | Liver | 15:1 | - | |||
| 90 days | 15:1 | - | ||||||
| Seawater | Oreochromis niloticus sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | Liver | - | × | [13] |
| Cornmeal + molasses (240 fish/m3) | - | × | ||||||
| Glutathione peroxidase (GPx) | ||||||||
| Freshwater | Oreochromis niloticus | Glucose | 15:1 | 8 weeks | Serum | 15:1 | + | [29] |
| Cyprinus carpio | Sugar beet molasses | 20:1 | 10 weeks | Serum | 20:1 | + | [44] | |
| Sugar | 20:1 | - | ||||||
| Corn starch | 20:1 | - | ||||||
| Sugar (6 kg/m3) | 15:1 | 49 days | Serum | 15:1 | + | [32] | ||
| Sugar (12 kg/m3) | 15:1 | - | ||||||
| Rice bran | 20:1 | 60 days | Serum | - | × | [33] | ||
| Sugarcane molasses | - | × | ||||||
| Rice bran + sugarcane molasses | 20:1 | + | ||||||
| Opsariichthys kaopingensis | Glucose | 15:1, 20:1, 25:1 | 28 days | Gills | 20:1, 25:1 | + | [68] | |
| Kidney | 20:1, 25:1 | + | ||||||
| Brain | 15:1, 20:1, 25:1 | + | ||||||
| Liver | 20:1 | + | ||||||
| Gut | 20:1 | + | ||||||
| Serum | 20:1, 25:1 | + | ||||||
| Carassius auratus | Anhydrous glucose | 10:1, 15:1, 20:1, 25:1 | 8 weeks | Gut | 10:1, 15:1, 20:1, 25:1 | + | [79] | |
| Kidney | 10:1, 15:1, 20:1, 25:1 | + | ||||||
| Liver | 15:1, 20:1, 25:1 | + | ||||||
| Channa argus | Glucose | 10:1, 15:1, 20:1 | 8 weeks | Liver | 15:1, | + | [80] | |
| Intestine | 10:1, 15:1, 20:1 | + | ||||||
| Seawater | Oreochromis niloticus sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | Liver | - | × | [13] |
| Cornmeal + molasses (240 fish/m3) | - | × | ||||||
| Glutathione reductase (GR) | ||||||||
| Freshwater | Oreochromis niloticus | Glycerol (140 fish/m3) | 15:1 | 98 days | Serum | 15:1 | + | [47] |
| Glycerol (280 fish/m3) | 15:1 | + | ||||||
| Molasses (140 fish/m3) | 15:1 | 98 days | Serum | - | × | |||
| Molasses (280 fish/m3) | 15:1 | + | ||||||
| Starch (140 fish/m3) | 15:1 | 98 days | Serum | - | × | |||
| Starch (280 fish/m3) | - | × | ||||||
| Seawater | Oreochromis niloticus sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | Liver | - | × | [13] |
| Cornmeal + molasses (240 fish/m3) | - | × | ||||||
| Reduced glutathione (GSH) | ||||||||
| Freshwater | Oreochromis niloticus | Glucose (166 organisms/m3) | 15:1 | 120 days | Liver | 15:1 | + | [41] |
| Glucose (333 organisms/m3) | 15:1 | + | ||||||
| Glucose (600 organisms/m3) | - | × | ||||||
| Glucose | 10:1, 15:1, 20:1 | 120 days | Liver | 10:1, 15:1 | + | [42] | ||
| Malondialdehyde (MDA) | ||||||||
| Freshwater | Oreochromis niloticus | Glucose (166 organisms/m3) | 15:1 | 120 days | Liver | 15:1 | + | [41] |
| Glucose (333 organisms/m3) | 15:1 | + | ||||||
| Glucose (600 organisms/m3) | - | × | ||||||
| Glucose | 10:1, 15:1, 20:1 | 120 days | Liver | 10:1, 15:1 | + | [42] | ||
| Cyprinus carpio | Sugar (6 kg/m3) | 15:1 | 49 days | Serum | 15:1 | + | [72] | |
| Sugar (12 kg/m3) | 15:1 | - | ||||||
| Rice bran | 20:1 | 60 days | Serum | - | × | [33] | ||
| Sugarcane molasses | 20:1 | - | ||||||
| Rice bran + sugarcane molasses | - | × | ||||||
| Clarias gariepinus | Sucrose | 15:1 | 6 weeks | Muscle | - | × | [20] | |
| Glycerol | - | × | ||||||
| Rice bran | - | × | ||||||
| Opsariichthys kaopingensis | Glucose | 15:1, 20:1, 25:1 | 28 days | Gills | - | × | [68] | |
| Kidney | 20:1 | - | ||||||
| Brain | - | × | ||||||
| Liver | 20:1, 25:1 | - | ||||||
| Gut | 20:1 | - | ||||||
| Serum | 15:1, 20:1, 25:1 | - | ||||||
| Carassius auratus | Anhydrous glucose | 10:1, 15:1, 20:1, 25:1 | 8 weeks | Gut | 20:1, 25:1 | - | [79] | |
| Kidney | 20:1, 25:1 | - | ||||||
| Liver | 15:1, 20:1, 25:1 | - | ||||||
| Channa argus | Glucose | 10:1, 15:1, 20:1 | 8 weeks | Liver | 15:1 | - | [80] | |
| Intestine | 10:1, 15:1, 20:1 | - | ||||||
| Seawater | Oreochromis niloticus sp. | Cornmeal + molasses (120 fish/m3) | 15:1 | 7 weeks | Liver | - | × | [13] |
| Cornmeal + molasses (240 fish/m3) | - | × | ||||||
5. Immune Responses
| Species | Carbon Source | C:N Ratio | Period | Target Organs | Response C:N Ratio | Response | Reference | |
|---|---|---|---|---|---|---|---|---|
| Phagocytosis | ||||||||
| Freshwater | Oreochromis niloticus | Wheat-milling by-product | 15:1 | 10 weeks | Macrophages | 15:1 | + | [28] |
| Rice bran | 15:1 | + | ||||||
| Clarias gariepinus | Tapioca flour (4 fish/L) | 10:1 | 20 days | Blood | - | × | [19] | |
| Tapioca flour (6 fish/L) | - | × | ||||||
| Tapioca flour (8 fish/L) | 10:1 | + | ||||||
| Total immunoglobulin | ||||||||
| Freshwater | Oreochromis niloticus | Wheat-milling by-product | 15:1 | 10 weeks | Plasma | 15:1 | + | [28] |
| Rice bran | 15:1 | + | ||||||
| Molasses (15% food reduction) (500 fish/m3) | 15:1 | 53 days | Skin mucus | - | × | [87] | ||
| Molasses (30% food reduction) (500 fish/m3) | - | × | ||||||
| Molasses (45% food reduction) (500 fish/m3) | - | × | ||||||
| Molasses (100% food reduction) (500 fish/m3) | 15:1 | - | ||||||
| Molasses (15% food reduction) (1000 fish/m3) | 15:1 | 53 days | Skin mucus | - | × | |||
| Molasses (30% food reduction) (1000 fish/m3) | - | × | ||||||
| Molasses (45% food reduction) (1000 fish/m3) | - | × | ||||||
| Molasses (100% food reduction) (1000 fish/m3) | - | × | ||||||
| Labeo rohita | Tapioca | 15:1 | 20 days | Plasma | - | × | [45] | |
| 40 days | 15:1 | + | ||||||
| 60 days | 15:1 | + | ||||||
| Wheat | 15:1 | 20 days | Plasma | - | × | |||
| 40 days | 15:1 | + | ||||||
| 60 days | 15:1 | + | ||||||
| Corn | 15:1 | 20 days | Plasma | - | × | |||
| 40 days | 15:1 | + | ||||||
| 60 days | - | × | ||||||
| Sugar bagasse | 15:1 | 20 days | Plasma | 15:1 | - | |||
| 40 days | 15:1 | + | ||||||
| 60 days | - | × | ||||||
| Cyprinus carpio | Sugar beet molasses | 20:1 | 10 weeks | Serum | 20:1 | + | [44] | |
| Sugar | 20:1 | + | ||||||
| Corn starch | 20:1 | + | ||||||
| Immunoglobulin M (IgM) | ||||||||
| Freshwater | Oreochromis niloticus | Glycerol | 15:1 | 12 weeks | Serum | 15:1 | + | [24] |
| Mannan oligosaccharides | 15:1 | + | ||||||
| Cyprinus carpio | Rice bran | 20:1 | 60 days | Serum | 20:1 | + | [33] | |
| Sugarcane molasses | 20:1 | + | ||||||
| Rice bran + sugarcane molasses | 20:1 | + | ||||||
| Channa argus | Glucose | 10:1, 15:1, 20:1 | 8 weeks | Serum | - | × | [80] | |
| Kidney | 15:1, 20:1 | + | ||||||
| Seawater | Paralichthys olivaceus | Glucose | 10:1 | 4 months | Plasma | 10:1 | + | [28] |
| Lysozyme activity | ||||||||
| Freshwater | Oreochromis niloticus | Wheat-milling by-product | 15:1 | 10 weeks | Plasma | 15:1 | + | [28] |
| Rice bran | 15:1 | + | ||||||
| Sucrose | >10:1 | 87 days | Hepatopancreas | - | × | [25] | ||
| Head kidney | - | × | ||||||
| Serum | - | × | ||||||
| Glucose | 15:1 | 8 weeks | Serum | 15:1 | + | [29] | ||
| Glucose (166 organisms/m3) | 15:1 | 120 days | Liver | 15:1 | + | [41] | ||
| Glucose (333 organisms/m3) | 15:1 | + | ||||||
| Glucose (600 organisms/m3) | 15:1 | - | ||||||
| Wheat flour (200 fish/m3) | 15:1 | 90 days | Serum | 15:1 | + | [30] | ||
| Wheat flour (250 fish/m3) | 15:1 | + | ||||||
| Wheat flour (300 fish/m3) | 15:1 | + | ||||||
| Wheat flour (350 fish/m3) | 15:1 | + | ||||||
| Glycerol | 15:1 | 12 weeks | Serum | 15:1 | + | [24] | ||
| Mannan oligosaccharides | 15:1 | + | ||||||
| Glucose | 10:1, 15:1, 20:1 | 120 days | Liver | 10:1, 15:1, 20:1 | + | [42] | ||
| Molasses (15% food reduction) (500 fish/m3) | 15:1 | 53 days | Skin mucus | - | × | [87] | ||
| Molasses (30% food reduction) (500 fish/m3) | 15:1 | - | ||||||
| Molasses (45% food reduction) (500 fish/m3) | 15:1 | - | ||||||
| Molasses (100% food reduction) (500 fish/m3) | 15:1 | - | ||||||
| Molasses (15% food reduction) (1000 fish/m3) | 15:1 | 53 days | Skin mucus | - | × | |||
| Molasses (30% food reduction) (1000 fish/m3) | - | × | ||||||
| Molasses (45% food reduction) (1000 fish/m3) | 15:1 | - | ||||||
| Molasses (100% food reduction) (1000 fish/m3) | 15:1 | - | ||||||
| Labeo rohita | Tapioca | 15:1 | 20 days | Serum | - | × | [45] | |
| 40 days | 15:1 | + | ||||||
| 60 days | 15:1 | + | ||||||
| Wheat | 15:1 | 20 days | Serum | 15:1 | + | |||
| 40 days | 15:1 | + | ||||||
| 60 days | 15:1 | + | ||||||
| Corn | 15:1 | 20 days | Serum | - | × | |||
| 40 days | - | × | ||||||
| 60 days | 15:1 | - | ||||||
| Sugar bagasse | 15:1 | 20 days | Serum | - | × | |||
| 40 days | - | × | ||||||
| 60 days | - | × | ||||||
| Cyprinus carpio | Sugar beet molasses | 20:1 | 10 weeks | Serum | - | × | [44] | |
| Sugar | - | × | ||||||
| Corn starch | - | × | ||||||
| Sugar (6 kg/m3) | 15:1 | 49 days | Serum | 15:1 | × | [32] | ||
| Sugar (12 kg/m3) | 15:1 | × | ||||||
| Rice bran | 20:1 | 60 days | Serum | 20:1 | + | [33] | ||
| Sugarcane molasses | - | × | ||||||
| Rice bran + sugarcane molasses | - | × | ||||||
| Opsariichthys kaopingensis | Glucose | 15:1, 20:1, 25:1 | 28 days | Gills | 20:1 | + | [68] | |
| Kidney | 15:1, 20:1 25:1 | + | ||||||
| Brain | 20:1 | + | ||||||
| Liver | 20:1, 25:1 | + | ||||||
| Gut | 15:1, 20:1, 25:1 | + | ||||||
| Carassius auratus | Anhydrous glucose | 10:1, 15:1, 20:1, 25:1 | 8 weeks | Gut | 15:1, 20:1, 25:1 | + | [79] | |
| Kidney | 15:1, 20:1, 25:1 | + | ||||||
| Liver | 20:1, 25:1 | + | ||||||
| Channa argus | Glucose | 10:1, 15:1, 20:1 | 8 weeks | Serum | 10:1, 15:1, 20:1 | + | [80] | |
| Kidney | 15:1, 20:1 | + | ||||||
| Myeloperoxidase (MPO) | ||||||||
| Freshwater | Oreochromis niloticus | Wheat-milling by-product | 15:1 | 10 weeks | Blood | 15:1 | + | [28] |
| Rice bran | 15:1 | + | ||||||
| Wheat flour (200 fish/m3) | 15:1 | 90 days | Serum | 15:1 | + | [30] | ||
| Wheat flour (250 fish/m3) | 15:1 | + | ||||||
| Wheat flour (300 fish/m3) | 15:1 | + | ||||||
| Wheat flour (350 fish/m3) | - | × | ||||||
| Spentwash | 10:1 | 180 days | Serum | 10:1 | + | [73] | ||
| Labeo rohita | Tapioca | 15:1 | 20 days | Serum | 15:1 | - | [45] | |
| 40 days | 15:1 | + | ||||||
| 60 days | 15:1 | + | ||||||
| Wheat | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | - | × | ||||||
| 60 days | 15:1 | + | ||||||
| Corn | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | - | × | ||||||
| 60 days | - | × | ||||||
| Sugar bagasse | 15:1 | 20 days | Serum | 15:1 | - | |||
| 40 days | - | × | ||||||
| 60 days | - | × | ||||||
| Respiratory burst activity | ||||||||
| Freshwater | Oreochromis niloticus | Wheat-milling by-product | 15:1 | 10 weeks | Leucocytes | 15:1 | + | [28] |
| Rice bran | 15:1 | + | ||||||
| Wheat flour (200 fish/m3) | 15:1 | 90 days | Phagocytes | 15:1 | + | [30] | ||
| Wheat flour (250 fish/m3) | 15:1 | + | ||||||
| Wheat flour (300 fish/m3) | 15:1 | + | ||||||
| Wheat flour (350 fish/m3) | 15:1 | + | ||||||
| Labeo rohita | Tapioca | 15:1 | 20 days | Leucocytes | 15:1 | + | [45] | |
| 40 days | 15:1 | + | ||||||
| 60 days | 15:1 | + | ||||||
| Wheat | 15:1 | 20 days | Leucocytes | 15:1 | + | |||
| 40 days | 15:1 | + | ||||||
| 60 days | 15:1 | + | ||||||
| Corn | 15:1 | 20 days | Leucocytes | - | × | |||
| 40 days | 15:1 | + | ||||||
| 60 days | 15:1 | + | ||||||
| Sugar bagasse | 15:1 | 20 days | Leucocytes | 15:1 | × | |||
| 40 days | 15:1 | + | ||||||
| 60 days | 15:1 | + | ||||||
| Brackish water | Chanos chanos | Sorghum | 15:1 | 45 days | Phagocytes | 15:1 | + | [58] |
| 90 days | 15:1 | + | ||||||
| Potato | 15:1 | 45 days | Phagocytes | 15:1 | + | |||
| 90 days | 15:1 | + | ||||||
| Yam | 15:1 | 45 days | Phagocytes | 15:1 | + | |||
| 90 days | 15:1 | + | ||||||
| Glucose | 15:1 | 45 days | Phagocytes | 15:1 | + | |||
| 90 days | 15:1 | + | ||||||
| Alterative complement activity (ACH50) | ||||||||
| Freshwater | Oreochromis niloticus | Wheat-milling by-product | 15:1 | 10 weeks | Plasma | 15:1 | + | [28] |
| Rice bran | 15:1 | + | ||||||
| Cyprinus carpio | Sugar beet molasses | 20:1 | 10 weeks | Serum | - | × | [44] | |
| Sugar | - | × | ||||||
| Corn starch | - | × | ||||||
| Sugar (6 kg/m3) | 15:1 | 49 days | Serum | 15:1 | × | [32] | ||
| Sugar (12 kg/m3) | 15:1 | + | ||||||
| Rice bran | 20:1 | 60 days | Serum | 20:1 | + | [30] | ||
| Sugarcane molasses | 20:1 | + | ||||||
| Rice bran + sugarcane molasses | 20:1 | + | ||||||
| Complement 3 (C3) | ||||||||
| Freshwater | Oreochromis niloticus | Glucose (166 organisms/m3) | 15:1 | 120 days | Liver | 15:1 | + | [41] |
| Glucose (333 organisms/m3) | 15:1 | + | ||||||
| Glucose (600 organisms/m3) | 15:1 | - | ||||||
| Glucose | 10:1, 15:1, 20:1 | 120 days | Liver | 15:1, 20:1 | + | [42] | ||
| Channa argus | Glucose | 10:1, 15:1, 20:1 | 8 weeks | Serum | 10:1, 15:1 | + | [80] | |
| Kidney | 15:1 | + | ||||||
| Complement 4 (C4) | ||||||||
| Freshwater | Channa argus | Glucose | 10:1, 15:1, 20:1 | 8 weeks | Serum | 10:1, 15:1, 20:1 | + | [80] |
| Kidney | 10:1, 15:1, 20:1 | + | ||||||
6. Disease Resistance
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guerrera, M.C.; Arfuso, F.; Rizzo, M.; Saoca, C.; Fazio, F.; Fortino, G.; Santulli, A.; Piccione, G. Gonadal sexual differentiation of European sea bass (Dicentrarchus labrax, L. 1758) of fingerlings in different size classes. Mar. Freshw. Behav. Physiol. 2016, 49, 347–354. [Google Scholar] [CrossRef]
- Arfuso, F.; Guerrera, M.C.; Fortino, G.; Fazio, F.; Santulli, A.; Piccione, G. Water temperature influences growth and gonad differentiation in European sea bass (Dicentrarchus labrax, L. 1758). Theriogenology 2017, 88, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Erkinharju, T.; Dalmo, R.A.; Hansen, M.; Seternes, T. Cleaner fish in aquaculture: Review on diseases and vaccination. Rev. Aquac. 2021, 13, 189–237. [Google Scholar] [CrossRef]
- Santos, L.; Ramos, F. Antimicrobial resistance in aquaculture: Current knowledge and alternatives to tackle the problem. Int. J. Antimicrob. Agents. 2018, 52, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Sohn, S.; Kim, S.K.; Hur, Y.B. Effects on hematological parameters, antioxidant and immune responses, AChE, and stress indicators of olive flounders, Paralichthys olivaceus, raised in bio-floc and seawater challenged by Edwardsiella tarda. Fish Shellfish. Immunol. 2020, 97, 194–203. [Google Scholar] [CrossRef]
- Ragasa, C.; Agyakwah, S.K.; Asmah, R.; Mensah, E.T.D.; Amewu, S.; Oyih, M. Accelerating pond aquaculture development and resilience beyond COVID: Ensuring food and jobs in Ghana. Aquaculture 2022, 547, 737476. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, S.; Liu, J.; Wang, H.; Zhu, J.; Li, D.; Zhao, R. Application of machine learning in intelligent fish aquaculture: A review. Aquaculture 2021, 540, 736724. [Google Scholar] [CrossRef]
- Liu, H.; Li, H.; Wei, H.; Zhu, X.; Han, D.; Jin, J.; Yang, Y.; Xie, S. Biofloc formation improves water quality and fish yield in a freshwater pond aquaculture system. Aquaculture 2019, 506, 256–269. [Google Scholar] [CrossRef]
- Abakari, G.; Luo, G.; Kombat, E.O. Dynamics of nitrogenous compounds and their control in biofloc technology (BFT) systems: A review. Aquacult. Fisheries. 2021, 6, 441–447. [Google Scholar] [CrossRef]
- Ekasari, J.; Rivandi, D.R.; Firdausi, A.P.; Surawidjaja, E.H.; Zairin, M., Jr.; Bossier, P.; De Schryver, P. Biofloc technology positively affects Nile tilapia (Oreochromis niloticus) larvae performance. Aquaculture 2015, 441, 72–77. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.K.; Hur, Y.B. Temperature-mediated changes in stress responses, acetylcholinesterase, and immune responses of juvenile olive flounder Paralichthys olivaceus in a bio-floc environment. Aquaculture 2019, 506, 453–458. [Google Scholar] [CrossRef]
- Diatin, I.; Shafruddin, D.; Hude, N.; Sholihah, M.A.; Mutsmir, I. Production performance and financial feasibility analysis of farming catfish (Clarias gariepinus) utilizing water exchange system, aquaponic, and biofloc technology. J. Saudi Soc. Agric. Sci. 2021, 20, 344–351. [Google Scholar] [CrossRef]
- Banuelos-Vargas, I.; de Oca, G.A.R.M.; Martinez-Montano, E.; Perez-Jimenez, A.; Mendoza-Gamboa, O.A.; Estrada-Godínez, J.A.; Hernandez, C. Antioxidant and immune response of juvenile red tilapia (Oreochromis sp.) cultured at different densities in sea water with biofloc plus probiotics. Aquaculture 2021, 544, 737112. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Y.; Xu, D.H.; Qiao, G.; Zhang, J.; Qi, Z.; Li, Q. Effect of different water biofloc contents on the growth and immune response of gibel carp cultured in zero water exchange and no feed addition system. Aquac. Res. 2018, 49, 1647–1656. [Google Scholar] [CrossRef]
- Deb, S.; Noori, M.T.; Rao, P.S. Application of biofloc technology for Indian major carp culture (polyculture) along with water quality management. Aquac. Eng. 2020, 91, 102106. [Google Scholar] [CrossRef]
- Khanjani, M.H.; Alizadeh, M.; Sharifinia, M. Effects of different carbon sources on water quality, biofloc quality, and growth performance of Nile tilapia (Oreochromis niloticus) fingerlings in a heterotrophic culture system. Aquac. Int. 2021, 29, 307–321. [Google Scholar] [CrossRef]
- Khanjani, M.H.; Alizadeh, M.; Mohammadi, M.; Sarsangi Aliabad, H. Biofloc system applied to Nile tilapia (Oreochromis niloticus) farming using different carbon sources: Growth performance, carcass analysis, digestive and hepatic enzyme activity. Iran. J. Fish. Sci. 2021, 20, 490–513. [Google Scholar] [CrossRef]
- Fauji, H.; Budiardi, T.; Ekasari, J. Growth performance and robustness of African Catfish Clarias gariepinus (Burchell) in biofloc-based nursery production with different stocking densities. Aquac. Res. 2018, 49, 1339–1346. [Google Scholar] [CrossRef]
- Dauda, A.B.; Romano, N.; Ebrahimi, M.; Karim, M.; Natrah, I.; Kamarudin, M.S.; Ekasari, J. Different carbon sources affects biofloc volume, water quality and the survival and physiology of African catfish Clarias gariepinus fingerlings reared in an intensive biofloc technology system. Fish. Sci. 2017, 83, 1037–1048. [Google Scholar] [CrossRef]
- Haridas, H.; Chadha, N.K.; Sawant, P.B.; Deo, A.D.; Ande, M.P.; Syamala, K.; Sontakke, R.; Lingam, S.S. Growth performance, digestive enzyme activity, non-specific immune response and stress enzyme status in early stages of grey mullet reared in a biofloc system. Aquac. Res. 2021, 52, 4923–4933. [Google Scholar] [CrossRef]
- Azim, M.E.; Little, D.C. The biofloc technology (BFT) in indoor tanks: Water quality, biofloc composition, and growth and welfare of Nile tilapia (Oreochromis niloticus). Aquaculture 2008, 283, 29–35. [Google Scholar] [CrossRef]
- De Alvarenga, E.R.; Alves, G.F.D.O.; Fernandes, A.F.A.; Costa, G.R.; da Silva, M.A.; Teixeira, E.D.A.; Turra, E.M. Moderate salinities enhance growth performance of Nile tilapia (Oreochromis niloticus) fingerlings in the biofloc system. Aquac. Res. 2018, 49, 2919–2926. [Google Scholar] [CrossRef]
- Kishawy, A.T.; Sewid, A.H.; Nada, H.S.; Kamel, M.A.; El-Mandrawy, S.A.; Abdelhakim, T.M.; El-Murr, A.E.I.; Nahhas, E.N.; Hozzein, W.N.; Ibrahim, D. Mannanoligosaccharides as a carbon source in Biofloc boost dietary plant protein and water quality, growth, immunity and Aeromonas hydrophila resistance in Nile tilapia (Oreochromis niloticus). Animals 2020, 10, 1724. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Gao, Q.; Wang, C.; Liu, W.; Sun, D.; Li, L.; Tan, H. Growth, digestive activity, welfare, and partial cost-effectiveness of genetically improved farmed tilapia (Oreochromis niloticus) cultured in a recirculating aquaculture system and an indoor biofloc system. Aquaculture 2014, 422, 1–7. [Google Scholar] [CrossRef]
- Mirzakhani, N.; Ebrahimi, E.; Jalali, S.A.H.; Ekasari, J. Growth performance, intestinal morphology and nonspecific immunity response of Nile tilapia (Oreochromis niloticus) fry cultured in biofloc systems with different carbon sources and input C: N ratios. Aquaculture 2019, 512, 734235. [Google Scholar] [CrossRef]
- Wang, G.; Yu, E.; Xie, J.; Yu, D.; Li, Z.; Luo, W.; Qiu, L.; Zheng, Z. Effect of C/N ratio on water quality in zero-water exchange tanks and the biofloc supplementation in feed on the growth performance of crucian carp, Carassius auratus. Aquaculture 2015, 443, 98–104. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.K.; Kim, J.H. Bio-floc technology application in flatfish Paralichthys olivaceus culture: Effects on water quality, growth, hematological parameters, and immune responses. Aquaculture 2018, 495, 703–709. [Google Scholar] [CrossRef]
- Mansour, A.T.; Esteban, M.Á. Effects of carbon sources and plant protein levels in a biofloc system on growth performance, and the immune and antioxidant status of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2017, 64, 202–209. [Google Scholar] [CrossRef]
- Long, L.; Yang, J.; Li, Y.; Guan, C.; Wu, F. Effect of biofloc technology on growth, digestive enzyme activity, hematology, and immune response of genetically improved farmed tilapia (Oreochromis niloticus). Aquaculture 2015, 448, 135–141. [Google Scholar] [CrossRef]
- Haridas, H.; Verma, A.K.; Rathore, G.; Prakash, C.; Sawant, P.B.; Babitha Rani, A.M. Enhanced growth and immuno-physiological response of Genetically Improved Farmed Tilapia in indoor biofloc units at different stocking densities. Aquac. Res. 2017, 48, 4346–4355. [Google Scholar] [CrossRef]
- Klanian, M.G.; Díaz, M.D.; Solís, M.J.S.; Aranda, J.; Moral, P.M. Effect of the content of microbial proteins and the poly-β-hydroxybutyric acid in biofloc on the performance and health of Nile tilapia (Oreochromis niloticus) fingerlings fed on a protein-restricted diet. Aquaculture 2020, 519, 734872. [Google Scholar] [CrossRef]
- Adineh, H.; Naderi, M.; Hamidi, M.K.; Harsij, M. Biofloc technology improves growth, innate immune responses, oxidative status, and resistance to acute stress in common carp (Cyprinus carpio) under high stocking density. Fish Shellfish Immunol. 2019, 95, 440–448. [Google Scholar] [CrossRef]
- Ebrahimi, A.; Akrami, R.; Najdegerami, E.H.; Ghiasvand, Z.; Koohsari, H. Effects of different protein levels and carbon sources on water quality, antioxidant status and performance of common carp (Cyprinus carpio) juveniles raised in biofloc based system. Aquaculture 2020, 516, 734639. [Google Scholar] [CrossRef]
- Tabarrok, M.; Seyfabadi, J.; Salehi Jouzani, G.; Younesi, H. Comparison between recirculating aquaculture and biofloc systems for rearing juvenile common carp (Cyprinus carpio): Growth performance, haemato-immunological indices, water quality and microbial communities. Aquac. Res. 2020, 51, 4881–4892. [Google Scholar] [CrossRef]
- Bakhshi, F.; Najdegerami, E.H.; Manaffar, R.; Tukmechi, A.; Farah, K.R. Use of different carbon sources for the biofloc system during the grow-out culture of common carp (Cyprinus carpio L.) fingerlings. Aquaculture 2018, 484, 259–267. [Google Scholar] [CrossRef]
- Najdegerami, E.H.; Bakhshi, F.; Lakani, F.B. Effects of biofloc on growth performance, digestive enzyme activities and liver histology of common carp (Cyprinus carpio L.) fingerlings in zero-water exchange system. Fish Physiol. Biochem. 2016, 42, 457–465. [Google Scholar] [CrossRef]
- Dauda, A.B.; Romano, N.; Chen, W.W.; Natrah, I.; Kamarudin, M.S. Differences in feeding habits influence the growth performance and feeding efficiencies of African catfish (Clarias gariepinus) and lemon fin barb hybrid (Hypsibarbus wetmorei ♂ × Barboides gonionotus ♀) in a glycerol-based biofloc technology system versus a recirculating system. Aquac. Eng. 2018, 82, 31–37. [Google Scholar] [CrossRef]
- Zafar, M.A.; Talha, M.A.; Rana, M.M. Effect of biofloc technology on growth performance, digestive enzyme activity, proximate composition, and hematological parameters of Asian stinging catfish (Heteropneustes fossilis). J. Appl. Aquac. 2021, 34, 1–19. [Google Scholar] [CrossRef]
- Fischer, H.; Romano, N.; Renukdas, N.; Egnew, N.; Sinha, A.K.; Ray, A.J. The potential of rearing juveniles of bluegill, Lepomis macrochirus, in a biofloc system. Aquacult. Rep. 2020, 17, 100398. [Google Scholar] [CrossRef]
- Mahanand, S.S.; Moulick, S.; Rao, P.S. Optimum formulation of feed for rohu, Labeo rohita (Hamilton), with biofloc as a component. Aquac. Int. 2013, 21, 347–360. [Google Scholar] [CrossRef]
- Liu, G.; Ye, Z.; Liu, D.; Zhao, J.; Sivaramasamy, E.; Deng, Y.; Zhu, S. Influence of stocking density on growth, digestive enzyme activities, immune responses, antioxidant of Oreochromis niloticus fingerlings in biofloc systems. Fish Shellfish Immunol. 2018, 81, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhu, S.; Liu, D.; Ye, Z. Effect of the C/N ratio on inorganic nitrogen control and the growth and physiological parameters of tilapias fingerlings, Oreochromis niloticus reared in biofloc systems. Aquac. Res. 2018, 49, 2429–2439. [Google Scholar] [CrossRef]
- Hwihy, H.; Zeina, A.; Abu Husien, M.; El-Damhougy, K. Impact of Biofloc technology on growth performance and biochemical parameters of Oreochromis niloticus. Egypt. J. Aquat. Biol. Fish. 2021, 25, 761–774. [Google Scholar] [CrossRef]
- Bakhshi, F.; Najdegerami, E.H.; Manaffar, R.; Tokmechi, A.; Rahmani Farah, K.; Shalizar Jalali, A. Growth performance, haematology, antioxidant status, immune response and histology of common carp (Cyprinus carpio L.) fed biofloc grown on different carbon sources. Aquac. Res. 2018, 49, 393–403. [Google Scholar] [CrossRef]
- Verma, A.K.; Rani, A.B.; Rathore, G.; Saharan, N.; Gora, A.H. Growth, non-specific immunity and disease resistance of Labeo rohita against Aeromonas hydrophila in biofloc systems using different carbon sources. Aquaculture 2016, 457, 61–67. [Google Scholar] [CrossRef]
- Puente-Marin, S.; Nombela, I.; Ciordia, S.; Mena, M.C.; Chico, V.; Coll, J.; Ortega-Villaizan, M.D.M. In silico functional networks identified in fish nucleated red blood cells by means of transcriptomic and proteomic profiling. Genes 2018, 9, 202. [Google Scholar] [CrossRef]
- Shourbela, R.M.; Khatab, S.A.; Hassan, M.M.; Van Doan, H.; Dawood, M.A. The effect of stocking density and carbon sources on the oxidative status, and nonspecific immunity of Nile tilapia (Oreochromis niloticus) reared under biofloc conditions. Animals 2021, 11, 184. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, D.; Zhao, J.; Chen, X. Fish red blood cells express immune genes and responses. Aquac. Fish. 2018, 3, 14–21. [Google Scholar] [CrossRef]
- Modesto, K.A.; Martinez, C.B. Effects of Roundup Transorb on fish: Hematology, antioxidant defenses and acetylcholinesterase activity. Chemosphere 2010, 81, 781–787. [Google Scholar] [CrossRef]
- Firouzbakhsh, F.; Noori, F.; Khalesi, M.K.; Jani-Khalili, K. Effects of a probiotic, protexin, on the growth performance and hematological parameters in the Oscar (Astronotus ocellatus) fingerlings. Fish Physiol. Biochem. 2011, 37, 833–842. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. The selenium accumulation and its effect on growth, and haematological parameters in red sea bream, Pagrus major, exposed to waterborne selenium. Ecotox. Environ. Saf. 2014, 104, 96–102. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. The chromium accumulation and its physiological effects in juvenile rockfish, Sebastes schlegelii, exposed to different levels of dietary chromium (Cr6+) concentrations. Environ. Toxicol. Pharmacol. 2014, 41, 152–158. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. Changes in hematological parameters, plasma cortisol, and acetylcholinesterase of juvenile rockfish, Sebastes schlegelii supplemented with the dietary ascorbic acid. Aquacult. Rep. 2016, 4, 80–85. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. Toxic effects on bioaccumulation and hematological parameters of juvenile rockfish Sebastes schlegelii exposed to dietary lead (Pb) and ascorbic acid. Chemosphere 2017, 176, 131–140. [Google Scholar] [CrossRef]
- Nageswari, P.; Verma, A.K.; Gupta, S.; Jeyakumari, A.; Mallikarjun Hittinahalli, C. Haematological, serum biochemical and anti-oxidative enzymes responses of sutchi catfish (Pangasianodon hypophthalmus) against Aeromonas hydrophila using various carbon sources in biofloc system. Aquac. Res. 2022, 53, 1851–1861. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Kumar, S.; Das, D.; Sarbajna, A.; Chakraborty, S.B. Enzymatic, non enzymatic antioxidants and glucose metabolism enzymes response differently against metal stress in muscles of three fish species depending on different feeding niche. Ecotox. Environ. Saf. 2020, 202, 110954. [Google Scholar] [CrossRef]
- Makaras, T.; Razumienė, J.; Gurevičienė, V.; Šakinytė, I.; Stankevičiūtė, M.; Kazlauskienė, N. A new approach of stress evaluation in fish using β-d-Glucose measurement in fish holding-water. Ecol. Indic. 2020, 109, 105829. [Google Scholar] [CrossRef]
- Sontakke, R.; Tiwari, V.K.; Paniprasad, K.; Rani, A.B.; Ande, M.P. Nonspecific immune and antioxidant status of milkfish, Chanos chanos varies with the carbon source used in the biofloc system. J. Exp. Zool. India 2018, 22, 109–118. [Google Scholar]
- Kim, J.H.; Kang, J.C. Effects of dietary chromium exposure to rockfish, Sebastes schlegelii are ameliorated by ascorbic acid. Ecotox. Environ. Saf. 2017, 139, 109–115. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, J.Y.; Lim, L.J.; Kim, S.K.; Choi, H.S.; Hur, Y.B. Effects of waterborne nitrite on hematological parameters and stress indicators in olive flounders, Paralichthys olivaceus, raised in bio-floc and seawater. Chemosphere 2018, 209, 28–34. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.K.; Hur, Y.B. Hematological parameters and antioxidant responses in olive flounder Paralichthys olivaceus in biofloc depend on water temperature. J. Therm. Biol. 2019, 82, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Choi, H.; Sung, G.; Seo, S.A.; Kim, K.I.; Kang, Y.J.; Kang, J.C. Toxic effects on hematological parameters and oxidative stress in juvenile olive flounder, Paralichthys olivaceus exposed to waterborne zinc. Aquacult. Rep. 2019, 15, 100225. [Google Scholar] [CrossRef]
- Narra, M.R.; Rajender, K.; Reddy, R.R.; Murty, U.S.; Begum, G. Insecticides induced stress response and recuperation in fish: Biomarkers in blood and tissues related to oxidative damage. Chemosphere 2017, 168, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Gholipour Kanani, H.; Nobahar, Z.; Kakoolaki, S.; Jafarian, H. Effect of ginger-and garlic-supplemented diet on growth performance, some hematological parameters and immune responses in juvenile Huso huso. Fish Physiol. Biochem. 2014, 40, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, M.; Kim, J.Y.; Hur, K.J.; Ramesh, M.; Hur, J.H. Responses of the freshwater fish Cyprinus carpio exposed to different concentrations of butachlor and oxadiazon. Biocatal. Agric. Biotechnol. 2017, 11, 275–281. [Google Scholar] [CrossRef]
- Adel, M.; Dawood, M.A.; Shafiei, S.; Sakhaie, F.; Shekarabi, S.P.H. Dietary Polygonum minus extract ameliorated the growth performance, humoral immune parameters, immune-related gene expression and resistance against Yersinia ruckeri in rainbow trout (Oncorhynchus mykiss). Aquaculture 2020, 519, 734738. [Google Scholar] [CrossRef]
- Kim, J.H.; Cho, J.H.; Kim, S.R.; Hur, Y.B. Toxic effects of waterborne ammonia exposure on hematological parameters, oxidative stress and stress indicators of juvenile hybrid grouper, Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀. Environ. Toxicol. Pharmacol. 2020, 80, 103453. [Google Scholar] [CrossRef]
- Yu, Z.; Li, L.; Zhu, R.; Li, M.; Wu, L.F. Effects of bioflocs with different C/N ratios on growth, immunological parameters, antioxidants and culture water quality in Opsariichthys kaopingensis Dybowski. Aquac. Res. 2020, 51, 805–815. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeong, E.H.; Jeon, Y.H.; Kim, S.K.; Hur, Y.B. Salinity-mediated changes in hematological parameters, stress, antioxidant responses, and acetylcholinesterase of juvenile olive flounders (Paralichthys olivaceus). Environ. Toxicol. Pharmacol. 2021, 83, 103597. [Google Scholar] [CrossRef]
- Kim, J.H.; Yu, Y.B.; Choi, J.H. Toxic effects on bioaccumulation, hematological parameters, oxidative stress, immune responses and neurotoxicity in fish exposed to microplastics: A review. J. Hazard. Mater. 2021, 413, 125423. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. The toxic effects on the stress and immune responses in juvenile rockfish, Sebastes schlegelii exposed to hexavalent chromium. Environ. Toxicol. Pharmacol. 2016, 43, 128–133. [Google Scholar] [CrossRef]
- Vargas-Chacoff, L.; Martínez, D.; Oyarzún, R.; Nualart, D.; Olavarría, V.; Yáñez, A.; Bertrán, C.; Ruiz-Jarabo, I.; Mancera, J.M. Combined effects of high stocking density and Piscirickettsia salmonis treatment on the immune system, metabolism and osmoregulatory responses of the Sub-Antarctic Notothenioid fish Eleginops maclovinus. Fish Shellfish Immunol. 2014, 40, 424–434. [Google Scholar] [CrossRef]
- Menaga, M.; Felix, S.; Charulatha, M.; Gopalakannan, A.; Panigrahi, A. Effect of in-situ and ex-situ biofloc on immune response of Genetically Improved Farmed Tilapia. Fish Shellfish Immunol. 2019, 92, 698–705. [Google Scholar] [CrossRef]
- Kamilya, D.; Debbarma, M.; Pal, P.; Kheti, B.; Sarkar, S.; Singh, S.T. Biofloc technology application in indoor culture of Labeo rohita (Hamilton, 1822) fingerlings: The effects on inorganic nitrogen control, growth and immunity. Chemosphere 2017, 182, 8–14. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. Oxidative stress, neurotoxicity, and metallothionein (MT) gene expression in juvenile rock fish Sebastes schlegelii under the different levels of dietary chromium (Cr6+) exposure. Ecotox. Environ. Saf. 2016, 125, 78–84. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. Effects of sub-chronic exposure to lead (Pb) and ascorbic acid in juvenile rockfish: Antioxidant responses, MT gene expression, and neurotransmitters. Chemosphere 2017, 171, 520–527. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Doan, H.V.; Ashouri, G.; Gioacchini, G.; Maradonna, F.; Carnevali, O. Oxidative stress and antioxidant defense in fish: The implications of probiotic, prebiotic, and synbiotics. Rev. Fish. Sci. Aquac. 2020, 29, 198–217. [Google Scholar] [CrossRef]
- Zhang, Y.; Roh, Y.J.; Han, S.J.; Park, I.; Lee, H.M.; Ok, Y.S.; Lee, B.C.; Lee, S.R. Role of selenoproteins in redox regulation of signaling and the antioxidant system: A review. Antioxidants 2020, 9, 383. [Google Scholar] [CrossRef]
- Yu, Z.; Li, L.; Zhu, R.; Li, M.; Duan, J.; Wang, J.Y.; Liu, Y.H.; Wu, L.F. Monitoring of growth, digestive enzyme activity, immune response and water quality parameters of Golden crucian carp (Carassius auratus) in zero-water exchange tanks of biofloc systems. Aquacult. Rep. 2020, 16, 100283. [Google Scholar] [CrossRef]
- Yu, Z.; Zhao, Y.Y.; Jiang, N.; Zhang, A.Z.; Li, M.Y. Bioflocs attenuates lipopolysaccharide-induced inflammation, immunosuppression and oxidative stress in Channa argus. Fish Shellfish Immunol. 2021, 114, 218–228. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, Y.J.; Kim, K.I.; Kim, S.K.; Kim, J.H. Toxic effects of nitrogenous compounds (ammonia, nitrite, and nitrate) on acute toxicity and antioxidant responses of juvenile olive flounder, Paralichthys olivaceus. Environ. Toxicol. Pharmacol. 2019, 67, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, K.; Pereira, E.; Duarte, A.C.; Ahmad, I. Glutathione and its dependent enzymes’ modulatory responses to toxic metals and metalloids in fish-a review. Environ. Sci. Pollut. Res. 2013, 20, 2133–2149. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Choi, H.; Hwang, U.K.; Kang, J.C.; Kang, Y.J.; Kim, K.I.; Kim, J.H. Toxic effects of lead exposure on bioaccumulation, oxidative stress, neurotoxicity, and immune responses in fish: A review. Environ. Toxicol. Pharmacol. 2019, 68, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.C.; Choi, Y.J.; Kim, J.H. Toxic effects of waterborne cadmium exposure on hematological parameters, oxidative stress, neurotoxicity, and heat shock protein 70 in juvenile olive flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2022, 122, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kang, Y.J.; Lee, K.M. Effects of Nitrite Exposure on the Hematological Properties, Antioxidant and Stress Responses of Juvenile Hybrid Groupers, Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀. Antioxidants 2022, 11, 545. [Google Scholar] [CrossRef]
- Dilmi, A.; Refes, W.; Meknachi, A. Effects of C/N Ratio on Water Quality, Growth Performance, Digestive Enzyme Activity and Antioxidant Status of Nile tilapia Oreochromis niloticus (Linnaeus, 1758) in Biofloc Based Culture System. Turk. J. Fish. Quat. Sci. 2021, 22, TRJFAS19754. [Google Scholar] [CrossRef]
- Aliabad, H.S.; Naji, A.; Mortezaei, S.R.S.; Sourinejad, I.; Akbarzadeh, A. Effects of restricted feeding levels and stocking densities on water quality, growth performance, body composition and mucosal innate immunity of Nile tilapia (Oreochromis niloticus) fry in a biofloc system. Aquaculture 2022, 546, 737320. [Google Scholar] [CrossRef]
- Doan, H.V.; Lumsangkul, C.; Hoseinifar, S.H.; Tongsiri, S.; Chitmanat, C.; Musthafa, M.S.; El-Haroun, E.; Ringo, E. Modulation of growth, innate immunity, and disease resistance of Nile tilapia (Oreochromis niloticus) culture under biofloc system by supplementing pineapple peel powder and Lactobacillus plantarum. Fish Shellfish Immunol. 2021, 115, 212–220. [Google Scholar] [CrossRef]
- Ahmad, I.; Babitha Rani, A.M.; Verma, A.K.; Maqsood, M. Biofloc technology: An emerging avenue in aquatic animal healthcare and nutrition. Aquac. Int. 2017, 25, 1215–1226. [Google Scholar] [CrossRef]
- Elayaraja, S.; Mabrok, M.; Algammal, A.; Sabitha, E.; Rajeswari, M.V.; Zágoršek, K.; Ye, Z.; Zhu, S.; Rodkhum, C. Potential influence of jaggery-based biofloc technology at different C:N ratios on water quality, growth performance, innate immunity, immune-related genes expression profiles, and disease resistance against Aeromonas hydrophila in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2020, 107, 118–128. [Google Scholar] [CrossRef]
- Jensch-Junior, B.E.; Pressinotti, L.N.; Borges, J.C.S.; da Silva, J.R.M.C. Characterization of macrophage phagocytosis of the tropical fish Prochilodus scrofa (Steindachner, 1881). Aquaculture 2006, 251, 509–515. [Google Scholar] [CrossRef]
- Esteban, M.Á.; Cuesta, A.; Chaves-Pozo, E.; Meseguer, J. Phagocytosis in teleosts. Implications of the new cells involved. Biology 2015, 4, 907–922. [Google Scholar] [CrossRef]
- Fazelan, Z.; Vatnikov, Y.A.; Kulikov, E.V.; Plushikov, V.G.; Yousefi, M. Effects of dietary ginger (Zingiber officinale) administration on growth performance and stress, immunological, and antioxidant responses of common carp (Cyprinus carpio) reared under high stocking density. Aquaculture 2020, 518, 734833. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. The immune responses in juvenile rockfish, Sebastes schlegelii for the stress by the exposure to the dietary lead (II). Environ. Toxicol. Pharmacol. 2016, 46, 211–216. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. The immune responses and expression of metallothionein (MT) gene and heat shock protein 70 (HSP 70) in juvenile rockfish, Sebastes schlegelii, exposed to waterborne arsenic (As3+). Environ. Toxicol. Pharmacol. 2016, 47, 136–141. [Google Scholar] [CrossRef]
- Teng, Y.; Fu, Q.; Tan, Y.; Ding, Y.; Ding, Y.; Chen, X. Identification and characterization of immunoglobulin T heavy chain in large yellow croaker (Larimichthys crocea). Fishes 2022, 7, 29. [Google Scholar] [CrossRef]
- Lee, J.W.; Kim, J.H.; Lee, D.C.; Lim, H.J.; Kang, J.C. Toxic effects on oxidative stress, neurotoxicity, stress, and immune responses in juvenile olive flounder, Paralichthys olivaceus, exposed to waterborne hexavalent chromium. Biology 2022, 11, 766. [Google Scholar] [CrossRef]
- Akrami, R.; Gharaei, A.; Mansour, M.R.; Galeshi, A. Effects of dietary onion (Allium cepa) powder on growth, innate immune response and hemato–biochemical parameters of beluga (Huso huso Linnaeus, 1754) juvenile. Fish Shellfish Immunol. 2015, 45, 828–834. [Google Scholar] [CrossRef]
- Chen, C.; Yang, B.; Raza, S.H.A.; Zhang, D.; Wu, T.; Zhang, Z.; Ullah, I.; Khan, R.; Yang, G.; Wang, C.; et al. Role of Myeloperoxidase of northern snakehead (Channa argus) in Aeromonas veronii infection. Microb. Pathog. 2019, 135, 103622. [Google Scholar] [CrossRef]
- Buchan, K.D.; Prajsnar, T.K.; Ogryzko, N.V.; De Jong, N.W.; Gent, M.V.; Kolata, J.; Foster, S.J.; van Strijp, J.A.G.; Renshaw, S.A. A transgenic zebrafish line for in vivo visualisation of neutrophil myeloperoxidase. PLoS ONE 2019, 14, e0215592. [Google Scholar] [CrossRef]
- Kumar, S.; Sahu, N.P.; Pal, A.K.; Choudhury, D.; Yengkokpam, S.; Mukherjee, S.C. Effect of dietary carbohydrate on haematology, respiratory burst activity and histological changes in L. rohita juveniles. Fish Shellfish Immunol. 2005, 19, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Biller-Takahashi, J.D.; Takahashi, L.S.; Saita, M.V.; Gimbo, R.Y.; Urbinati, E.C. 2013 Leukocytes respiratory burst activity as indicator of innate immunity of pacu Piaractus mesopotamicus. Braz. J. Biol. 2013, 73, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Bavia, L.; Santiesteban-Lores, L.E.; Carneiro, M.C.; Prodocimo, M.M. Advances in the complement system of a teleost fish, Oreochromis niloticus. Fish Shellfish Immunol. 2022, 123, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.; Portugal, I.; Zilberg, D. Analyzing complement activity in the serum and body homogenates of different fish species, using rabbit and sheep red blood cells. Vet. Immunol. Immunopathol. 2018, 199, 39–42. [Google Scholar] [CrossRef]
- Yousefi, M.; Hoseini, S.M.; Vatnikov, Y.A.; Kulikov, E.V.; Drukovsky, S.G. Rosemary leaf powder improved growth performance, immune and antioxidant parameters, and crowding stress responses in common carp (Cyprinus carpio) fingerlings. Aquaculture 2019, 505, 473–480. [Google Scholar] [CrossRef]
- Kuroda, N.; Naruse, K.; Shima, A.; Nonaka, M.; Sasaki, M. Molecular cloning and linkage analysis of complement C3 and C4 genes of the Japanese medaka fish. Immunogenetics 2000, 51, 117–128. [Google Scholar] [CrossRef]
- Kim, J.H.; Sohn, S.; Kim, S.K.; Kim, S.R.; Kim, S.K.; Kim, S.M.; Kim, N.Y.; Hur, Y.B. Effects on the survival rates, hematological parameters, and neurotransmitters in olive flounders, Paralichthys olivaceus, reared in bio-floc and seawater by Streptococcus iniae challenge. Fish Shellfish Immunol. 2021, 113, 79–85. [Google Scholar] [CrossRef]
- Gutiérrez, S.M.; Dosta, M.D.C.M.; Partida, A.H.; Mejía, J.C.; Rodríguez, G.A.; de Oca, M. Effect of two carbon sources in microbial abundance in a Biofloc culture system with Oreochromis niloticus (Linnaeus, 1758). Int. J. Fish. Aquat. 2016, 4, 421–427. [Google Scholar] [CrossRef]

| Species | Pathogen (Strain) | CFU | Carbon Source | C/N Ratio | Period | Response C/N Ratio | Response | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| Survival rates | |||||||||
| Freshwater | Oreochromis niloticus | Vibrio harveyi | 4 × 108 CFU/mL | Glucose (166 organisms/m3) | 15:1 | 14 days | 15:1 | + | [41] |
| Glucose (333 organisms/m3) | 15:1 | + | |||||||
| Glucose (600 organisms/m3) | - | × | |||||||
| Aeromonas hydrophila (ATCC 7966) | 3 × 108 CFU/mL | Mannan oligosaccharides + plant-protein-based diet | 15:1 | 3 days | 15:1 | + | [24] | ||
| Mannan oligosaccharides + fish-protein-based diet | 15:1 | + | |||||||
| Glycerol + plant-protein-based diet | 15:1 | + | |||||||
| Glycerol + fish-protein-based diet | 15:1 | + | |||||||
| Aeromonas hydrophila (ATCC 7966) | 3 × 108 CFU/mL | Mannan oligosaccharides + plant-protein-based diet | 15:1 | 7 days | 15:1 | + | |||
| Mannan oligosaccharides + fish-protein-based diet | 15:1 | + | |||||||
| Glycerol + plant-protein-based diet | 15:1 | + | |||||||
| Glycerol + fish-protein-based diet | 15:1 | + | |||||||
| Aeromonas hydrophila (ATCC 7966) | 106 CFU/mL | Wheat flour (200 fish/m3) | 15:1 | 3 days | 15:1 | + (83.33%) | [30] | ||
| Wheat flour (250 fish/m3) | 15:1 | + (83.33%) | |||||||
| Wheat flour (300 fish/m3) | 15:1 | + (75%) | |||||||
| Wheat flour (350 fish/m3) | 15:1 | + (62.5%) | |||||||
| Clarias gariepinus | Aeromonas hydrophila | 106 CFU/mL | Tapioca (4 fish/L) | 10:1 | 7 days | 10:1 | + | [19] | |
| Tapioca (6 fish/L) | 10:1 | + | |||||||
| Tapioca (8 fish/L) | 10:1 | + | |||||||
| Labeo rohita | Aeromonas hydrophila (ATCC 7966) | 1.8 × 107 CFU/mL | Tapioca | 15:1 | 14 days | 15:1 | + | [45] | |
| Wheat | 15:1 | + | |||||||
| Corn | 15:1 | + | |||||||
| Sugar bagasse | 15:1 | + | |||||||
| Cyprinus carpio | Aeromonas hydrophila (ATCC 897) | 1.1 × 107 CFU/mL | Sugar beet molasses | 15:1 | 14 days | 15:1 | + | [44] | |
| Sugar | 15:1 | + (50%) | |||||||
| Corn starch | 15:1 | + (50%) | |||||||
| Seawater | Paralichthys olivaceus | Edwardsiella tarda (FP 5060) | 6.61 × 104 CFU/g fish | - | - | 7 days | - | × | [5] |
| 6.61 × 105 CFU/g fish | - | + (100%) | |||||||
| 6.61 × 106 CFU/g fish | - | + (33%) | |||||||
| 6.61 × 107 CFU/g fish | - | + (33%) | |||||||
| Streptococcus iniae (FP 5228) | 3.36 × 106 CFU/g fish | Glucose | 10:1 | 96 h | 10:1 | + (100%) | [70] | ||
| 3.36 × 107 CFU/g fish | 10:1 | + (80%) | |||||||
| 3.36 × 108 CFU/g fish | 10:1 | + (70%) | |||||||
| 3.36 × 109 CFU/g fish | 10:1 | × (20%) | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.-B.; Choi, J.-H.; Lee, J.-H.; Jo, A.-H.; Lee, K.M.; Kim, J.-H. Biofloc Technology in Fish Aquaculture: A Review. Antioxidants 2023, 12, 398. https://doi.org/10.3390/antiox12020398
Yu Y-B, Choi J-H, Lee J-H, Jo A-H, Lee KM, Kim J-H. Biofloc Technology in Fish Aquaculture: A Review. Antioxidants. 2023; 12(2):398. https://doi.org/10.3390/antiox12020398
Chicago/Turabian StyleYu, Young-Bin, Jae-Ho Choi, Ju-Hyeong Lee, A-Hyun Jo, Kyung Mi Lee, and Jun-Hwan Kim. 2023. "Biofloc Technology in Fish Aquaculture: A Review" Antioxidants 12, no. 2: 398. https://doi.org/10.3390/antiox12020398